RESULTS
1 Characterization of macrophages and DCs
Macrophages and DCs play an essential role in the initiation and maintenance of immune response to intracellular pathogens such as Mtb. Although macrophages are known to be the primary host cells for mycobacteria and serve as the long-term host for the bacilli, Mtb can readily infect DCs as well (Bodnar, Serbina et al. 2001; Jiao et al., 2002).
Thus, to investigate the effects of the initial interactions between several ESX-5 mutant strains of Mtb and the immune cells, two human monocyte derived macrophage populations, and MoDCs were generated.
1.1 Morphologic and phenotypic analysis of the cell populations
Type-1 (M1) and type-2 (M2) were obtained by differentiation of purified human CD14 monocytes in the presence of GM-CSF or M-CSF, respectively. Examination of these macrophages by microscope, after 6 days of culture, has revealed striking differences. The majority of M1 consistently had a classical adherent “fried egg” morphology. M2, on the contrary, primarily appeared as adherent cells with a stretched, spindle-like morphology and some cells remained small and round (Fig.11 A, B).
Fig.11 Human blood monocytes were polarized for 6 days in the presence of
GM-CSF or M-CSF to obtain M1 and M2, respectively, and GM-CSF and IL-4 to obtain DCs. (A) M1 typically appear as adherent cells with a classical fried egg morphology, (B) M2 as adherent and stretched, “spindle-like” cells; (C) DCs as non-adherent, with dendritic and veiled surface projections, cells (Bar = 100m).
To investigate possible differences in the expression of several relevant cell surface markers, M1 and M2 were stained with a series of phagocyte-marker-specific antibodies and appropriate isotype controls (Tab.1). Although absolute expression levels differed among the cells obtained from various donors, the pattern and regulation of expression were consistent throughout.
Both macrophage subsets expressed CD14, the typical monocyte-macrophage marker, on their cell surface; CD14 was expressed consistently higher on M2 than M1. Of all cell surface molecules analyzed, CD163, which is a cysteine-rich scavenger receptor family member, was the only distinctive marker expressed by M2 but not by M1, since it has been reported in literature to be associated with non classical macrophage phenotype (Goerdt and Orfanos, 1999; Verreck et al., 2006). Both polarized M1 and M2 expressed high levels of HLA-DR.
MoDCs were generated by differentiation of purified human CD14 monocytes in the presence of GM-CSF and IL-4 (Gerosa et al., 2008). We provided evidence that GM-CSF (100ng/ml) in combination with IL-4 (5ng/ml) was efficient in the generation of immature, non-adherent, with dendritic and veiled surface projections, MoDCs (Fig.11 C).
DC-SIGN (also known as CD209) expression was high on MoDCs as expected, whereas it was not detected on M1 and M2 macrophages. CD14, which was highly expressed on the surface of both M1 and M2, was virtually not present on MoDCs. The expression of costimulatory molecules, such as CD86, was low on immature MoDCs. Thus, on the basis of cell morphology and surface marker expression, polarized M1 and M2 and immature MoDCs represent three clearly different cell populations. M1 M2 iDCs CD14 820180 1507379 17.64.6 CD83 21±6 38±18 20.55±4.97 CD86 134±54 106±55 65.65±32.11 CD163 5111 520133 NA CD209 8±1.3 17.5±4 99.53±46.07 HLA-DR 514±170 560±125 795.97±117.39
Tab.1 Surface marker on differentiated macrophage populations M1, M2 and MoDCs were stained with specific monoclonal antibodies and analysed by flow cytometry. Indicated are the mean values of MFI ± the mean standard error (SEM) (n=8). NA= not analysed.
1.2 Functional analysisof the cell populations
To characterize the functional activity of the three cell populations we analysed the cytokine profile after 24h of in vitro stimulation with LPS from Escherichia coli (Fig.12).
Activated M1 produced considerable levels of pro-inflammatory cytokines IL-1, IL-6, and TNF-. In contrast, M2 displayed no, or a weak, production of these cytokines while the anti-inflammatory cytokine IL-10 production by activated M2 was significantly higher than activated M1 and MoDCs. Together, these data confirm the pro-inflammatory nature of M1 and anti-inflammatory/regulatory
properties of M2. MoDCs secreted lower amounts of both pro-inflammatory and anti-pro-inflammatory cytokines as compared to macrophage subsets.
