responsabili dell’allungamento del tronco. Naturalmente le due spiegazioni non si escludono a vicenda.
1.3 I geni Xotx e la Gastrulazione di Xenopus laevis
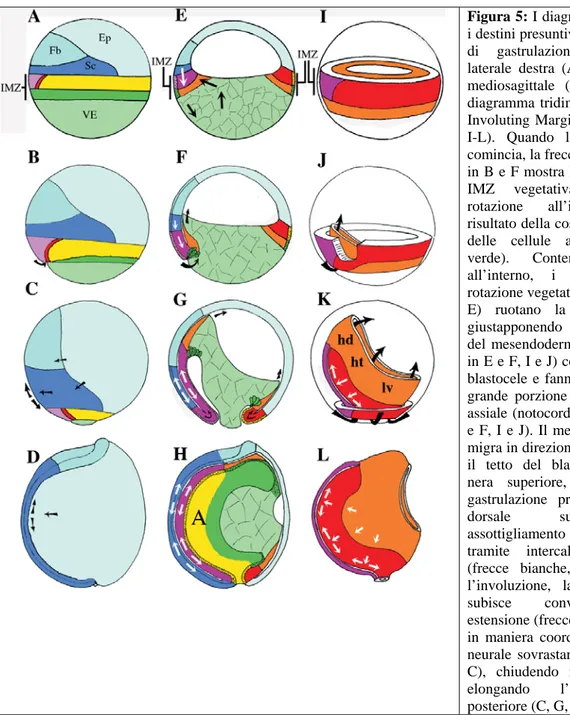
La migrazione cellulare che avviene durante la gastrulazione è senza dubbio uno degli eventi fondamentali dello sviluppo embrionale. La regione dell’embrione in cui inizia la gastrulazione si trova in posizione dorsale e corrisponde al territorio del labbro dorsale del blastoporo e alla notocorda presuntiva. Il mesoderma si forma all’interno della Involuting Marginal Zone (IMZ; Fig. 5A,E,I).
La IMZ è un anello di tessuto formato da uno strato superficiale di cellule epiteliali e una più spessa e profonda regione sottostante di cellule mesenchimali. La parte superficiale della IMZ è formata da notocorda presuntiva (in rosa), da una piccola porzione di mesoderma somitico presuntivo (in rosso), endoderma presuntivo (in giallo) e l’endoderma delle cellule a bottiglia presuntivo (in verde scuro; Fig. 5A-C, E-H). La regione multistratificata delle cellule mesenchimali sottostanti è formata da notocorda presuntiva (in magenta), mesoderma somitico presuntivo (in rosso), mesoderma della testa, cuore e mesoderma latero-ventrale presuntivi (in arancione; Fig. 5E-H, I-L). La IMZ si trova alla periferia dell’endoderma vegetativo (verde chiaro, Fig. 5E-H).
Durante la gastrulazione, movimenti morfogenetici diversi rivoluzionano drasticamente la struttura dell’embrione. Il primo di questi movimenti è l’involuzione della IMZ, le cui cellule ruotano intorno a una fessura detta labbro dorsale del blastoporo e successivamente si muovono lungo il tetto del blastocele (frecce nere, Fig. 5F-G, J-K). L’involuzione comincia dorsalmente
dopo la formazione tramite invaginazione di una depressione locale, il labbro dorsale del blastoporo. Essa deriva dal ripiegamento all’interno di uno strato epiteliale, mentre le cellule a bottiglia presuntive (cuboidali) subiscono una costrizione apicale e diventano a forma di cuneo. (Fig. 5A-B, E-F, 6A-C).
