D
IPARTIMENTO DIS
CIENZEA
RCHEOLOGICHE DOTTORATO DI RICERCA IN ARCHEOLOGIA CURRICULUM PALEOANTROPOLOGIA E PATOCENOSI(XIX ciclo) BIO/08
Tesi di dottorato
Paleogenetica, analisi di coalescenza seriale e paleonutrizione in resti umani
preistorici e storici del territorio lucchese
Dottoranda
Dott.ssa Annalisa Conzato
Coordinatore Tutore
Prof. Carlo Tozzi Prof. Francesco Mallegni
Progetto di ricerca: Paleoantropologia, paleonutrizione e paleogenetica dei resti umani medievali rinvenuti intorno e dentro le chiese lucchesi di S. Luca, di via S. Paolino e di S. Pietro in Gottelle, viste nell’ambito della genetica di popolazione attuale cittadina e del territorio.
Indice
1 IL CASO LUCCHESE. SCOPO DELLA RICERCA ... 4
1.1 Uno sguardo alla storia... 4
1.2 Cenni di demografia ... 8
1.2.1 Demografia toscana tra XIV - XX secolo ... 12
1.3 Provenienza dei campioni osteologici. Qualche considerazione ... 12
1.4 Finalità del lavoro... 16
2 ANTROPOLOGIA MOLECOLARE ... 19
2.1 La rivoluzione dell’antropologia molecolare ... 19
2.1.1 Molecole e fossili a confronto ... 21
2.1.2 Le molecole come congegni di datazione ... 23
2.1.3 Marcatori molecolari ... 26
2.1.4 Caratteristiche e potenzialità del DNA mitocondriale ... 28
2.1.5 Recupero e analisi del DNA mitocondriale... 31
2.1.6 Studi sul DNA antico e sue applicazioni... 33
2.2 Materiali e Metodi... 35
2.2.1 Campionamento... 35
2.2.2 Un problema su tutti: la contaminazione... 37
2.2.3 Studio dei reperti antichi. Principali tecniche biomolecolari ... 38
Trattamento dei campioni... 39
Valutazione del grado di racemizzazione degli amminoacidi... 39
Estrazione del Dna antico ... 40
La reazione a catena della polimerasi... 40
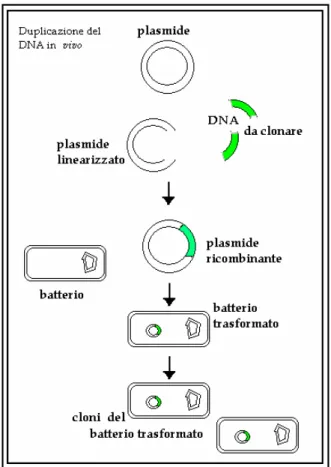
Il clonaggio ... 43
Il sequenziamento ... 44
Lettura e ricostruzione della sequenza nucleotidica... 45
2.2.4 Metodi di autenticazione ... 47
2.3 Analisi dei dati e Risultati ... 49
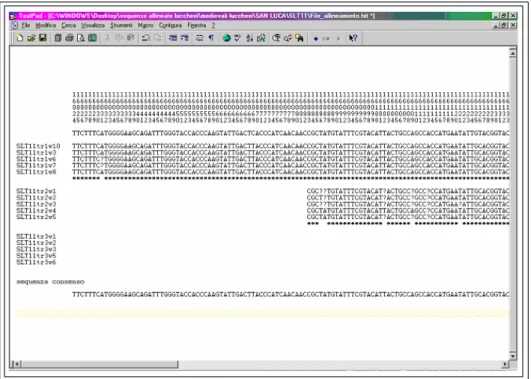
2.3.1 Polimorfismi e sequenze consenso... 49
2.3.2 Elaborazione dei dati. Analisi a livello intra- e infra- popolazionistico... 51
2.3.3 Simulazione di coalescenza... 52
La teoria della coalescenza ... 52
Il programma Serial SimCoal ... 53
2.3.4 MDS ... 58
2.4 Analisi dei risultati ... 59
2.4.1 Modelli di coalescenza a confronto... 59
2.4.2 Confronto interpopolazionistico. MDS ... 74
2.4.3 Valutazione del grado di racemizzazione... 75
2.5 Discussione... 76
3 PALEONUTRIZIONE ... 80
3.1 La chimica dell’osso e la diagenesi... 81
3.2.1 Metodi d’indagine ... 83
3.2.2 Preparazione dei campioni ossei ... 84
3.3 Gli oligoelementi... 86
3.4 Elaborazione dei dati... 90
3.4.1 Riconoscimento della dieta ... 91
3.5 Risultati ... 93
3.5.1 Eneolitico... 93
3.5.2 Etruschi e I secolo a.C... 97
3.5.3 Secoli V - VII d.C... 98
3.5.4 Secoli XVI - XVIII d.C.. ... 101
3.6 Discussione... 104
Bibliografia ... 108
1 Il caso Lucchese. Scopo della ricerca... 108
2 Antropologia molecolare... 110
1 IL CASO LUCCHESE. SCOPO DELLA RICERCA
1.1 Uno sguardo alla storia
Per capire in che misura l’arrivo di popolazioni alloctone e l’apporto nei secoli dei contatti con altre culture ha influenzato la struttura della popolazione attuale è necessario fare un percorso a ritroso nel tempo che ci conduce oltre la storia.
Il paleoambiente della conca lucchese era assai diverso rispetto alla situazione odierna, se è vero che l’ambiente nel corso dei secoli ha influenzato l’evoluzione e la diffusione dell’uomo, è pure vero che un popolo vi si può introdurre solo se possiede capacità adattative e culturali capaci di colonizzarlo e modificarlo. Le scoperte relative ad insediamenti del Paleolitico, del Mesolitico, del Neolitico e di numerose grotticelle sepolcrali eneolitiche, testimoniano l’antica frequentazione umana dell’area lucchese. I più antichi manufatti litici appartengono al paleolitico inferiore (350.000 anni fa). Tra 80.000 e 10.000 anni fa durante la glaciazione würmiana, prima l’uomo di Neanderthal, e poi il Sapiens sapiens lascerà testimonianze della sua presenza nella parte sud-orientale della pianura lucchese (Mencacci et al., 1976). In questo periodo la conca lucchese compresa tra gli alvei dei fiumi Serchio e Arno, si presentava ricca di dorsi alluvionali, di insulae tra braccia di fiume mutevoli, che negli ultimi millenni facilitarono lo sviluppo di insediamenti (Mencacci et al., 1982). Il ritrovamento di manufatti litici rappresentano gran parte delle culture preistoriche sino al Bronzo medio, dopo di che scompaiono le tracce di frequentazione che si ripresentarono nel VIII secolo a.C. (Villanoviano) per proseguire ininterrottamente con gli Etruschi e i Romani. I secoli non rappresentati da testimonianze archeologiche furono probabilmente caratterizzati da un peggioramento delle condizioni climatiche, particolarmente umido e piovoso, che determinarono l’allagamento della piana spingendo i gruppi umani verso le alture. La situazione iniziò a ristabilirsi con la ‘rotta’ del Paleoserchio che probabilmente a partire dal XIV – XII secolo si aprì una seconda via attraverso le strette di Ripafratta permettendo il deflusso delle acque dalla piana. La nuova rotta del fiume verso sud-ovest determinò radicali trasformazioni ambientali che influenzarono l’abitabilità della zona (Mencacci et al., 1982).
Le ultime testimonianze archeologiche prima del gap temporale, a parte le due piroghe del Bronzo medio rinvenute nella torbiera di Orentano, risalgono all’età del Rame. La sostanziale omogeneità del quadro culturale eneolitico, cioè il seppellimento in grotta dei
cadaveri dovuto a fattori più o meno intenzionali (Cocchi Genick et al., 1985), ha preservato parte degli scheletri, per la gran parte recuperati a cavallo del Novecento, che oggigiorno costituiscono il substrato più antico dai cui parte la presente indagine antropologica su Lucca e il suo territorio, in particolare quella paleogenetica e paleonutrizionale. L’uso di grotte naturali e di crepacci per deporvi i cadaveri, che nella gran parte dei casi si ritrovano sconvolti a causa delle successive frequentazioni degli ambienti, era il rituale di seppellimento tipico delle genti eneolitiche nella Toscana nord-occidentale a nord dell’Arno. Un’usanza assai diversa dalle culture centro-meridionali di Rinaldone, del Gaudo e di Laterza (grotticella artificiale) e dalla zona padana e mineraria toscana (tomba a fossa) (Cocchi Genick et. al., 1985). Le vicende del popolamento dell’area si perdono nella nebulosa protostoria che vedono l’alternanza di genti diverse, se non nel genotipo, sicuramente sotto l’aspetto culturale. Intorno all’anno mille a.C. tribù germaniche provenienti dal nord Europa presero stabile dimora nella pianura che si estendeva ad ovest del lago di Bietina percorsa dai corsi d’acqua Serchio e Ozzeri (Giovannini, 2003); ed infatti, l’etimologia del toponimo Lucca, non avrebbe radici etrusche, ligure o celtoligure, ‘Luck’ luogo paduligno così come un tempo fu accreditata (Mancini, 1975), ma sarebbe di origine indoeuropea, ‘Lawk’ successivamente latinizzato in ‘Luca’. Essendo terra di confine la pianura lucchese passò a più riprese in mano ligure ed etrusca e successivamente anche romana. A partire dal IV secolo a.C. la romanizzazione dell’area Toscana determinò un relativo spopolamento dei prosperi insediamenti etruschi collocati principalmente nel sud della regione a favore del nord ponendo la base per la nascita di nuovi centri (Pisa, Firenze, Fiesole,Volterra, Lucca ecc.) (Francovich, 1999). Nel 180 a.C. Lucca fu costituita colonia latina.
