CHAPTER 7
PALAEOBIOGEOGRAPHY
7.1 INTRODUCTION
The Neogene Euro-North African sirenians were fully aquatic herbivorous mammals strictly linked to coastal environments. They inhabited the Euro-North African coasts (Paratethys, Mediterranean, southern North Sea and northeastern Atlantic coasts) during a crucial period characterized by a long-term trend of climatic cooling (e.g. Zachos et al., 2001) and of decreasing marine influence with correlative reduction in size of the marine depositional domains (e.g. Rögl, 1998; Meulenkamp et al., 2000; Meulenkamp & Sissingh, 2003; Popov et al., 2004), which had repercussions on sirenian evolution that can be observed in the fossil record.
All the Euro-North African coasts were influenced during the Neogene by the effects of the increasing African-Eurasian convergence: the North African shoreline, constituting the southwestern Peri-Tethys margin, underwent a long-term northward movement; while the European Mediterranean coasts, constituting the northwestern Peri-Tethys margin, underwent a more complex development due to the final stages of Alpine convergence and continent-continent collision with back-arc post-orogenic extensions and island-arc formation (e.g. Meulenkamp & Sissingh, 2003). The Paratethys realm underwent progressive isolation with salinity variations (Popov et al., 2006; Meulenkamp & Sissingh, 2003, Jolivet et al., 2006); the northeast Atlantic shorelines underwent a long-term northwestward movement; and the southern coasts of the North Sea Basin moved northward with a consequent reduction in marine influence, and remained separated from the European Atlantic coasts until the Pliocene (Vliet-Lanoë et al., 2002; Meulenkamp & Sissingh, 2003).
The following schematic palaeogeographic maps (Figg. 1-4), covering four selected time slices, including the Early Miocene (early Burdigalian: 20.5-19 Ma), the Middle Miocene (early Langhian: 16.4-15.5 Ma), the Late Miocene (late Tortonian: 8-7 Ma), and the Early-Middle Pliocene (Zanclean-Piacenzian 5.4-2.6 Ma), illustrate the palaeogeographic evolution of the Euro-North African coasts during the Neogene, with indications of the known sirenian record (listed in detail in Appendix 2).
7.2 EARLY MIOCENE
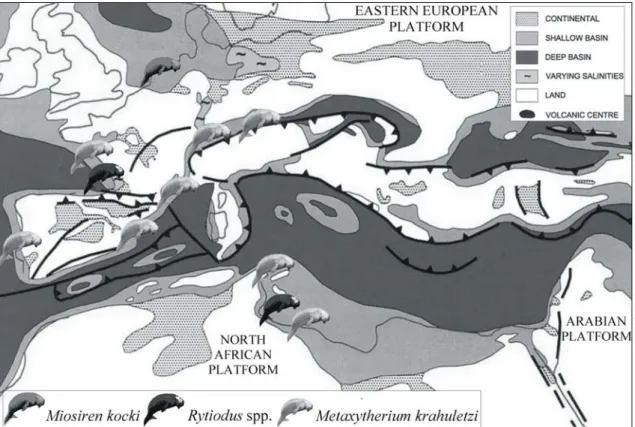
(Fig. 1)Fig. 1: Schematic palaeogeographic map of the Euro-North African coasts during the Early Miocene (modified from Meulenkamp & Sissingh, 2003); with indications of the main known sirenian records.
The Early Miocene was a period of global warming culminating in a climatic optimum from 17 to 15 Ma (Zachos et al., 2001). During this time taxa representing three different sirenian lineages lived along the Euro-North African coasts: Miosiren kocki (Trichechidae, Miosireninae); Metaxytherium krahuletzi (Dugongidae, Halitheriinae); and the genus Rytiodus (Dugongidae, Dugonginae).
Sirenian remains referred, or which could be referred, to Metaxytherium krahuletzi have been found in several lower Miocene shallow marine deposits of the Alpine Molasse - Carpathian Foreland Basins (Switzerland, Austria, Slovakia and maybe Germany), of the Rhône Graben region (France), and of the Alboran Sea (Spain).
