1. INTRODUZIONE
1.1 La Acetil-L-Carnitina (ALC)
La Acetil-L-Carnitina (ALC) (Fig. 1), una molecola naturale ubiquitariamente presente nell’organismo, rappresenta l’estere di L-carnitina a corta catena più abbondante nel cervello (Shug et al., 1982), in particolare a livello dell’ipotalamo (Bresolin et al., 1982).
CH
3H
3C N
+CH
2CH
2CH
2COOH
CH
3O
CO CH
3Figura 1.Struttura dell’acetil-l-carnitina
1.1.1 Aspetti molecolari e biochimici: “il sistema carnitina”
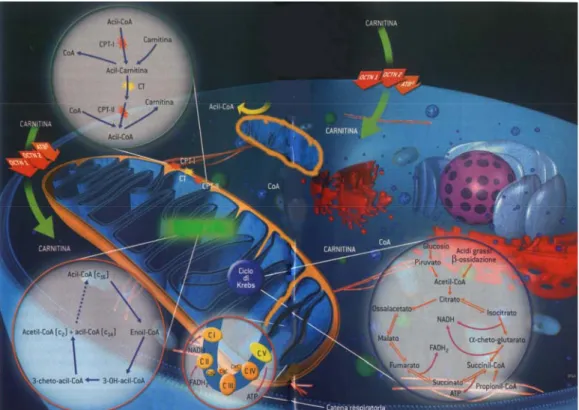
ALC, unitamente alle altre acil-carnitine e alla carnitina libera, fa parte del “sistema della carnitina” (Fig. 2), in cui si trova anche un complesso enzimatico, localizzato a livello della membrana mitocondriale, che comprende: Carnitina Palmitoil Transferasi I-II (CPT I-II),
Carnitina/Acil-carnitina Traslocasi (CT), Carnitina Acetil Transferasi (CAT). ALC si forma dall’acetilazione della carnitina ad opera della CAT, enzima localizzato nella membrana interna del mitocondrio, che catalizza il trasferimento del gruppo acetile dall’Acetil-CoenzimaA (Acetil-CoA) alla carnitina (Edwards et al., 1974); questa reazione è reversibile: l’enzima, infatti, può catalizzare anche la reazione inversa, con conseguente liberazione di Acetil-CoA, composto chiave nelle interazioni metaboliche fra cellule gliali e neuroni (Sonnewald et
al., 1993). ALC può essere trasportata all’interno del mitocondrio per mezzo
della CT, localizzata a livello della membrana mitocondriale interna, mediante un trasporto uniporto (Indiveri et al., 1994).
β-ossidazione
Il sistema della carnitina permette il controllo della concentrazione degli esteri del CoA e la permeabilità del gruppo acile attraverso la membrana mitocondriale interna, impermeabile al CoA e ai suoi derivati (Zammit, 1999). Infatti, la L-carnitina è essenziale per il trasporto degli acidi grassi all’interno del mitocondrio dove subiscono β-ossidazione con conseguente produzione di ATP. Proprio per il ruolo essenziale che riveste, una carenza di L-carnitina comporta un blocco della β-ossidazione (Edwards et al., 1974).
1.1.2 Meccanismi di trasporto
L-carnitina e ALC vengono trasportate attraverso le membrane plasmatiche grazie a trasportatori cationici appartenenti alla superfamiglia di trasportatori di ioni organici (Koepsell, 1998). Due di questi trasportatori sono stati isolati e caratterizzati nel ratto, nel topo e nell’uomo (Wu et al., 1998): OCTN1 (Organic Cation Transporter N1) con bassa affinità per carnitina e ALC, e OCTN2 (Organic Cation Transporter N2), con alta affinità per queste molecole (Tamai et al., 1998). Nel cervello, ALC viene trasportata da OCTN2 attraverso la barriera ematoencefalica (BEE) (Kim et al., 1990), anche se sono in corso studi su altri trasportatori (Mroczkowska et al., 2000) che operano con un meccanismo Na+-dipendente (Kido et al., 2001).
