CHAPTER 5
CLADISTIC ANALYSIS
5.1 INTRODUCTION
The systematic revisions presented in Chapter 4 consider as valid the following Neogene Euro-North African sirenian species: Miosiren kocki, Rytiodus capgrandi, Rytiodus sp., Prohalicore dubaleni, Metaxytherium krahuletzi, M. medium, M. petersi, M. serresii and M. subapenninum.
In order to clarify the relationships among these and the other known sirenian species, a cladistic analysis was undertaken.
I excluded from the cladistic analysis Metaxytherium petersi (under study by Domning and Pervesler) and Prohalicore dubaleni known only from a partial mandible. On the other hand, I include in the analysis the Oligocene “Halitherium” bellunense, here revised (see Chapter 4).
The main aims were to:
1. Clarify the origin and the relationships among the Metaxytherium species (Node 26 from Domning, 1994, unresolved).
2. Clarify the relationships of the genus Rytiodus (Node 4 from Bajpai & Domning, 1997, unresolved).
3. Determine the systematic position of “Halitherium” bellunense, considered by Domning (1989b, 1996) a dugongine, but whose affinities remain in doubt (Cladogram from Domning, 1989b: 425).
5.2 MATERIAL AND METHODS
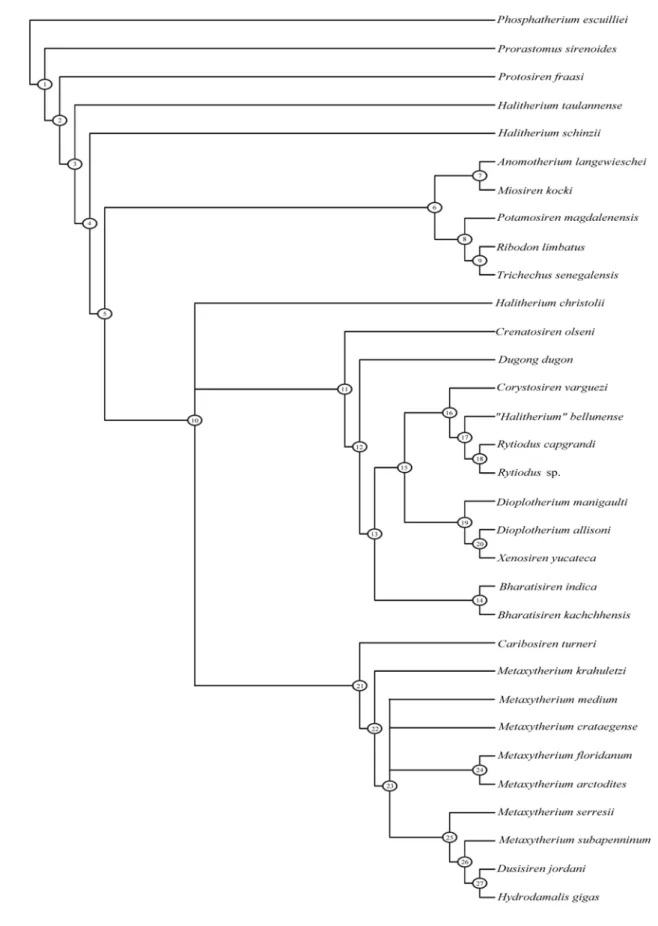
The cladistic analysis has been developed for 31 species of sirenians and one outgroup taxon (Tab. 1), following the previous analyses carried out by Domning (1994) and Bajpai and Domning (1997). Some characters come from these two studies, others were used in Domning and Thomas (1987) or in Sagne (2001), and the remaining are new. Some characters used in previous studies are redefined.
SPECIES REFERENCES Ma Nr
1 Phosphatherium escuilliei Gheerbrant et. al, 2005 53-56 4 2 Prorastomus sirenoides Savage et al., 1994; pers. obs. 46-50 1 3 Protosiren fraasi Gingerich et al., 1994 43-45 6 4 Anomotherium langewieschei Siegfried, 1965 23-25 1 5 Potamosiren magdalenensis Reinhart, 1951 12-16 1 6 Ribodon limbatus Ameghino, 1893; Heal, 1973 4-11 2
7 Miosiren kocki pers. obs. 21-23 1
8 Trichechus senegalensis Domning, 1994; pers. obs. 0 4 9 Halitherium taulannense Sagne, 2001; pers obs. 34-37 2 10 Halitherium schinzii Lepsius 1882; pers. obs. 28-31 22 11 Halitherium christolii Fitzinger, 1842 23-28 2 12 "Halitherium" bellunense pers. obs. 23-28 1
13 Crenatosiren olseni Domning, 1997 23-28 3
14 Dugong dugon pers. obs. 0 12
15 Bharatisiren indica Bajpai et al., 2006 23-28 1 16 Bharatisiren kachchhensis Bajpai & Domning, 1997 20-23 1 17 Rytiodus capgrandi Lartet, 1866; pers. obs. 18-23 1 18 Rytiodus sp. Heal, 1973; pers. obs. 17-23 2
19 Corystosiren varguezi Domning, 1990 3-5 4
20 Dioplotherium manigaulti Domning, 1989a 16-23 9 21 Dioplotherium allisoni Domning, 1987; Toledo & Domning, 1991 14-16 2
22 Xenosiren yucateca Domning, 1989b 4-11 1
23 Caribosiren turneri Reinhart, 1959 23-28 1
24 Metaxytherium krahuletzi Domning & Pervesler, 2001; pers. obs. 16-20 5 25 Metaxytherium medium Cottreau, 1928; pers. obs. 8-16 14 26 Metaxytherium floridanum Domning, 1988 11-13 52 27 Metaxytherium crataegense Kellogg, 1966; Aranda-Manteca et al., 1994 14-15 3 28 Metaxytherium arctodites Aranda-Manteca et al., 1994 14-15 3 29 Metaxytherium serresii Carone & Domning, 2007; pers. obs. 5-8 10
30 Metaxytherium subapenninum pers. obs. 3-5 6
31 Dusisiren jordani Domning, 1978; 5-11 8
32 Hydrodamalis gigas Domning, 1978; 1994; pers. obs. 0 3
Table 1: List of sirenian taxa and outgroup analyzed; with references; distribution range, approximated in millions of years; and the maximum number of specimens of each taxon for which any character was scored.
Three sirenian species, not included in the previous cladistic analyses, have been added: Halitherium taulannense Sagne, 2001; Metaxytherium arctodites Aranda-Manteca et al., 1994 and Metaxytherium crataegense (Simpson, 1932) (= M. calvertense Kellogg, 1966; see Aranda-Manteca et al., 1994).
Phosphatherium escuilliei has been chosen as outgroup as the most primitive representative of Proboscidea; Prorastomus sirenoides and Protosiren fraasi are representative respectively of Prorastomidae and Protosirenidae. Most of the Oligocene-Recent sirenian species have been included; while among the Eocene dugongid species only Halitherium taulannense is considered, being close to the Oligocene species of Halitherium. The study of the Eocene species is not the aim of this analysis.
The analysis is based on 57 unweighted morphological characters of cranium, mandible and dentition. Cheek-tooth cusp patterns are excluded owing to their high intraspecific variability.
These characters were analyzed with the PAUP version 40b10 computer program (Swofford, 1998). Multistate characters 31, 37, 46, 47, 82, 102, 122, 125, 126 and 136 have been treated as unordered because in these cases there are significant doubts that the states formed a single transformation series.
Some characters used showed intraspecific variation for some species. These polymorphisms have been scored unambiguously as one of two states, following the procedure adopted by Domning (1994: 177-178).
5.3 CHARACTERS USED
For this cladistic analysis, the following 57 unweighted morphological characters of cranium, mandible and dentition were scored. The data matrix for the 57 characters is shown in Table 2. 0 designates the most primitive state among taxa studied; 1, 2, and 3 are alternate derived states; ? indicates unknown or missing data; - indicates character not applicable (? and – have been treated alike).