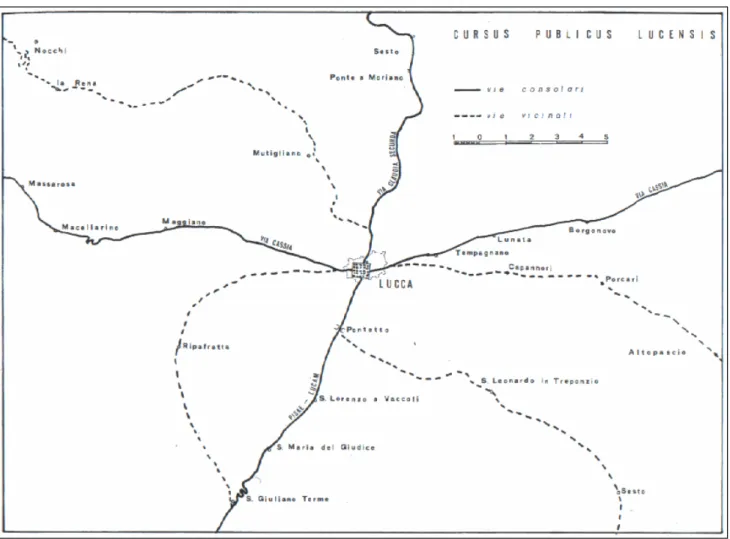
La presenza romana costituì una nuova compagine sociale che assorbì la popolazione locale Etrusca e Ligure, i territori vennero bonificati e riorganizzati e fondamentale fu la realizzazione di una grande rete di strade consolari che collegavano la regione a Roma e all’Italia settentrionale. ‘Luca’ assunse ben presto un ruolo di primaria importanza nei rapporti militari, amministrativi e commerciali, era servita da quattro coppie di strade, due per ogni porta, una primaria e una secondaria (Fig. 1.1a). Le otto strade disposte a raggiera erano collegate tra loro da una rete di diverticola e di vie secondarie, le vie ‘vicinales’ che collegavano il municipium lucchese con i pagi più lontani. Una strada consolare probabilmente esistente già nel II-I sec a.C., detta Cassia o Clodia, collegava la regione da un
lato ad Arezzo e di qui a Roma, dall’altro alla viabilità costiera. Verso nord, attraverso la valle del Serchio, dove ancora si conservano alcuni toponimi viari (Sesto di Moriano, Valdottavo, Diecimo), si snodava la via Luca - Parma. Altre strade permettevano ancora di raggiungere la costa: verso Pisa, aggirando i Monti Pisani, e verso Luni, attraverso la Val Freddana (http://luccapro.sns.it/ita/non/arc), la via Faetina metteva in comunicazione l’Etruria con l’Emilia (Riparbelli A, 1982). Quindi è indubbio che Lucca già dal tempo dei romani fosse un importante nodo viario.
La divisione dell’impero Romano in quelli d’Occidente e d’Oriente e i grandi spostamenti di popolazioni che si muovevano a partire dalle vaste pianure mongoliche segnarono la decadenza della supremazia romana.
L’Alto Medioevo quindi s’apre all’insegna dei grandi movimenti migratori forieri di trasformazioni che si intersecano con i cambiamenti apportati da un altro tipo di migrazione, quella a breve raggio. Essa è determinata da spostamenti continui come ad esempio quelli per matrimoni, o quelli legati al commercio, alla transumanza, o ai movimenti di inurbamento notoriamente legati a periodi non prosperi di carestie e instabilità politica. In modo fluttuante la migrazione a breve raggio determina il mescolamento di culture, beni materiali e persone. Per registrare una stasi dei grandi flussi migratori in Toscana si dovrà aspettare l’XI secolo; infatti, è proprio dopo l’anno Mille che gli studiosi individuano gli elementi di continuità ancora presenti nella struttura della popolazione attuale (Francovich, 1999).
I mongoli spingevano le popolazioni germaniche a spostarsi verso ovest, sud-ovest, forzando i confini nord-orientali dell’impero; iniziarono così le invasioni barbariche che si spinsero ben due volte sino a Roma. “Quando Lucca divenisse Longobarda al certo non si
sa, ma può essere stato verisimilmente regnando Alboino”, vale a dire prima del 574
(Mazzarosa, 1841) e la dominazione si protrasse per circa due secoli (Giovannini, 2003). Lucca era il centro del potere della Tuscia longobarda, gli insediamenti nella lucchesia e nel pistoiese erano sparsi e pare che le genti d’origine longobarda fossero esigue (Frankowich, 1999). L’irruzione longobarda in Italia fu dal punto di vista demico scarsamente incisivo, si parla di qualche centinaio di migliaia di invasori (Del Panta et al., 1996). Sotto il regno del re Autari (584-590) fu aperta, ricalcando i preesistenti tracciati romani, la via Francigena per collegare il regno di Pavia con la Tuscia attraverso percorsi alternativi a quelli controllati dai bizantini, per cui il passo della Cisa rimase la scelta obbligata per attraversare l’Appennino e quindi la Parma-Lucca, che usava tale valico, rimase la sola strada praticabile (Stopani,
1998); l’uso del passo della Cisa anticamente detta via di monte Bardone è ricordato pure dal cronista Paolo Diacono “… per Alpem Bardonis Tusciam ingressus”. Il percorso seguiva gli itinerari dei pellegrini diretti a Roma e in Terrasanta snodandosi longitudinalmente lungo la penisola. Molti furono i monasteri istituiti che fungevano da spedali per l’accoglienza dei viandanti piuttosto che da istituti religiosi. Essi si trovavano nei punti nodali della via per Roma e anche in Lucca e negli immediati dintorni della città sorsero molti xenodochia (luogo di ricetto per forestieri) e molte abbazie, alcune delle quali già in epoca longobarda (Stopani, 1998). La via Francigena viene considerata la strada più antica d’Europa, proveniva dalla Francia, varcava le Alpi e gli Appennini in corrispondenza del Passo della Cisa. Prese il nome di ‘Francigena’ o ‘Francesca’ quando, con la costituzione del Sacro Romano Impero e la discesa di Carlo Magno a Roma per l’incoronazione nella notte del Natale del 800, i Franchi cominciarono a percorrerla; solo più tardi raccordandosi con la Romea fu chiamata anch’essa Romea, perché conduceva a Roma. Non si arrestarono le fondazioni di abbazie, l’assistenza ai pellegrini e ai viandanti costituiva un compito di carattere pubblico. Durante il X e XI secolo il numero di pellegrini, mercanti e persone di ogni estrazione sociale che si mettevano in viaggio, percorrendo tutta l’Italia, per motivi religiosi o commerciali, divenne sempre più consistente. Tra i secoli XI e XII la strada è il punto d’incontro delle tre peregrinationes maiores della cristianità: Roma, sede delle tombe di San Pietro e Paolo, la Terrasanta e il Santo Sepolcro e, infine, Compostela per la venerazione delle spoglie di San Giacomo (Stopani, 1998). Era una strada variabile nei percorsi a seconda delle stagioni, il tracciato divenne più stabile con i Franchi, però presentava alcuni punti che costituivano passaggi obbligati per chiunque; Lucca, infatti, era una meta obbligata per la presenza del Volto Santo. La statua di Gesù Cristo detta la Santa Croce o Volto Santo “… fatta in parte da Nicodemo e in parte da gli Angeli” era custodita e adorata nel duomo di Lucca, la devozione coinvolgeva la gente del luogo e numerosi forestieri (Paolucci, 1718). Tra l’altro la città si trova vicina al porto di Pisa che in epoca Alto medievale era il più importante della costa Tirrenica tra Civitavecchia e Genova.
Lucca sin dall’epoca Tardo antica è al centro di una viabilità importante, la centralità, l’arrivo e il passaggio di popolazioni e de gli eserciti favorirono scambi commerciali e culturali e un apporto umano differenziato.
Appare chiaro che le lontane, lontanissime origini della popolazione sono miste e profondamente segnate dalle diverse dominazioni.
Figura 1.1a –Lucca romana: rete viaria primaria e secondaria (da Riparbelli, 1982).
1.2 Cenni di demografia
La vicenda demografica in dimensione diacronica della città di Lucca e dintorni è piuttosto difficile da seguire, in quanto prima del XIII secolo la documentazione si fa molto sporadica e non esauriente (Beloch). Quindi per il periodo antecedente a tale secolo, si cercherà di fare una panoramica demografica proiettando su Lucca l’evoluzione della popolazione della Toscana e dell’Italia. L’andamento della popolazione nel corso dei secoli sembra rispecchiare le caratteristiche proprie delle popolazioni di antico regime demografico, nelle quali la bassa speranza di vita, le ricorrenti epidemie e crisi di mortalità frenavano lo sviluppo demografico.
A partire dall’anno 0 la fase d’espansione demografica dell’era cristiana termina nel 200, segue poi una lunga caduta che porterà la popolazione italiana ben al di sotto del valore
iniziale. La recessione demografica della tarda antichità e del primo medioevo ridusse la popolazione di circa la metà. La crisi demografica delle città è attestata a partire dalla metà del III secolo quando alcune di esse, tra cui Lucca, ricostruirono le mura lasciando fuori quartieri un tempo popolati per difendersi meglio (Barni, in Del Panta et al., 1996). All’irruzione armata dei popoli germanici nel primo Medioevo fece seguito lo stanziamento sulle terre conquistate e la progressiva fusione con le antiche popolazioni.
La lenta ripresa ci metterà quasi mille anni prima di raggiungere intorno al 1200 un numero di 8,5 milioni di abitanti all’incirca uguale ai valori iniziali dei primi due secoli dopo Cristo; la crescita prolungata raggiunse il culmine nel 1300. La ripresa della popolazione nelle città intorno al X-XI secolo segna il trionfo dell’urbanesimo all’inizio del Trecento. I sistemi urbani più importanti erano situati lungo le principali direttrici; nella Toscana centro settentrionale ci sono grandi e importanti città che, in età classica, avevano svolto un ruolo secondario (Pinto, 1993); città quali Firenze, Pisa e Siena e un reticolo di centri minori, tra cui Lucca (Del Panta et al. 1996), una delle poche città medievali rilevanti a vantare significativi trascorsi romani (Pinto, 1993). Tra il 1300-1338 Lucca contava molto probabilmente più di 20-25.000 abitanti, 30.000 secondo il Pinto (1988), nei secoli precedenti era stata quasi sicuramente il principale polo urbano della regione svolgendo un ruolo di primo piano nel commercio e nella finanza internazionale (Ginatempo et al., 1990).