On the northern Peri-Tethys platform, by the Oligocene-Miocene boundary, the anticlockwise rotation of the Corsica-Sardinia block contributed to the collapse of the Gulf of Lions and the formation of the Valencia Trough. This successively determined the oceanization of the Algero-Provençal basin, constituting in this way a western basin connected with the Alpine Molasse Basin in the east and with the Alboran Sea in the
west (Roca, 2001). During the Aquitanian the Western Alpine Molasse Basin was closed in response to regional uplift. Contemporaneously, the rifting of the Rhône Graben ended and, in this way, the saline corridor between the North Sea and the Western Mediterranean was closed. However, a widespread marine transgression occurred during the Burdigalian in the North Alpine Molasse Basin and in the southernmost Rhône Graben, coeval with the Algero-Provençal oceanization (Sissingh, 2001), and this allowed sirenians to spread along the coasts of these internal basins. In the Eastern Paratethys, on the contrary, deep basins with anoxic conditions persisted throughout most of the Early Miocene (Meulenkamp & Sissingh, 2003), and no sirenian remains have been found in this area.
The Northeast Atlantic coasts underwent a regressive phase with a progressive shift from marine to fluvio-lacustrine sedimentation: the Paris Basin and the Aquitaine Basin closed respectively during the Oligocene-Miocene transition and in the Early Miocene (Meulenkamp & Sissingh, 2003); and the southern border of the North Sea Basin moved northward, affected by tectonic uplift and by fluctuating sea level (Vandenberghe et al., 1998). This area, separated from the European Atlantic coasts, was inhabited by large trichechids belonging to the genus Miosiren, whose remains were found in the Aquitanian deposits of the Edegem Sands Member of the Berchem Formation in Belgium and in the reworked sediment of the Red Crag Formation in England. Meanwhile, in the Aquitaine Basin, dugongines and halitheriines seem to have lived sympatrically, represented by the large-tusked dugongine Rytiodus capgrandi and by the generalist small-tusked halitheriine Metaxytherium krahuletzi.
Some other indeterminate sirenian remains come from Lower Miocene deposits of Portugal, testifying to a palaeobiogeographic continuity of sirenian records along the northeastern Atlantic coasts and the Mediterranean ones.
On the southern Peri-Tethys platform, the most relevant event during the Early Miocene was the separation between the African and Arabian Plates, whose initial rifting started around 30-27 Ma and ended around 16 Ma (Meulenkamp & Sissingh, 2003). Sirenian remains have been found in the Lower Miocene Gebel Zelten Formation (Libya). Some of these were referred to Metaxytherium sp. indet. by Heal (1973) and by Domning and Pervesler (2001), and others to Rytiodus sp. by Heal (1973); therefore also in this area dugongines and halitheriines seem to have lived sympatrically. Some fragmentary remains referable to Metaxytherium come also from Malta, Gozo and Sicily.
7.3 MIDDLE MIOCENE
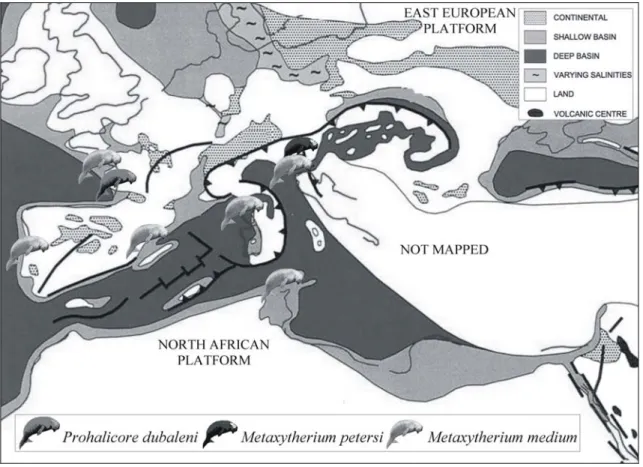
(Fig. 2)Fig. 2: Schematic palaeogeographic map of the Euro-North African coasts during the Middle Miocene (modified from Meulenkamp & Sissingh, 2003); with indications of the main known sirenian records.
The Middle Miocene was characterized by stepwise cooling. After this time, the Eastern Antarctic Ice Sheet was permanently established, causing a eustatic sea level drop of about 100 m (Zachos et al., 2001); the ocean circulation patterns became similar to the modern ones; and the world became globally and permanently cooler (Flower & Kennett, 1994).
In the North Sea, after the deposition of the Berchem Formation and before the deposition of the Diest Formation (Middle Miocene), a SW-NE oriented deep trough was formed on the continental shelf of the southern coast and a regressive phase occurred (Spiegler, 2001). Sirenians seem to have disappeared from the Belgian deposits, but they were found in the northern deposits of The Netherlands.