CRANIUM
1 Nasal process of premaxilla: (0) does not contact lacrimal; (1) contacts lacrimal (from
Sagne, 2001).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 ? 0 0 ? ? ? 0 0 1 1 ? ? 1 1 ? 1 1 1 1 1 1 ? 1 1 1 1 1 0 1 0 0 0
3 Rostrum: (0) length of premaxillary symphysis < 0.27 X condylobasal skull length;
(1) length of premaxillary symphysis > 0.27 X condylobasal skull length (from Domning, 1994).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 0 1 ? ? ? 0 0 1 1 ? ? 1 1 1 1 1 1 1 1 1 ? 1 1 1 1 1 1 1 1 1 1
6 Nasal process of premaxilla: (0) tapering at the posterior end, having long overlap
with frontal and/or nasal; (1) tapering but thickened at posterior end, with long overlap of frontal and/or nasal; (2) broadened and bulbous at the posterior end, having a more or less vertical joint surface in contact with frontal (redefined from Domning, 1994 and Baipaj & Domning, 1997).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 0 0 ? ? ? 0 0 0 0 ? 2 0 0 1 1 2 2 ? 2 2 2 0 0 0 0 0 0 0 0 0 0
7 Nasal process of premaxilla: (0) long; (1) short (from Domning, 1994).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 0 0 ? ? ? 0 0 0 0 ? 1 0 0 0 0 1 1 1 0 0 ? 0 0 0 0 0 0 0 0 0 0
8 External nares: (0) not retracted; (1) retracted and enlarged, reaching to the level of
the anterior margin of the orbit; (2) retracted and enlarged beyond the front of the orbit (redefined from Domning, 1994).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 1 1 ? ? ? 1 2 1 1 ? ? 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2
9 Premaxilla: (0) does not contact frontal; (1) contacts frontal (from Domning, 1994).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 1 1 ? ? ? 1 1 1 1 ? ? 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1
10 Premaxillary symphysis: (0) not laterally compressed; (1) laterally compressed
(redefined from Baipaj & Domning, 1997).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 0 ? ? ? ? 1 1 1 1 ? 1 1 1 1 1 ? 1 1 1 1 ? 1 1 1 1 1 1 1 1 1 1
11 Zygomatic-orbital bridge of maxilla: (0) nearly level with palate; (1) elevated above
palate, with its ventral surface lying ≥ 1 cm above the alveolar margin (from Domning, 1994).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 1 1 0 ? 1 1 1 1 0 0 0 ? 0 0 0 0 ? 0 ? 0 0 0 0 1 1 1 1 1 1 1 1 1
13 Infraorbital foramen: (0) small (diametres about 15 by 10 mm or less); (1) large
(diametres greater than 15 by 10 mm but less than about 30 mm); (2) very large (diametres greater than about 30 mm) (from Domning, 1994, redefined by Baipaj & Domning, 1997).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 0 0 ? ? 1 1 1 1 1 ? ? 1 2 1 1 1 2 ? 1 1 ? 1 1 1 1 1 1 1 2 1 1
16 Palate: (0) thin or incomplete at level of penultimate cheek tooth; (1) > 1 cm thick at
level of penultimate cheek tooth (from Domning, 1994).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 0 0 ? 0 1 1 1 0 0 0 ? 0 1 0 0 ? 0 ? 0 1 1 0 0 0 0 0 0 0 0 0 0
20 Infraorbital foramen: (0) not obstructed; (1) partly obstructed by a transverse bony
ridge (from Baipaj & Domning, 1997).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 0 0 ? ? 0 0 0 0 0 ? ? 0 0 0 1 0 0 0 1 1 1 0 0 0 0 0 0 0 0 0 0
31 Nasals: (0) meet in midline; (1) separated in midline by nasal incisure and/or
frontals; (2) absent or fused with the frontals (redefined from Domning, 1994. Unordered).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 0 0 0 ? ? 0 2 0 0 ? ? 2 2 2 2 1 1 2 2 2 2 1 1 1 1 1 1 1 1 1 2
36 Supraorbital process of frontal: (0) well developed, dorsoventrally thin (< 1 cm
thick), with prominent, dorsoventrally flattened posterolateral corner; (1) dorsoventrally thickened, with posterolateral corner variably developed (from Domning, 1994, redefined by Baipaj & Domning, 1997).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 0 0 0 ? ? 1 0 0 0 0 ? 0 1 1 0 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1
37 Nasal incisure at posterior end of mesorostral fossa: (0) absent (1) small (does not
extend posterior to the supraorbital process, but separating the nasals); (2) deep and narrow (extends posterior to the supraorbital process); (3) comparably deep but broad, with the anterior frontal margin displaying a median convexity (redefined from Domning, 1994. Unordered).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 0 0 0 ? ? 0 0 0 0 0 ? 2 3 2 2 2 2 3 2 2 2 0 0 1 1 1 2 1 0 0 0
38 Lamina orbitalis of frontal: (0) thin or absent; (1) 1 cm thick (from Domning, 1994).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 0 0 1 ? 0 1 0 0 0 0 ? 0 0 0 0 0 0 0 0 ? 0 0 0 0 0 0 0 0 0 0 0
42 Frontal roof: (0) convex, or more or less flat between temporal crests (if latter
present); (1) deeply concave, sloping steadily ventrad to anterior margin (from Domning, 1994).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 0 0 0 ? ? 0 0 0 0 0 0 0 1 0 1 1 1 1 1 1 1 0 0 0 0 0 0 0 0 0 0
43 Supraorbital process of frontal: (0) flattened in more or less horizontal plane, with
dorsal surface inclined relatively gently ventrolaterad; (1) turned markedly downward, with dorsal surface inclined strongly ventrolaterad and posterolateral corner projecting posteriorly (from Domning, 1994).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 0 0 0 ? ? 0 0 0 0 0 ? 1 0 1 1 1 1 ? 1 1 1 0 0 0 0 0 0 0 0 0 0
45 Parietal roof: (0) convex; (1) concave or flat (new).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 0 1 0 ? ? 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 ? 1 1 1 1 1 1 1 1 1 1
46 Intertemporal constriction: (0) present, anterior to the braincase; (1) present, at the
centre of the braincase; (2) absent (redefined from Domning & Thomas, 1987. Unordered).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 0 0 2 ? ? 2 2 1 1 2 2 2 2 1 1 1 1 2 2 2 ? 2 2 2 2 2 2 2 2 2 2
47 Bony falx cerebri: (0) absent; (1) present (new).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 ? 0 ? ? ? 1 1 1 1 1 ? 1 1 ? ? 1 1 1 1 1 ? 1 1 1 1 1 1 1 1 1 1
51 Sagittal crest: (0) present; (1) absent (from Domning, 1994).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 1 1 1 ? ? 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 ? 1 1 1 1 1 1 1 1 1 1
65 Paroccipital process: (0) long; (1) short, does not reach as far ventrally as occipital
condyle, or absent (new).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 0 0 1 ? ? 0 1 0 0 ? ? 1 1 1 1 ? ? ? 1 1 ? ? 1 1 1 1 1 1 1 1 1
66 Exoccipitals: (0) meet in suture dorsal to foramen magnum; (1) do not meet in a
suture (from Domning, 1994).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 0 0 0 ? ? 0 0 0 0 0 ? 0 1 0 0 1 1 0 0 0 ? ? 0 1 1 1 1 1 1 1 1
67 Supracondylar fossa of exoccipital: (0) absent; (1) deep and extending across entire
width of occipital condyle; (2) shallow or reduced and located dorsomedial to condyle (redefined from Domning, 1994. Unordered).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 0 2 1 ? ? 1 0 1 1 ? ? 1 1 1 1 1 ? 2 1 1 ? ? 1 1 1 1 1 2 2 0 0
70 Dorsolateral border of exoccipital: (0) rounded and more or less smooth, not
flangelike; (1) thick and overhanging posteriorly as a flange (redefined from Domning, 1994).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 0 0 0 ? ? 0 0 0 0 ? ? 0 0 0 0 0 0 0 0 0 ? ? 0 0 0 0 0 0 0 1 1
73 Posttympanic process of squamosal: (0) absent (i.e. no facet projecting for
sternomastoid muscle); (1) present (redefined from Domning, 1994).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 0 0 1 ? ? 1 1 1 1 1 ? 1 1 1 1 1 1 1 ? ? ? 1 1 1 1 1 1 1 1 1 1
76 Squamosal: (0) does not extend to temporal crest; (1) extend to temporal crest (from
Domning, 1994).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 0 0 1 ? ? 1 1 1 1 1 0 1 1 1 1 ? 0 0 1 0 ? 1 1 1 1 1 1 1 1 1 1
77 Processus retroversus of squamosal: (0) absent; (1) present (redefined from
Domning, 1994).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 0 0 0 ? ? 0 0 1 1 1 1 1 1 1 1 1 1 1 ? 1 1 1 1 1 1 1 1 1 1 1 0
82 External auditory meatus of squamosal: (0) narrow and slitlike (anteroposterior