Ma a metà del secolo la mano delle carestie, delle guerre e la falce della Peste Nera causarono il declino demografico, determinando soprattutto lo svuotamento del contado, che si protrasse fino al 1450. Le epidemie che colpirono Lucca e le Sei Miglia (territorio che si estendeva intorno alla città per 250 kmq. Il Repetti la definì come “Una campagna
d’irregolare periferia”) furono assai numerose: 1348, 1363-64, 1373, 1383, 1392, 1400 e
1418. L’epidemia del 1400 fu la più terribile, secondo le cronache del tempo in città morivano fino a 140 persone al giorno, ed i suoi effetti si evidenziano chiaramente nella piramide dell’età del periodo, che mostra una netta strozzatura nella classe tra i 15 e i 19 anni. In generale la ciclicità degli eventi pestilenziali interrompeva puntualmente le riprese demografiche; ma per il contado lucchese la bassa età al matrimonio, la limitata differenza d’età tra i coniugi e il lungo periodo fecondo favorirono l’espansione demografica che continuò anche alcuni anni dopo la pestilenza del 1400 (Leverotti, 1984). Sulla base dei libri del debito pubblico e dell’estimo degli ultimi decenni del XIV secolo la città avrebbe contato
circa 10.000 abitanti, per le vicarie circa 30.000 anime e per le Sei Miglia una popolazione di 9.000 unità, per un totale quindi poco meno di 50.000 abitanti (Pinto, 1988).
All’inizio del 1400 i pivieri (la campagna) si trovavano in uno stato di abbandono e di spopolamento dovuto alle guerre e alle pestilenze, tale da indurre l’emanazione di un decreto che premiava con l’immunità fiscale per dieci anni tutti i forestieri che avessero accettato di coltivare e abitare le Sei Miglia. Ben presto i forenses, pistoiesi, pisani, fiorentini, napoletani, tudertini e albanesi, si sostituirono ai lucchesi. Sempre in questo periodo, l’apporto di stranieri d’altro tipo riguardava la città di Lucca, pare che, secondo un vecchio studio intitolato ‘Le schiave orientali in Italia’, le schiave in Lucca fossero molto numerose; ed infatti il numero degli infanti abbandonati, che venivano raccolti ed esposti all’ospedale di San Luca, erano, secondo i vecchi registri dello spedale, per la maggior parte figli di schiave (Leverotti, 1984). Soprattutto dopo la pestilenza del Quattrocento la popolazione dello stato di Lucca dovette subire un calo. Se per la città appare improbabile un’ulteriore flessione rispetto ai 10.000 abitanti della fine del Trecento, che rappresentavano di già un limite molto basso, si può invece supporre per il contado un calo ulteriore del 10-20% che avrebbe portato la popolazione intorno al terzo decennio del XV secolo a circa 30-35.000 abitanti (Pinto, 1988).
Lucca alla metà del XV secolo contava da 6.000 a 10.000 abitanti. La ripresa delle città toscane arriva tardivamente Lucca, invece, vive un vivace recupero e nel 1540 torna ad avere circa 20.000 abitanti (Ginatempo et al., 1990). L’ipotesi di 40-45.000 abitanti per lo stato lucchese all’inizio del Quattrocento sembra accettabile (Pinto, 1988). Sebbene l’incompletezza delle fonti non consenta di seguire nitidamente lo sviluppo demografico della città e del contado è certo che nella prima metà del Cinquecento la popolazione cresce a seguito dell’impulso delle attività mercantili cittadine tanto che intorno alle antiche mura si moltiplicano le abitazioni e la città si distende nei borghi. Nel 1540 la ‘summa buccharum’ conta più di 18.000 unità in città e quasi 32.000 nelle Sei Miglia (Berengo, 1962; Pinto, 1988).
Verso la metà del XVI secolo, malgrado la ripresa quattro-cinquecentesca, la Toscana esercitò un ruolo secondario nello sviluppo demografico ed economico italiano dell’età moderna registrando un popolamento urbano pressoché stabile rispetto a quello in crescita delle città del Nord, di Roma e delle ‘capitali’ del Mezzogiorno, ed infatti gli abitanti di Lucca si aggiravano ancora intorno a 15-20.000 (Del Panta et al., 1996; Ginatempo et al.,
1990). In generale l’incremento della popolazione italiana in questi secoli va un po’ a singhiozzo, si ricorda infatti la crisi tra Cinque e Seicento (l’ondata di peste del 1629-31 coinvolse anche Lucca). Prenderà a crescere in modo continuativo a partire dalla seconda metà del XVII secolo, una lunga fase di lenta evoluzione caratterizzata da un inizio di modernizzazione del tessuto economico sociale. (Del Panta et. al., 1996).
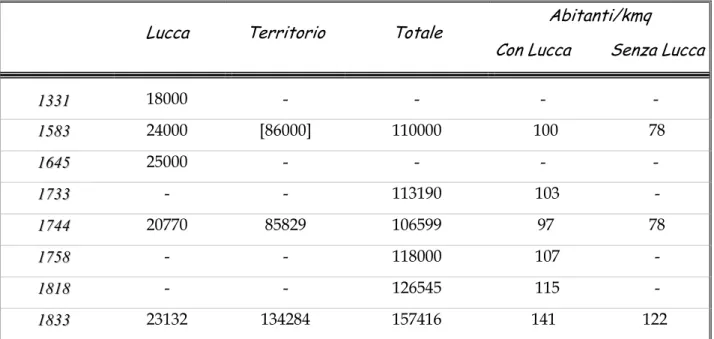
Lo sviluppo demografico a partire dal XVI secolo della Repubblica di Lucca è riportato nella tabella 1.2a tratta dal lavoro dello storico Beloch. La peste del 1630 deve aver fatto 15.000 vittime nel contado, pertanto nel corso del XVII secolo non può essersi verificato un incremento della popolazione, tutt’al più saranno state recuperate perdite. Di conseguenza la popolazione avrebbe compreso già verso il 1600 un numero di circa 85.000 abitanti e inclusa la capitale circa 110.000.
Da questa breve panoramica ci si rende conto di quanto il territorio lucense, sin dai tempi antichi, ebbe una compagine umana variegata, plasmata dal determinismo degli eventi naturali e dalle azioni umane.
Lucca Territorio Totale Abitanti/kmq
Con Lucca Senza Lucca
1 1333311 18000 - - - - 1 1558833 24000 [86000] 110000 100 78 1 1664455 25000 - - - - 1 1773333 - - 113190 103 - 1 1774444 20770 85829 106599 97 78 1 1775588 - - 118000 107 - 1 1881188 - - 126545 115 - 1 1883333 23132 134284 157416 141 122
1.2.1 Demografia toscana tra XIV - XX secolo
Nel periodo preso in considerazione, l’evoluzione demografica della Toscana, è simile a quella dell’Italia, e salvo alcune particolarità, a quella del resto d’Europa (Corsini, 1988).
La popolazione Toscana dei primi del Trecento supera certamente il milione di abitanti. Intorno al 1430 ha circa 420 - 430.000 abitanti. Per quasi cinquant’anni i livelli demografici non subirono modificazioni sostanziali, solo nell’ultimo terzo del XV secolo i segni della ripresa cominciarono a manifestarsi in modo univoco in tutte le parti della regione (Pinto, 1988). Seguì nel XVII secolo un periodo di ristagno, circa 700.000 persone, e il successivo lungo processo di crescita demografica, accelerato a partire dalla seconda metà del XVIII secolo - ai primi dell’Ottocento supera il milione, agli inizi del Novecento è intorno ai due milioni e mezzo - fino a buona parte del XX e in fase di visibile rallentamento in quest’ultimo periodo. Oggigiorno la popolazione è di circa tre milioni e mezzo (Corsini, 1988).
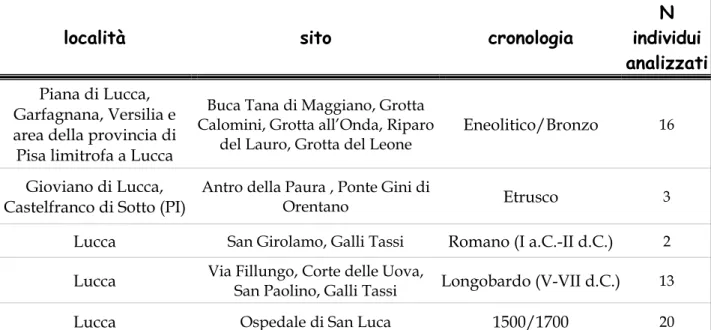
1.3 Provenienza dei campioni osteologici. Qualche considerazione Eneolitico, Bronzo: Grotte
I resti scheletrici preistorici provengono dalle grotte della regione della bassa Versilia, della Garfagnana e del pisano e sono nel complesso attribuibili a genti vissute a cavallo tra la fine del Neolitico e l’inizio dell’Età dei Metalli. Si tratta di materiale piuttosto frammentario che dal punto di vista morfologico ha rilevato elementi di eterogeneità antropologica della regione, dato che non meraviglia visto l’ampio spazio cronologico preso in considerazione. Il Formicola (1980) riscontrò differenze nei caratteri morfologici dei reperti ossei rinvenuti nella zona costiera e in quella interna, tanto da ipotizzare una persistenza di modelli più antichi nelle valli interne poco accessibili alle forme rinnovatrici che si manifestavano nei siti costieri. La mobilità di questi pastori cercatori di metallo invece, sembra essere la chiave per interpretare la presenza di elementi diversi, il substrato mediterranoide di tipo antico che testimonia il contributo di popolazioni paleo-mesolitica e la presenza di caratteri nuovi giunti probabilmente da vie di terra o di mare (Mallegni, 1985). Non conoscendo la corretta successione cronologica delle varie stazioni non c’è la possibilità di cogliere eventuali
cambiamenti di popolazione o di conoscere se e quando si sono avute infiltrazioni di nuovi tipi umani.
