PARTE PRIMA
Introduzione
“Ieri abbiamo ascoltato il re delle scoregge, sapeva di dolce come il miele sulle tartine…!”
[lettera di W. A. Mozart alla madre]
Le lettere di Mozart alla madre, alla cugina e all’amata Costanze erano piene di giochi di parole, termini osceni e scurrili che rompevano la lettera come certi passaggi delle sue fughe rompono le regole musicali. Alla coprografia si aggiungeva un comportamento bizzarro e ossessivo; scriveva di lui Paoline Pichler: “i suoi rapporti
sociali non erano contraddistinti da alcun sintomo di insolita potenza intellettuale o culturale, né di erudizione o altri interessi di alto livello. Un’attitudine mentale piuttosto ordinaria, scherzi infantili e ……un modo di vita irresponsabile erano tutto ciò che lo distingueva in società. Ciononostante, quali profondità, quali mondi di fantasia, armonia, melodia e sentimento si celavano dietro questa esteriorità poco promettente!”
Molto probabilmente, se Mozart fosse vissuto oggi, gli sarebbe stata diagnosticata la patologia dei tic multipli o sindrome di Gilles de la Tourette con depressione, deficit ADHD e idee psicotiche.
Un altro storico esempio di sindrome di Tourette è Samuel Johnson, l’autore del primo dizionario inglese. Molti di coloro che lo incontravano rimanevano sorpresi dai suoi movimenti bruschi e dai frammenti di parole che era solito produrre nel bel mezzo di un discorso. Le “involontarie cattive abitudini”, come Johnson definiva i suoi comportamenti, non si limitavano a suoni o movimenti bizzarri: la vita dell’autore inglese era intrisa di strani rituali, come passare inginocchiato attraverso le porte, e di periodi di profonda depressione. I medici e studiosi del tempo avranno sicuramente pensato che Johnson fosse affetto da una corea reumatica o da chissà quale disturbo psicologico. Oggi quei singolari comportamenti sarebbero probabilmente interpretati come sintomi della sindrome di Tourette.
I La sindrome di Gilles de la Tourette
I.1 La storia
La prima descrizione di un paziente affetto da sindrome di Tourette si trova nei diari del medico francese Itard (1775-1838) e riguarda il caso della giovane marchesa de Dampierre, nota nell’ambiente dell’alta società parigina per il comportamento bizzarro e le improvvise esplosioni di parole oscene ed inappropriate per la sua condizione sociale. La nobildonna manifestava movimenti convulsivi dall’età di sette anni. Durante la pubertà aveva sviluppato anche strane e bizzarre vocalizzazioni che successivamente, per un periodo di 18-20 mesi, si erano interrotte, per ricomparire poco dopo il suo matrimonio accompagnate da coprolalia che ha persistito fino alla morte a 85 anni.
Sessant’anni più tardi un allievo del neurologo francese Charcot, Georges Gilles de la Tourette, dopo aver analizzato nove soggetti affetti dallo stesso e strano male della marchesa de Dampierre, fornì la prima descrizione della sindrome come un’entità distinta dall’isteria e dalla corea. Prima di giungere a tale conclusione, il giovane medico francese aveva letto, oltre al caso della marchesa de Dampierre, tre articoli che poi si rivelarono molto importanti per la caratterizzazione della malattia dei tic multipli. Nel primo articolo, del neurologo Beard, erano descritti alcuni individui maschi affetti dalla sindrome del Jumping Frenchmen of Maine che producevano strani movimenti e rumori anche su imitazione. Beard concludeva l’analisi affermando che questo tipo di disturbo era più frequente nei maschi, era ereditario ed esordiva nell’infanzia.
La sindrome di Gilles de la Tourette (TS) è una malattia neurologica ereditaria caratterizzata da movimenti stereotipati e involontari e da emissioni sonore incontrollabili.
I primi sintomi appaiono generalmente durante l’adolescenza o nell’infanzia, e possono subire una riduzione prima dell’età adulta.
Il sintomo d’esordio più comune è un tic del viso, ma col tempo ne possono apparire altri (spasmo del capo, simile ad uno strappo, lo stiramento del collo, il battere i piedi, piegare il corpo, ecc.). I pazienti con TS presentano anche tic sonori: vocalizzazioni, grugniti, grida, parole, coprolalia o ecolalia. Queste manifestazioni possono variare d’intensità, di durata e frequenza, di caratteristiche e modalità espressive nel corso degli anni. La sindrome di Tourette si accompagna anche ad una serie di disturbi affini più o meno gravi, come l’OCD e l’ADHD.
L’eziopatogenesi è sicuramente genetica, ereditata come anomalia di un gene con tratto autosomico dominante. L’alterazione del gene determina anormale metabolismo della dopamina. Gli studi di neuroimmagine hanno messo in luce anche una riduzione volumetrica dei gangli della base.
Nel secondo articolo, del neurologo O’Brien, si descriveva il Latah1 osservato in Malesia. Secondo O’Brien il latah è caratterizzato da bruschi movimenti, da imitazioni di suoni, ad esempio il latrato di un cane, e da un linguaggio osceno.
Nel terzo articolo, di uno studioso di queste problematiche, Hammond, si descriveva un comportamento simile a quello dei soggetti affetti da Jumping Frenchmen of
Maine e osservato in Siberia nella regione dello Yakutsk, detto myriachit2. Le persone affette da myriachit, però, non emettevano strani suoni o frasi.
Nella descrizione di Gilles de la Tourette la sindrome si articolava in tre fasi: la prima caratterizzata da tic facciali, in particolare dall’ammiccamento, la seconda da comparsa di tic sonori, che possono variare da piccoli rumori a forme di tic sonoro più complesse come l’ecolalia; la terza fase, da coprolalia.
Nel secolo successivo si consolidò la convinzione che la sindrome di Tourette fosse causata da una lesione cerebrale simile a quella che si ha nell’encefalite letargica. Comunque, nell’età della psicoanalisi freudiana, prese piede l’idea che il tic fosse un’ossessione e, dunque, curabile con terapia psicoanalitica. Il problema maggiore di queste teorie stava nel fatto che si basavano su un campione di pazienti molto ristretto: di solito uno, o al massimo, due casi e, per lo più, analizzati solo per supportare le teorie precedentemente esposte.
Negli anni ’70 del 900, negli Stati Uniti si abbandonò la psicoanalisi e ci si rivolse ad una cura farmacologica basata essenzialmente sull’aloperidolo3. Alla luce dei risultati positivi, da quel momento si impose l’idea della sindrome di Tourette non più come problema emozionale o psicologico, ma come disturbo neurologico.
I.2 Caratteristiche cliniche
La sindrome è principalmente caratterizzata da tic motori e sonori, semplici e complessi, che possono manifestare cambiamenti di sede, di frequenza e di intensità e che si alternano con periodi di relativa quiescenza. I tic motori, solitamente, iniziano nella fascia di età fra i 3 e gli 8 anni.
Molti pazienti sperimentano anche alterazioni sensoriali che, di solito, precedono la manifestazione ticcosa motoria: il cosiddetto premonitory urge; affermano di sentirsi
1
In malese latah significa “imprevedibile”. Il termine latah fu usato per la prima volta nel manoscritto Hykayat Koris del XV secolo per descrivere la sindrome sussultoria che colpiva una regina malese.
2
Myriachit nel dialetto locale vuol dire “comportarsi da stupido”.