Fig.12 Following 24h in vitro stimulation with E. coli LPS, M1, unlike M2, produced high levels of pro-inflammatory cytokines. MoDCs secreted measurable but lower amounts of both pro-inflammatory and anti-inflammatory cytokines as compared to macrophage subsets (n=5).
n g/m L n g/m L Unstimulated E. coli LPS IL-1 M1 M2 MoDCs 0 0.4 0.8 1.2 1.6 2 0 20 40 60 80 100 120 TNF- M1 M2 MoDCs 0 5 10 15 20 IL-10 M1 M2 MoDCs 0 20 40 60 80 100 IL-6 M1 M2 MoDCs
2 Maturation marker expression and cytokine profiles
2.1 MoDC maturation marker expression and cytokine profiles
MoDCs are professional antigen-presenting cells essential for the initiation and regulation of T cell responses (Banchereau and Steinman, 1998; Sallusto and Lanzavecchia, 1999).
In order to investigate the effect of ESX-5 mutant strains on the maturation of MoDCs, immature MoDCs were infected for 24h with Mtbwt or ESX-5 mutants MtbeccD5ko, Mtbppe25-pe19 and
Mtbrv1794ko. Following the infection, surface expression intensity of DC maturation markers CD83, CD80, CD86, HLA-DR and the amount of cytokines secreted into culture supernatants were analyzed. MoDCs stimulated with LPS from E. coli and unstimulated cultures were established as positive and negative controls, respectively.
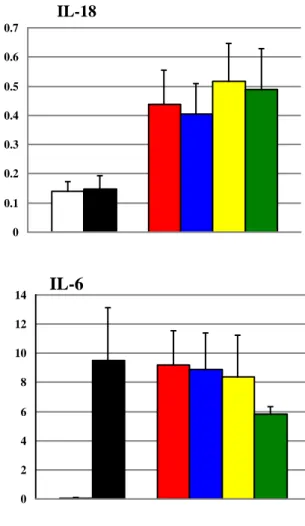
Although expression of CD83 was decreased on MoDCs infected with Mtbwt compared to LPS stimulated cells, we found no difference among MoDCs infected with Mtbwt and the ESX-5 mutants for any of the maturation markers analyzed (Fig.13). Furthermore, the profile of cytokines of MoDCs infected with ESX-5 mutants MtbeccD5ko,
Mtbppe25-pe19, or Mtbrv1794ko was comparable to that of Mtbwt infected cells (Fig.14) indicating that inactivation or deletion of various ESX-5 components do not appear to have an impact on the maturation and cytokine secretion of human MoDCs in vitro.
Fig.13 Maturation marker expression on MoDC infected with ESX-5 mutants
is comparable to that of Mtbwt infected cells. The levels of marker expression were evaluated by MFI analyses (n=4).
0 10 20 30 40 50 60 CD80 0 20 40 60 80 100 120 140 160 180 CD83 0 20 40 60 80 100 120 CD86 0 200 400 600 800 1000 1200 1400 1600 1800 HLA-DR M F I M F I Unstimulated E. coli LPS
Fig. 14 Cytokine profile of MoDC infected with ESX-5 mutants is comparable
to that of Mtbwt infected cells (n=3).
0 1 2 3 4 5 6 7 IL-1 0 0.1 0.2 0.3 0.4 0.5 0.6 0.7 IL-18 0 10 20 30 40 50 60 70 80 TNF- 0 2 4 6 8 10 12 14 IL-6 0 1 2 3 4 5 IL-10 ng /mL ng /mL ng /mL
IL-1
IL-18
TNF-
ng
/m
l
IL-1
IL-18
TNF-
ng
/m
l
2.2 Cytokine production by macrophage populations
Macrophages secrete a number of pro-inflammatory cytokines that are essential in eliciting a protective immune response against intracellular pathogens such as mycobacteria (Orme and Cooper, 1999; Flynn and Chan, 2001; van Crevel et al., 2002; Ottenhoff et al., 2002; Raja, 2004). To determine whether the inactivation/deletion of ESX-5 genes have an impact on these cytokine responses, two macrophage subsets M1 (pro-inflammatory) and M2 (regulatory) were infected with Mtbwt or with the ESX-5 mutants for 24h and the cytokines secreted into culture supernatants were measured.