Lo studio del materiale osteologico, frammentario e non sempre in un buon stato di conservazione, ha suggerito la presenza di un substrato mediterranoide di tipo arcaico quale testimonianza di vecchie popolazioni di ascendenza paleo-mesolitica già presenti su tutto il territorio italiano (Mallegni, 1985). In particolare si riscontra un’affinità con i neolitici della Liguria anche se nella Toscana centrale il fenomeno è mascherato dalla presenza di maggior brachicefalia dovuta ad apporti genetici di popolazioni molto probabilmente dell’Italia meridionale (Mallegni, 1985).
Buca tana di Maggiano (colline di Maggiano, Lucca): è il giacimento eneolitico più importante dell’entroterra lucense (Mencacci et al., 1976), costituito da una cavità naturale utilizzata come luogo di sepolture collettive (Cocchi Genick et al., 1989). La grotta venne usata come sepolcreto da genti appartenenti alla cultura di Remedello (età del Rame) e in possesso della cultura di Lagozza (Neolitico finale, seconda metà del IV millennio a.C.) (Corazza, 1969). Il deposito di ossa caotiche venne attribuito all’eneolitico (Puccioni, 1914). Il materiale osteologico presenta strette somiglianze con resti eneolitici particolarmente con quelli della vicina grotta di Agnano (Paoli, 1969). Prevalente dolicomorfia (cranio stretto e allungato in senso antero-posteriore), bassa statura e struttura gracile. Reminescenze mesolitiche potrebbero esser state ereditate dalle più antiche popolazioni liguri (Parenti et al., 1960). Anche la distribuzione dei gruppi sanguigni del sistema ABO è risultata simile a quella del campione di Agnano (Grotta del Leone) che si discosta dalle distribuzioni della Toscana e dell’Italia (Paoli, 1969). Un recente riesame dei materiali archeologici ha individuato una frequentazione anche medievale che probabilmente ha inquinato e alterato il sito (Ciampoltrini, 2000).
Grotta all’Onda (Versilia. Camaiore, Lucca): cavità naturale utilizzata anche a scopo abitativo per stanziamenti occasionali (Cocchi Genick et al., 1989), ha restituito industria musteriana, ceramiche neolitiche, eneoliteiche e dell’età del bronzo (Formicola, 1980; Amadei, 1985). Frequentazione pressoché ininterrotta dalla preistoria sino all’età del Bronzo, sporadica in età storica. Secondo lo studio delle industrie litiche la frequentazione è ridotta
nel paleolitico e invece più evidente in età eneolitica; tra le due culture esiste una netta separazione stratigrafica (Graziosi, 1931).
Grotta Calomini (Garfagnana, Lucca): cavità naturale utilizzata per sepolture collettive (Cocchi Genick et al., 1989). Sui reperti scheletrici si riconoscono elementi di arcaicità e tratti di tipo paleomediterraneo che ricordano soprattutto i neo-eneolitici liguri (Formicola, 1980).
Riparo del Lauro (Versilia. Candalla, Lucca): il riparo sottoroccia fu utilizzato a scopo abitativo per stanziamento occasionale; riferibile al Bronzo medio (Cocchi Genick, 1987). Gli insediamenti di Candalla furono scoperti dal gruppo archeologico di Camaiore nel 1981 e pare risalgano all'età del Bronzo (da 3.500 a 3.000 anni fa), ma la troviamo annoverata anche tra i siti eneolitici della provincia di Lucca (Cocchi Genick et al., 1989).
Grotta del leone (Agnano, Pisa): ritrovata industria litica del Paleolitico superiore, pochi strumenti in osso e in metallo inquadrabili nell’ambito dell’età dei metalli o in un momento iniziale di essa (Radi, 1985), utilizzata nell’Eneolitico per sepolture collettive (Cocchi Genick et al., 1989). Le ossa umane sono state rinvenute nello strato intermedio (Eneolitico) del deposito. Molte erano incluse nella stalagmite che separava lo strato superiore da quello medio; ciò fa pensare che esse siano rimaste in superficie dopo che la grotta cessò di essere frequentata dagli uomini dell’Eneolitico. Se ciò fosse vero si dovrebbe escludere una vera e propria inumazione a favore di una semplice deposizione dei corpi. Secondo Parenti (1960) il gruppo presenta un insieme di caratteristiche, probabilmente arcaiche, che si discostano dal tipo mediterraneo classico e ipotizza una possibile forma di ‘scivolamento’ di geni dalle popolazioni mesolitiche della Liguria a quelle successive. Da una recente revisione del materiale osteologico è emerso un tipo fisico unitario senza elementi intrusivi e una significatività e/o endogamia degli individui esaminati (Turbiani, 2005).
Secondo un recente lavoro di Cocchi Genick (2002) le sepolture collettive in grotta sarebbero il riflesso di un cambiamento avvenuto nelle maglie del tessuto sociale e tracciano una linea di separazione con il precedente Neolitico. Nel corso del III millennio si assiste ad
uno sviluppo delle pratiche funerarie associate ad elementi magico-rituali e da un flusso di scambi di conoscenze e di esperienze, attivato in gran parte dalla richiesta di beni e armi metalliche che attestano le acquisite capacità metallurgiche.
Periodo Etrusco
Villaggio Etrusco di Ponte Gini di Orentano, località del comune di Castelfranco di Sotto (Pisa) all’estrema punta di ponente della Valdinievole, ad un passo da Lucca, Montecatini terme e la Versilia.
Antro della paura di Gioviano (Lucca): nel 1974 scoperti all’interno dell’antro resti umani, resti di fauna, frammenti fittili etruschi e liguri databili tra VI e III sec a.C.
Periodo Romano: Lucca, sepolture intramoenia
Complesso San Girolamo. Il campione osteologico proviene da una sepoltura femminile scoperta nel terrapieno annesso alle mura del complesso (fine I secolo a.C.).
Complesso Galli Tassi. Sepolture riferibili all’età Romana (II secolo d.C.).
Alto Medioevo, periodo longobardo: Lucca, sepolture intramoenia Via Fillungo
Corte delle Uova Complesso Galli Tassi
Non si sono ottenute dagli archeologi informazioni riguardo a queste sepolture, se non la collocazione temporale (V – VIII sec. d.C.).
Secoli XVI-XVIII : Lucca, sepolture intramoenia, l’Ospedale di San Luca
La posizione di Lucca, alla quale facevano capo le grandi vie romee per le quali affluivano forestieri e numerosi pellegrini che si recavano a Roma, e la facilità con cui divampavano le epidemie, favorirono il sorgere di opere di carità e istituti di beneficenza tra i quali l’Ospedale di San Luca. Il titolo di ospedale non stava sempre ad indicare il luogo per la cura dei malati, ma dove si offriva assistenza e ospitalità. Le prime notizie certe relative all’Ospedale risalgono all’atto di fondazione concesso dal Vescovo Enrico a Bonansegna Rettore e agli operai dello Spedale il 27 settembre 1262.
Il fiorire di istituti assistenziali avvenivano per opera di Compagnie e Confraternite, non sempre di carattere prettamente religioso, come l’Arte dei mercanti lucchesi che si occupava non solo di cose inerenti la sua istituzione, ma si prodigava nell’assistenza ai più bisognosi, assumendo il patronato e la direzione dell’ospedale detto della Misericordia prima e di San Luca poi. La prima sede dell’ospedale sorse contemporaneamente alla chiesa di San Luca, così com’era l’usanza dell’epoca in modo tale che i malati potessero beneficiare del conforto spirituale; la chiesa ebbe molta importanza e fu nel corso della storia ingrandita e restaurata nel 1600, fu però distrutta nel 1912 (Torchiana, 1954). L’Ospedale fu il più importante centro d’assistenza della provincia sino in epoca moderna. Durante il XVI sec. al San Luca si accorparono numerosi altri ospizi e nel 1563 l’Ospedale si dotò di un collegio medico.
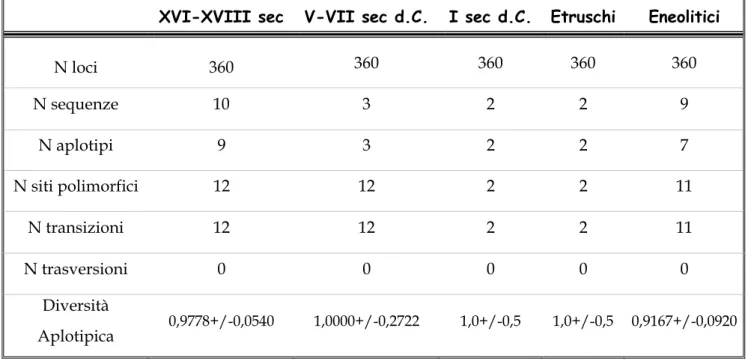
Le sepolture rinvenute nella chiesa sono 10. Il numero minimo di individui è 31 di cui 5 infantili.
1.4 Finalità del lavoro
L’ indagine è un contributo alternativo che s’intende dare alla ricerca multidisciplinare sul problema delle origini e della storia di Lucca che, come scrisse S. Ferri nel 1961, si perdono nel confuso ginepraio delle civiltà antiche. Il filo conduttore della tesi è la biodiversità umana che si esprime nel binomio genotipo-fenotipo, e la vita che conserva la traccia degli esperimenti evolutivi. Gli aspetti paleoantropologici indagati sono la dieta e la genetica di alcuni gruppi umani che abitarono o transitarono nell’area di Lucca. Le analisi di paleogenetica sono state condotte presso il Laboratorio di Antropologia Molecolare dell’Università di Firenze e quelle di paleonutrizione presso il Laboratorio di Paleonutrizione dell’Università di Pisa.