breadth less than dorsoventral); (1) about as wide anteroposteriorly as high; (2) very broad and shallow, wider anteroposteriorly than high (from Domning, 1994. Unordered).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 0 0 2 ? ? 2 2 1 1 1 ? 1 1 1 1 ? ? ? ? ? ? 1 1 1 1 1 1 1 1 1 1
83 Zygomatic process of squamosal: (0) narrow throughout its length; (1)
lozenge-shaped, broader posteriorly than anteriorly (new).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 ? 0 ? ? ? 1 0 1 1 1 1 1 1 0 ? 1 1 1 ? ? ? 1 0 1 1 1 1 1 1 1 1
85 Ventral extremity of jugal: (0) lies posterior to orbit; (1) lies approximately under
posterior edge of orbit, but forward of jugal’s postorbital process (if present); (2) lies ventral to orbit (from Domning, 1994).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 0 0 ? ? ? 2 0 0 1 ? 2 1 2 2 2 2 2 2 2 2 1 ? 1 2 2 2 2 2 2 2 2
86 Postorbital process of jugal: (0) absent; (1) present (new).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 ? 1 ? ? ? 1 1 1 1 ? 1 0 0 0 0 ? 1 ? 0 1 1 ? 0 0 1 0 1 0 0 0 0
87 Preorbital process of jugal: (0) does not contact premaxilla; (1) contacts premaxilla
(from Domning, 1994).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 0 0 ? ? ? 0 0 0 0 ? ? 0 1 0 0 0 0 ? 0 0 ? 0 0 0 0 0 1 0 1 1 1
88 Preorbital process of jugal: (0) absent; (1) relatively flat and thin
(posteromedial-anterolateral breadth of portion lateral to maxillojugal suture > anteromedial-posterolateral thickness; (2) thick and robust (breadth < thickness) (redefined from Domning, 1994).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 1 1 ? ? ? 1 1 1 1 ? ? 1 2 2 2 ? 1 2 2 2 2 ? 1 1 1 1 1 1 1 1 1
89 Zygomatic process of jugal: (0) with a posterior longitudinal process; (1) without a
posterior longitudinal process (redefined from Domning, 1994).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 ? 0 ? ? ? 0 0 0 0 ? 0 0 0 0 0 0 0 0 1 1 1 0 0 0 0 0 0 0 0 0 0
90 Ventral rim of orbit: (0) does not distinctly overhang lateral surface of jugal; (1)
does distinctly overhang (from Baipaj & Domning, 1997).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 0 0 ? ? ? 0 0 0 0 ? 0 1 1 ? 1 ? 1 ? 1 1 1 0 0 0 0 0 0 0 0 0 0
91 Lacrimals: (0) with nasolacrimal canal; (1) without nasolacrimal canal, but still
large; (2) vestigial or absent (from Domning, 1994).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 ? 0 ? ? ? ? 1 2 0 ? ? ? 1 1 1 1 ? 1 1 1 1 ? 1 1 1 1 1 1 1 1 2 2
97 Posterior border of palate: (0) not incised, merely shallowly concave; (1) incised or
deeply indented (redefined from Domning, 1994).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 0 0 ? ? ? 0 1 1 1 ? ? ? 1 1 1 1 1 ? 1 1 ? 1 1 1 1 1 1 1 1 1 1
99 Palatines: (0) extend anteriorly beyond posterior edge of zygomatic-orbital bridge;
(1) do not extend so far forward (from Domning, 1994).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 0 0 ? 1 1 0 1 0 0 ? ? 0 1 0 0 ? 0 ? 0 0 0 0 0 0 0 0 0 1 1 1 1
101 Alisphenoid canal: (0) present; (1) absent (from Domning, 1994).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 0 0 1 ? ? 1 1 1 1 1 ? 1 1 1 1 ? 1 ? 1 1 ? 1 1 1 1 1 1 1 1 1 1
102 Pterygoid fossa: (0) absent; (1) present, extending above level of roof of internal
nares; (2) reduced, confined to below roof of internal nares (from Domning, 1994, redefined by Baipaj & Domning, 1997. Unordered).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 1 0 1 ? ? 1 1 1 1 1 ? 1 1 1 1 ? 1 ? 2 2 ? 1 1 1 1 1 1 1 1 1 1
MANDIBLE
122 Ventral border of horizontal mandibular ramus: (0) straight or only slightly
concave; (1) moderately concave, sharply downturned anteriorly; (2) moderately and evenly concave; (3) strongly concave (from Domning, 1994. Unordered).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 0 1 1 2 2 ? 2 1 1 3 ? 3 3 3 ? ? 3 ? ? 3 ? ? 3 3 3 3 3 3 3 2 2
123 Accessory mental foramina: (0) present, in addition to and usually posterior to the
large principal foramen; (1) absent (from Doming, 1994).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 0 0 1 0 0 ? 0 0 0 1 ? 1 1 ? ? ? 1 ? ? 0 ? ? 1 1 1 1 1 1 1 1 1
125 Posterior border of mandible: (0) descends ventrally or posteroventrally from
condyle without marked interruption or abrupt change of direction; (1) bears a processus angularis superior below condyle; (2) has not a distinct processus angularis superior, but does have broadly convex outline beginning well below condyle (from Domning, 1994. Unordered).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 0 ? 2 ? ? ? 2 2 2 2 ? 2 2 1 ? ? ? ? ? 2 ? ? 2 2 2 2 2 2 2 2 2
126 Anterior border of coronoid process: (0) approximately vertical; (1) extends
slightly anterior to base of process; (2) extends very far anterior to base (from Domning, 1994. Unordered).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 0 ? ? ? ? ? 2 1 1 1 ? 1 1 1 ? ? 1 ? ? 1 ? ? 1 1 1 1 1 1 1 1 0
128 Horizontal ramus of mandible: (0) slender (minimum dorsoventral height < 0.25 X
length of mandible); (1) broad dorsoventrally (height > 0.25 X length) (from Domning, 1994).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 0 0 0 0 0 ? 0 0 0 1 ? 1 1 1 ? ? 1 ? ? 1 ? ? 1 1 1 1 1 1 1 0 0
129 Ventral border of horizontal ramus of mandible: (0) tangent to angle; (1) not
tangent to angle (from Domning, 1994).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 0 ? 0 0 0 ? 1 0 1 1 ? 1 1 ? ? ? ? ? ? 1 ? ? 1 1 1 1 1 1 1 1 1
DENTITION
136 First upper incisors: (0) with enamel on all sides, forming complete enamel crown;
(1) with enamel mainly on lateral side; (2) with enamel mainly on medial side (redefined from Domning, 1994. Unordered).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 1 0 ? ? ? 0 - 0 0 ? 0 0 2 ? ? 2 2 2 ? 2 2 ? 0 0 0 0 0 0 0 -
-137 First upper incisors: (0) enamel crown distinct from root; (1) enamel extends entire
length of tusk (from Domning, 1994).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 1 0 ? ? ? 0 - 0 0 ? ? 0 1 ? ? 1 1 1 1 1 1 ? 0 0 0 0 0 0 0 -
-139 First upper incisors: (0) present; (1) vestigial or absent (from Domning, 1994).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 0 0 ? ? ? 0 1 0 0 ? 0 0 0 0 0 0 0 0 0 0 0 ? 0 0 0 0 0 0 0 1 1
140 Depth of I1 alveolus: (0) much less than half the length of the premaxillary
symphysis; (1) about half the length of the symphysis; (2) much greater than half length of the symphysis (from Domning, 1994).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 1 0 ? ? ? 2 - 1 1 ? 1 1 2 2 2 2 2 2 2 2 2 0 0 0 0 0 0 1 2 - -
141 Cross section of I1 crown: (0) suboval or subelliptical; (1) lens-shaped or
lozenge-shaped; (2) broad and extremely flattened mediolaterally (modified from Domning, 1994. Unordered).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 0 0 ? ? ? 0 - 0 0 ? 1 0 0 0 0 2 2 2 1 1 ? ? 0 0 0 0 0 0 0 - -
143 I2; I3; i1; i2; i3: (0) present; at least in part (1) all absent (from Domning, 1994).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 0 0 1 1 1 1 1 0 1 1 1 1 1 1 1 ? 1 1 1 1 ? 1 1 1 1 1 1 1 1 1 1
144 Canines: (0) present; (1) absent (redefined from Domning, 1994).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 0 0 1 1 1 1 1 0 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1 1
150 Supernumerary molars: (0) absent; (1) present and replenished indefinitely by
horizontal replacement (from Domning, 1994).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 0 0 0 0 1 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
155 Postcanine dental formula: (0) P/p1-4, M/m 1-3; (1) P/p1-5, M/m1-3; (2)
secondarily reduced from P/p1-5, M/m1-3, by loss of anterior premolars (redefined from Domning, 1994).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 1 1 2 2 2 2 2 1 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2
157 Permanent premolars: (0) some double- or triple-rooted; (1) all single-rooted; (2)
all absent (from Domning, 1994).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 0 0 1 1 2 2 1 2 1 1 ? 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2 2
Table 2: Matrix of character-state data for sirenian taxa and outgroup analyzed (following page).