In a first set of experiments we analysed the cytokine production of the two macrophage subsets infected with Mtbwt or the Mtb ESX-5 mutants MtbeccD5ko and Mtbppe25-pe19, which have been reported to be
attenuated in the mouse model (Bottai et al., 2012). We observed that the secretory profiles of M1 and M2 infected with MtbeccD5ko and
Mtbppe25-pe19 were comparable to that of Mtbwt infected cells (Fig.15).
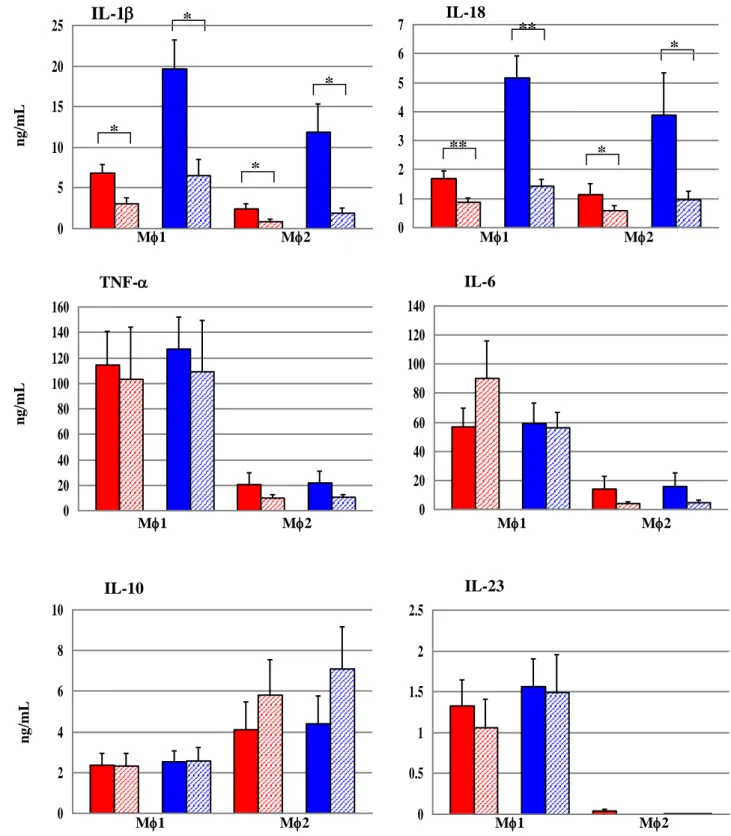
Then we analysed the macrophage response to the mutant strain Mtbrv1794ko, whose orthologue in M. marinum (MMAR_2676) has been demonstrated to modulate cytokine secretion by human macrophages (Abdallah et al., 2008). In contrast to the mutant strains previously analysed, we observed that disruption of Mtb rv1794 gene resulted in significant increase of secretion of pro-inflammatory cytokines IL-1 and IL-18 from both human macrophage populations (Fig.16).