Sono stati prelevati campioni d’osso di individui provenienti da sepolture intramoenia e siti fuori dalle porte di Lucca localizzati nel territorio circostante. Si è cercato di coprire nel miglior modo possibile la scala temporale raggiungendo un numero di 54 individui campionati su di un totale di 79 reperti rinvenuti; cioè è stata fatta una scelta che desse garanzia di risultati sulla base dello stato di conservazione del materiale. L’intento è quello di ricostruire, per quanto possibile, alcuni aspetti antropologici del territorio ripercorrendo la scala del tempo, dall’epoca moderna sino alla fumosa preistoria, immergendosi il più possibile nel quadro ambientale e nel contesto storico-culturale dei vari periodi considerati e
prendendo in considerazione l’andamento demografico e gli eventi che presumibilmente influirono sul pool genetico della popolazione del territorio.
Alcuni elementi chimici presenti in traccia nell’osso sono considerati elementi guida per la ricostruzione delle abitudini alimentari delle popolazioni antiche. La spettroscopia ad assorbimento atomico è la tecnica d’indagine che si è utilizzata per l’analisi paleonutrizionale. Gli elementi indagati sono il calcio, quale parte integrante della matrice ossea, zinco e stronzio, in quanto sono quelli maggiormente discriminanti del tipo di dieta, rame e magnesio. Il fine è quello di tracciare il quadro alimentare delle popolazioni dell’area lucchese nel corso del tempo, e confrontarlo con l’andamento delle linee evolutive della storia dell’alimentazione italiana dall’antico ai tempi più recenti.
Per quanto riguarda l’aspetto paleogenetico, che ha avuto un peso significativo nel corso dei tre anni di dottorato in termini di tempo, di know how acquisito e impiegato, si è indagata una particolare regione del DNA (acido desossiribonucleico) mitocondriale. Il DNA è un concentrato di istruzioni unico in natura che gli individui mantengono quasi costante e che si trasmette da una generazione a quella successiva.
Il DNA, che costituisce il genoma, conferisce all’uomo e a tutti gli organismi viventi la capacità di riprodurre se stessi. Nella riproduzione interviene la casualità per cui compaiono le mutazioni che, se non sono prive di conseguenze, innescano il meccanismo della selezione, il cambiamento registrato nella doppia elica potrà essere ereditato dalle generazioni successive; il fenomeno è automatico ed inevitabile e, generazione dopo generazione, determina la trasformazione della specie. Ma è il ritmo dell’accumularsi delle mutazioni neutre su cui si basa il concetto di orologio molecolare fondamentale per determinare i tempi della divergenza tra specie. La filogenesi descritta dalle molecole è il rapporto di parentela antenato-discendente che si basa sulle mutazioni neutrali alla selezione naturale. Il DNA residuo recuperato dai resti antichi rappresenta una fonte che racconta una storia iniziata più di 100 mila anni fa, il cui studio rappresenta un diverso punto di vista da dove osservare il nostro passato.
Oltre al DNA contenuto nei cromosomi, tutti gli organismi superiori sono dotati di un pacchetto suppletivo di informazione genetica sottoforma di piccole molecole circolari di DNA, che nell’uomo si trova nei mitocondri. La caratteristica genetica fondamentale dei geni extracellulari è che, essendo dislocati nel citoplasma cellulare, vengono ereditati solo per via materna, quindi in modo non mendeliano. Inoltre l’assenza di ricombinazione, l’elevato tasso
di mutazione e la presenza in numerose copie per ogni cellula, per cui lo si può recuperare facilmente anche in campioni antichi degradati, fanno si che il DNA mitocondriale sia lo strumento più usato negli studi di evoluzione. Ed infatti, il piccolo anello genomico, ed in particolare la regione ipervariabile HVR-I sono l’oggetto d’indagine del presente studio.
La novità di questo progetto sta nell’innovativo approccio molecolare all’antropologia, e in seno a questo, il fatto di aver utilizzato un programma di simulazione di coalescenza. Il software permette di simulare scenari demografici complessi e di includere campioni antichi nel passato ed eventi storici, quali quelli di bottleneck, che probabilmente hanno influenzato il patrimonio genetico delle popolazioni; il tutto confrontato con i valori osservati ottenuti dalle sequenze della regione ipervariabile del DNA mitocondriale. Questo approccio moderno fa fronte a due fondamentali problemi, il primo è che l’esperimento evolutivo non è replicabile, è una ‘corsa’ di sola andata, e secondo che le condizioni iniziali dell’esperimento sono sconosciute (Rosemberg et al., 2002).
Il fatto che il materiale più antico abbia dato dei risultati è un presupposto per indagare certi problemi di popolamento in una maniera del tutto nuova; ciò potrà permettere una risposta alle tante domande su questo aspetto relativo all’umanità del territorio lucchese, che, come è noto, è sempre stato contraddistinto da percorsi viari che lo hanno messo in comunicazione sia con il Nord che con il Centro Italia. Sin dai tempi dei romani il sistema viario di fondovalle da essi concepito, costituì un’alternativa vantaggiosa alla viabilità trasversale di cui si servirono i popoli protostorici, definendo la centralità dell’area lucense e della città di Lucca. Il sistema viario si dimostrò, tra l’altro, determinante per la conquista delle popolazioni italiche, permettendo attraverso le valli l’accesso militare e commerciale (Riparbelli, 1982). I tracciati romipeti, la via francigena, le vie garfagnine e pedemontana delle alture pisane sono stati capaci di favorire un apporto umano anche differenziato, il quale è poi l’aspetto che in definitiva la ricerca vuole evidenziare. Si cercherà di contribuire alla ricostruzione della storia ‘antropologica’ della lucchesia sfruttando parte delle informazioni che l’archivio biologico costituito dallo scheletro è capace di restituirci.
L’intento è abbastanza arduo, sino al punto al quale siamo riusciti è già buona cosa, soprattutto per quanto riguarda la paleogenetica che richiede dovizia di tempo e attenzione.
2 ANTROPOLOGIA MOLECOLARE
2.1 La rivoluzione dell’antropologia molecolare
La natura delle differenze tra due o più popolazioni va ricercata nella matrice biologica e nell’ensemble culturale. I caratteri biologici seguono una trasmissione solo verticale vale a dire dai genitori ai figli, mentre quelli culturali hanno anche una diffusione orizzontale tra individui non consanguinei. Le differenze culturali legate all’apprendimento sono più facilmente soggette a cambiamenti rispetto a quelle genetiche, cioè quelle ereditate biologicamente. Le differenze morfologiche tra gli individui sono in parte di origine genetica e in parte determinate dall’ambiente in cui vivono. Alcuni fenotipi, quali il colore della pelle, la forma del naso, l’altezza e in generale la struttura scheletrica, rappresentano una fedele descrizione dell’adattamento dei gruppi umani alle diverse situazioni climatiche. Pertanto i caratteri antropologici mostrano l’azione selettiva dei diversi ambienti in cui l’uomo anatomicamente moderno s’è imbattuto durante il suo cammino di diffusione sulla superficie terrestre. L’adattamento fu biologico e pure culturale, sviluppando nel corso di migliaia d’anni una differenziazione in entrambi i contesti. Nello studio dell’evoluzione dell’uomo associare i cambiamenti evolutivi a quelli climatici non è cosa semplice visto la scarsità delle testimonianze fossili, per cui non si è riusciti a definire con precisione l’origine e l’estinzione delle specie di ominini che compaiono nel cespuglio evolutivo. I nostri avi sotto il profilo ecologico dovevano essere poco specializzati e perciò dovevano essere capaci di tollerare un’ampia rosa di condizioni climatiche diverse. Si può dunque pensare che l’uomo abbia colonizzato i vari ambienti indipendentemente dai caratteri selettivamente vantaggiosi, i quali erano compatibili con l’ambiente, ma solo successivamente si sarebbero rilevati vantaggiosi. Le evidenze morfologiche, culturali e/o archeologiche registrate nelle popolazioni attuali e in quelle antiche mostrano notevoli dissomiglianze tra le popolazioni, c’è da chiedersi se a queste differenze corrisponde un adattamento biologico, cioè mutazioni nel corredo genetico, di pari entità. La risposta è che siamo una specie giovanissima e che, malgrado le diversità visibili, a guardare l’invisibile, e cioè i geni, si scopre una grande omogeneità tra le popolazioni anche molto distanti geograficamente. Non solo, a partire dagli anni ’60 del XX secolo l’approccio molecolare all’antropologia rivela una parentela stretta tra uomo, scimpanzé e un po’ più distante anche con il gorilla.
La disciplina di cui mi occupo, l’antropologia molecolare, prende vita ufficialmente nei primi anni ‘60 e fu così denominata da Emile Zuckerland che per la prima volta lancia l’idea dell’ ‘orologio molecolare’. Si ricorda però che sin dai primi del Novecento iniziarono gli esperimenti di antropologia molecolare, ma nulla ancora si sapeva della molecola responsabile della trasmissione dei caratteri ereditabili. Le prime intuizioni si ebbero negli anni ‘40 e alla molecola venne dato il nome di DNA (acido desossiribonucleico), ma com’è noto, della sua struttura a doppia elica se ne seppe nel ’53, ed è solo dagli anni ’70 -’80 che la biologia molecolare ha strumenti potenti ed efficaci per lo studio di questa importante macromolecola. La storia dell’approccio molecolare allo studio dell’evoluzione è fatta di false partenze e timidi tentativi. Sicuramente il lavoro destinato a scuotere il campo delle scienze antropologiche, e che ha fatto letteralmente storia, è ‘Mitochondrial Dna and human
evolution’ che venne pubblicato su Nature nel 1987 da Allan Wilson e collaboratori. La fonte
microscopica ricercata e interrogata dagli antropologi molecolari è il DNA mitocondriale nelle diverse popolazioni del globo, che secondo gli studiosi deriverebbe da un’unica genitrice africana vissuta 200.000 anni fa.