3
obbligati a compiere un tic dalle sensazioni corporee che si manifestano nell’area anatomica che sarà interessata dal tic. Questi impulsi creano una sensazione di frustrazione perché il soggetto cerca continuamente di trattenerli.
Oltre ai tic, nei soggetti affetti da sindrome di Tourette si manifestano anche il disturbo ossessivo-compulsivo (OCD) e il disturbo dell’attenzione e dell’iperattività (ADHD).
Si distinguono tre tipi di sindrome tourettiana: pura in cui si manifestano tic motori e sonori, full blown con presenza anche di coprolalia, e plus in cui compaiono, oltre ai tic, anche il disturbo ossessivo-compulsivo e il disturbo dell’attenzione e dell’iperattività.
I dati epidemiologici del Nord America e dell’Europa hanno evidenziato che la sindrome è più comune nei maschi, con un rapporto di tre a uno. L’età media d’esordio dei sintomi è sette anni, con un range che va dai due ai quindici anni.
I sintomi iniziali possono essere diversi, ma, per lo più, si manifesta un singolo tic o tic multipli localizzati nel viso e nella testa, che si diffondono poi alle spalle e al tronco. I tic sonori raramente rappresentano il sintomo d’esordio, ma, generalmente, si manifestano dopo i tic motori, attorno agli undici anni. I tic sonori complessi, come la coprolalia e l’ecolalia, sono presenti come sintomo iniziale solo nel 10% dei casi.
Il disturbo dell’attenzione e dell’iperattività e quello ossessivo-compulsivo hanno un esordio tardivo e, secondo alcuni, come conseguenza dello stress causato dai tic.
Gli studi interculturali condotti negli anni ’90 (Staley et al., 1997:6-16) hanno evidenziato le diversità di manifestazione della sindrome di Tourette: i soggetti coreani, ad esempio, manifestano rari sintomi di disturbo ossessivo-compulsivo rispetto ai soggetti europei e nordamericani. In Giappone la coprolalia è un sintomo molto raro: ciò è interpretato come effetto della specificità culturale nell’espressione delle emozioni. Una tourettiana di undici anni di Shangai presentava un tic sonoro complesso con la seguente vocalizzazione: “Abbasso Chou-En-lai”4. E’ pertanto legittimo l’interesse di alcuni studiosi (Earls, 1992:55-59) per i fattori culturali e psicosociali dei tourettiani.
Va, tuttavia, ribadito che le altre caratteristiche cliniche, come la sede dei primi tic motori e la manifestazione dei tic sonori semplici, mostrano un alto grado di similarità interculturale.
4
I.3 Diagnosi
Il manuale dell’associazione psichiatrica americana (DSM IV) riconosce tre tipi di patologie ticcose: la sindrome di Tourette, il disturbo cronico da tic, il disturbo transitorio da tic. I criteri diagnostici sono così schematizzabili:
Sindrome di Tourette
- co-presenza in determinati periodi di tic motori e sonori semplici e/o complessi;
- evoluzione fluttuante, con periodi di esacerbazione e remissione non superiori a tre mesi;
- presenza di tic per almeno un anno;
- esordio della malattia prima del diciottesimo anno d’età.
Disturbo cronico da tic
- presenza di tic o solo motori o solo sonori;
- evoluzione piuttosto stabile sia per l’intensità del tic sia per il tempo di manifestazione;
- presenza di tic per un periodo superiore ad un anno; - esordio della malattia prima del diciottesimo anno d’età;
Disturbo transitorio da tic
- presenza di tic motori e/o sonori semplici o complessi; - durata della malattia non superiore ad un anno;
- esordio della malattia prima del diciottesimo anno d’età.
La diagnosi di sindrome di Tourette richiede la combinazione di tic motori multipli e di uno o più tic sonori. Sintomi come l’ecolalia e la coprolalia non sono, tuttavia, necessari per la diagnosi. Ciò che principalmente caratterizza la sindrome è la variazione di frequenza, di intensità, di localizzazione e di forma dei tic, infatti, come ricorda Porta:
“(Questi) tic motori possono inoltre mutare nella loro espressione motoria, oppure ‘migrare’ immutati da un segmento del corpo a un altro. La loro caratteristica è quella di costituire un movimento per lo più caricaturale eseguito in maniera veloce. Si tratta della goffa copia di un movimento che parrebbe avere una sua finalità, di fatto è ingiustificato”
(Porta, 1996:25) e ancora: “i tic migrano in molte parti del corpo, potendo completamente
sparire dal distretto prima coinvolto” (Porta, 1996:46).
Molti studi (Leckman et al., 1998:14-19; Pappert et al., 2003:936-940) suggeriscono che il tic diminuisce durante la tarda adolescenza; Leckman e colleghi, tramite un modello statistico matematico, hanno constatato che il grado di severità del tic aumenta fra gli 8 e i 12 anni, mentre diminuisce negli anni successivi.
Solitamente, assieme ai tic si presentano anche altri disturbi del movimento come l’ecoprassia, riscontrata anche nel Jumping Frenchmen of Maine, nel latah e nel myriachit.
I.4 Sintomi correlati
Un soggetto affetto da sindrome di Tourette, come si è detto, può presentare, oltre ai tic, anche sintomi psicopatologici, come il disturbo ossessivo-compulsivo e il disturbo dell’attenzione e dell’iperattività. Secondo Shapiro et al. (1998), nel 25-35% dei bambini affetti da sindrome di Tourette sono presenti anche il disturbo ossessivo-compulsivo e quello ADHD: nella maggior parte dei casi, questi sintomi costituiscono un deficit più grave del tic, specialmente per quanto riguarda il rendimento scolastico.
Il disturbo ossessivo-compulsivo è caratterizzato da pensieri o comportamenti frequenti e prolungati che determinano un grave stato d’ansia. Un esempio di pensiero ossessivo può essere la paura di fare del male a qualcuno o la paura di essere contaminati da germi e batteri. Le compulsioni sono, invece, atti che il malato si sente costretto a compiere per rimuovere momentaneamente lo stato di stress. Molti tourettiani affetti da OCD riferiscono che i loro atti compulsivi sono quasi del tutto spontanei e non sono necessariamente effettuati in risposta ad una ossessione, perciò non producono e non sono prodotti da ansia o stress. Secondo Singer (2005:149-159) è possibile distinguere nei tourettiani due tipi di disturbo ossessivo-compulsivo: quello giovanile, legato a comportamenti aggressivi ed impulsivi, e quello adulto, caratterizzato da atti autolesivi.
Il disturbo dell’attenzione e dell’iperattività (ADHD) è una sindrome complessa i cui sintomi primari sono l’iperattività, l’impulsività e la difficoltà a mantenere l’attenzione. Il suo esordio è intorno ai 4-5 anni e precede l’inizio dei sintomi ticcosi di circa due anni. Sono state molte le teorie che hanno cercato di dare una spiegazione a questo disturbo: nel 1947 Strauss e Lehtinen ipotizzarono che i bambini affetti da ADHD fossero danneggiati nella capacità di progettare e svolgere un determinato compito. Distrazione e disattenzione sarebbero la reazione ad uno stimolo superiore alla capacità di concentrazione, generando
così un comportamento disorganizzato, risultato di un sistema cognitivo che non “sopporta carichi eccessivi”.