Studi condotti sul macaco Rhesus (Kuratsune et al., 1997) hanno evidenziato che l’ingresso di ALC avviene in maggior quantità a livello della corteccia
cerebrale rispetto alle altre aree cerebrali e che il gruppo acetile si distacca da ALC subito dopo esser entrato nel cervello. L’elevata capacità di ALC di attraversare la BEE sottolinea come questa sostanza possa agire in modo rilevante su aspetti funzionali del sistema nervoso attraverso proprietà di tipo neuroprotettivo e neuromodulatorio.
1.2 Azioni neuroprotettive e anti-aging
Numerosi studi hanno riconosciuto gli effetti benefici di trattamenti con ALC in alcune patologie neurodegenerative la cui comparsa è stata messa in relazione con disfunzioni mitocondriali e con processi a cui l’organismo va incontro naturalmente durante l’invecchiamento (Zbigniew and Ashraf, 2003; Hinerfeld et al., 2004; Liu et al., 2004). E’ stato accertato che ALC riduce la quantità dei radicali liberi e aumenta la disponibilità di energia mitocondriale, ed ha effetti benefici nella stabilizzazione delle membrane (Calvani et al., 1992). Questi effetti neuroprotettivi sono stati rilevati principalmente nell’ippocampo, nella substantia nigra, nello striato e nelle zone cerebrali ricche di recettori muscarinici (Bonavita, 1986; Calvani et al., 1992; Pettegrew et al., 1995). Il fatto che ALC possa in parte proteggere dall’azione dei radicali liberi è stato ulteriormente dimostrato in studi condotti sull’impiego di questa molecola in relazione al morbo di Parkinson (Mazzio
et al., 2003; Steffen et al., 1995; Virmani et al., 2004). ALC, infatti, presenta
un’attività antiossidante primaria, che inibisce la generazione di radicali liberi e contribuisce al termine delle reazioni di propagazione radicalica, e una attività antiossidante secondaria che ripara le catene di grassi polinsaturi (Arduini et al., 1994).
Studi di tipo biomolecolare condotti su ratti giovani trattati cronicamente con ALC hanno evidenziato che tale sostanza modula positivamente la trascrizione di un membro della famiglia delle heat shock protein, la hsp 72 (Traina et al., 2004). Questa famiglia di proteine contribuisce a stabilire uno stato citoprotettivo in molte patologie umane come nelle malattie neurodegenerative. Recentemente, Calabrese e coll. (2005) hanno dimostrato che negli astrociti il trattamento con ALC induce un aumento dell’espressione dell’eme-ossigenasi 1 (heme oxigenase 1) e della hsp 60, membri della famiglia delle heat shock protein. E’, inoltre, emerso che ALC stimola anche l’espressione del fattore di trascrizione Nrf2 nel nucleo delle cellule trattate (Calabrese et al., 2005).
Studi condotti su topi SAMP8 (Senescence-Acceleration-Prone Mice 8) hanno evidenziato che il trattamento cronico con ALC riduce i livelli di perossidazione lipidica (Yasui et al., 2002). L’aumento della perossidazione lipidica è considerato uno degli eventi correlati con l’invecchiamento, così come la diminuzione dell’attività della Na+/K+-ATPasi e la diminuzione della glutatione S-transferasi; a tale aumento è dovuto anche l’irrigidimento delle
membrane plasmatiche e l’accumulo di lipofuscine. In esperimenti condotti su ratti di 24 mesi, si è osservato che la somministrazione di ALC antagonizza alcuni di questi eventi (Kaur et al., 2001; Ghirardi et al., 1988; Aureli et al., 1990; Aureli et al., 1994).