Il principio su cui si basa l’antropologia molecolare è che il patrimonio genetico di una cellula che va incontro a divisione, viene copiato e trasmesso a ogni cellula figlia. A ogni passaggio però sono possibili errori di copiatura che alterano la sequenza delle basi lungo il filamento di DNA. Le mutazioni si accumulano di generazione in generazione; pertanto le differenze tra le popolazioni sono tanto più numerose quanto più lungo è il tempo trascorso da quando hanno avuto l’ultimo antenato comune. Questo processo è fondamentale per capire l’utilità dei marcatori molecolari per determinare le distanze evolutive.
I messaggeri fisici dell’ereditarietà sono i cromosomi, complessi di DNA e di proteine. Il DNA è una molecola complessa a forma di doppia elica, variamente avvolta e ripiegata i cui quattro costituenti elementari sono i nucleotidi (basi azotate): A (adenina), C (citosina), G (guanina) e T (timina). Un piccolo segmento di DNA, il gene, ne costituisce l’unità fondamentale. Molecole di RNA messaggero trascrivono il contenuto dei geni, che verrà tradotto dalle strutture cellulari nelle proteine corrispondenti; a ogni tripletta di basi corrisponde un amminoacido della molecola proteica. La perdita o l’aggiunta di uno o più nucleotidi lungo il segmento può cambiare la funzione del gene corrispondente. Questo cambiamento, detto mutazione, avviene spontaneamente e con bassa probabilità ed è trasmesso alla discendenza.
Il DNA determina e controlla il meccanismo dell’espressione genica cellulare, in quanto contiene le sequenze dei geni e le sequenze che regolano l’on/off di una particolare funzione durante la vita della cellula. La stabilità della molecola, la capacità di resistere ai cambiamenti, di replicarsi esattamente e di trasmettere le informazioni genetiche alle cellule figlie sono le proprietà fondamentali del DNA.
2.1.1 Molecole e fossili a confronto
A partire dagli anni ‘60 le testimonianze fossili macroscopiche fondamentali per ricostruire la storia evolutiva umana sono state affiancate da un nuovo tipo di fonte, il DNA, che conserva la traccia degli esperimenti evolutivi che hanno condotto all’uomo moderno. Sin dagli albori dell’antropologia molecolare il problema fu quello di conciliare la storia registrata nelle molecole con quella raccontata dalle testimonianze fossili e archeologiche. L’approccio metodologico sperimentale degli antropologi molecolari si incontra e si scontra con il linguaggio dell’ antropologia fisica e dell’archeologia nel campo interdisciplinare dello studio dell’evoluzione umana. Il sistema morfometrico della paleoantropologia e il metodo sperimentale dell’antropologia molecolare non sono altro che due orizzonti scientifici il cui fine ultimo è la ricostruzione naturale della storia dell’uomo, pertanto il dialogo tra fossili e molecole è possibile e necessario.
La critica principale mossa dai biologi evoluzionisti alla paleontologia è che quest’ultima è una scienza descrittiva e inferenziale che si basa sulla morfologia comparata e pertanto è soggettiva. A rimarcare questa arbitrarietà sta il fatto che, per quanto riguarda l’evoluzione umana, i fossili che attualmente sono nelle mani degli scienziati sono per lo più frammenti di scheletri, e sono questi la base su cui si tenta di ricostruire le fattezze dell’individuo e quindi i caratteri della specie appartenente. Inoltre i caratteri influenzabili dall’ambiente di vita forniscono una descrizione parziale della storia evolutiva, cioè la storia degli ambienti in cui hanno vissuto e vivono i diversi gruppi umani.
D’altra parte il funzionamento non perfetto degli orologi molecolari, la proposta di ricostruire la storia dell’umanità sulla base di piccole porzioni del corredo genetico, come quella del DNA mitocondriale che tutto intero non rappresenta che un duecentomillesimo del nostro genoma, l’inquinamento della fonte con DNA eterologo e la tendenza al riduzionismo e all’eccessiva frammentazione che perde di vista l’insieme, sono alcune delle critiche mosse
alle indagini molecolari. Una considerazione risaputa, ma che vale sempre la pena ribadire, è che tanto l’indagine sui fossili quanto quella sulle molecole hanno i loro limiti. La ricostruzione dell’evento evolutivo necessita quindi dell’interdigitazione di tutte le informazioni fornite dai diversi campi di studio.
Le evidenze molecolari sono una documentazione che vanno a riempire, per lo meno parzialmente, gli hiatus delle testimonianze fossili che purtroppo lasciano scoperte molte tappe dell’evoluzione delle specie. È bene ricordare che gli studi molecolari non sono in grado di stabilire l’origine genetica delle differenze nei caratteri morfologici osservabili e non possono neppure stabilire se una mutazione che determina un carattere ne influenza anche altri.
Allo stato attuale l’antropologia molecolare è come un fiume in piena che immette nel mare della conoscenza innumerevoli dati. Credo che la chiave di volta per la ricostruzione dell’albero genealogico umano stia proprio nel materiale genetico che rappresenta un bacino d’informazioni, che non sono migliori o più fondamentali di quelle ricavabili dalla morfologia, dalla linguistica o da altre discipline, ma sono abbondanti, e lo saranno sempre più, e quindi paragonabili. Ora come ora non credo che le molecole siano tanto più obbiettive dei fossili perché la loro complessità nasconde insidie e trabocchetti per l’analisi e l’interpretazione delle divergenze evolutive. Ad ogni modo le discrepanze temporali tra i calcoli evolutivi degli antropologi fisici e quelli dei biologi evoluzionisti, che solitamente anticipano i tempi rispetto ai primi, a mio avviso è, entro certi limiti, più che plausibile visto che l’evoluzione molecolare precede necessariamente quella morfologica; la manifestazione del carattere, riconoscibile nelle testimonianze fossili, non è immediata, a volte addirittura le mutazioni possono rimanere silenti e con molto ritardo tradursi in caratteri morfologici (Meldolesi, 1999).
Oggigiorno il recupero e lo studio del DNA da reperti scheletrici e tessuti antichi è la frontiera più avanzata negli studi di antropologia molecolare. Il DNA residuo recuperato dai resti antichi rappresenta una fonte che racconta una storia iniziata molti anni fa, il cui studio rappresenta un diverso punto di vista da dove osservare il nostro passato.
2.1.2 Le molecole come congegni di datazione
La mutazione, un cambiamento del DNA, è normalmente un fenomeno molto raro, ed è necessario che lo sia, essa infatti può essere dannosa all’organismo. Vi sono meccanismi di controllo e di correzione delle copie della doppia elica. Le copie, se eseguite male, vengono corrette con il risultato che le mutazioni sono giustappunto rare.
Due fattori che possono variare nello spazio e nel tempo hanno un ruolo determinante nella storia evolutiva: la selezione naturale e la deriva genetica. La selezione può variare da un gene all’altro in ogni popolazione, mentre la deriva è sempre presente e ha in media la stessa intensità per tutti i geni; le dimensioni demografiche della popolazione determinano la deriva genetica, perciò essa è una proprietà della popolazione stessa, ed è un cambiamento evolutivo indipendente dalla selezione naturale.
Le evidenze sui polimorfismi del DNA mostrano quanto sia grande la variazione genetica intraspecifica, ed è naturale chiedersi quale siano i modi e i tempi con i quali questa variazione si fissa nel genoma. Se si desse un’interpretazione ‘selezionista’ il problema parrebbe risolto introducendo la selezione naturale: le mutazioni si fissano perché danno vantaggio nelle diverse condizioni ambientali in cui vivono le popolazioni. Ma gli innumerevoli dati molecolari registrano una gran quantità di mutazioni, esse sono decisamente troppo numerose per poter essere tutte favorevoli. Questa fondamentale intuizione la si deve a Mooto Kimura (1970) il quale osservò che la gran parte delle mutazioni è neutrale dal punto di vista della selezione naturale; il destino di queste variazioni all’interno della popolazione è lasciato alla volontà del caso, ai processi stocastici, ovvero alla deriva genetica. Una mutazione neutrale è una variazione equivalente dal punto di vista selettivo, il fatto che un allele sia più diffuso rispetto ad altri non vuol dire che sia automaticamente quello più vantaggioso. L’introduzione della ‘teoria neutrale dell’evoluzione molecolare’ ha ridimensionato il ruolo della selezione naturale, sino a supporre che l’evoluzione si attui prevalentemente per mezzo del caso. La teoria neutrale, dunque, scardinò il mito della selezione naturale ridimensionandone l’onnipresenza. Il dibattito tra selezionisti e neutralisti fu acceso e non s’è tuttora spento poiché il postulato di Kimura è alla base del concetto di orologio molecolare e dei ticchettii sottoforma di mutazioni che sono uno strumento preziosissimo nel campo dell’evoluzione, ma la cui affidabilità sembra non essere totale. Tuttavia è unanime l’ammissione che nelle mani
dell’evoluzione ci siano molteplici strumenti, perciò la selezione naturale non esclude la deriva genetica e viceversa. Piuttosto è da considerare a che livello della struttura del vivente esse agiscono; sicuramente la manifestazione del carattere, il fenotipo, è il bersaglio diretto della selezione, mentre le molecole o meglio il genotipo è l’oggetto su cui opera la casualità.