Negli anni ’80, per spiegare l’ADHD, Tim Shallice (1988) propose il modello
information processing, che prevede quattro unità: unità cognitive, gli schemi, l’inventario
dei contenuti (contention scheduling) e il SAS (supervisory attentional system). Le unità cognitive sono funzioni correlate a sistemi specifici, come ad esempio quello visivo, mentre gli schemi sono programmi altamente specializzati che controllano tutte le attività (guidare, lavorare, ecc.). Con l’inventario dei contenuti si intende la selezione degli schemi adeguati per costruire i comportamenti quotidiani. Il SAS, infine, si occupa del raggiungimento di nuovi scopi tramite la creazione di risposte flessibili. In altre parole, il SAS si attiva quando l’inventario non trova schemi appropriati al compito. Nei soggetti affetti da ADHD il deficit potrebbe essere localizzato a livello del SAS.
Interessante è il modello proposto da Barkley nel 1998, secondo il quale l’ADHD è un deficit che colpisce l’inibizione comportamentale nei suoi tre processi: il primo, che inibisce la “risposta prepotente”5 ad un evento; il secondo è la “risposta campione”, una fase di ritardo riguardante la decisione della risposta; l’ultimo protegge la fase di ritardo da possibili distrazioni o interruzioni. Insieme a questi processi cooperano le funzioni esecutive, fra cui la non-verbal working memory, cioè la capacità di mantenere rappresentata nella mente l’informazione a cui dare una risposta, e la verbal working
memory, che permette all’individuo di verbalizzare l’informazione. Nell’ADHD si
riscontra una disfunzione esecutiva che dà come risultato l’impulsività, la facilità di distrazione e l’iperattività. Questi tre sintomi invalidano il malato a livello scolastico: i tourettiani affetti da ADHD, infatti, perdono facilmente l’attenzione ed hanno molte difficoltà a concentrarsi. Ciò genera frustrazione non riuscendo a mantenere il normale ritmo scolastico.
Accanto a questi sintomi ne compaiono altri, come depressione, disturbi della personalità e comportamento antisociale.
5
Con “risposta prepotente” si intende una risposta immediatamente disponibile perché già precedentemente associata a quell’evento.
I.5 Eziopatogenesi
La sindrome di Tourette è un disturbo di natura neurologica su base ereditaria6 caratterizzato da alterazioni della neurotrasmissione sinaptica, in particolare dopaminergica, che causa lo sbilanciamento dei circuiti cortico-striato-talamico-corticali. La dopamina è nota per avere effetti modulatori ed è prodotta nella sostanza nera e nell’area ventrale segmentale. La dopamina nei gangli della base contribuisce ad attivare i movimenti; nei tourettiani il suo principale recettore, D2, quando è stimolato, causa movimenti stereotipati e iperattività. Sulla base di questi dati si può ritenere che sia coinvolta nella sindrome di Tourette un’alterazione del sistema dopaminergico.
L’ipotesi dopaminergica si basa sui risultati ottenuti nella cura dei tic con l’utilizzo di agenti farmacologici antagonisti della dopamina, quali l’aloperidolo, il pimozide e la flufenazina.
Le analisi strutturali del cervello hanno evidenziato diversità morfologiche nei tourettiani. Innanzitutto, tramite la risonanza magnetica, si è constatato che il tourettiano presenta minori asimmetrie nei gangli della base rispetto ai soggetti normali: l’asimmetria che vede il lato sinistro più ampio è meno evidente nei tourettiani (Peterson et al., 1993:941-949).
Altre ricerche7 hanno mostrato che altre zone dei gangli della base, come il putamen ed il globo pallido, sono più piccole. I gangli della base svolgono un ruolo molto importante nel controllo dei movimenti: queste riduzioni potrebbero essere coinvolte nell’incapacità del soggetto a controllare alcune attività muscolari. Inoltre, i gangli della base fanno parte del percorso cortico-striato-talamico-corticale ad azione inibitoria regolato dalla dopamina, dalla serotonina e dalla norepinefrina8.
Interessante è il dato che, durante le fasi di soppressione del quadro ticcoso, le strutture extrapiramidali sottocorticali sono ipoattive, mentre aumenta l’attività delle aree corticali con funzione inibitoria.
Precisamente, dai dati della PET sono emersi due pattern metabolici: aumento di attività nelle aree premotoria laterale e motoria supplementare, ipofunzione nel caudato e nelle aree talamiche.
6
I dati più convincenti riguardo all’ereditarietà e alla presenza di un gene portatore nella sindrome di Tourette vengono dagli studi sui gemelli. Hyde et al. (1992) hanno dimostrato che esiste una concordanza della sindrome del 56% nei gemelli omozigoti. Sono stati anche esaminati fattori non genetici e a riguardo Singer (1994) sostiene che lo stress materno durante la gravidanza, l’ipertermia e l’infezione da streptococco possono svolgere un ruolo molto importante nell’espressione della sindrome di Tourette
7
http://www.macalester.edu/~psych/whathap/UBNRP/tourettes/.
8
Tra i fattori ambientali implicati nella sindrome, un ruolo di rilievo spetta alle infezioni causate dallo streptococco beta emotilico di gruppo A (SBEGA). Questo batterio, molto diffuso e responsabile di oltre il 50% delle faringotonsilliti o “placche alla gola”, può innescare un processo auto-immunitario, cioè la formazione di anticorpi che per debellarlo si ritorcono contro l’organismo stesso: nello specifico, contro i gangli della base. Questo quadro patologico è definito PANDAS (Pediatric Autoimmune Neuropsychiatric
Disorders Associated with Streptococcal infections). Vi rientrano casi di disturbo
ossessivo-compulsivo, di tic e di sindrome di Tourette.
I.6 Trattamento dei tic
I.6.I Trattamento non farmacologico
Sono state sperimentate numerose tecniche come approcci terapeutici alternativi alla farmacoterapia. I risultati ottenuti, tuttavia, sono scarsi. Tra le tecniche più usate compaiono il training autogeno, l’ipnosi e l’agopuntura: il primo, praticato nella maggior parte dei casi in un contesto di terapia di gruppo, porta benefici soltanto momentanei (Bergin et al., 1998:136-142). Per quanto riguarda le altre terapie, non è possibile dare alcun giudizio poiché i dati pubblicati sono insufficienti (Wu & Kang, 1996:211-213).
I.6.2 Trattamento farmacologico e neurochirurgico
Se è vero che in molti casi di tic semplici è possibile far accettare il disturbo ai pazienti ed ai loro familiari, nella sindrome di Tourette, in cui molti tic sono invalidanti dal punto di vista sociale e fisico, il trattamento di base resta quello farmacologico. I farmaci più efficaci sono quelli attivi sul sistema dopaminergico, detti neurolettici, come la pimozide9, la sulpiride e l’aloperidolo.
Di sicuro interesse sono i dati (Jankovic & Beach, 1997:358-362) riguardanti un potente depletore dopaminergico, la tetrabenazina, ancora non presente in Italia. Questo farmaco ha il vantaggio, rispetto ai neurolettici, di non causare discinesia tardiva, di non sedare, deprimere e far aumentare il peso.
Una tecnica neurochirurgica usata di recente per curare i tic è la Deep brain stimulation10 (stimolazione profonda del cervello): nel nucleo centro-mediano-talamico, la zona cerebrale che
controlla i movimenti, si impiantano in anestesia locale degli elettrodi che stimolano leggermente le cellule nervose e rimuovono i movimenti involontari ticcosi.