CHARACTER STATE SPECIES 1 1 0 1 1 0 2 1 2 2 1 2 3 1 2 1 2 6 1 2 8 1 2 9 1 3 6 1 1 3 6 7 8 9 1 0 1 1 1 3 1 6 2 0 3 1 3 6 3 7 3 8 4 2 4 3 4 5 4 6 4 7 5 1 6 5 6 6 6 7 7 0 7 3 7 6 7 7 8 2 8 3 8 5 8 6 8 7 8 8 8 9 9 0 9 1 9 7 9 9 5 3 7 1 3 9 1 4 0 1 4 1 1 4 3 1 1 1 5 5 4 5 5 7 4 0 Phosphatherium escuilliei 1 ? 0 0 0 0 0 0 1 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 ? 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 2 Prorastomus sirenoides 0 0 0 0 1 1 0 1 0 0 0 0 0 0 0 0 0 0 0 ? 1 0 0 0 0 0 0 0 0 ? 0 ? 0 1 ? 0 0 0 0 0 1 0 0 0 0 0 0 1 1 0 1 0 0 0 0 1 0 3 Protosiren fraasi 0 1 0 0 1 1 ? 0 0 0 0 0 0 0 0 0 0 1 0 0 1 0 0 2 0 0 0 0 0 0 0 1 0 1 0 0 ? 0 0 0 0 1 0 ? ? 0 ? 0 0 0 0 0 0 0 0 1 1 4 Anomotherium langewieschei ? ? ? ? ? ? ? ? ? ? ? 0 0 0 1 0 0 0 2 ? 1 1 0 1 0 1 1 0 2 ? ? ? ? ? ? ? ? ? ? 1 1 1 1 2 ? 0 0 ? ? ? ? ? 1 1 0 2 1 5 Potamosiren magdalenensis ? ? ? ? ? ? ? 1 ? 0 ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? 1 ? ? 2 0 ? ? 0 0 ? ? ? ? ? 1 1 0 2 2 6 Ribodon limbatus ? ? ? ? ? ? ? 1 1 1 0 ? ? ? 0 ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? ? 1 ? ? 2 0 ? ? 0 0 ? ? ? ? ? 1 1 1 2 2 7 Miosiren kocki 0 0 0 0 1 1 1 1 1 1 0 0 1 0 1 0 0 0 2 1 1 0 0 1 0 1 1 0 2 1 2 1 0 1 0 0 1 0 0 1 1 ? ? ? ? ? ? 0 0 0 2 0 1 1 0 2 1 8 Trichechus senegalensis 0 0 0 0 2 1 1 1 1 1 0 2 0 0 0 0 0 1 2 1 1 1 0 0 0 1 1 0 2 0 0 1 0 1 0 0 2 1 1 1 1 2 0 2 2 0 1 - - 1 - - 1 1 1 2 2 9 Halitherium taulannense 1 1 0 0 1 1 1 0 1 0 0 0 0 0 0 0 0 1 1 1 1 0 0 1 0 1 1 1 1 1 0 1 0 1 0 0 0 1 0 1 1 1 0 2 1 0 0 0 0 0 1 0 0 0 0 1 1 10 Halitherium schinzii 1 1 0 0 1 1 1 0 1 0 0 0 0 0 0 0 0 1 1 1 1 0 0 1 0 1 1 1 1 1 1 1 0 1 0 0 ? 1 0 1 1 1 0 2 1 0 1 0 0 0 1 0 1 1 0 2 1 11 Halitherium christolii ? ? ? ? ? ? ? 0 ? 0 ? ? 0 0 0 0 0 1 2 1 1 ? ? ? ? 1 1 1 1 1 ? ? ? ? ? ? ? ? ? 1 1 3 1 2 1 1 1 ? ? ? ? ? 1 1 0 2 ? 12 "Halitherium" bellunense ? ? 2 1 ? ? 1 ? ? ? ? ? ? ? ? 0 ? 1 2 ? 1 ? ? ? ? ? 0 1 ? 1 2 1 ? ? 0 0 ? ? ? ? ? ? ? ? ? ? ? 0 ? 0 1 1 1 1 0 2 2 13 Crenatosiren olseni 1 1 0 0 2 1 1 0 1 0 0 2 0 2 0 0 1 1 2 1 1 1 0 1 0 1 1 1 1 1 1 0 0 1 0 1 1 ? 0 1 1 3 1 2 1 1 1 0 0 0 1 0 1 1 0 2 2 14 Dugong dugon 1 1 0 0 2 1 1 0 2 1 0 2 1 3 0 1 0 1 2 1 1 1 1 1 0 1 1 1 1 1 2 0 1 2 0 1 1 1 1 1 1 3 1 2 1 1 1 2 1 0 2 0 1 1 0 2 2 15 Bharatisiren indica ? 1 1 0 2 1 1 0 1 0 0 2 1 2 0 0 1 1 1 ? 1 1 0 1 0 1 1 1 1 0 2 0 0 2 0 ? 1 1 0 1 1 3 ? 1 1 1 ? ? ? 0 2 0 1 1 0 2 2 16 Bharatisiren kachchhensis 1 1 1 0 2 1 1 0 1 0 1 2 0 2 0 1 1 1 1 ? 1 1 0 1 0 1 1 1 1 ? 2 0 0 2 0 1 1 1 0 1 1 ? ? ? ? ? ? ? ? 0 2 0 1 1 0 2 2 17 Rytiodus capgrandi 1 1 2 1 2 1 ? ? 1 ? 0 1 1 2 0 1 1 1 1 1 1 ? 1 1 0 1 ? 1 ? 1 2 ? 0 ? 0 ? ? 1 ? ? ? ? ? ? ? ? ? 2 1 0 2 2 ? 1 0 2 2 18 Rytiodus sp. 1 1 2 1 2 0 1 0 2 0 0 1 1 2 0 1 1 1 1 1 1 ? 1 ? 0 1 0 1 ? 1 2 1 0 1 0 1 1 1 0 1 1 3 1 ? 1 1 ? 2 1 0 2 2 1 1 0 2 2 19 Corystosiren varguezi 1 1 ? 1 2 1 1 ? ? ? 0 2 1 3 0 1 ? 1 2 1 1 ? 0 2 0 1 0 1 ? 1 2 ? ? 2 0 ? 1 ? ? ? ? ? ? ? ? ? ? 2 1 0 2 2 1 1 0 2 2 20 Dioplotherium manigaulti 1 1 2 0 2 1 1 0 1 0 1 2 1 2 0 1 1 1 2 1 1 1 0 1 0 ? 1 ? ? ? 2 0 0 2 1 1 1 1 0 1 2 ? ? ? ? ? ? ? 1 0 2 1 1 1 0 2 2 21 Dioplotherium allisoni 1 1 2 0 2 1 1 0 1 1 1 2 0 2 ? 1 1 1 2 1 1 1 0 1 0 ? 0 1 ? ? 2 1 0 2 1 1 1 1 0 1 2 3 0 2 1 1 1 2 1 0 2 1 1 1 0 2 2 22 Xenosiren yucateca ? ? 2 ? 2 1 ? 0 ? 1 1 2 1 2 0 1 1 ? ? ? ? ? ? ? ? ? ? 1 ? ? 1 1 ? 2 1 1 ? ? 0 ? ? ? ? ? ? ? ? 2 1 0 2 ? ? 1 0 2 2 23 Caribosiren turneri 1 1 0 0 2 1 1 0 1 0 0 1 1 0 0 0 0 1 2 1 1 ? ? ? ? 1 1 1 1 1 ? ? 