Nei ratti anziani, un decremento dell’attività dell’enzima di membrana Na+/K+-ATPasi in diverse regioni del cervello (corteccia, ippocampo, striato) è associato ad una riduzione dell’efficacia sinaptica (Tanaka and Ando, 1990; Kaur et al., 1998). Il trattamento cronico con ALC determina un aumento dell’attività della Na+/K+-ATPasi in diverse regioni del cervello e ciò contribuisce ad attribuire ad ALC effetti anti-invecchiamento (Kaur et al., 2001). Inoltre, ALC porta all’annullamento delle disfunzioni mitocondriali correlate all’età avanzata e al conseguente aumento della disponibilità di ATP che potrebbe perciò determinare, anch’esso, un aumento dell’attività della Na+/K+-ATPasi (Hagen et al., 1998).
Cambiamenti nella struttura e nella funzione del sistema nervoso centrale sono associati al declino cognitivo proprio dell’invecchiamento. In studi comportamentali effettuati su ratti anziani, la somministrazione cronica di ALC si è dimostrata in grado di migliorare l’apprendimento e la memoria compromessi dall’invecchiamento. Ciò è stato osservato sottoponendo gli animali a differenti tests sperimentali, quali il “Morris water maze” (Caprioli
et al., 1995; Taglialatela et al., 1996), l’“AKON-1 maze” (Ando et al., 2001),
Il trattamento con ALC migliora anche la memoria a lungo termine e la memoria di riferimento spaziale in ratti anziani sottoposti a “split-stem T-maze” (Barnes et al., 1990).
1.2.1 ALC e neurotossicità indotta dal glutammato
ALC ha anche un ruolo protettivo nei confronti della neurotossicità indotta da glutammato: quando tale trasmettitore è in eccesso provoca un malfunzionamento dei recettori NMDA, a cui si devono numerosi disturbi neurodegenerativi. ALC previene questo effetto, in quanto aumenta l’affinità tra il glutammato e i suoi recettori metabotropici (Minana et al., 1996).
Prickaerts e coll. (1995), in uno studio condotto su giovani ratti trattati con strepzotocina (STREP), sostanza che provoca un’alterazione del metabolismo del glucosio, hanno dimostrato che il trattamento cronico con ALC (75mg/Kg/dì) produce un aumento delle vie energetiche alternative. In questo modo si riducono gli effetti da neurodegenerazione indotti da STREP, deficit nella percezione di informazioni spaziali e riduzione della colina acetiltransferasi (AChT) nell’ippocampo. Proprio per questa sua proprietà, ALC è utilizzata nel trattamento delle neuropatie diabetiche, in cui è stata riscontrata una ridotta velocità dell’impulso nervoso nelle fibre nervose periferiche. Studi condotti su animali in cui era stato sperimentalmente indotto il diabete, hanno dimostrato che la somministrazione di ALC aumenta
la velocità di conduzione dell’impulso nervoso (Merry et al., 1995; Lowitt et
al., 1995).
Da tutte queste considerazioni emerge il perché ALC abbia applicazioni terapeutiche in un ampio spettro di patologie.
1.3 Effetti analgesici
Il fatto che ALC sia in grado di modulare l’attività dei recettori è stato dimostrato anche da studi condotti su ratti anziani con neuropatie dolorose croniche. Uno dei meccanismi comunemente riconosciuto alla base del dolore cronico è un aumento della sensibilità dei neuroni sensoriali delle corna dorsali (Sandkuhler, J., 2000). In modelli animali con neuropatie dolorose, si è dimostrato che tale aumento è dovuto al rilascio di elevate quantità di glutammato, con conseguente potenziamento a lungo termine della sinapsi tra la fibra afferente e il secondo neurone delle corna dorsali (Sandkuhler, J., 2000). E’ anche stata individuata la causa di questo potenziamento in una attivazione simultanea dei recettori metabotropici e N-Metil-D-Aspartato (NMDA) del glutammato. In particolare, a questo riguardo, è stato dimostrato che ALC stimola l’up-regulation dei recettori metabotropici del glutammato di tipo 2 (mGlu2R) nelle corna dorsali del midollo e nella corteccia
prefrontale, inducendo analgesia (Chiechio et al., 2002). ALC è, perciò, un composto utilizzato nel trattamento di neuropatie del dolore per le sue
proprietà analgesiche che si esplicano in casi di dolore cronico ed acuto (Chiechio et al., 2002).