9
Gli effetti collaterali riscontrati con la pimozide riguardano alterazioni elettrocardiache, quindi è necessario monitorare il paziente ed effettuare controlli cardiologici accurati.
10
I.7 Il tic
La parola tic, di origine onomatopeica, rievoca il rumore di un colpo secco. Nel ‘600 era usato in medicina veterinaria per denominare il fenomeno, presente in alcuni cavalli, dell’interruzione del respiro con conseguente emissione di uno strano rumore, definito appunto tic. Il cavallo che produceva il tic era, per così dire, “difettoso” e, se venduto, l’acquirente aveva la possibilità di recedere dal contratto di acquisto una volta scoperto il problema. Nel 1664 si pubblicò un volume sulla compra-vendita dei cavalli in cui si consigliava: “Prima di concludere l’acquisto di un cavallo occorre notare se non
sia ticcoso, cioè se non presenti tic. Lo si potrà vedere dal fatto che avrà i denti di sopra e di sotto usati, ma meglio sarà osservarlo mentre mangia, poiché appoggia l’apice dei denti contro la mangiatoia, e produce come un’eruttazione; questo si chiama tic; e per molti versi non vorrei un cavallo come questo, cioè con tale difetto. Prima di tutto il cavallo ticcoso, mangiando, perde una parte della sua avena. In secondo luogo, a forza di avere tic si riempie il corpo d’aria, che spesso gli causa malessere fino a farlo morire. In terzo luogo, diviene magro e non lo si può far ingrassare, perché ordinariamente ha le viscere strette; ed infine questa malattia non si diffonde per contagio ma i cavalli, soprattutto più giovani imparano questo l’uno dall’altro” (Porta, 1996:21).
Successivamente, nel XVIII secolo, il medico Boissier de Sauvages (Arzimanoglou, 1998:761-765) classificò 19 varietà di tic considerandoli movimenti viziosi e ripetitivi.
Con “tic”11 si intende, attualmente, un movimento motorio o una vocalizzazione improvvisa12, rapidi e ricorrenti. I tic si classificano in base alla localizzazione anatomica, alla frequenza, alla complessità e all’intensità. Esempi di tic sono il battito degli occhi, la contrazione del collo o dei muscoli delle spalle, lo schiarimento della gola, cui si aggiungono anche tic complessi, sonori e motori. Secondo Jankovic (1997:267-275), i tic sono così suddivisibili:
11
Ricordo che già nella letteratura dell’800 si distingueva tra tic 'materiali' e tic 'psichici': i primi rappresentano la produzione di un movimento inarrestabile e involontario che può essere strutturato in una serie di atti motori variabili nel tempo. I tic “psichici”, invece, si esprimono in termini di comportamento, gesto o movimento ripetitivo e incoercibile (Porta, 1996). Inoltre, in molti casi di sindrome di Tourette i tic sono rappresentati da successioni variabili di movimenti spasmodici, scosse brusche che colpiscono un gruppo di muscoli.
12
I tic sonori semplici sono causati principalmente dalla contrazione della muscolatura respiratoria, nasale od orale.
a) Tic motori semplici
- clonici13 brevi (100 msec.);
- distonici di durata superiore ai 300 msec.;
- tonici di durata superiore ai 500 msec. e consistenti principalmente in contrazioni isometriche;
b) Tic motori complessi
- movimenti articolati apparentemente non finalistici; - movimenti compulsivi di natura finalistica;
c) Tic sonori semplici
- rumori semplici, colpi di tosse, vocalizzazioni velari;
d) Tic sonori complessi - ecolalia; - palilalia; - coprolalia.
TIC MOTORI TIC SONORI
Semplici:
battito degli occhi; smorfie; protrusione della lingua; torsione del collo; spallucce; tensione addominale; fronte aggrottata; sussulto degli arti.
Semplici:
schiarimento della gola; tosse; sniffing (annusare); fischio; grugnito; versi degli animali
Complessi:
salto; applauso; accovacciamento; bacio; toccare oggetti; coproprassia; ecoprassia.
Complessi:
ecolalia; palilia; imitazione dell’accento; coprolalia.
Tabella 1: Tipologie di tic secondo il DSM IV
13
I tic clonici sono sensibili ai bloccanti della dopamina, mentre i tic distonici sono più difficili da trattare con la farmacoterapia, ma possono rispondere positivamente a locali iniezioni di tossina botulinica.
A loro volta i movimenti ticcosi sono suddivisibili in (Jankovic, 1997):
1) volontari
- intenzionali, cioè iniziati senza una sollecitazione esterna; - responsivi, indotti da stimoli esterni.
2) semivolontari
- indotti da sensazioni fisiche interne; - indotti da compulsioni.
3) involontari
- non arrestabili, come le crisi comiziali o i miocloni; - sopprimibili, come i tremori, le distonie, le stereotipie ecc.
4) automatici - appresi;
- attivati inconsciamente, come l’atto del camminare.
Tutti i tic sono esacerbabili dall’ira o dallo stress, mentre durante il sonno o in una condizione che richiede gran concentrazione possono attenuarsi fin quasi a scomparire del tutto. Molti pazienti sono in grado di controllare i propri tic a costo di un enorme sforzo di volontà o di concentrazione, ma il tic ricompare quando viene meno tale volontà.
I tic, solitamente, sono preceduti da brevi esperienze focali sensorie definite tic sensori14. Questi si riferiscono, nello specifico, a sensazioni spiacevoli, temporaneamente alleviabili con un movimento. I tic sensori complessi, detti anche “tic fantasma”, sono invece associati a proiezioni mentali di impressioni sensoriali provenienti da persone o oggetti e possono essere placati, ad esempio, toccando o annusando la persona o l’oggetto scatenanti.
I tic complessi sono distinguibili dai rituali compulsivi poiché non sono né del tutto consci né finalizzati. Nella pratica, può, tuttavia, risultare difficile distinguere i tic dalle compulsioni. Shapiro e Shapiro (1986) chiamano i tic “compulsioni impulsive”15 mettendo così in luce il fatto che nei tic non ci può essere né consapevolezza né controllo totale. Le compulsioni impulsive o
14
Attualmente al termine tic sensorio si preferisce premonitory sensation.
15
Con “compulsioni impulsive” si vogliono indicare degli atti che sono obbligatori per l’individuo ma sono compiuti in modo automatico, mentre le compulsioni sono atti attentamente controllati.
“impulsioni” secondo i due autori americani inizialmente producono sollievo e soddisfazione, mentre le compulsioni del OCD causano ansia e tensione. George et al. (2000:327-331) sostengono che le impulsioni dei tourettiani sono dirette verso l’Io, mentre le compulsioni sono azioni più elaborate e dirette verso l’esterno, come ad esempio pulire, controllare, ecc. I tourettiani talora presentano “tic ideici”: i pazienti riferiscono, cioè, di avere un’idea fissa che non li abbandona mai e li rende ansiosi ed agitati. E’ possibile confondere il tic ideico con l’ossessione, anche se, come sottolinea Cath et al.(2001:171-185), il pensiero ossessivo è irrazionale e privo di significato, mentre il tic ideico o cognitivo è un pensiero razionale e fondato.
II Il Sistema extrapiramidale
La funzione motoria è elaborata da strutture corticali e sottocorticali complesse che permettono lo svolgimento corretto di un movimento inibendo o facilitando l’azione dei motoneuroni: i movimenti che si creano sono azionati da impulsi nervosi che si trasformano in energia fisica appena raggiungono il muscolo; l’esecuzione di un movimento richiede, accanto alla stimolazione dei muscoli agonisti, anche l’inibizione di tutti i muscoli antagonisti.