IL-10
IL-1
IL-18
TNF-
ng
/m
l
ng
/m
l
Fig.15 Cytokine profiles of M1 and M2 infected with ESX-5 mutants MtbeccD5ko and Mtbppe25-pe19 is comparable to that of Mtbwt infected cells
(n=6). 0 1 2 3 4 5 6 7 IL-1 0 0.2 0.4 0.6 0.8 1 1.2 1.4 IL-18 0 10 20 30 40 50 60 70 80 90 TNF- 0 5 10 15 20 25 30 35 40 45 50 IL-6 0 0.2 0.4 0.6 0.8 1 1.2 1.4 1.6 IL-10 0 1 2 3 4 5 IL-23 ng /mL ng /mL ng /mL
IL-10
ng
/m
l
IL-1
IL-6
ng
/m
l
TNF-
M
M
1
1
M
M
2
2
IL-18
M
M
1
1
M
M
2
2
M
M
1
1
M
M
2
2
M
M
1
1
M
M
2
2
M
M
1
1
M
M
2
2
Fig.16 IL-1 and IL-18 production was significantly higher in macrophages infected with Mtbrv1794ko mutant than in macrophages infected with Mtbwt. Student’s t-test for paired samples (*****P<0.001, * ***P<0.01, *P<0.05) (n=20). *
M1 M2 M1 M2 M1 M2 M1 M2 M1 M2 M1 M2 0 5 10 15 20 25 IL-1 0 1 2 3 4 5 6 7 IL-18 0 20 40 60 80 100 120 140 160 TNF- 0 10 20 30 40 50 60 70 80 90 IL-6 0 2 4 6 8 10 12 14 16 IL-10 0 0.4 0.8 1.2 1.6 2 IL-23 *** *** ** * ng /mL ng /mL ng /mL
IL-10
ng
/m
l
IL-1
IL-6
ng
/m
l
TNF-
M
M
1
1
M
M
2
2
IL-18
M
M
1
1
M
M
2
2
M
M
1
1
M
M
2
2
M
M
1
1
M
M
2
2
M
M
1
1
M
M
2
2
2.3 Cytokine secretion upon infection of macrophages with live or heat killed bacteria
To determine whether the observed effect on IL-1 and IL-18 production necessitate the presence of metabolically active bacteria, we compared the cytokine profiles of live and heat-killed (hk) bacteria (Fig.17).
When the two macrophage subsets were infected with hk Mtbwt or Mtbrv1794ko, the IL-1 and IL-18 levels were significantly reduced as compared to cells infected with live bacteria, indicating that metabolically active bacteria appear to be essential to induce IL-1 and IL-18 secretion from human macrophage populations. However, hk bacteria elicited amounts of IL-6, TNF-, IL-10 and IL-23 similar to the live bacteria.
IL-10
IL-1
IL-18
TNF-
ng
/m
l
ng
/m
l
Fig. 17 Macrophage subsets infected with hk bacteria showed a significant
reduction of extracellular IL-1 and IL-18 levels as compared with cells infected with live bacteria. Student’s t-test for paired samples (***P<0.01, *
*P<0.05) (n=8). 0 5 10 15 20 25 IL-1 0 1 2 3 4 5 6 7 IL-18 0 20 40 60 80 100 120 140 160 TNF-
0 20 40 60 80 100 120 140 IL-6 0 2 4 6 8 10 IL-10 IL-23 0 0.5 1 1.5 2 2.5 * * * ** ** * * * M1 M2 M1 M2 M1 M2 M1 M2 M1 M2 M1 M2 ng /mL ng /mL ng /mL Mtbwt Mtbrv1794ko Mtbwt hk Mtbrv1794ko hk3 Construction of the complemented strain
For the Mtbrv1794ko a complemented strain (Mtbrv1794ko-C) was constructed, in which the complementing plasmid carrying the rv1794 gene was introduced into Mtbrv1794ko mutant strain.
The Mtbrv1794ko-C complemented strain restored the effects on the level of cytokine production similar to that of Mtbwt. We observed that M1 and M2 infected with the complemented strain (Mtbrv1794ko-C) secreted IL-1 and IL-18 at levels that were comparable to cells infected with Mtbwt (Fig.18).
Fig.18 M1 and M2 infected with Mtbrv1794ko-C secreted IL-1and IL-18 at levels comparable to that of Mtbwt infected cells. Student’s t-test for paired samples (*****P<0.001; *P<0.05) (n=12). * ng /mL ng /mL M1 IL-1 IL-18 * * * * 0 10 20 30 *** *** *** *** 0 1 2 3 4 5 6 7 8 IL-18 0 5 10 15 20 25 30 35 IL-1 0 10 20 30 40 50 M2
IL-10
IL-1
IL-18
TNF-
ng
/m
l
ng
/m
l
4 Growth of Mtbrv1794ko strain in macrophage subsets
To test the hypothesis that the higher levels of IL-1 and IL-18 in culture supernatants of macrophages infected with Mtbrv1794ko as compared to Mtbwt infected cells did not depend on an altered ability of the mutant strain to infect/grow inside macrophages, the number of bacteria (CFU count) were assessed following the infection of M1 and M2 with Mtbrv1794ko or Mtbwt after 24h. We observed no significant differences in CFU number between the macrophage populations infected with Mtbrv1794ko and Mtbwt (Fig.19).Fig.19 No significant differences in CFU numbers were observed between
Mtbwt and Mtbrv1794ko infected macrophage populations following 24h infection.