In uno studio recente, condotto in parallelo su un gruppo di pazienti con neuropatia diabetica e un gruppo di controllo, è stato dimostrato che ALC è efficace nel ridurre i sintomi e nell’alleviare il dolore di tale patologia, e nel migliorare la rigenerazione delle fibre nervose (Sima et al., 2005).
1.4 Azioni neuromodulatorie
ALC ha anche un ruolo protettivo nei confronti della neurotossicità indotta da glutammato: quando tale neurotrasmettitore è in eccesso provoca un malfunzionamento dei recettori NMDA a cui si devono numerosi disturbi neurodegenerativi (Minana et al., 1996). ALC previene questo effetto, in quanto aumenta l’affinità tra il glutammato e i suoi recettori metabotropici (Minana et al., 1996).
ALC svolge molti ruoli modulatori. Le proprietà biologiche di ALC sono state spesso studiate in relazione a funzioni cerebrali, come apprendimento, attenzione e memoria e se ne è ricercata l’influenza soprattutto sul sistema colinergico e glutammatergico.
Esperimenti effettuati in vitro su sinaptosomi isolati da ratti trattati cronicamente con ALC hanno evidenziato un aumento dell’uptake di colina operato da ALC (Curti et al., 1989). Esperimenti di microdialisi cerebrale
condotti su ratti svegli e in grado di muoversi, hanno dimostrato che ALC stimola il rilascio di acetilcolina in alcune regioni del cervello, come l’ippocampo e lo striato (Imperato et al., 1989).
ALC, inoltre, gioca un ruolo neuromodulatorio sul trofismo neuronale durante le prime fasi dell’embriogenesi del sistema nervoso (Kentroti et al., 1992).
1.5 Effetti su apprendimento e memoria
1.5.1 Effetti sulla long-term-potentiation (LTP) e sui recettori
glutammatergici.
Un meccanismo molto studiato è il fenomeno sinaptico chiamato potenziamento a lungo termine (LTP), un persistente incremento nella forza sinaptica, come risulta dalla misurazione dell’ampiezza del potenziale postsinaptico eccitatorio (EPSP), che può essere rapidamente indotto da una breve attività neuronale. Questa forma di plasticità sinaptica è implicata nel processo di memoria in parecchie regioni del cervello ma gli studi si sono principalmente soffermati sull’ippocampo. All’interno dell’ippocampo, la sinapsi più studiata è quella dell’input della collaterale/comminsurale di Schaffer ai neuroni piramidali della regione CA1. L’LTP si può descrivere come un potenziamento delle risposte sinaptiche eccitatorie registrate nel campo CA1 dell’ippocampo, in seguito a stimolazione tetanica (treni di 100
stimolazioni a 100 Hz) delle vie afferenti di Schaffer. Se somministriamo ALC 30 minuti prima della stimolazione tetanica, a concentrazioni comprese tra 10 μM e 1 mM, in preparati ippocampali di ratti anziani si determina un aumento della trasmissione sinaptica basale del 20% e dell’ LTP di circa il 150%. Questi dati ci suggeriscono che il trattamento con ALC è in grado di aumentare la trasmissione sinaptica eccitatoria in ratti anziani (Castorina et
al., 1993).
Il meccanismo di azione dell’ALC sulla trasmissione sinaptica sembra coinvolgere i recettori del glutammato NMDA e i recettori muscarinici. Infatti, l’aggiunta di antagonisti dei recettori del glutammato di tipo NMDA riducono significativamente l’aumento del EPSP indotto dall’ALC (Castorina
et al., 1993).