Le funzioni motorie si presentano organizzate secondo un sistema gerarchico anche se la capacità di organizzare assieme i diversi movimenti dipende essenzialmente dalle aree corticali motorie che influenzano il sistema corticale e quello extrapiramidale, di cui fanno parte i gangli della base. Il sistema extrapiramidale è un elemento essenziale per la corretta esecuzione di un movimento, poiché ha il compito di orientare l’azione, regolare il tono muscolare e inibire movimenti involontari patologici. Le sindromi da lesione al sistema extrapiramidale, in particolare ai gangli della base, causano squilibri di mediatori chimici16 inibitori ed eccitatori.
II.1 I gangli della base
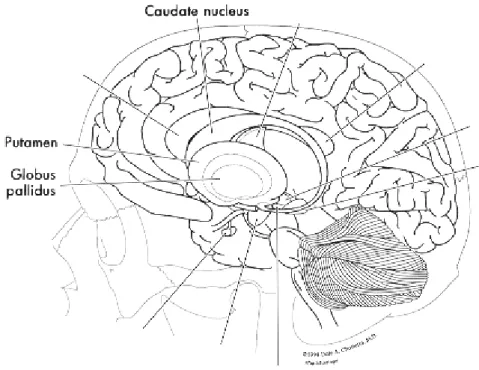
I gangli della base sono un gruppo di nuclei di sostanza grigia: il nucleo caudato, il putamen, il globo pallido interno ed esterno, il nucleo subtalamico e la sostanza nera divisa in parte reticolata e parte compatta ricca di neuroni dopaminergici.
16
Con il termine mediatori chimici inibitori ed eccitatori si fa riferimento a tre gruppi neurali: neuroni gabaergici inibitori con sostanza P, neuroni gabaergici inibitori contenenti encefalite e neuroni colinergici eccitatori.
I gangli della base o sistema extrapiramidale sono costituiti da 3 strutture, il nucleo caudato, il putamen e il globo pallido, poste alla base della corteccia cerebrale e circondate dal talamo e dall’ipotalamo, interconnesse con le aree prefrontali. Queste connessioni sono di importanza essenziale nella regolazione dei movimenti volontari e nell’esecuzione e apprendimento dei programmi motori. I gangli della base sono generalmente considerati responsabili della selezione, esecuzione e inibizione di diversi programmi motori e cognitivi, tra cui quelli coinvolti nella produzione del linguaggio (Marsden, 1982; Mink, 1996).
All’interno del sistema extrapiramidale, ricopre particolare importanza nella adeguata organizzazione sequenziale delle azioni assume il complesso del neostriato (striato), che sembra abbia svolto un ruolo fondamentale nell’evoluzione del pensiero e del linguaggio umano (Lieberman et al., 1992).
Secondo alcuni l’origine neurale della sindrome di Gilles de la Tourette coinvolge il sistema striato-pallido. Del resto, tutti i disturbi in cui si manifesta un problema con la sequenzialità di azioni, parole, pensieri, il deficit vedono coinvolto un errato funzionamento del sistema striato.
Il complesso costituito da putamen e caudato è detto neostriato o semplicemente striato, mentre il globo pallido è detto paleostriato. Il complesso dei nuclei della base integra informazioni afferenti allo striato e trasmette l’elaborazione attraverso proiezioni talamo-corticali.
Figura 1: I gangli della base( www.psychoeducation.org)
Fra le più importanti proiezioni ai gangli della base ci sono quelle provenienti dalle aree prefrontali preposte al controllo dell’attenzione, alla progettazione e all’astrazione. L’interruzione fra le aree prefrontali e i gangli della base, in particolare con lo striato, crea un deficit nelle funzioni esecutive.
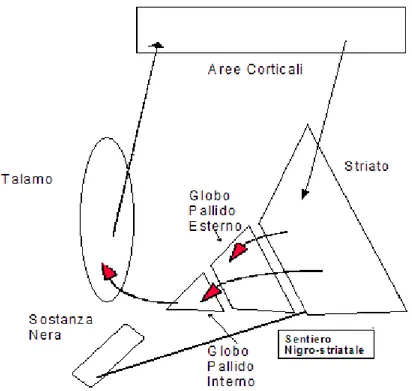
Le connessioni fra i gangli della base e le aree prefrontali risultano di essenziale importanza nella regolazione dei movimenti volontari permettendo ai gangli della base di modulare aree corticali specifiche inibendole o stimolandole tramite i circuiti corticali e sottocorticali. I circuiti partono con proiezioni che vanno dalle aree corticali allo striato, il quale proietta poi al globo pallido che, a sua volta, proietta al talamo, che chiude il cerchio riproiettando alla corteccia.
Figura 2: Connessioni fra gangli della base e corteccia (riadattamento da Thulin, 2002:4)
Cummings nel 1993 individuò 5 circuiti paralleli: un circuito motorio, che si origina nell’area motoria supplementare, un circuito oculomotorio e tre circuiti che nascono nella corteccia prefrontale. Questi ultimi tre circuiti regolano i vari aspetti motori, cognitivi ed emotivi nell’uomo.
Organigramma 1: Organizzazione di tre circuiti dei gangli della base che regolano il controllo motorio, le funzioni cognitive e l'emotività (adattamento da www.epistemic-forum.com)
Corteccia laterale orbitale Corteccia prefrontale dorsolaterale Corteccia anteriore cingolata Caudato (dorsolaterale) Caudato (ventromediano) Nucleo accumbens Globus pallido (laterale dorsomediano) Globus pallido (mediano dorsomediano) Globus pallido (rostrolaterale)
In base alle regioni coinvolte sono individuabili le seguenti connessioni:
1) Il caudato mediano e il nucleo accumbens ricevono input dalla corteccia frontale e dalle aree limbiche;
2) Il caudato e il putamen sono reciprocamente interconnessi con la sostanza nera;
3) Il caudato ed il putamen inviano input al globo pallido; 4) Il globo pallido esterno e il globo pallido interno ricevono input dal caudato e dal putamen, ed entrambi sono in comunicazione con il nucleo subtalamico;
5) Il globo pallido interno invia output inibitori al talamo.
In questo sistema di proiezioni subentra anche il fascio nigro-striatale, formato da proiezioni dopaminergiche della sostanza nera sullo striato. Le proiezioni cambiano effetto secondo il recettore dopaminergico espresso: se le proiezioni GABAergiche dello striato al globo pallido interno esprimono il recettore D1, si avrà un effetto eccitatorio; se, invece, le
proiezioni GABAergiche dello striato al globo pallido esterno esprimono il recettore D2, si
avrà un effetto inibitorio.
Altre attività dei gangli della base includono “the integration of feeling and
movement, the shifting and smoothing of fine motor behavior, the suppression of unwanted motor behaviors, setting the level of body anxiety, controls pleasure and ecstasy and enhances motivation”(St. Clair et al., 2005:3).
