CAPITOLO IV
VITAMINA B12
4.1 CENNI STORICI
Nel 1926, due medici di Harvard, George Minot e William Murphy, durante gli studi sull’anemia perniciosa, scoprirono che i sintomi della malattia potevano essere alleviati nutrendo i pazienti con grandi quantità di fegato crudo. Il materiale attivo nel fegato, che fu chiamato vitamina B12,
era presente in quantità molto basse e passarono molti anni prima che fosse possibile isolarne abbastanza per caratterizzare la molecola.
Nel 1964, in Inghilterra, Dorothy Hodgkin e i suoi collaboratori usarono la cristallografia ai raggi X e riuscirono a completare la determinazione della struttura di questa sostanza attiva: per questo lavoro la Hodgkin fu insignita del Premio Nobel.
4.2 BIOCHIMICA
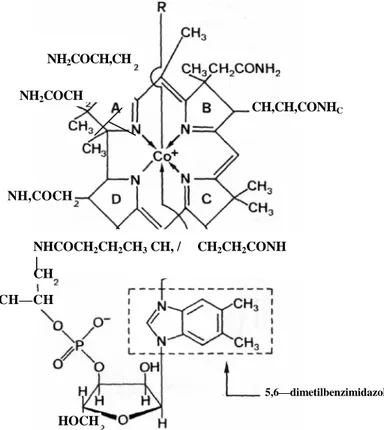
Per vitamina B12 si intende un gruppo di sostanze chimicamente
affini (cobalamine), la cui struttura è costituita da un gruppo planare (anello corrinico) che si lega ad un nucleotide (5,6-dimetilbenzimidazolo legato a ribofosfato) disposto inferiormente e quasi perpendicolarmente ad esso (Figura 3).
5,6—dimetilbenzimidazolo NH2COCH,CH NH2COCH CH,CH,CONHC NH,COCH NHCOCH2CH2CH3 CH, / CH2CH2CONH CH CH—CH HOCH
Figura 3. Struttura della Vitamina B12
L'anello corrinico è simile a quello delle porfirine ed è formato da 4 anelli pirrolici che legano centralmente un atomo di cobalto (Co).
Il Co può legare covalentemente, al di sopra dell’anello corrinico, diversi radicali, a seconda dei quali prende nome il particolare composto cobalaminico:
¾ -CN (cianocobalamina); ¾ -OH (idrossicobalamina); ¾ -CH3 (metilcobalamina);
¾ 5-desossi-5'-adenosina (adenosilcobalamina);
L'adenosilcobalamina, scoperta da H.A.Parker, è la principale forma della vitamina B12 in natura; una seconda forma enzimaticamente attiva è la
metilcobalamina o metil-B12. L'esposizione alla luce converte rapidamente
l'idrossicobalamina in adenosil e metilcobalamina, che sono i coenzimi fisiologici. La cianocobalamina è la forma più stabile e perciò è quella prodotta commercialmente dalla fermentazione batterica ed è solubile in acqua e stabile al calore.
Nell'uomo vi sono solo due reazioni enzimatiche (Ludwige Matthews, 1997) in cui la cobalamina entra come coenzima:
¾ La metilazione dell'omocisteina a metionina, che avviene nel citoplasma ed utilizza la metilcobalamina come coenzima ed il N5 -metil-tetraidrofolato come sorgente di metili.
La metionina sintetasi lega la cobalamina e l’N5-metiltetraidrofolato, trasferendo il gruppo metilico al gruppo prostetico cobalaminico.
Il gruppo metilico è quindi trasferito all’omocisteina, generando metionina; l'assenza di cobalamina esercita un blocco in questa reazione ed un accumulo di N5-metiltetraidrofolato, cofattori necessari per numerose reazioni biochimiche, che comportano il trasferimento di un gruppo monocarbonioso.