Around the Early-Middle Miocene transition, the principal rifting of the European Cenozoic Rift System, begun during the Middle-Late Eocene, had come to an end; the Corsica-Sardinia block ceased its rotation; the Carpathian Foreland Basin stopped
subsiding; the Vienna Basin and the Pannonian back-arc basin system opened; and the Greater Caucasian archipelago emerged. On the south, meanwhile, the Arabian Platform, which had been emergent since about the Eocene-Oligocene transition, underwent a marine transgression; parts of the African platform domain collapsed (Meulenkamp & Sissingh, 2003); the Red Sea Basin prograded northward; while the Suez Rift increased its isolation from the Mediterranean with evaporite accumulation in the Gulf of Suez-Red Sea domain around 14 Ma (Orszag-Sperber et al., 1998).
Only the generalist genus Metaxytherium survived along the coasts of the Mediterranean Sea and northeast Atlantic (Italy, southern Spain, Portugal, France, Netherlands; Tunisia), and also in the Western and Central Paratethys region (Austria; Switzerland and maybe Hungary). All the Atlantic and Mediterranean specimens are referred to M. medium, while some of the Paratethys specimens are referred to M.
medium and others to M. petersi (Domning, pers. comm.).
The only exception is represented by Prohalicore dubaleni, represented only by a partial mandible found in the Serravallian deposits of Audon, near Tartas (Landes, SW France) (Flot, 1887). This formation represents the last important transgression of the Atlantic Ocean in Aquitaine (Gardère, 2005)
7.4 LATE MIOCENE (
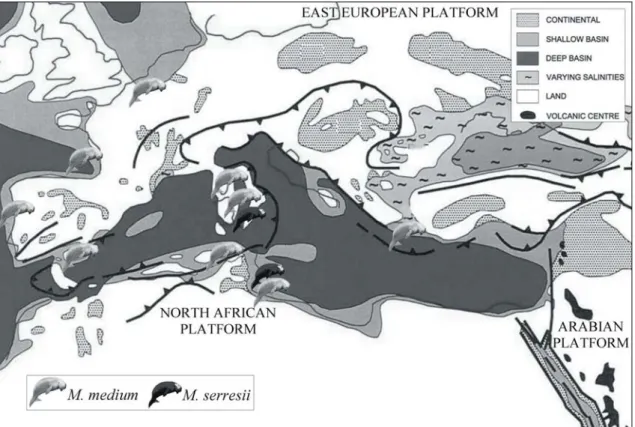
Fig. 3)Fig. 3: Schematic palaeogeographic map of the Euro-North African coasts during the Late Miocene (modified from Meulenkamp & Sissingh, 2003); with indications of the main known sirenian records.
In the course of the late Middle and Late Miocene, the cooling trend continued (Zachos et al., 2001), and the overall uplift affected the western and central domains of the Northern Peri-Tethys Platform (Meulenkamp & Sissingh, 2003). These processes seem to have had repercussions for sirenian palaeobiogeography; in fact, during this period sirenians seem to have gone extinct in the Paratethys and become reduced along the northeast Atlantic coasts; while in the Mediterranean Sea, they have been found in Spain, Italy, Greece, Tunisia, Libya, Malta and Gozo. All of these sirenian specimens are referred, or could be referred, to the generalist halitheriine Metaxytherium medium. The uplift was particularly effective around 11 Ma and around 8 Ma and caused the final break-up of the sedimentation realm across Europe (Meulenkamp & Sissingh, 2003); the evaporite accumulation in the Gulf of Suez-Red Sea domain terminated (Orszag-Sperber et al., 1998); the intramontane basin on the Iberian block decreased in size; sedimentation in the peri-Alpine Molasse Basin ended; the lateral migration of the foredeep depocenter in the Outer Carpathians ended; and the Pannonian back-arc basin
was interrupted by a rifting followed by a post-rift thermal subsidence 9-8 Ma that caused the termination of marine/brackish influence and the interruption of the connection with the foreland basins of the Outer Carpathians (Meulenkamp & Sissingh, 2003).
Meanwhile, around 8 Ma, the Betic and Rifian regions were uplifted, the Tyrrhenian and the Dead Sea basins opened, stable marine deposition in the Aegean region began, and the fore-Apennine basin depocenter migrated southward. These processes determined the main features of the Messinian scenario (Popov et al., 2006).
The only sirenian records from just before the Messinian Salinity Crisis (MSC) are those referred to the dwarfed species Metaxytherium serresii from southern Italy (at about 7.24 Ma) and from the Sahabi Formation of Libya (at about 6.8 Ma)
During the Messinian the Mediterranean basin underwent progressive isolation from the Atlantic Ocean, with consequent dessication. This isolation was caused principally by tectonic uplift along the African and Iberian continental margins that determined the closure of the Betic and Rifian corridors, respectively, between 7.6 Ma and 6.8 Ma (Garcés et al., 2001) and between at least 6.0 Ma and 5.65-5.4 Ma (Krijgsman et al., 1996, 1999; Warny et al., 2003).