II.2 Le funzioni del Neostriato
All’interno del sistema dei gangli della base, particolare importanza nella corretta organizzazione sequenziale delle azioni assume il complesso del neostriato. Tradizionalmente si attribuivano al neostriato le funzioni di inizio ed esecuzione dei movimenti (Denny-Brown, 1962). Negli anni ’80 diversi studi (Marsden, 1982:524-532; Berridge & Fentress, 1987:336-342) hanno invece messo in luce che il neostriato partecipa a funzioni motorie più complesse, come la sequenza dei movimenti. Un supporto a questa ipotesi viene dagli studi riguardanti soggetti parkinsoniani (Harrington & Haaland, 1991:99-115) che hanno evidenziato la difficoltà di questi soggetti ad eseguire azioni da combinare in un ordine preciso. Anche le analisi sulla sintassi dei gesti di pulizia degli animali, il cosiddetto grooming, sostengono che il neostriato si occupa di ordinare
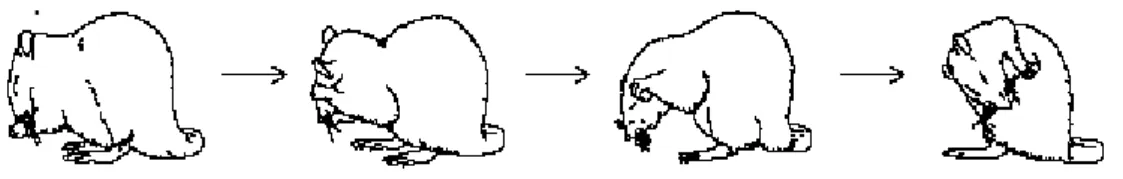
sintatticamente i movimenti. In un articolo di Cromwell e Berridge (1996:3444-3458) si è analizzata la sequenza dei gesti di grooming nei ratti dopo che questi avevano subito piccole lesioni cerebrali nelle aree dorsolaterali e ventrolaterali del neostriato con acido quinolico. Nonostante il deficit, i ratti mantenevano la capacità di compiere azioni di pulizia ma perdevano la corretta sequenza delle azioni. Lesioni, invece, al globo pallido hanno avuto un effetto contrario: danneggiato il movimento, ma conservata la sequenza. Due anni più tardi, Aldridge e Berridge (1998:1-14) studiarono le connessioni neurali coinvolte nella catena sintattica delle attività motorie del grooming dei ratti, una catena di 25 movimenti connessi con un ordine fisso di 4 fasi.
Figura 3: Le quattro fasi principali della grooming dei topi: movimenti ellittico, unilaterale, bilaterale e leccata laterale (Aldridge & Berridge, 1998:2778)
All’inizio della sequenza si attivano immediatamente i neuroni striati. I gangli della base facilitano l’azione sequenziale, ma non prendono parte all’azione di inizio. L’importanza di questo studio è quella di evidenziare che i gangli della base hanno la funzione di ordinare le azioni, mentre l’inizio e la programmazione reclutano altre aree cerebrali, collegate alle strutture subcorticali da una serie di network neurali.
Sulle basi di quanto finora detto è possibile pensare che, oltre alla discinesia, alla corea e all’acinesia, i danni al sistema neostriato coinvolgono anche funzioni mnestiche e cognitive. Si è anche ipotizzato che la funzione di sequenziazione delle azioni svolta dal neostriato abbia svolto un ruolo fondamentale nell’evoluzione del pensiero e del linguaggio umano. Infatti, una lesione al sistema del neostriato o, più in generale, al sistema striato, causa difficoltà di comprensione sintattica17 non solo delle azioni ma anche del linguaggio. Lieberman et al. (1992:169-189) riferiscono che, negli stadi finali del morbo di Parkinson, i malati hanno difficoltà nel comprendere le frasi con sintassi complessa (es: il cane era inseguito dal gatto), mentre comprendono facilmente frasi con sintassi semplice (es: il gatto inseguiva il cane).
17
Marsden (1992:171-178), proprio a partire dai sintomi del morbo di Parkinson, affermava che “the normal operation of the basal ganglia……may be the orderly and
rational sequencing of the individual components of motor and cognitive plans”.
I disturbi come la sindrome di Gilles de la Tourette o il disturbo ossessivo-compulsivo sono caratterizzati da una scorretta sequenzialità di azioni, frasi, pensieri. Per quanto non sia ancora chiara la correlazione anatomica, si pensa sempre più spesso al sistema striato-pallido. E’ plausibile, dunque, che, quando si manifesti un problema con la successione di azioni, parole, pensieri, il deficit possa essere causato da un errato funzionamento del sistema striato.
II.3 La citoarchitettura cerebrale
Fisiologi e neurologi sono concordi nell’affermare che non esiste un centro specializzato del linguaggio, poiché il cervello è un sistema di sistemi fra loro interconnessi. Le regioni o aree macroscopiche, come ad esempio le aree corticali o subcorticali, sono composte di piccoli circuiti locali, neuroni connessi tramite sinapsi. I gangli della base ed il sistema limbico sono strutture macroscopiche fra loro collegate. Appena ci si muove dal livello macro a quello micro, ci si rende conto che ogni area è collegata ad un'altra, attraverso una catena di neuroni. I neuroni possiedono gli assoni che inviano segnali (neurotrasmettitori) ad altri neuroni attraverso le sinapsi. I messaggi sono invece ricevuti attraverso le sinapsi dai dendriti, appartenenti anch’essi ai neuroni. Ogni neurone contiene una media di 1000 sinapsi, un numero molto più ampio delle connessioni neurali.
III Il sistema extrapiramidale e il linguaggio: l’ipotesi di Lieberman
III.1. Alcune considerazioni
Nel 1993 Philip Lieberman osservò che i cinque membri della spedizione americana sul monte Everest, Sagarmatha, durante l’ascesa presentavano notevoli problemi di controllo delle azioni motorie del linguaggio e di comprensione sintattica. Tra tali deficit temporanei compariva il deterioramento della produzione linguistica ed un lieve danno nella memoria a breve termine. Alcuni anni prima erano stati condotti degli esperimenti (Kennedy et al.,1989:99-104) che in una camera ipobarica simulavano l’ascesa: emerse sperimentalmente un deficit nel ragionamento grammaticale che durò alcuni giorni.
Una possibile spiegazione dei disturbi che si presentavano è l’ipossia cerebrale, determinata da un basso contenuto di ossigeno nell’aria inspirata. Gli studi istologici sull’ipossia (Brierley, 1976) hanno segnalato le aree bersaglio dell’ipossia: ippocampo, cervelletto e gangli della base.
Interessante è notare che gli stessi tipi di disturbo presenti nell’ipossia si hanno anche nel morbo di Parkinson, delle cui difficoltà di comprensione sintattica si è già detto, possibile espressione di un danno ai circuiti subcorticali diretti alla corteccia prefrontale. Già Grossman et al. (1991:1620-1628) avevano evidenziato difficoltà di comprensione sintattica e ricorso a frasi brevi e con sintassi semplificata nei parkinsoniani. Nello studio del 1991, Grossman e la sua équipe constatarono un’analogia tra i deficit linguistici nel morbo di Parkinson e quelli nell’afasia di Broca. Testarono, dunque, i soggetti parkinsoniani con un compito di abilità manuale che consisteva nell’eseguire sequenze manuali non familiari. I risultati hanno evidenziato nei parkinsoniani gravi difficoltà nell’esecuzione di sequenze, sia linguistiche sia manuali. Nel 1993 Kimura ripeté lo stesso test su soggetti afasici di Broca e notò difficoltà nel riprodurre sequenze motorie. Secondo
Negli ultimi anni l’ipotesi modulare che sostiene la presenza di aree specializzate per la funzione ‘linguaggio’ ha subito molte critiche ed è stata soppiantata da altri pareri, tra cui quella dello scienziato cognitivo e linguista Philip Lieberman, che sostiene l’esistenza di un sistema funzionale del linguaggio (FLS), sviluppatosi nel corso dell’evoluzione e composto da una serie di strutture e sottosistemi cerebrali fra loro interagenti e direttamente connessi ai gangli della base.