La progressiva riduzione età-dipendente del numero dei recettori di tipo NMDA è correlata con la comparsa e lo sviluppo di disturbi della memoria associati all’invecchiamento e ad alcune condizioni patologiche come la demenza. In particolare, si assiste ad una riduzione del 19.5% dei recettori NMDA nell’ippocampo di ratti anziani. Il trattamento per sei mesi con ALC (75 mg/kg/dì) a ratti anziani attenua la perdita neuronale dei recettori NMDA a livello dell’ippocampo. Mentre una singola dose di trattamento con ALC a ratti anziani riduce di poco la perdita neuronale e non ha effetti su ratti giovani. Questi risultati suggeriscono che l’ALC possa esercitare un’azione
trofica e neuroprotettiva quando è somministrata cronicamente e un’azione stimolatoria quando è somministrata a singole dosi(Castorina et al., 1994).
1.5.2
Effetti sull’apprendimento negli invertebrati
Gli invertebrati sono utili modelli per analizzare gli eventi cellulari e molecolari che riguardano l’apprendimento. Il sistema nervoso di molti invertebrati contiene pochi neuroni ben identificabili, nei quali è possibile studiare in parallelo le attività neuronali alla base di semplici atti comportamentali. E’ così possibile studiare anche i meccanismi alla base di forme elementari di apprendimento non associativo (disabitudine e sensitizzazione) e apprendimento associativo (condizionamento Pavloviano e condizionamento operante). Studi comportamentali sull’invertebrato Hirudo
medicinalis hanno dimostrato che una singola somministrazione di ALC
induce cambiamenti comportamentali, in particolare blocca la sensitizzazione e riduce la disabitudine indotte da stimolazione nocicettiva (brushing) nell’induzione al nuoto, dopo 2 ore dalla somministrazione, e che questi effetti permangono per oltre 11 giorni (Ristori, et al., 2004).
Studi elettrofisiologici hanno analizzato l’effetto dell’ALC sui neuroni di H.
medicinalis. Questi neuroni sono localizzati all’interno dei gangli che
costituiscono il sistema nervoso dell’invertebrato. In tali cellule la stimolazione dei campi afferenti o stimolazioni intracellulari evocano una
scarica di potenziali d’azione, seguita da una iperpolarizzazione postuma (AHP). AHP è un parametro elettrofisiologico caratteristico di molte cellule nervose sia di vertebrato che di invertebrato.
Nei neuroni tattili T di H. medicinalis, l’AHP è sostenuta dall’attività della
pompa Na+,K+/ATPasi, e in modo minore, per circa un 20-25%,
dall’attivazione di una conduttanza K+/Ca2+-dipendente (gK/Ca) (Jansen and
Nicholls, 1973; Van Essen, 1973). La ripetuta stimolazione dei neuroni T determina un incremento dell’ampiezza dell’AHP che è sostenuto da un potenziamento dell’attività della Na+,K+/ATPasi. L’incremento dell’ampiezza dell’AHP porta ad una durevole depressione delle connessioni sinaptiche tra le cellule T e i neuroni postsinaptici (Scuri et al., 2002). Ciò ha suggerito che questo fenomeno possa essere coinvolto nella plasticità a breve termine associato a processi di apprendimento non associativo (Zaccardi et al., 2001, 2004). Il trattamento con ALC 2mM determina un prolungato aumento dell’ampiezza dell’AHP, suggerendo un possibile ruolo della pompa elettrogenica Na+,K+/ATPasi nella modulazione della trasmissione sinaptica. Infatti in presenza di apamina, un antagonista del canale K+/Ca2+-dipendente (gK/Ca) (Mozzachiodi et al., 2001), l’ALC è ancora in grado di produrre un
aumento dell’ampiezza dell’AHP suggerendo che ALC possa modulare positivamente e in modo sostenuto l’attività della Na+,K+/ATPasi (Scuri et al., 2002). Tali effetti si registrano anche dopo 11 giorni dal trattamento (Lombardo et al., 2004).