0 ? 0 0 1 1 0 1 1 ? ? ? ? ? ? ? ? ? 0 ? 1 1 0 2 2 24 Metaxytherium krahuletzi 1 1 0 0 2 1 1 1 1 0 0 1 1 0 0 0 0 1 2 1 1 1 0 1 0 1 1 1 1 0 1 0 0 1 0 0 1 1 0 1 1 3 1 2 1 1 1 0 0 0 0 0 1 1 0 2 2 25 Metaxytherium medium 1 1 0 0 2 1 1 1 1 0 0 1 1 1 0 0 0 1 2 1 1 1 1 1 0 1 1 1 1 1 2 0 0 1 0 0 1 1 0 1 1 3 1 2 1 1 1 0 0 0 0 0 1 1 0 2 2 26 Metaxytherium floridanum 1 1 0 0 2 1 1 1 1 0 0 1 1 1 0 0 0 1 2 1 1 1 1 1 0 1 1 1 1 1 2 1 0 1 0 0 1 1 0 1 1 3 1 2 1 1 1 0 0 0 0 0 1 1 0 2 2 27 Metaxytherium crataegense 1 1 0 0 2 1 1 1 1 0 0 1 1 1 0 0 0 1 2 1 1 1 1 1 0 1 1 1 1 1 2 0 0 1 0 0 1 1 0 1 1 3 1 2 1 1 1 0 0 0 0 0 1 1 0 2 2 28 Metaxytherium arctodites 0 1 0 0 2 1 1 1 1 0 0 1 1 2 0 0 0 1 2 1 1 1 1 1 0 1 1 1 1 1 2 1 1 1 0 0 1 1 0 1 1 3 1 2 1 1 1 0 0 0 0 0 1 1 0 2 2 29 Metaxytherium serresii 1 1 0 0 2 1 1 1 1 0 0 1 1 1 0 0 0 1 2 1 1 1 1 2 0 1 1 1 1 1 2 0 0 1 0 0 1 1 1 1 1 3 1 2 1 1 1 0 0 0 1 0 1 1 0 2 2 30 Metaxytherium subapenninum 0 1 0 0 2 1 1 1 2 0 0 1 1 0 0 0 0 1 2 1 1 1 1 2 0 1 1 1 1 1 2 0 1 1 0 0 1 1 1 1 1 3 1 2 1 1 1 0 0 0 2 0 1 1 0 2 2 31 Dusisiren jordani 0 1 0 0 2 1 1 1 1 0 0 1 1 0 0 0 0 1 2 1 1 1 1 0 1 1 1 1 1 1 2 0 1 1 0 0 2 1 1 1 1 2 1 2 1 0 1 - - 1 - - 1 1 0 2 2 32 Hydrodamalis gigas 0 1 0 0 2 1 1 1 1 0 0 2 1 0 0 0 0 1 2 1 1 1 1 0 1 1 1 0 1 1 2 0 1 1 0 0 2 1 1 1 1 2 1 2 0 0 1 - - 1 - - 1 1 0 2 2
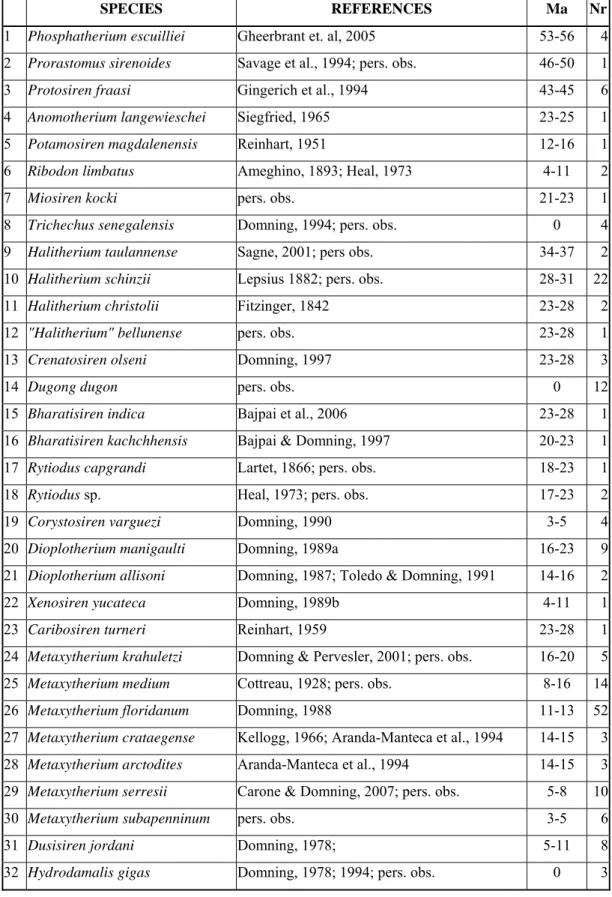
Fig. 1: Strict consensus tree of sirenians taxa and outgroup, generated by PAUP version 40b10. Tree length 157 steps; consistency index (CI) 0.50; retention index (RI) 0.74. Note that node 26 is probably spurious (see text).
5.4 RESULTS
The matrix of character-state data for 31 species of sirenians is given in Table 2; in addition, these characters were scored for one outgroup taxon, Phosphatherium escuilliei, the most primitive known proboscidean (Gheerbrant et al., 2005).
This cladistic analysis produced 6 maximally parsimonious trees, all of them 157 steps long, with a consistency index (CI) of 0.50 and retention index (RI) of 0.74.
The the strict consensus tree is a tree in which all the nodes are supported at 100%, and in which nodes 10 and 23 are unresolved (see discussion below).
The character transformations supporting the nodes of the tree are listed below.
The letters “r” and “c” after character-state changes indicate reversals and convergences respectively; the numbers after the letter c indicate the other nodes at which the convergence occurred (or, in the case of convergences in terminal taxa, the nodes under which the convergence is discussed below). Convergences and reversals are summarized in Table 3 Autapomorphies of terminal taxa are listed if any are known. Some other significant cranial and postcranial characters, not included in this cladistic analysis, are also listed in order to support various part of the tree. Some significant polymorphisms are noted where they occur. Unresolved nodes and missing data are also discussed.
Node 1 (Sirenia; one branch forms the possibly paraphyletic taxon Prorastomidae):
Non homoplastic characters: 51(1), 155(1).
Homoplastic characters: 8(1) ri = 0.80, 9(1) ri = 0.00; 88(1) ri = 0.83, 102(1) ri = 0.66. Autapomorphies of Prorastomus sirenoides:
Homoplastic characters: 136(1) ri = 0.80, 137(1) ri = 0.85, 140(1) ri = 0.64 c 3 16 25.
Node 2 (Sirenia excluding Prorastomidae; one branch forms the possibly paraphyletic taxon
Protosirenidae):
Non homoplastic characters: 10(1), 125(2).