This caused the destruction of the Mediterranean marine ecosystem with relevant effects on world oceanographic and climatic patterns (Thunnell et al., 1987). The Messinian event is documented in the stratigraphic record by (1) cyclic diatomite-sapropel sedimentation (7.25–5.96 Ma), (2) thick and extensive evaporite deposition (5.96–5.59 Ma) followed by a short time interval characterized by non-deposition and erosion (5.59-5.52 Ma, the “Messinian gap”), a small hiatus which was probably caused by the definitive isolation (Krijgsman et al. 1999); and (3) fresh- to brackish-water sedimentation (5.52–5.33 Ma, the “Lago-Mare” facies).
The second and third phases correspond to the Messinian Salinity Crisis (MSC) (Krijgsman et al., 1999).
At the end of the Messinian age (5.33 Ma), the Mediterranean returned to fully marine conditions throughout (the so-called “Terminal Messinian Flooding” of Krijgsman et al., 1999).
The scarcity of fossils and the heterogeneity of sediment that characterize the MSC deposits make difficult the understanding of the physico-chemical and palaeoenvironmental conditions of the water body and the resulting palaeogeographical setting of the Mediterranean during this event (Cita & Corselli, 1990). The classic model envisions a total desiccation of the whole basin (Hsü, 1984), but recent years
have brought a wealth of new data suggesting that normal marine conditions were present in the Mediterranean during the evaporitic episode and in the later portion of the post-evaporitic ‘Lago-mare’ phase, thereby implying marine connexions during the MSC and a marine refilling of the basin before the Messinian–Zanclean boundary. This evidence includes:
1) More or less stenohaline marine fish remains found in several Messinian intra-evaporitic series: Epinephelus sp., Zeus sp., and Harengula sp. from Monte Castellaro (Pesaro, Italy) (Sorbini, 1988); Trachurus sp. from Vena del Gesso (Romagne, Italy) (Gaudant, 2002); Mugil sp. from Cherasco (Piedmont, Italy) (Cavallo & Gaudant, 1987); Sardina? crassa from western Crete (Greece) (Gaudant, 1980); Sardinia crassa from Djebel Murdjadjo (Algeria) (Gaudant et al., 1997); Lepidopus sp. and
Spratelloides gracilis from Castagnito (Piedmont, northern Italy) (Fourtanier et al.,
1991); and Sardinia crassa from Molinos de Aguas quarry (Sorbas basin, SE Spain) (Néraudeau et al., 2002).
2) Leaves of seagrasses referred to Posidonia oceanica above the Messinian gypsum bed of the Piedmont basin (Sturani, 1976: 18, Fig. 5c).
3) Stenohaline marine bivalve association from the last levels of the Messinian gypsum of Los Yesos (Sorbas basin, SE Spain) (Lacour et al., 2002).
4) Stenohaline echinoid Brissopsis lyrifera from levels interbedded in the Messinian evaporitic series of Molinos de Aguas quarry (Néraudeau et al., 2002) and of Los Yesos (Lacour & Néraudeau, 2000) (Sorbas basin, SE Spain).
5) Fish otolith assemblage (Gobiidae; Myctophidae; Sciaenidae) from “Lago-mare” deposits ot the Colombacci Formation (central Italy) (Carnevale et al., 2006a).
6) Fish remains (also Myctophidae and Bythitidae) from the upper portion of the ‘Lago-mare’ sequence from Cava Serredi (central Italy) (Carnevale et al., 2006b).
7) Oxygen, carbon and strontium isotopic composition of fish otoliths and other fossils, coming from upper Messinian post-evaporitic deposits of Cava Serredi and Podere Torricella (central Italy) (Carnevale et al., 2008).
All these data support the existence of normal marine conditions at various locations in the Mediterranean Basin at least periodically during the MSC; therefore suitable habitats for sirenians could also have existed during the MSC. Stable isotope values for
Metaxytherium species, and in particular for M. serresii, indicate a high adaptability in
diet and habitats; therefore M. serresii appears to have been well adapted for living in small basins, lagoons and deltas, feeding on seagrasses and/or other aquatic plants. Moreover, M. serresii specimens are recorded at about 7.24 Ma from southern Italian
deposits; at about 6.8 Ma in Libyan deposits; at 5.25 - 4.98 Ma in Spanish deposits; and at 4 – 5.3 Ma in French deposits. From a palaeontological point of view, the M. serresii record appears nearly continuous in the Mediterranean Basin. On the contrary, sirenians appear to have died out on the northeastern Atlantic coasts during the Late Miocene: just some rib fragments from (probably) Messinian deposits of Portugal are signalled by Estevens (2000). Sirenians appear to return along the northeastern Atlantic coasts just in the Middle Pliocene, as recorded by Ennouchi (1954) in Morocco; however, the latter specimen might not belong to Metaxytherium but to a dugongine (pers. obs. and Domning, pers. comm.).