IL-10
IL-1
IL-18
TNF-
ng
/m
l
ng
/m
l
0.00E+00 2.00E+06 4.00E+06 6.00E+06 8.00E+06 1.00E+07M
1
M
2
Mtbrv1794ko Mtbwt Mtbwt Mtbrv1794ko CF U/mL5 Induction of cell death and apoptosis
As both M1 and M2 macrophage populations similarly produced higher levels of IL1 and IL-18 upon infection with Mtbrv1794ko, we focused on M1 population for further experiments.
It has been proposed that Mtb inhibits apoptosis and triggers cell death by necrosis of host macrophages to evade innate immunity (Behar et al., 2010; Behar et al., 2011).
To investigate the ability of the mutant strain to induce cell death in human macrophages, M1 were infected with Mtbwt, Mtbrv1794ko or Mtbrv1794ko-C and cell death was assessed by flow cytometry following staining with a membrane-impermeant fluorescence dye, which infiltrates selectively into dead cells because of their damaged membranes. The dye covalently binds to nucleic acids of dead cells upon exposure to visible light. In this way dead cells can be detected by FACS analysis. At 24h following infection we observed that Mtbwt was able to induce cell death in 92 % (meanSEM) of cells. Comparable results were obtained in macrophages infected with Mtbrv1794ko or Mtbrv1794ko-C (Fig.20).
Moreover, the capacity of the mutant strain to induce apoptosis in M1 was investigated following 24h of infection. We observed that the proportion of apoptotic cells were comparable for the three strains tested (Fig.20).
We can conclude that the differences in IL-1 production between Mtbwt and Mtbrv1794ko, are not due to a different ability of the mutant strain to infect M1 and induce cell death, since the percentage of dead or apoptotic cells was not significantly different between both strains.
Fig.20 Percentage of dead and apoptotic cells following 24h infection of M1 with Mtbwt, Mtbrv1794ko or Mtbrv1794ko-C.
0
2
4
6
8
10
12
14
P
ercent
a
g
e
o
f p
o
si
ti
v
e ce
ll
s
Mtbwt Mtbrv1794ko Mtbrv1794ko-C6 Analysis of IL-1
pathway
The processing and secretion of active IL-1 by innate immune cells is dependent, first, on induction of pro–IL-1 through PRR-signaling and then on activation of a multiprotein complex called “inflammasome” which, in turn, triggers the caspase-1, an enzyme required for cleavage of pro-IL-1 into their mature active forms.
Therefore, we first investigated the effect of Mtbwt, Mtbrv1794ko and Mtbrv1794ko-C on the synthesis of pro-IL-1, as well as inflammasome components (i.e. NLRP3, ASC and caspase-1). Next, we examined the capacity of the mutant strain, compared to the Mtbwt, to activate the caspase-1.
6.1 Quantitative measurement of pro-IL-1 mRNA levels by Real Time PCR
We have shown that IL-1 levels were significantly higher in culture supernatants of M1 infected with Mtbrv1794ko as compared to the cells infected with Mtbwt or Mtbrv1794ko-C.
To investigate whether this difference was due to a major stimulation of the inactive cytokine precursors, relative mRNA levels of pro-IL-1 in M1 infected with Mtbwt, Mtbrv1794ko or Mtbrv1794ko-C were analysed.
All the bacterial strains strongly induced pro-IL-1 mRNA expression in M1 at 4h and 24h after infection. However, there were no significant differences in pro-IL-1 expression from Mtbrv1794ko infected M1 as compared to Mtbwt or Mtbrv1794ko-C infected ones (Wilcoxon matched pairs signed-rank test)(Fig.21).