Homoplastic characters: 3(1) ri = 0.66, 11(0)r ri = 0.83, 45(1) ri = 0.66, 67(1) ri = 0.42, 86(1) ri = 0.60 c 16 20 24, 122(1) ri = 0.87, 126(1) ri = 0.50, 157(1) ri = 0.85.
Autapomorphies of Protosiren fraasi:
Homoplastic characters: 67(2) ri = 0.42 c 17 25, 102(0)r ri = 0.50.
Node 3 (Halitherium taulannense + Oligocene–Recent Sirenia):
Non homoplastic characters: 47(1), 73(1), 82(1), 101(1).
Homoplastic characters: 1(1) ri = 0.57, 13(1) ri = 0.50, 46(1) ri = 0.71, 76(1) ri = 0.66 c 19, 77(1) ri = 0.66, 83(1) ri = 0.25, 97(1) ri = 0.66, 140(1) ri = 0.64 c 1 16 25.
Non homoplastic characters: 143(1), 144(1), 155(2).
Homoplastic characters: 85(1) ri = 0.66, 91(1) ri = 0.66, 129(1) ri = 0.60 c 9.
Node 5 (Oligocene – Recent Sirenia excluding Halitherium schinzii):
Homoplastic characters: 46(2) ri = 0.71, 65(1) ri = 0.80, 123(1) ri =, 128(1) ri = 0.75 c 5.
Node 6 (Trichechidae):
Non homoplastic character: 82(2).
Homoplastic characters: 1(0)r ri = 0.57 c 24 26, 3(0)r ri = 0.66, 11(1) ri = 0.83 c 22, 16(1) ri = 0.40 c 12 20, 77(0)r ri = 0.66 c 27, 126(2) ri = 0.50, 129(0)r ri = 0.60.
In this analysis Trichechidae appear to be stably rooted well within the Oligocene-Recent Dugongidae; and characters 1, 3, 77 and 129 are here regarded as reversals, but they could be also regarded as primitive.
Node 7 (Miosireninae):
Non homoplastic character: 38(1). Homoplastic character: 45(0)r ri = 0.66. Autapomorphies of Miosiren kocki:
Homoplastic characters: 36(1) ri = 0.60 c 10, 65(0)r ri = 0.80, 85(2) ri = 0.66 c 12 23, 97(0)r ri = 0.66, 140(2) ri = 0.64 c 12 26. Also posttympanic process of squamosal enlarged and clublike and reduction and simplification of M3. Characters 85, 97 and 140 are not scored for Anomotherium langewieschei, so I can not rule out that these transformations occur in the whole Miosireninae clade, at node 7. Moreover, the lower dentition of Anomotherium langewieschei also shows a tendency to reduction of the third molar (see Siegfried, 1965: tab. 11, fig 4).
Node 8 (Trichechinae):
Homoplastic characters: 99(1) ri = 0.71 c 12 25, 122(2) ri = 0.87 c 27, 123(0)r ri = 0.75 c 20, 157(2) ri = 0.85 c 10.
Autapomorphies of Potamosiren magdalenensis: Homoplastic character: 16(0)r ri = 0.40.
Node 9 (Ribodon limbatus + Trichechus senegalensis):
Non homoplastic character: 150(1).
Autapomorphies of Trichechus senegalensis:
Homoplastic characters: 8(2) ri = 0.80 c 10, 31(2) ri = 0.80 c 11 27, 67(0)r ri = 0.42 c 27, 83(0)r ri = 0.25 c 14 22, 85(0)r ri = 0.66, 91(2) ri = 0.66 c 27, 129(1) ri = 0.60 c 4, 139(1) ri = 0.50 c 27.
either in Ribodon limbatus or in Potamosiren magdalenensis, so I can not rule out that the trasformations occur at node 9 or at node 8.
Node 10 Unresolved (Halitherium christolii + Dugonginae + Caribosiren
turneri-Metaxytherium spp.-Hydrodamalinae):
Homoplastic characters: 122(3) ri = 0.87, 128(1) ri = 0.90.
Moreover the homoplastic characters 8(2) ri = 0.80 c 9, 86(0)r ri = 0.60, and 157(2) ri = 0.85 c 8 are autoapomorphies of the clade Dugonginae + Caribosiren turneri-Metaxytherium spp.-Hydrodamalinae, but they are not scored in Halitherium christolii.
The position of Halitherium christolii is problematic, the only character that could shift this poorly known species farther down, to be sister taxon of the Dugonginae + Caribosiren turneri-Metaxytherium spp.-Hydrodamalinae clade is character 36 ri = 0.60, scored as primitive in Halitherium christolii and as derived in Dugonginae and in Caribosiren turneri-Metaxytherium spp.-Hydrodamalinae. But this character is not derived in all the Dugonginae; in fact it is scored as primitive in Bharatisiren kachchhensis, in Dioplotherium allisoni and above all in Crenatosiren olseni. C. 36(0) is regarded as a reversal in Bharatisiren kachchhensis and in Dioplotherium allisoni (see below), but in Crenatosiren olseni, the most primitive dugongine, it could be regarded with the same probability either as primitive or as a reversal. Therefore node 10 remains unresolved, awaiting better knowledge of Halitherium christolii.
Node 11 (Dugonginae):
Homoplastic characters: 31(2) ri = 0.80 c 9 27, 37(2) ri = 0.75 c 24, 43(1) ri = 0.85, 90(1) ri = 0.83.
Node 12 (Dugonginae excluding Crenatosiren olseni):
Homoplastic characters: 42(1) ri = 0.71, 85(2) ri = 0.66 c 7 23, 88(2) ri = 0.83, 136(2) ri = 0.80, 137(1) ri = 0.85, 140(2) ri = 0.64 c 7 26.
Autapomorphies of Dugong dugon:
Homoplastic characters: 13(2) ri = 0.50 c 18 26, 16(1) ri = 0.40 c 6 20, 37(3) ri = 0.75 c 17, 43(0)r ri = 0.85, 66(1) ri = 0.80 c 18 23, 87(1) ri = 0.50 c 24 26, 99(1) ri = 0.71 c 8 25. Domning (1994) also includes: shortened zygomatic-orbital bridge of maxilla [c. 14(1)], strong inflection of processus retroversus of squamosal, constant presence in juveniles of deciduous I1, and frequent presence in adults of vestigial lower incisors; sexual dimorphism in size and in eruption of permanent I1 tusks; functional loss of enamel crowns of cheek teeth; persistently open roots in m/M 2-3. Dugong dugon is a Recent species, considered aberrant, probably neotenic, without a known fossil record (Domning, 1994).
Non homoplastic character: 6(1).
Node 14 (Bharatisiren spp.):
Homoplastic character: 46(1) ri = 0.71 c 18. Autapomorphies of Bharatisiren indica: Non homoplastic character: 125(1).
Homoplastic characters: 42(0)r ri = 0.71 c 16, 83(0)r ri = 0.25 c 9 22.
Characters 83 and 125 are not scored in Bharatisiren kachchhensis, so I can not rule out that they are autapomorphies of the whole taxon Bharatisiren.
Autapomorphies of Bharatisiren kachchhensis:
Homoplastic characters: 20(1) ri = 0.66 c 19, 36(0)r ri = 0.60 c 10? 11? 20; also posterior end of premaxillary symphysis dorsally flattened (c. 10(2) from Bajpai & Domning, 1997), palatal incisure posteriorly constricted, and maxillojugal suture located lateral to anterior part of ridge for origin of masseter muscle (Bajpai & Domning, 1997).
Node 15 (Dugonginae excluding Crenatosiren olseni, Dugong dugon and Bharatisiren spp):
Non homoplastic characters: 6(2), 141(1). Homoplastic character: 76(0)r ri = 0.66.
Node 16 (“Halitherium” bellunense + Corystosiren varguezi + Rytiodus spp.):
Non homoplastic character: 7(1).
Homoplastic character: 86(1) ri = 0-60 c 2 20 24. Autapomorphies of “Halitherium” bellunense:
Homoplastic characters: 42(0)r ri = 0.71 c 14, 90(0)r ri = 0.83, 136(0)r ri = 0.80, 140(1)r ri = 0.64 c 1 3 25. These reversals could represent ontogenetic and not phylogenetic differences, because of the very juvenile age of the only known “Halitherium” bellunense specimen.