La fissazione casuale delle mutazioni neutrali è il punto focale della teoria, se poi si stabilisce il ritmo con cui esse s’accumulano, diventa un ottimo strumento per misurare il trascorrere del tempo. Le molecole, in particolare il DNA, sono paragonabili a un orologio capace di misurare il passare del tempo in termini di accumulo di mutazioni neutrali, ovvero di variazioni non visibili sotto il profilo selettivo, che si fissano ad una velocità che sembra essere costante almeno nel lungo periodo. La teoria neutrale è la base teorica dell’orologio molecolare; il processo di mutazione è casuale e la fissazione delle mutazioni non deve essere disturbata dalla selezione naturale. Le mutazioni neutrali sono quelle in cui l’imparzialità del caso ha la meglio su qualsiasi criterio di selezione che andrebbe ad alterare la costanza della fissazione delle mutazioni rimuovendole o fissandole a seconda che esse conferiscano handicap o vantaggio agli individui in cui si presentano. Infatti, ad esempio, i geni che determinano i caratteri somatici fortemente soggetti alla selezione per mano dell’ambiente nel quale le popolazioni vivono, non sono in grado di fornirci i tempi necessari per la fissazione di un carattere sensibile a determinate condizioni ambientali.
Dunque appare vera l’intuizione darwiniana per cui i caratteri più utili per ricostruire l’evoluzione sono quelli inutili e insignificanti “… la variabilità (degli organi rudimentali)
sembra derivare dalla loro inutilità, e di conseguenza dal fatto che la selezione naturale non ha avuto la possibilità di frenare la deviazione di struttura”. Gli elementi insignificanti, o in
parte sprovvisti di attività biologica, che oggigiorno l’antropologia molecolare ricerca e studia sono contenuti nei segmenti non codificanti, cioè quelli che non hanno alcuna informazione da trasmettere all’organismo, almeno apparentemente, presenti nel materiale genetico.
Il tempo scandito dall’orologio molecolare fornisce la stima delle separazioni tra specie. Quanto più due popolazioni sono separate tanto più è l’accumulo di mutazioni, ciò vuol dire che il numero di differenze genetiche che separa due popolazioni è direttamente proporzionale al tempo a partire dal quale hanno condiviso lo stesso antenato. Così come il tasso di sostituzione aminoacidica delle proteine viene utilizzato per calcolare i tempi di divergenza tra specie differenti, il tasso di sostituzione nucleotidica del DNA, in particolare
di quello mitocondriale, può essere usato per calcolare il tempo di divergenza tra due linee evolutive. Quindi è possibile, analizzando le differenze nelle sequenze di più individui, risalire non solo all’antenato comune più recente da cui discende l’attuale variazione, ma anche stimare il numero di generazioni che separa l’antenato dal presente.
Trovo necessario e doveroso riflettere sui problemi legati all’uso delle molecole come sistemi di datazione, perché non sono strumenti di precisione ma possono essere corretti e interpretati. In particolare nel prossimo paragrafo (2.1.4) mi occuperò delle grandi potenzialità e dei limiti peculiari di un orologio molecolare molto speciale basato sul genoma dei mitocondri, su cui, per altro, si basa l’indagine genetica che ho condotto sui campioni lucchesi.
L’azione selettiva per ogni gene è diversa e determina perciò per ciascuno di essi ritmi diversi di mutazione. Ne segue che la probabilità che un cambiamento colpisca una sequenza di basi è diversa per ognuna di esse. Ci sono ad esempio delle bande di DNA più o meno corte, ripetute in tandem e non codificanti (mini- microsateliti), un tempo etichettate come DNA spazzatura, che a causa della loro ripetitività durante la replicazione acquisiscono o perdono una unità ripetuta finendo per allungarsi o accorciarsi. E’ stato stimato che nel corso di una generazione è 10.000 volte più probabile assistere a una variazione di questo tipo piuttosto che registrare una mutazione puntiforme nel gene dell’emoglobina che causa l’anemia falciforme (Meldolesi, 1999). Quindi il cambiamento di alcuni siti è legato anche alla probabilità che a mutare siano altri punti del DNA in cui si concentrano le mutazioni multiple, i così detti hot spot, quali le sequenze ripetute e gli pseudogeni. Questi ultimi, definiti come relitti evolutivi, sono copie dei geni originari che però non hanno alcuna funzione codificante; essi cambiano più in fretta dei geni funzionali. Ad esempio lo pseudogene dell’emoglobina ha un ritmo di mutazione 5 volte maggiore a quello del gene funzionante (Meldolesi, 1999). C’è da chiedersi se dietro a quella che appare come mutazione non si celino variazioni non più apprezzabili perché sovrapposte da altri cambiamenti e/o da correzioni da parte di altre mutazioni che ripristinano la sequenza originaria (retromutazioni); o se una sequenza che si allunga non si sia prima accorciata e viceversa. Gli scienziati affrontano il problema apportando correzioni di tipo statistico ai calcoli e scegliendo il gene giusto a seconda degli scopi e del periodo evolutivo che si desidera indagare.
Una delle assunzioni della teoria neutrale è che uno stesso gene mostri un ugual tasso di mutazioni in specie diverse; è stato dimostrato l’esatto contrario e cioè che la velocità di mutazione può essere anche molto diversa. Si giustifica questa differenza con la diversa efficienza dei meccanismi di riparazione del DNA e con i diversi tempi di generazione: in linea di massima più generazioni si traduce in un numero maggiore di processi di replicazione e quindi in più errori di copiatura. La questione è arginata ribadendo che un orologio valido per tutte le specie non esiste. Esistono degli orologi molecolari ad hoc per ogni confronto in determinati intervalli di tempo e di determinati gruppi tassonomici. Un orologio che ticchetti velocemente utile per studiare avvenimenti recenti, come la comparsa dell’uomo anatomicamente moderno, è il DNA mitocondriale, valevole anche per confronti tra scimmie antropomorfe e uomo; mentre per indagare eventi più antichi risalenti a milioni di anni fa è conveniente scegliere come orologio molecolare un gene che cambia lentamente, come ad esempio quello che codifica per l’RNA ribosomiale, usato ad esempio per lo studio della comparsa degli animali moderni.
2.1.3 Marcatori molecolari
La storia biologica delle specie è indagata utilizzando dei marcatori molecolari.
Quando di un gene si osservano due o più forme diverse, chiamate alleli, si parla di polimorfismo genetico. I polimorfismi genetici comunemente definiti classici sono quelli relativi al prodotto dei geni, le proteine (es. sistemi AB0 e Rh +/-), sono marcatori che furono largamente studiati prima che si scoprissero i polimorfismi che permettono lo studio diretto dei geni (DNA). Il vantaggio nell’analisi dei marcatori del DNA sta nel sondare parti di genoma che sono sopravvissute di generazione in generazione sino ad oggi. Tale tramandare consente di tratteggiare le linee evolutive che da un passato più o meno remoto sono giunte al presente.
Tra i marcatori molecolari negli studi sul DNA antico si ricorda il DNA mitocondriale quale principale strumento d’indagine, riferendosi specialmente alla regione non codificante
D-loop e a quella del gene che codifica il Citocromo C ossidasi sub unità II, e il DNA
nucleare, in particolare gli STRs (microsatelliti) autosomici e sessuali. Il DNA mitocondriale umano è costituito da regioni codificanti e regioni neutre non codificanti. La regione codificante rappresenta la frazione maggiore e contiene i geni per rRNA, tRNA e per
proteine. La D-Loop (displacement-loop) è una regione che consiste in circa 600 paia di basi, di essa fanno parte le regioni ipervariabili HVRI e HVRII, è localizzata vicino all’origine della molecola, non codifica per alcuna proteina ed evolve con velocità maggiore rispetto alle regioni codificanti per cui è un mezzo d’indagine efficace per studiare avvenimenti evolutivi. Le strutture respiratorie del mitocondrio contengono numerosi trasportatori di elettroni come i citocromi, che sono proteine contenenti un gruppo prostetico eme che trasporta elettroni. Il citocromo C, evolutosi circa 1,5 miliardi di anni fa, è poco variabile ed è presente in tutti gli organismi che possiedono la catena respiratoria mitocondriale (Stayer, 1989).
Nelle regioni non codificanti del genoma animale e vegetale sono presenti corte sequenze ripetute in tandem chiamate STR (Short Tandem Repeat) o più comunemente microsatelliti. Sono delle vere e proprie balbuzie sparse su tutto il genoma. Essi marcano il 20% del genoma umano e si trovano sia nei cromosomi autosomici che in quelli sessuali. Gli STRs autosomici consentono la tipizzazione genetica dell’individuo. Quelli sessuali sono utili per la determinazione del sesso e per lo studio di filogenesi per via paterna, i microsatelliti del cromosoma X materno sono ereditati da entrambi i sessi, mentre gli STRs del cromosoma X paterno sono ereditabili solo dalle figlie e quelli del cromosoma Y sono esclusivamente trasmissibili di padre in figlio. Gli STRs sono un buon metodo di datazione genetica: la differenza media tra il numero di ripetizioni di due popolazioni tende a crescere all’ aumentare del tempo di separazione tra le popolazioni (Cavalli Sforza, 1996).
Il DNA contenuto nei mitocondri è una piccola porzione di genoma oggetto di molte attenzioni da parte degli studiosi di genetica di popolazione, poiché le sue caratteristiche quali l’eredità di tipo materno, l’assenza di ricombinazione, l’elevato tasso di mutazione e la presenza in numerose copie per ogni cellula per cui lo si recupera facilmente anche in campioni degradati, lo rendono lo strumento più estesamente e proficuamente utilizzato nello studio dell’evoluzione della nostra specie e per comprendere la dinamica della diffusione nel mondo e i rapporti tra le varie popolazioni.