On the whole, there is no evidence to support the disappearance of M. serresii from the Mediterranean Basin during the MSC and its recolonization after the crisis from the northeastern Atlantic coasts; therefore the M. serresii record is here considered continuous from the uppermost Tortonian to the early Pliocene in the Mediterranean Basin.
7.5 PLIOCENE
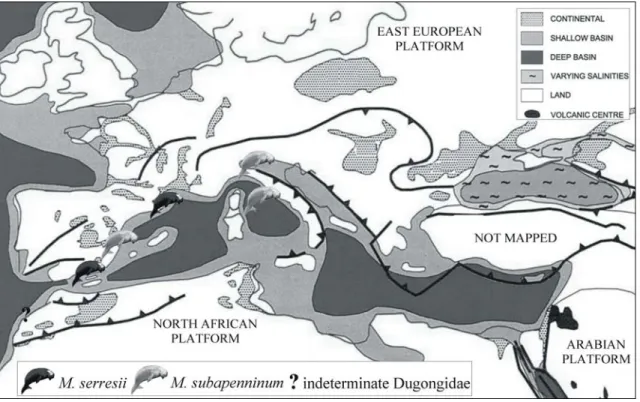
(Fig. 4)Fig. 4: Schematic palaeogeographic map of the Euro-North African coasts during the Pliocene (modified from Meulenkamp & Sissingh, 2003); with indications of the main known sirenian records.
During the Pliocene the overall uplift of Western and Central Europe continued: the second phase of rifting of the Rhine Graben from around 4-3 Ma created the present-day Rhine river system; the southern part of the Carpathian arc closed during the Middle and Late Pliocene; while on the contrary the Eastern Paratethys underwent a subsidence regime with formation of new basins (Meulenkamp & Sissingh, 2003).
Immediately after the end of the Messinian, sirenians were still about the same small size, as documented by the lower Pliocene record of Metaxytherium serresii from the sands of Montpellier (France). However, the subsequent species from Italy and Spain,
Metaxytherium subapenninum, was larger.
The Pliocene is marked by a subtle warming trend until 3.2 Ma when δ18O again increased, reflecting the onset of Northern Hemisphere glaciation (Zachos et al., 2001). Capozzi and Picotti (2003) recorded an acceleration of climatic cooling after 3 Ma and it is just during (or before?) this cold period that sirenians seem to die out in the Mediterranean Sea. Sirenians (of uncertain identity) also lived along the Atlantic coast of Morocco during the Middle Pliocene.
7.6 MAJOR CHANGE IN SIRENIAN DISTRIBUTION AND
EVOLUTION
The sirenian palaeobiogeographic evolution, here presented, encompassed changes which could be correlated with palaeogeographic and palaeoclimatic events related to the Neogene African-Eurasian convergence.
Eight major features of sirenian distribution and evolution are summarized below:
1. Presence of three sirenian subfamilies along the Euro-North African coasts during the Early Miocene: the Trichechidae Miosireninae represented by the genus Miosiren; the Dugongidae Dugonginae represented by the genus Rytiodus and the Dugongidae Halitheriinae represented by the genus Metaxytherium. 2. Sympatric presence of the large-tusked dugongine Rytiodus and the generalist
small-tusked halitheriine Metaxytherium in the Aquitaine Basin and along the Libyan coasts during the Early Miocene.
3. Survival of only the genus Metaxytherium at the Early-Middle Miocene transition, with the exception of Prohalicore dubaleni (referred to the Trichechidae Miosireninae) in the Serravallian deposits of the Aquitaine Basin. 4. Disappearance of sirenians from the Paratethys region during the Middle-Late
Miocene.
5. Disappearance of sirenians from the Euro-northeastern Atlantic coasts at the end of the Miocene.
6. Record of the dwarfed Metaxytherium serresii considered continuous from the uppermost Tortonian to the early Pliocene in the Mediterranean Basin.
7. Extinction of the last Mediterranean sirenians (Metaxytherium subapenninum) during the Pliocene at about 3.1 Ma.
8. Survival of sirenians along the Atlantic coast of Morocco during the Middle Pliocene.