1.6. Regolazione dell’espressione genica
E’ stato ipotizzato che l’ALC possa regolare l’espressione genica. Studi condotti, sia su ratti adulti che anziani, hanno dimostrato che la somministrazione di ALC causa un innalzamento dei trascritti mitocondriali che codificano per la subunità I della citocromo c ossidasi, sia a livello cardiaco che a livello del tessuto cerebrale (Gadaleta et al., 1990, Traina et
al., 2004). L’ALC ha scarsi effetti se questo tipo di somministrazione viene
effettuata su ratti giovani (Gadaleta et al., 1998). La sintesi di proteine mitocondriali così come la trascrizione del DNA mitocondriale (mtDNA) sono regolate da proteine e fattori di trascrizione codificati da geni nucleari (Attardi and Schatz, 1988). In alcune linee cellulari l’ALC potrebbe modulare l’espressione genica, tramite l’acetilazione dell’istone H4. Due fattori di
trascrizione, NRF-1 e NRF-2 possono riconoscere sequenze presenti in geni codificanti per importanti proteine mitocondriali (Gugneja et al., 1996). Alla luce di questi risultati è stato ipotizzato che l’ALC aumenti l’acetilazione dell’istone H4 che a sua volta modula l’azione dei fattori di trascrizione
NRF-1 e NRF-2. Inoltre l’acetilazione dell’istone H4 potrebbe regolare la sintesi
della cardiolipina, un importante componente della membrana mitocondriale (Pisano et al., 1996). La sintesi di cardiolipina è regolata dall’azione di ormoni tiroidei (Hostetler, 1991) e studi condotti su ratti ipotiroidei hanno mostrato che la somministrazione di ALC determina un innalzamento del
mtRNA per la subunità I della citocromo c ossidasi a livello epatico (Mawal
et al., 1998; Rao et al., 1997).
Dunque, la struttura e la funzione della permeabilità della membrana mitocondriale, che potrebbe essere preservata dall’azione dell’ALC, sembra avere un importante ruolo sulla trascrizione del mtDNA poiché influenza il trasporto di alcuni metaboliti e proteine richieste per la sintesi dell’mtRNA (Fisher et al., 1987).
Gli effetti dell’ALC sulla regolazione dell’espressione genica, a livello cerebrale, è stata recentemente valutata in ratti trattati cronicamente con questa molecola (Traina et al., 2004, 2006). La costruzione di due librerie sottrattive di cDNA, utilizzando la tecnica della ibridazione sottrattiva soppressiva (SSH), ha permesso di identificare geni differenzialmente espressi nei due campioni. Questa analisi ha fornito una serie interessante di risultati, che suggeriscono possibili meccanismi molecolari responsabili delle proprietà neuroprotettive e neuromodulatorie dell’ALC. In particolare è emerso che l’ALC modula positivamente la trascrizione del gene che codifica per la heat shock protein 72 (Hsp-72), suggerendo che probabilmente l’ALC è in grado di potenziare i meccanismi di protezione cellulare anche in assenza di stress particolari rendendo la cellula più resistente agli insulti di vario tipo. E’, inoltre, emerso che l’ALC modula positivamente la trascrizione per la isoforma γ della proteina 14-3-3, mentre modula negativamente la trascrizione
del gene per la l’ATP synthase lipid-binding protein P3 precursor, un componente della ATPasi mitocondriale (Traina et al., 2004).
Studi molto recenti hanno ipotizzato che ALC è anche coinvolta nell’omeostasi del calcio e in fenomeni di plasticità sinaptica, suggerendo, come possibile meccanismo molecolare, una modulazione positiva del canale anionico voltaggio-dipendente mitocondriale, VDCA1 (Traina et al., 2006).