Fig.21 Pro-IL-1 expression in Mtbrv1794ko infected M1 was comparable to that of Mtbwt infected cells. Quantitative measurement of mRNA levels was performed by real time PCR and expressed as fold increase compared to unstimulated cells. IL-1 Fold i n cr ea se r el a ti ve t o e xpres si on in un st im u lat ed ce ll s ( L og ) 4h 1.00E+00 1.00E+01 1.00E+02 1.00E+03 1.00E+04 1.00E+05 1.00E+06 24h 1.00E+00 1.00E+01 1.00E+02 1.00E+03 1.00E+04 1.00E+05 1.00E+06
Mtbwt Mtbrv1794ko Mtbrv1794ko-C Mtbwt Mtbrv1794ko Mtbrv1794ko-C
6.2 Analysis of pro- and active-IL-1 protein levels by immunoblotting
Based on the level of cytokine transcript, it appeared that pro-IL-1 was similarly induced after infection with all the bacterial strains.
Immunoblotting of lysates of M1 infected 24h with Mtbwt, Mtbrv1794ko or Mtbrv1794ko-C as well as non-infected cells, confirmed the data obtained by the quantitative Real Time-PCR. The inactive precursor of IL-1 (31 kDa) was detected equally in M1 infected with Mtbwt, Mtbrv1794ko or Mtbrv1794ko-C. As most of the IL-1 activation occurs during its exocytosis, the active fragment is not readily visible on Western blots of cell lysates (Master et al., 2008). On the contrary, the active form of IL-1 (17 kDa) was detected in higher amounts in the culture supernatants of M1 infected with Mtbrv1794ko as compared to cells infected with Mtbwt or Mtbrv1794ko-C. -actin (41 kDa) was used to normalize the amount of the total protein in each experimental condition (Fig.22).
Fig.22 Mtbrv1794ko induces equal amounts of pro-IL-1 in M1 cell lysates but an increased release of mature IL-1 into culture supernatants as compared to Mtbwt or Mtbrv1794ko-C.
Cell lysates
Culture supernatants
-actin Pro-IL-1 IL-1 41 kDa 31 kDa 17 kDa IL-1 -actin ratio % 0 73 67 64
RPMI Mtbwt Mtbrv1794ko Mtbrv1794ko-C RPMI Mtbwt Mtbrv1794ko Mtbrv1794ko-C
6.3 Caspase-1 inhibition
Since caspase-1 is generally required for the processing of IL-1 and IL-18 in human monocyte derived macrophages infected with Mtb (Ciaramella et al., 2002), we used a caspase-1 specific inhibitor to validate this observation in our system. We observed that the amount of IL-1 and IL-18 detectable in culture supernatants of infected macrophages was drastically reduced when cells were pre-treated with caspase-1 inhibitor Ac-YVAD-cmk (Fig.23). On the contrary, the production of IL-10 was unaffected by the inhibition of caspase-1. These results suggest that IL1 and IL-18 production of infected macrophages is dependent on caspase-1. Therefore, we investigated the effect of the Mtbrv1794ko strain on pro-caspase-1 synthesis and caspase-1 activation.
Fig.23 Inhibition of caspase-1 strongly reduces IL-1 and IL-18 production. M1 were infected with Mtbwt, Mtbrv1794ko or Mtbrv1794ko-C in the presence of 50M of caspase-1 inhibitor YVAD. IL-1 and IL-18 levels in culture supernatants were determined 24h after infection (Data from a representative experiment is depicted).
0 2 4 6 8 10 IL-1 Control + YVAD IL-18 0 2 4 6 8 10 Control + YVAD IL-10 0 1 2 3 4 5 Control + YVAD ng /mL ng /mL E. coli LPS Mtbwt
6.4 Quantitative measurement of mRNA levels of inflammasome components by Real Time PCR
Distinct inflammasome complexes, consisting of the adaptor ASC and caspase-1 coupled to a cytosolic pattern-recognition receptor, are involved in the processing of pro-IL-1. Although the inflammasome assembly and activation is a hallmark of the innate immune response against intracellular pathogens (Mariathasan and Monack, 2007; Stutz et al., 2009), the role of these scaffolding proteins during mycobacterial infection has not been studied in detail. Nevertheless, recent works have identified a NLRP3 and ASC-containing complex as the major inflammasome activated in macrophages by Mtb infection (Koo et al., 2008; Carlsson et al., 2010; Mishra et al., 2010; Wong et al., 2011; Dorhoi et al., 2012).