2.1.4 Caratteristiche e potenzialità del DNA mitocondriale
Secondo l’ipotesi ‘simbiontica’ di Margulis (1970) i mitocondri derivano da batteri che in un tempo remoto assunsero rapporti simbiotici con cellule eucariotiche ancestrali; l’idea che il primordiale ospite e inquilino fossero rispettivamente un protobatterio e un archeobatterio fu via via confermata da prove molecolari, tant’è che alla fine degli anni novanta il confronto tra i geni dell’ RNA ribosomiale di ceppi batterici con quelli dei mitocondri suggerì che il batterio Rickettsia prowazekii, famoso nella storia umana perché è colui che causa il tifo, fosse il più simile ai mitocondri (Anderson et al., 1998). I mitocondri sono organelli citoplasmatici che hanno mantenuto una certa autonomia rispetto alla cellula, infatti, hanno un loro anello a doppia elica di DNA e i ribosomi per la sintesi proteica. Essi sono indispensabili alla cellula perché forniscono energia attraverso la fosforilazione ossidativa, una funzione fondamentale visto che l’energia liberata dalla rottura dei legami fosforici delle molecole di ATP sostiene gran parte dei processi biochimici fondamentali per la vita.
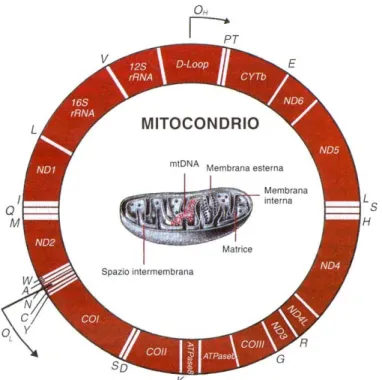
Il DNA contenuto nei mitocondri (mtDNA) è il marcatore genetico più utilizzato dagli antropologi e genetisti di popolazione per comprendere la storia evolutiva dell’uomo. E’ peraltro il genoma eucariotico più conosciuto che è stato sequenziato in numerosi organismi e ha caratteristiche che lo rendono particolarmente adatto allo studio dei resti antichi. Il DNA mitocondriale umano si presenta sottoforma di una breve doppia elica chiusa ad anello costituita da 16.569 coppie di basi che corrispondono allo 0,0006% del genoma totale. L’anello è costituito da due filamenti denominati H (heavy) e L (light) per la loro differente costituzione nucleotidica: il filamento H contiene in prevalenza basi puriniche A e G (di peso molecolare maggiore), il filamento L basi pirimidiniche C e T (di peso molecolare minore) (Fig. 2.1.4a).
Oltre ad essere breve, a differenza del DNA nucleare, quello mitocondriale è piccolo e organizzato in maniera parsimoniosa difatti è privo di complicanze quali introni e sequenze ripetute. Le porzioni non codificanti sono estremamente ridotte, il 7% a fronte del 95% del DNA cromosomico; una di queste è la così detta ‘regione di controllo’ dove ha inizio la replicazione e la trascrizione della molecola. In ogni cellula sono presenti migliaia di copie di mtDNA e solo due copie di quello nucleare, questa abbondanza consente di recuperare la molecola anche da campioni particolarmente depauperati e incompleti. Nel 1981 Anderson e collaboratori presso il MRC Laboratory of Molecular Biology di Cambridge ottennero da un
soggetto di nazionalità inglese la sequenza completa del mtDNA umano. La sequenza di riferimento universalmente usata è convenzionalmente riferita alla catena L
(CRS-Cambridge Reference Sequence).
Le caratteristiche strutturali della molecola ne consentono una facile estrazione, purificazione ed analisi, ma la peculiarità che determina l’importanza del mtDNA come strumento nel campo dell’evoluzione molecolare è l’ereditarietà di tipo uniparentale. Il genoma mitocondriale dei mammiferi ha una trasmissione di tipo materno, il piccolo genoma circolare durante la fusione di ovocita e spermatozoo non va incontro a crossing
over come i cromosomi perché il gamete maschile apporta allo zigote solo materiale
cromosomico, mentre l’ovocita contribuisce non solo con il materiale cromosomico ma anche con l’intero citoplasma. Risulta quindi che il mtDNA della prole è identico a quello della madre, che è uguale a quello della nonna e così via. Pertanto per ogni famiglia è possibile risalire indietro nel tempo lungo la linea di ascendenza femminile. Tanto più indietro si va nell’albero genealogico tanto più la parentela si allarga sino a che solo poche donne possono rappresentare l’umanità vivente. Il tasso di evoluzione del mtDNA umano, ovvero il processo di accumulo di mutazioni, è di una o due mutazioni puntiformi ogni cento nucleotidi per milione d’anni, ed è da dieci a venti volte più veloce del tasso di evoluzione dei geni del DNA nucleare; in particolare le regioni non codificanti dell’anello (es. D-loop) hanno un tasso di divergenza (numero medio di mutazioni in una popolazione) da uno a quattro-cinque volte maggiore rispetto a quelle codificanti. Il ticchettio è così veloce perché il sistema di riparazione non è così efficace e il numero dei cicli di replicazione è maggiore. Ci si attende quindi che la popolazione ancestrale presenti la massima quantità di mutazioni, ed infatti la maggiore eterogeneità del DNA mitocondriale africano indica che ha accumulato variazioni più degli altri perché più lunga è stata la sua storia evolutiva; una prova in più a favore dell’ Africa come origine dell’umanità (Biondi et al., 2004). Fu proprio lo studio del DNA mitocondriale e la sua caratteristica di ‘orologio molecolare’ a far scaturire l’ipotesi della Eva nera che avallò la teoria dell’origine monocentrica e africana dell’umanità intorno ai 200.000 anni fa.
L’mtDNA è inoltre un buon indicatore di eventi di variazione genetica dovuta a fluttuazioni casuali, a colli di bottiglia, ad effetto del fondatore e a colonizzazioni, eventi cioè indipendenti dall’attività della selezione naturale, perché la popolazione che contribuisce alla generazione successiva per l’mtDNA è ¼ di quella del genoma nucleare. Questi eventi
favoriscono la differenziazione locale e sono tanto più visibili quanto più sono ridotte le dimensioni della popolazione (Biondi et al., 2004).
In questo tipo d’analisi si parte dal presupposto che in un individuo tutte le copie di mtDNA siano uguali tra loro (omoplasia), ma l’eteroplasmia, seppur rara, è stata evidenziata negli organismi, primo tra tutti è stato il caso dello zar di Russia Nicolai II. Il motivo della presenza di sequenze diverse di mtDNA in uno stesso individuo non è ancora chiaro ma l’ipotesi al momento formulata è che mitocondri paterni riescano ad entrare al momento della fecondazione nell’uovo ricombinando il loro materiale genetico con quello materno. È bene inoltre ricordare che l’orologio mitocondriale non è metronomico, o più semplicemente svizzero, le lancette non girano sempre in modo regolare. Quest’imperfezione non può che far correre il pensiero alla teoria di Kimura e quindi al carattere neutrale della regione di controllo del mtDNA che invece, alla luce di quanto detto, potrebbe non essere inviolata dai processi selettivi. Questa imperfezione è causata dal fenomeno delle mutazioni multiple negli
hot spot (cfr. 2.1.2) per cui nel breve periodo il tasso di mutazione sembra accelerare, pare
invece rallentare in tempi più lunghi. Gli studiosi assicurano che la media dei ticchettii è abbastanza regolare tanto da poter fare stime temporali affidabili sul lungo periodo.
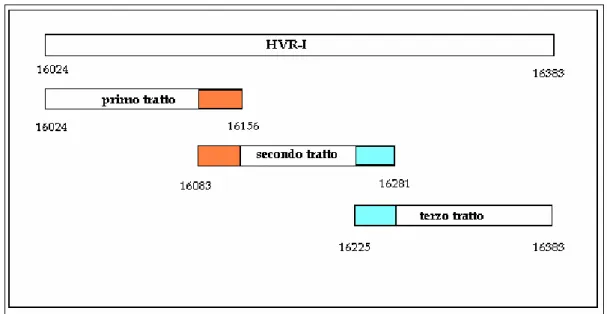
Come si è accennato precedentemente, le regioni che compongono il DNA mitocondriale hanno tassi di evoluzione diversi. Il segmento non codificante D-loop composto da 600 paia di basi, di cui fan parte le regioni ipervariabili HVR I e II, ha un tasso di divergenza molto elevato. Le HVRs hanno un alto numero di siti polimorfici e rilevano circa il 3% della variabilità tra gli individui (Stoneking, 2000). La regione HVR I è il segmento più informativo per confronti tra popolazioni separatesi in tempi evolutivamente recenti visto il grado di omologia con gli scimpanzé ed altri mammiferi, ed è questo il tratto oggetto delle analisi molecolari del presente studio.
Figura 2.1.4a - La molecola del DNA mitocondriale (mtDNA) è circolare ed è presente in molte
copie tutte uguali in ciascun mitocondrio. Essa contiene geni che codificano per RNA ribosomiale (12S e 16 rRNA), RNA transfer (P, T, E, L, S, H, R, G, K, S, D, Y, C, N, A, W, M, Q, I, L e V) ed enzimi (cytb, ND1, ND6, COI, COII, COIII, ATPase 6 e 8). Il DNA mitocondriale si replica autonomamente dal DNA nucleare; la replicazione dei due filamenti H e L inizia in due differenti siti d’origine: OH e OL (la freccia indica il senso della replicazione). La replicazione del filamento pesante inizia con un’ansa di dislocazione della D-loop (regione di controllo).
2.1.5 Recupero e analisi del DNA mitocondriale
Il materiale genetico antico può essere recuperato da numerosi tipi di materia biologica, quali resti mummificati, coproliti, preparati istologici e ossa. Nell’analisi del DNA antico gli ostacoli principali contro cui ci si deve misurare costantemente sono il basso numero di molecole endogene conservate e il loro stato di degradazione dovuto alla diagenesi e ai processi di demolizione spontanea che s’innescano alla morte cellulare, problemi in parte risolti dall’utilizzo della reazione a catena della polimerasi (pcr), una replicazione in vitro capace di copiare selettivamente un frammento di DNA e di aumentarne il numero in modo esponenziale in poco tempo; partendo da una manciata di molecole - in teoria ne basterebbe solo una - in poche ore si producono miliardi di molecole. L’invenzione della polymerase
chain reaction (Mullis et al., 1994) rivoluzionò letteralmente il campo della biologia