A sostegno del FLS Lieberman ricorda i dati emersi dall’analisi comparativa del linguaggio di afasici di Broca e di parkinsoniani, da cui emerge la non esclusività delle strutture corticali nella produzione linguistica accentuando il ruolo del sistema extrapiramidale nel processo sintattico.
Ulteriore supporto a questa tesi viene dagli studi sul gene FOXP2, isolato nei membri della famiglia KE, e che in prima istanza, convalidava l’ipotesi innatista. Approfondite indagini (Vargha-Khadem, 1995) hanno evidenziato che le difficoltà linguistiche dei membri della famiglia KE possano conseguire ad un disturbo nel controllo motorio.
Lieberman “deficits in sequencing manual motor movements and linguistic sequencing in
the sentence comprehension task were correlated. The correlation between sequencing complex manual motor movements and the cognitive operations implicated in the comprehension of syntax is consistent with Broca’s area playing a role in both verbal memory and manual motor control circuits supported by basal ganglia” (2002: 45).
Grossman et al. (2001:123-130) ipotizzarono che i deficit linguistici nei parkinsoniani fossero espressione di un deficit dell’attenzione, ipotesi confutata da Lieberman che afferma “… PD subjects…consistently perform better when faced with a
sentence such as ‘the banana was eaten by the boy’ than the sentence ‘the clown was poked by the cowboy’. They clearly have no attentional deficits regarding the fact that an inanimate banana cannot eat a boy” (2002:45).
Nei parkinsoniani, oltre ai problemi di comprensione, si manifesta un danno nella sequenza dei comandi motori coinvolti nell’esecuzione delle consonanti occlusive. Il segnale acustico che differenzia [b] da [p], quando precedono una vocale, è l’intervallo di tempo nella sequenza dei comandi motori nella produzione delle due consonanti18. Come ricorda Lieberman: “these speech sounds are produced by closing the lips, obstructing the
flow of air from a speaker’s mouth, and then abruptly opening the lips, which produces a ‘burst’ of turbulent air that has distinct acoustic properties. At the same time, the speaker must adjust the muscles of the larynx to produce phonation subsequent to the burst. In order to produce a [b], phonation must occur within 20 msec of lip opening; longer delays will yield the sound [p]” (2002:46). Nei soggetti affetti da morbo di Parkinson e negli
alpinisti della spedizione sull’Everest si manifesta un problema nel controllo delle sequenze motorie delle occlusive, per cui intendono [b] anche se il suono prodotto è più simile a [p].
Si può notare, da quanto finora detto, che i gangli della base svolgono un ruolo fondamentale nella produzione e comprensione linguistiche, diversamente da quanto si riteneva fino al 19° secolo, e cioè che le uniche aree cerebrali coinvolte nell’attività linguistica fossero l’area di Broca e l’area di Wernicke.
III.2 Partendo dal “gene del linguaggio”
Nel 2001 un équipe di genetisti di Monaco di Baviera individuò il “presunto” gene del linguaggio dopo aver analizzato il DNA dei membri della famiglia KE, affetti da deficit
18
Questo fenomeno prende il nome di Voice Onset Time (VOT) e descrive il tempo che intercorre fra il burst e l’inizio della fonazione periodica generata dalla laringe.
linguistici riguardanti la sequenza delle parole, e di un altro soggetto, CS, con deficit identici a quelli dei KE, e dopo essere giunti alla conclusione che il gene FOXP2 in questi pazienti manifesta una mutazione. Il FOXP2 appartiene ad un gruppo di geni che secernono la proteina FOX, che fa parte del gruppo proteico conosciuto come fattore di trascrizione e interessato nel controllo dei programmi genetici delle singole cellule.
Il gene FOXP2 è coinvolto nello sviluppo cerebrale e si ipotizza che prenda parte alla creazione di substrati neurali implicati nell’acquisizione del linguaggio, ma è coinvolto anche nello sviluppo di altri organi, come i polmoni ed il cuore. A fronte di queste considerazioni, è lecito domandarsi perché i deficit della famiglia KE e del soggetto CS riguardano solo le funzioni linguistiche. Marcus e Fisher (2003:257-262), uno psicologo ed un genetista americani che da qualche anno si occupano del caso FOXP2, rispondono a questa domanda sostenendo che esistono due coppie di uno stesso gene, perché i geni umani, duplici per natura (uno viene ereditato dalla madre ed uno dal padre), perciò, così “the problems of the KE family are associated with mutation in just one copy of the FOXP2
gene, which probably leaves affected members with only half the usual dosage of normally functioning FOXP2 protein” (Marcus et al., 2003:261).
Sono stati condotti anche alcuni studi (Shu, 2001:27488-27497) comparativi fra esseri umani, topi e scimmie da cui sembra risultare che questo gene abbia svolto un ruolo primario nello sviluppo cerebrale. La presenza di FOXP2 negli animali non ne diminuisce il significato rispetto al linguaggio, ma piuttosto fornisce un’ulteriore testimonianza delle modificazioni e adeguamenti verificatisi nel cervello nel corso dell’evoluzione. E’ probabile che i geni implicati nel linguaggio siano coinvolti, anche, in altre funzioni. Consideriamo, ad esempio, la funzione della gerarchia grammaticale: è probabile che i geni che vi operano influenzino anche la rappresentazione gerarchica dei “progetti”, anche se i circuiti coinvolti in queste funzioni sono situati in aree diverse. Emerge, quindi, l’ipotesi che aree cerebrali diverse elaborano funzioni simili, permettendo contemporaneamente la progettazione linguistica di una frase e di una sequenza motoria. A tale proposito, tornando al caso della famiglia KE, studi approfonditi hanno mostrato che nei membri malati di questa famiglia, circa 15 in tre generazioni, sono presenti, oltre ai disturbi linguistici anche disordini motori localizzati nell’area orofacciale e deficit cognitivi, nonché difficoltà nelle capacità percettive ed espressive. L’aprassia orofacciale è un aspetto prominente dei membri della famiglia KE: Vargha-Khadem (1995) ha dimostrato che l’aprassia orofacciale determina difficoltà nel controllo e nella coordinazione dei
muscoli del viso e della bocca, pertanto, tali difficoltà possono facilmente creare impedimenti nell’espressione linguistica.
L’ipotesi suggerita da Vargha-Khadem induce a pensare che le difficoltà espressive nel linguaggio orale di alcuni membri della famiglia KE potrebbero essere conseguenti ad un disturbo nel controllo motorio. Di contro, però, ci sono alcune considerazioni: innanzitutto, il deficit dei soggetti KE non riguarda solo il linguaggio orale, ma anche quello scritto. Inoltre, si presentano deficit di comprensione e di lettura per quanto riguarda espressioni di una certa lunghezza. Infine la fMRI ha evidenziato in questi soggetti anomalie nelle regioni corticali tradizionalmente legate alla funzione linguistica.