Node 17 (Corystosiren varguezi + Rytiodus spp.):
Non homoplastic character: 141(2).
Autapomorphies of Corystosiren varguezi:
Homoplastic characters: 37(3) ri = 0.75 c 12, 67(2) ri = 0.42 c 2 25.
Node 18 (Rytiodus spp.):
Homoplastic characters: 31(1)r ri = 0.80 c 21, 46(1) ri = 0.71 c 14, 66(1) ri = 0.80 c 12 23. Autapomorphies of Rytiodus sp. from Libya:
Homoplastic characters: 9(0)r ri = 0.00, 13(2) ri = 0.50 c 12 26, 88(1)r ri = 0.83. Character 88 is not scored in Rytiodus capgrandi, so I can not rule out that 88(1) is an autapomorphy of the whole taxon Rytiodus.
Character state 9(0) should be considered not as a reversal, but as a derived condition, because the lacrimal intrudes between the posterior extremity of the nasal process of premaxilla and
the anterior extremity of the supraorbital process of frontal, so that the dorsal contact between premaxilla and frontal is lost.
Node 19 (Dioplotherium spp. + Xenosiren yucateca):
Non homoplastic character: 89(1).
Homoplastic characters: 20(1) ri = 0.66 c 14, 102(2) ri = 0.50. Autapomorphy of Dioplotherium manigaulti:
Homoplastic characters: 76(1) ri = 0.66 c 3.
Node 20 (Dioplotherium allisoni + Xenosiren yucateca):
Homoplastic characters: 16(1) ri = 0.40 c 6 12, 86(1) ri = 0.60 c 2 16 24. Autapomorphies of Dioplotherium allisoni:
Non homoplastic character: 125(2).
Homoplastic characters: 36(0)r ri = 0.60 c 10? 11? 14, 123(0)r ri = 0.75 c 8; also posterior end of premaxillary symphysis dorsally flattened (c. 10(2) from Bajpai & Domning, 1997).
Characters 123 and 125 are not scored either in Xenosiren yucateca or in Dioplotherium manigaulti, so I can not rule out that they occur father down at node 19 or 20.
Autapomorphy of Xenosiren yucateca: Homoplastic character: 85(1)r ri = 0.66.
Node 21 (Caribosiren turneri + Metaxytherium spp. + Hydrodamalinae):
Homoplastic characters: 31(1) ri = 0.80 c 18, 140(0)r ri = 0.64.
It is uncertain whether the tusks of Caribosiren turneri were really absent (as autapomorphy) or merely small (Domning, 1994).
Node 22 (Metaxytherium spp. + Hydrodamalinae):
Homoplastic character: 11(1) ri = 0.83 c 6.
Metaxytherium krahuletzi is characterized by the following reversal: 83(0)r ri = 0.25 c 9 14. Moreover M. krahuletzi appears to be primitive among the Metaxytherium species in having a tripartite sternum (Domning & Pervesler, 2001: 30) and in having the following polimorphisms: nasals sometimes united in midline [c. 31(0 or 1)], exoccipitals sometimes meet in midline [c. 66(0 or 1)], and ventral extremity of jugal sometimes under the posterior edge of orbit [c. 85(1 or 2)].
Node 23 Unresolved (Metaxytherium spp. + Hydrodamalinae excluding Metaxytherium
krahuletzi):
Homoplastic characters: 37(1) ri = 0.75, 66(1) ri = 0.80 c 12 18, 85(2) ri = 0.66 c 7 12.
M. medium and M. crataegense appear not to be distinguishable morphologically and therefore a specific distinction appears questionable.
Homoplastic character: 86(1) ri = 0.60 c 2 16 20.
This node is supported only by one character, that is scored only on the holotype of M. arctodites and is polymorphic in M. floridanum.
Metaxytherium floridanum is characterized by a shortened zygomatic-orbital bridge of maxilla (c. 14(1) from Domning, 1994).
Autapomorphies of Metaxytherium arctodites:
Homoplastic characters: 1(0)r ri = 0.57 c 6 26, 37(2) ri = 0.75 c 11, 87(1) ri = 0.50 c 12 26.
Node 25 (Metaxytherium serresii + Metaxytherium subapenninum + Hydrodamalinae):
Homoplastic characters: 67(2) ri = 0.42 c 2 17, 99(1) ri = 0.71 c 8 12, 140(1) ri = 0.64 c 1 3 16.
The body of Metaxytherium serresii is smaller than that of the other Metaxytherium species (see Chapter 6).
Node 26 (Metaxytherium subapenninum + Hydrodamalinae):
Homoplastic characters: 1(0)r ri = 0.57 c 6 24, 37(0)r ri = 0.75, 87(1) ri = 0.50 c 12 24. Autapomorphies of Metaxytherium subapenninum:
Homoplastic characters: 13(2) ri = 0.50 c 12 18, 140(2) ri = 0.64 c 7 12.
Also a tendency to enlargement (broadening) of the nasal process of premaxilla. Moreover the body of M. subapenninum is larger than that of the other Metaxytherium species.
In agreement with Domning (1994), I regard this node as spurious, being not supported by palaeogeographic and chronostratigraphic evidences; and I regard these three changes as evolved in parallel by European Pliocene Metaxytherium subapenninum and Miocene-Recent North Pacific hydrodamalines.
It is crucial to note that two of these three changes characterized Miocene North Pacific Metaxytherium arctodites too: 1(0)r and 87(1). Moreover, Aranda-Manteca et al. (1994) consider this species as the sister taxon of the Hydrodamalinae, since it shares with them a preorbital process of jugal that contacts the premaxilla [c. 87(1)], a reduced lacrimal, and possibly a tendency to thinning of the zygomatic-orbital bridge. In order to test this hypothesis, I reran the analysis excluding M. subapenninum, and I then obtained a tree with the clade Metaxytherium arctodites + Hydrodamalinae supported at 100% and characterized by 1(0)r and 87(1), in accord with palaeogeographic and chronostratigraphic evidences.
Node 27 (Hydrodamalinae):
Non homoplastic character: 70(1).
Homoplastic characters: 67(0)r ri = 0.42 c 9, 91(2) ri = 0.66 c 9, 122(2) ri = 0.87 c 8, 128(0)r ri = 0.90, 139(1) ri = 0.50 c 9.
Homoplastic characters: 31(2) ri = 0.80 c 9 11, 77(0)r ri = 0.66 c 6, 126(0)r ri = 0.50.
In this cladistic analysis D. jordani does not show autapomorphies, anyway Domning (1994: 184) observed that D. jordani bears at least one autapomorphy, separation of palatines in midline, that rules it out of the ancestory of H. gigas.