Figura 5: I diagrammi mostrano i destini presuntivi e i movimenti di gastrulazione in visione laterale destra (A-D), nel piano mediosagittale (E-H), e in un diagramma tridimensionale della Involuting Marginal Zone (IMZ; I-L). Quando la gastrulazione comincia, la freccia nera in basso in B e F mostra la trazione della IMZ vegetativamente e la rotazione all’interno come risultato della costrizione apicale delle cellule a bottiglia (in verde). Contemporaneamente, all’interno, i movimenti di rotazione vegetativa (frecce nere, E) ruotano la IMZ dorsale, giustapponendo il bordo guida del mesendoderma (in arancione in E e F, I e J) contro il tetto del blastocele e fanno involvere una grande porzione del mesoderma assiale (notocorda, magenta in E e F, I e J). Il mesendoderma poi migra in direzione animale lungo il tetto del blastocele (freccia nera superiore, F, J). Nella gastrulazione precoce, la IMZ dorsale subisce un assottigliamento ed estensione tramite intercalazione radiale (frecce bianche, E, F). Dopo l’involuzione, la IMZ dorsale subisce convergenza ed estensione (frecce bianche, G, K) in maniera coordinata al tessuto neurale sovrastante (frecce nere,
Il labbro dorsale del blastoporo, poco profondo, definisce il sito di inizio della gastrulazione, ma contribuisce poco alla profondità finale dell’archenteron. Cambiamenti molto più significativi sono compiuti nello stesso tempo dai movimenti recentemente descritti di rotazione vegetativa (Winklbauer e Schürfeld, 1999). Durante tutti gli stadi tardivi della blastula e la gastrulazione, gli apici delle cellule dell’endoderma vegetativo si contraggono gradualmente (Keller, 1975), comprimendo le loro estremità basali nella parte animale e estendendo la IMZ nella parte vegetativa. All’inizio della gastrulazione, le cellule centrali dell’endoderma vegetativo si stanno già muovendo in direzione del polo animale, verso il pavimento del blastocele, mentre quelle sul pavimento si muovono nelle zone periferiche (più dorsali) contro il tetto del blastocele dorsale. Questo complesso insieme di movimenti ruota l’intera IMZ e inizia l’involuzione della parte vegetativa di essa intorno al labbro del blastoporo (frecce nere Fig. 5E-F;I-J; Fig. 6 A,B).
Sia la formazione delle cellule a bottiglia che la rotazione vegetativa cominciano dorsalmente e continuano lateralmente e ventralmente. Gran parte dei movimenti di involuzione precoce che si pensavano dovuti alla formazione delle cellule a bottiglia (Hardin e Keller, 1988) avvengono invece a causa di questi più sostanziali movimenti di rotazione vegetativa. La rotazione vegetativa porta il mesendoderma (il bordo superiore del foglietto mesodermico e l’endoderma associato) in giustapposizione al sovrastante tetto del blastocele (Fig. 5F). Una volta all’interno, migra direzionalmente verso il polo animale, usando il soprastante tetto del blastocele come superficie di scorrimento. (Holtfreter, 1943a,b; 1944; Nakatsuji, 1975; Nakatsuji e Johnson, 1982; Winklbauer, 1990; Davidson et al., 2002).