In particolare il depauperamento di tetraidrofolato previene la sintesi di adeguate quantità di deossitimidilato (dTMP) e delle purine necessarie per la sintesi del DNA nelle cellule in rapida divisione. L'accumulo del folato come
N5 metiltetraidrofolato, ed il depauperamento associato dei tetraidrofolati cofattori nella carenza di vitamina B12 sono stati indicati come "trappola del
metil-folato": questo è il passaggio biochimico che collega il metabolismo della vitamina BI2 con quello dell'acido folico (Jacques et al., 1999).
L'acido folico può essere ridotto a diidrofolato dall'enzima diidrofolato reduttasi e servire, perciò, come fonte di tetraidrofolati necessari per la sintesi di purine e dTMP, essenziali per la sintesi di DNA.
¾ La seconda reazione enzimatica che utilizza la cobalamina è l'isomerizzazione dell’ L-metilmalonil-CoA a succinil-CoA, mediante
l'enzima L-metilmalonil-CoA mutasi ed il coenzima 5'-deossiadenosilcobalamina.
La carenza di questo fattore provoca l’accumulo nei tessuti di metilmalonil-CoA e del suo precursore, il propionil-CoA; ne conseguono la sintesi e l’incorporazione neuronale di acidi grassi non fisiologici, contenenti un numero anomalo di carboni (Metz, 1992) ed anche questa anomalia biochimica, potrebbe contribuire alle complicanze neurologiche del deficit di cobalamina.
4.3 Aspetti Fisiopatologici
La vitamina B12 naturale è sintetizzata solo da alcuni microrganismi.
L’uomo la ottiene dai cibi: carne, fegato, pesce, uova, latte e prodotti caseari. Il fabbisogno giornaliero minimo di vitamina BB12 è di circa 2,5 μg;
le riserve di un uomo adulto normale si aggirano sui 5 mg ed il fegato ne contiene 1 μg/g.
La maggior parte della vitamina B12 si trova nei mitocondri come
adenosilcobalamina, mentre la metilcobalamina è la forma predominante nel sangue.
La perdita obbligatoria giornaliera è pari ad 1/1000 della riserva corporea e si riduce proporzionatamente al ridursi delle riserve; per cui occorrono da due a quattro anni prima che la mancanza completa di assorbimento della vitamina (per esempio per gastrectomia totale), si manifesti come carenza.
Nell'uomo esistono due meccanismi di assorbimento intestinale: uno attivo e molto efficiente, l'altro passivo ed a bassa efficienza.
Durante la digestione, a livello gastrico, la cobalamina dei cibi viene rilasciata e forma un complesso stabile con il vettore gastrico R, glicoproteina di significato incerto che si ritrova nelle secrezioni (saliva, latte, secrezioni gastriche, bile), nei fagociti e nel plasma. Penetrando nel
duodeno, il complesso cobalamina-proteina vettore R viene digerito, con liberazione della cobalamina che si lega al fattore intrinseco. Questa glicoproteina, di peso molecolare di 50 Kda, è prodotta dalle cellule parietali dello stomaco.
La secrezione del fattore intrinseco è generalmente proporzionale a quella dell’acido cloridrico. Il complesso cobalamina-fattore intrinseco è resistente alla digestione proteolica e può così raggiungere l'ileo termininale, dove recettori specifici, presenti sull'orletto a spazzola della mucosa, lo legano rendendo possibile l'assorbimento della vitamina. Così il fattore intrinseco, come del resto la transferrina, svolge il ruolo di vettore proteico verso le cellule. Il complesso cobalamina-fattore intrinseco, legato al recettore, si porta all’interno delle cellule della mucosa ileale dove, nel giro di alcune ore, il fattore intrinseco è distrutto e la cobalamina viene trasferita a un’altra proteina di trasporto, la transcobalamina II (TCII). Il complesso con la transcobalamina raggiunge i tessuti, si lega a specifici recettori di membrana ed entra nelle cellule mediante endocitosi. Nella frazione citoplasmatica solubile, la vitamina è liberata come idrossicobalamina e, o è convertita a metilcobalamina o entra nei mitocondri, dove il cobalto è ridotto e si forma 5’deossiadenosilcobalamina.