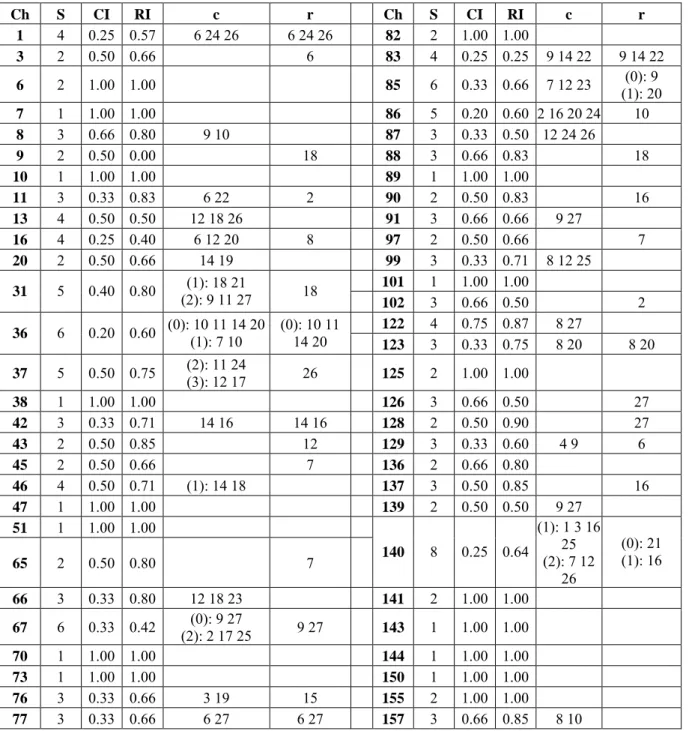
Ch S CI RI c r Ch S CI RI c r 1 4 0.25 0.57 6 24 26 6 24 26 82 2 1.00 1.00 3 2 0.50 0.66 6 83 4 0.25 0.25 9 14 22 9 14 22 6 2 1.00 1.00 85 6 0.33 0.66 7 12 23 (1): 20 (0): 9 7 1 1.00 1.00 86 5 0.20 0.60 2 16 20 24 10 8 3 0.66 0.80 9 10 87 3 0.33 0.50 12 24 26 9 2 0.50 0.00 18 88 3 0.66 0.83 18 10 1 1.00 1.00 89 1 1.00 1.00 11 3 0.33 0.83 6 22 2 90 2 0.50 0.83 16 13 4 0.50 0.50 12 18 26 91 3 0.66 0.66 9 27 16 4 0.25 0.40 6 12 20 8 97 2 0.50 0.66 7 20 2 0.50 0.66 14 19 99 3 0.33 0.71 8 12 25 101 1 1.00 1.00 31 5 0.40 0.80 (2): 9 11 27 (1): 18 21 18 102 3 0.66 0.50 2 122 4 0.75 0.87 8 27 36 6 0.20 0.60 (0): 10 11 14 20 (1): 7 10 (0): 10 1114 20 123 3 0.33 0.75 8 20 8 20 37 5 0.50 0.75 (2): 11 24 (3): 12 17 26 125 2 1.00 1.00 38 1 1.00 1.00 126 3 0.66 0.50 27 42 3 0.33 0.71 14 16 14 16 128 2 0.50 0.90 27 43 2 0.50 0.85 12 129 3 0.33 0.60 4 9 6 45 2 0.50 0.66 7 136 2 0.66 0.80 46 4 0.50 0.71 (1): 14 18 137 3 0.50 0.85 16 47 1 1.00 1.00 139 2 0.50 0.50 9 27 51 1 1.00 1.00 65 2 0.50 0.80 7 140 8 0.25 0.64 (1): 1 3 16 25 (2): 7 12 26 (0): 21 (1): 16 66 3 0.33 0.80 12 18 23 141 2 1.00 1.00 67 6 0.33 0.42 (2): 2 17 25 (0): 9 27 9 27 143 1 1.00 1.00 70 1 1.00 1.00 144 1 1.00 1.00 73 1 1.00 1.00 150 1 1.00 1.00 76 3 0.33 0.66 3 19 15 155 2 1.00 1.00 77 3 0.33 0.66 6 27 6 27 157 3 0.66 0.85 8 10
Table 3: Character fits, convergences and reversals for the tree in figure 1. Ch, character; S, steps; RI, retention index; c and r, nodes where respectively convergences and reversals appear.
5.5 DISCUSSION AND CONCLUSIONS
The results of this cladistic analysis show a close correspondence between palaeobiogeographic and chronostratigraphic distribution of the taxa and their positions on the tree. The only clear exception is represented by node 26 (Metaxytherium subapenninum + Hydrodamalinae), here considered spurious in accord with Domning, 1994 (see discussion above).
Eocene sirenians are not the object of this analysis, nevertheless I have to point out that some autapomorphies scored at node 3 (Halitherium taulannense + Oligocene–Recent Sirenia) were also scored by Domning (1994) in other Eocene sirenian species besides Halitherium taulannense.
H. taulannense is regarded as the oldest representative of the genus Halitherium (Sagne, 2001) and could represent the link between the Eocene and post-Eocene sirenians in this analysis. Concerning the whole Late Eocene–Oligocene genus Halitherium, it appears to be polyphyletic.
Trichechidae appear to be stably rooted well within the Oligocene-Recent Dugongidae. However, the monophyly of this clade is supported by some primitive states, here interpreted as reversals, but which could really be primitive characters and the clade could be rooted farther down in the cladogram.
The position of Halitherium christolii is problematic.
The introducion of more Eocene sirenian species in this cladistic analysis could clarify the systematic position of the Halitherium species, their relationships, and probably change the point of origin of trichechids. Moreover, a better knowledge of H. christolii, especially in comparison with the basal species of Dugonginae and the other Oligocene Halitheriinae, could resolve node 10.
The Dugonginae clade appears well defined in this cladistic analysis. Crenatosiren olseni is confirmed to be the most primitive dugongine. The living species Dugong dugon is confirmed to be the second-earliest member of the dugongines in terms of clade rank, therefore it appears to be a species with no fossil record (see Domning, 1994). Bharatisiren indica and Bharatisiren kachchhensis constitute a dugongine taxon independently evolved in the Indian Ocean. The other dugongine species are characterized by modified and specialized tusks; these dugongines are divided into two branches: one including Dioplotherium spp. and Xenosiren yucateca, and the other including “Halitherium” bellunense, Corystosiren varguezi and Rytiodus spp.
“Halitherium” bellunense is represented only by an incomplete and very juvenile specimen, here revised (see chapter 4). Although it lacks 34 characters, it shows derived characters diagnostic for specialized dugongines, which establish its systematic position as sister taxon of the clade Corystosiren varguezi + Rytiodus spp. Moreover it is characterized by four reversals, which could represent ontogenetic and not phylogenetic differences, because of the very juvenile age of the specimen.
Rytiodus capgrandi and Rytiodus sp. from Libya constitute a clade distict from Corystosiren varguezi. Rytiodus sp. from Libya, here revised (see chapter 4), appears to be a valid species different from Rytiodus capgrandi in having lost the dorsal contact between premaxilla and frontal [c. 9(0) ri = 0.00] and in having a very large infraorbital foramen [c. 13(2) ri = 0.50]. The node 21 representes a taxon including Caribosiren turneri, the paraphyletic genus Metaxytherium and the subfamily Hydrodamalinae, that appears to be the sister taxon of Dugonginae.
Caribosiren olseni is confirmed to be the sister taxon of the Metaxytherium spp. + Hydrodamalinae clade and Metaxytherium krahuletzi is confirmed to be the most primitive species of the paraphyletic genus Metaxytherium in accord with Domning (1994).
Among the Miocene Metaxytherium species M. arctodites appears to be well distinct in having three autapomorphies.
On the contrary, M. medium and M. crataegense appear not to be distinguishable morphologically; and M. floridanum is distinct from them only by having a shortened zygomatic-orbital bridge of maxilla (c. 14(1) from Domning, 1994) and a postorbital process of jugal usually present (13/18 specimens, from Domning, 1988: 407).
A specific distinction among M. medium, M. crataegense, and probably also M. floridanum, seems to be questionable and these species could represent distinct populations of only one species with a wide distribution comparable to that of the modern dugongine Dugong dugon. Metaxytherium floridanum and Metaxytherium arctodites form a clade which appears poorly supported; in fact, the only apomorphy supporting this clade is c. 86(1) ri = 0.60, scored only on the holotype of M. arctodites and scored as polymorphic in M. floridanum.
Node 26 (Metaxytherium subapenninum + Hydrodamalinae) is regarded as spurious, in accord with Domning (1994), being unsupported by palaeogeographic and chronostratigraphic evidences. The European Pliocene Metaxytherium subapenninum and the Miocene-Recent North Pacific hydrodamalines are considered to have evolved in parallel.
The same cladistic analysis, modified to exclude M. subapenninum, shows a tree in which Metaxytherium arctodites is the sister taxon of Hydrodamalinae and the respective node is
supported at 100% by two apomorphies, in agreement with Aranda-Manteca et al. (1994) and palaeogeographic and chronostratigraphic evidences.
Metaxytherium serresii and Metaxytherium subapenninum are well distinct from the other Metaxytherium species in having: a supracondylar fossa of exoccipital located dorsomedial to condyle, palatines not extending anteriorly beyond posterior edge of zygomatic-orbital bridge, and tusks larger with their alveoli extending about half the length of the symphysis or more.