To define the role of the NLRP3 inflammasome components, we analysed the expression of NLRP3, ASC and pro-caspase-1 at mRNA level in M1 infected with Mtbwt, Mtbrv1794ko, or Mtbrv1794ko-C (Fig.24). We observed that the NLRP3 or ASC mRNA expression was equally induced in all the experimental conditions. Although the pro-caspase-1 mRNA expression was strongly induced following 4h and 24h of infection, no significant differences were detected between macrophages infected with Mtbrv1794ko and Mtbwt or Mtbrv1794ko-C infected cells (Wilcoxon matched pairs signed-rank test).
Fig.24 NLRP3, ASC and caspase-1 expression in Mtbrv1794ko infected M1 was comparable to that of Mtbwt infected cells. Quantitative measurement of mRNA levels was performed by real time PCR and expressed as fold increase compared to unstimulated cells.
NLRP3 4h 1.00E-03 1.00E-02 1.00E-01 1.00E+00 1.00E+01 1.00E+02 1.00E+03 24h 1.00E-01 1.00E+00 1.00E+01 1.00E+02 1.00E+03 ASC 4h 1.00E-03 1.00E-02 1.00E-01 1.00E+00 1.00E+01 1.00E+02 1.00E+03 24h 1.00E-02 1.00E-01 1.00E+00 1.00E+01 1.00E+02 1.00E+03 Caspase-1 4h 1.00E-02 1.00E-01 1.00E+00 1.00E+01 1.00E+02 1.00E+03 1.00E+04 1.00E+05 1.00E+06 24h 1.00E-02 1.00E-01 1.00E+00 1.00E+01 1.00E+02 1.00E+03 1.00E+04 1.00E+05
Mtbwt Mtbrv1794ko Mtbrv1794ko-C Mtbwt Mtbrv1794ko Mtbrv1794ko-C
Mtbwt Mtbrv1794ko Mtbrv1794ko-C Mtbwt Mtbrv1794ko Mtbrv1794ko-C
Mtbwt Mtbrv1794ko Mtbrv1794ko-C Mtbwt Mtbrv1794ko Mtbrv1794ko-C
Fold i n cr ea se r el a ti ve t o e xpres si on in un st im u lat ed ce ll s ( L og ) Fold i n cr ea se r el a ti ve t o e xpres si on in un st im u lat ed ce ll s ( L og ) Fold i n cr ea se r el a ti ve t o e xpres si on in un st im u lat ed ce ll s ( L og )
6.5 Caspase-1 activity
Cleavage of the pro-IL-1 to its active form is performed by the caspase-1 (Martinon and Tschopp, 2004; Dinarello, 2009). In resting cells, caspase-1 is present in an inactive state (pro-caspase-1) and, once recruited to an inflammasome complex, it is activated and, in turn, converts pro-IL-1 into active IL-1.
Data obtained by Real Time-PCR suggest that a possible biased processing of pro-IL-1 might be due to a difference in activation rather than production of caspase-1. Thus, we investigated whether a difference in the activation status of this enzyme might be responsible for the higher IL-1 levels observed in culture supernatants of macrophages infected with Mtbrv1794ko, as compared to Mtbwt or Mtbrv1794ko-C infected cells. To this purpose we directly monitored caspase-1 activation in M1 by using a FLuorochrome Inhibitor of active CAspase assay (FLICA). The FITC-labelled cell-permeable FLICA reagent FAM-YVAD-FMK enters the cells and forms a covalent link with activated caspase-1, therefore it can be used to measure the amount of active caspase-1 inside the cell by flow cytometry.
FLICA analyses demonstrated that caspase-1 activity was increased in M1 infected with Mtbrv1794ko as compared to cells infected with Mtbwt (Fig.25), suggesting that increased levels of IL-1 measured in supernatants of Mtbrv1794ko infected M1 might indeed be due to increased caspase-1 activity in these cells.
Fig.25 Higher caspase-1 activity was observed in M1 infected with Mtbrv1794ko as compared to Mtbwt.