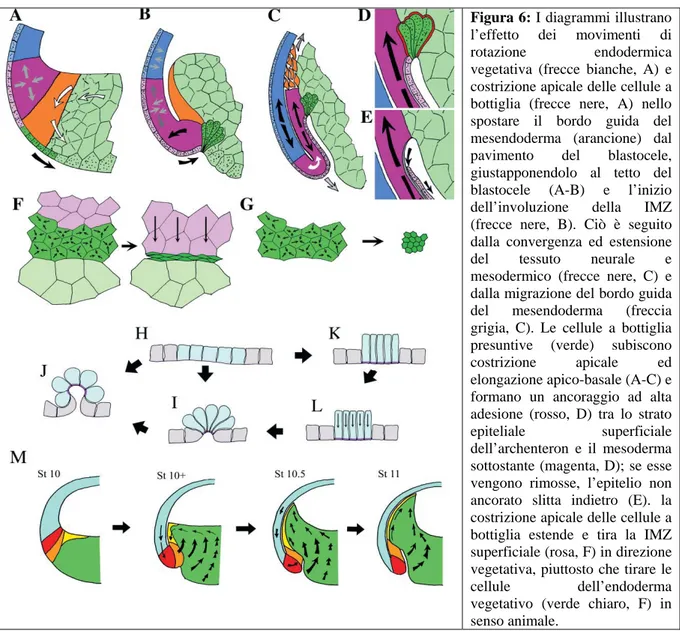
Figura 6: I diagrammi illustrano l’effetto dei movimenti di rotazione endodermica vegetativa (frecce bianche, A) e
costrizione apicale delle cellule a bottiglia (frecce nere, A) nello spostare il bordo guida del mesendoderma (arancione) dal pavimento del blastocele, giustapponendolo al tetto del blastocele (A-B) e l’inizio dell’involuzione della IMZ (frecce nere, B). Ciò è seguito dalla convergenza ed estensione del tessuto neurale e mesodermico (frecce nere, C) e dalla migrazione del bordo guida del mesendoderma (freccia grigia, C). Le cellule a bottiglia presuntive (verde) subiscono costrizione apicale ed elongazione apico-basale (A-C) e formano un ancoraggio ad alta adesione (rosso, D) tra lo strato
epiteliale superficiale dell’archenteron e il mesoderma
sottostante (magenta, D); se esse vengono rimosse, l’epitelio non ancorato slitta indietro (E). la costrizione apicale delle cellule a bottiglia estende e tira la IMZ superficiale (rosa, F) in direzione vegetativa, piuttosto che tirare le
cellule dell’endoderma vegetativo (verde chiaro, F) in
senso animale.
Nell’embrione la costrizione apicale è altamente anisotropa con piccole contrazioni nella direzione della circonferenza (direzione mediolaterale) e grandi contrazioni nella direzione animale-vegetativa, che risultano in una forma allungata delle cellule (F). In espianti la costrizione apicale avviene in tutte le direzioni (G). Cellule a bottiglia espiantate rimangono più rotonde (H-J). La costrizione apicale senza resistenza all’elongazione apico-basale produce colonnarizzazione (HK). Successivi accorciamenti apico-basali producono invaginazione (K-L-I). (Da Keller et al., 2003).
La sola involuzione della IMZ non è tuttavia sufficiente per la chiusura del blastoporo né è importante per l’elongazione dell’asse corporeo. Questi movimenti sono in larga parte compiuti dalla convergenza ed estensione. Per
notocorda e dei somiti e la soprastante piastra neurale posteriore si restringono (convergono) lungo la direzione mediolaterale (circumblastoporale), e si allungano (si estendono) in direzione antero-posteriore (frecce bianche, Fig. 5G-H, K-L). Questi movimenti accoppiati, spesso chiamati estensione
convergente, allungano l’asse corporeo e contribuiscono all’involuzione delle
cellule e alla chiusura del blastoporo (Keller, 1986; Keller et al., 2000). Durante tutti gli stadi di blastula e gastrula, la regione animale dell’embrione si espande per epibolia, estendendosi in direzione vegetativa nella regione lasciata vuota dalla IMZ quando quest’ultima si è internalizzata. L’epibolia si ha per assottigliamento e allargamento della porzione animale (Fig. 5A-C).
1.3.a Convergenza ed Estensione
Durante la gastrulazione e la neurulazione, il mesoderma presuntivo della notocorda e dei somiti e la sovrastante piastra neurale posteriore, convergono (si restringono) nella loro direzione mediolaterale (circumblastoporale) e si estendono (si allungano) lungo la loro dimensione antero-posteriore (Keller, 1975; 1976). Questi movimenti allungano l’asse antero-posteriore, guidano l’involuzione, portano a una chiusura asimmetrica il blastoporo e risultano nella formazione di un archenteron allungato (Keller, 1986 e Keller et al., 2000; frecce bianche, pannelli a sinistra, Fig. 7A, B).