7 The inflammasome activity
Caspase-1 conversion is achieved upon assembly and activation of inflammasomes, cytosolic multiprotein complexes conveting pro-caspase-1 into active pro-caspase-1. While inflammasome activation and subsequent IL-1 and IL-18 synthesis are strongly upregulated in Mtb infected macrophages (Giacomini et al., 2001; Montero et al., 2004), the mechanism by which Mtb activate the inflammasome for processing and release of these cytokines is not completely understood (Koo et al., 2008; Master et al., 2008; Kurenuma et al., 2009; Mishra et al., 2010). We investigated two of the major inflammasome triggering mechanisms, i.e. the K+ efflux and the release of the lysosomial protease cathepsin B, reported to be involved in Mtb infection (Kurenuma et al., 2009; Abdallah et al., 2011), to evaluate their role in the increased IL-1 production by Mtbrv1794ko infected cells.
7.1 Cytokine production via induction of the K+ efflux
K+ efflux has long been considered to be one of the main mechanisms for activation of NLRP3 inflammasome (Petrilli et al., 2007); furthermore, it has been shown to be important for inflammasome activation by Mtb (Kurenuma et al., 2009). To investigate whether this inflammasome triggering stimulus plays a role in the increased IL-1 production by Mtbrv1794ko infected cells, a high concentration of KCl (40mM) was added to the infection medium to inhibit potential K+ efflux.
The results obtained demonstrated that the amount of IL-1 and IL-18 was reduced in all the experimental conditions where KCl was added to the infection medium. Moreover, no differences were observed between M1 infected with Mtbrv1794ko and Mtbwt or Mtbrv1794ko-C in presence of KCl (Fig.26).
Fig.26 Increased intracellular [K+] decreases IL-1 and IL-18 secretion. M1 were infected with Mtbwt, Mtb1794ko or Mtbrv1794ko-C without ( ) or with the addition of 40mM KCl ( ) IL-1 and IL-18 levels in culture supernatants were determined following 24h of infection. (Data from a representative experiment is depicted).
RPMI Mtbwt Mtbrv1794ko Mtbrv1794ko-C
IL-1 0 5 10 15 20 25 IL-18 0 2 4 6 8 10
RPMI Mtbwt Mtbrv1794ko Mtbrv1794ko-C
ng
7.2 Cytokine production via release of cathepsin B
It has been shown that lysosomal destabilization and leakage of cathepsin B into the cytoplasm results in the activation of the NLRP3 inflammasome by an unidentified mechanism (Halle et al., 2008). Furthermore, inflammasome activation has recently been reported to be triggered by cathepsin B in the context of mycobacterial infection (Abdallah et al., 2011). To investigate the role of cathepsin B in inflammasome activation during infection with Mtbwt, Mtbrv1794ko or Mtbrv1794ko-C, 10M of the cathepsin B inhibitor Ca-074-Me, cathepsin D inhibitor Pepstatin A or cathepsin L inhibitor Z-Phe-Tyr(tBu)-dmk were added to the infection medium.
M1 infected with Mtbrv1794ko, Mtbwt, or Mtbrv1794ko-C in the presence of cathepsin B inhibitor Ca-074-Me failed to secrete IL-1 and IL-18. In contrast, the presence of cathepsin D or L inhibitors in the culture of infected macrophages did not cause a significant alteration in the IL-1 and IL-18 secretion (Fig.27). These results suggest a specific requirement for active cathepsin B for the production of pro-inflammatory cytokines IL-1 and IL-18 by infected macrophages.
Fig. 27 Inhibition of Cathepsin B decreases IL-1 and IL-18 secretion. M1 were infected with Mtbwt, Mtb1794ko or Mtbrv1794ko-C without or with the addition of 10M Cathepsin B inhibitor CA-074, 10M Cathepsin D inhibitor Pepstatin A, 10M Cathepsin L inhibitor Z-Phe-Tyr(tBu)-dmk into the cultures. IL-1 and IL-18 levels in culture supernatants were determined following 24h of infection (Data from a representative experiment is depicted).
0 2 4 6 8 10 IL-1
Control Ca-074 Pepstatin A Z-Phe- (tBu)-dmk IL-18 0 2 4 6 8 10
Control Ca-074 Pepstatin A Z-Phe- (tBu)-dmk
E. coli LPS Mtbwt
unstimulated Mtbrv1794ko Mtbrv1794ko-C
ng