Anche Watkins et al. (2002:465–478) concludono che i deficit verbali e non verbali della famiglia KE sono il risultato di un deficit di esecuzione di movimenti sequenziali. A sostegno di questa ipotesi ci sono i dati emersi dalla PET e dalla fRM che mostrano la presenza nei membri della famiglia KE di una riduzione volumetrica dei gangli della base. Il disturbo, quindi, si confermerebbe trasmesso geneticamente ma non sarebbe specificamente linguistico.
La presenza di deficit cognitivi, motori e linguistici in caso di un danno ad una delle componenti dei circuiti cortico-striato-corticali era già stato notata nel 1989 da Kimura e Watson, i quali, analizzando soggetti afasici con lesione focale, avevano riscontrato deficit nelle attività di coordinamento delle sequenze vocali e gestuali. Un anno prima, Dewey et
al. (1988:743-751) avevano osservato che i bambini affetti da aprassia19 mostravano gravi deficit cognitivi e linguistici.
Da quanto finora detto risulta un quadro in contrasto con l’idea locazionista stretta20, in quanto una stessa area cerebrale risulta in grado di intervenire in diverse funzioni in base all’attivazione di determinati circuiti neurali. Dai citati esperimenti di Kimura con soggetti afasici è emerso che aree solitamente considerate linguistiche prendono parte anche a funzioni di controllo motorio e viceversa. E’ chiaro che il cervello
19
L’aprassia è definita come l’incapacità di eseguire movimenti preordinati ed in sequenza.
20
L’ipotesi locazionista era già stata confutata agli inizi del’900 dallo psicologo russo Vygotskij che sosteneva che una sola funzione, ad esempio quella linguistica, era il risultato di un’attività complessa che coinvolgeva diverse strutture cerebrali. Vi era, perciò, una cooperazione dinamica controllata da strutture neurali in grado di monitorare diversi organismi.
Anche un altro psicologo russo, Lurija, ipotizzò una teoria in contrasto con l’ipotesi locazionista. Infatti, osservando i soggetti afasici e parkinsoniani, si accorse che era impossibile sostenere una rigida e meccanicistica corrispondenza fra funzione e area cerebrale specifica, mentre si poteva ipotizzare che le diverse funzioni cognitive fossero dei sistemi funzionali (concetto quest’ultimo ripreso in seguito da Lieberman nel suo FLS) che investono varie aree cerebrali interconnesse fra loro. Il cervello, secondo Lurija, è un sistema di connessioni dove un compito può essere eseguito da diversi meccanismi ottenendo lo stesso risultato. Per tali motivi è impossibile localizzare la funzione “linguaggio” in una determinata area.
non è organizzato secondo unità discrete o moduli specializzati nello svolgere una sola funzione; la “funzione linguaggio” non è circoscritta a determinate aree del cervello soltanto. Secondo Mesulman il cervello “is mapped at the level of multifocal neural system
rather than specific anatomical sites”(1990:589).
Partendo da queste considerazioni, Lieberman rigetta l’idea che il cervello è formato da “moduli” indipendenti ed atti esclusivamente ad una specifica funzione, e sostiene, invece, la teoria del sistema neurale funzionale (di cui fa parte il functional
linguistic system, FLS), un network di circuiti neurali che lavorano assieme per portare a
termine le diverse funzioni del comportamento umano. E’ importante notare che i network che collaborano per eseguire le attività linguistiche comprendono le strutture subcorticali oltre alle aree corticali considerate tradizionalmente.
III.3 Verbal Working Memory
Si è sempre ritenuto che l’area di Broca fosse implicata nella comprensione delle frasi. Secondo Lieberman questa affermazione non è del tutto accettabile. Le più recenti ricerche, soprattutto grazie alle tecniche di neuroimmagini, hanno mostrato che, durante i compiti di comprensione di frasi, viene coinvolto, oltre all’area di Broca, anche un magazzino neurale di memoria a breve termine, chiamato Verbal Working Memory. Questo sistema neurale è dinamico ed ingaggia risorse addizionali secondo la complessità del compito: ad esempio, possono essere impiegate anche aree addette al ragionamento astratto, come le regioni del lobo frontale. Uno studio di Awh et al. (1996:25-31), condotto utilizzando la PET, ha evidenziato che le strutture impiegate nella Verbal Working Memory e nel controllo motorio sono anche implicate nel processo di elaborazione sintattica di una frase. Tali risultati avallano nuovamente l’ipotesi che l’area di Broca non sia l’unico centro di produzione e comprensione linguistica: in altre parole, non è “l’organo della sintassi”21. Diverse sono le aree cerebrali che si attivano nella comprensione di frasi. Just et al. (1996:114-116), sottoponendo soggetti neurologicamente sani ad un test di comprensione sintattica con complessità crescente22, hanno dimostrato che si attivano la corteccia temporale, il giro superiore temporale, l’area di Wernicke e l’area di Broca. Inoltre, con
21
L’espressione “organo della sintassi” è stata coniata da Grodzinsky per riferirsi all’area di Broca che “is thought to house mechanisms that compute dependencies among nonadjacent sentential constituents, established by transformational relations” (2000:83). Tale ipotesi si fonda sull’analisi sintattica dell’eloquio dei soggetti affetti da afasia di Broca e sui dati offerti dalla fMR.
22
Esempi di frasi con complessità sintattica crescente sono: “The reporter attacked the senator and admitted the error”, “The reporter that attacked the senator admitted the error”, “The reporter that the senator attacked admitted the error”.
l’aumentare della complessità sintattica della frase si attivano anche nell’emisfero destro le aree omologhe all’area di Wernicke e Broca.
Prima si è accennato al “lessico mentale” che prende parte alle attività di comprensione della frase. Comprendere una frase significa prima di tutto identificare le parole, comprenderne il significato e le loro restrizioni sintattiche. E’ un processo complesso che richiede che “the concepts that are coded by a word result in the activation
of the brain mechanisms that concern the real-world attributes of the word in question”
(Lieberman, 2002:49). Un esempio ci viene dall’esperimento di Martin et al. (1995) in cui si nota, tramite la PET, che si attiva la corteccia motoria primaria se pensiamo ad un utensile, mentre si attiva l’area visiva primaria associata a forma e colore se si pensa ad un oggetto o ad un animale. Si può, quindi, usare la metafora del dizionario per comprendere l’attività svolta dal cervello quando sente una parola.
Damasio et al. (1996:409-505), hanno studiato 29 soggetti sia sani sia con lesioni cerebrali focali e hanno notato che i soggetti con danno cerebrale presentavano vari deficit in compiti di riconoscimento di immagini23: alcuni non riuscivano a denominare persone ed animali, altri gli utensili, altri ancora sia gli animali sia le persone sia gli utensili. Durante il test è anche emerso che, a seconda della categoria, si attivavano regioni differenti.
III.4 FLS: il pensiero di Lieberman
La facoltà del linguaggio è stata considerata per molto tempo una capacità innata, un sistema unico e specifico con principi propri (Chomsky, 1990). Philip Lieberman rivede la teoria innatista sostenendo che il linguaggio è un’abilità appresa che ha le sue radici in un impianto biologico definito Functional Language System (FLS).
Il FLS di Lieberman è un sistema che si è sviluppato nel corso dell’evoluzione, formato da diverse strutture fra loro interagenti. Questo sistema è composto da una serie di sottosistemi cerebrali molti dei quali sono direttamente connessi alle strutture subcorticali. Tra queste, specificatamente i gangli della base regolano le sequenze motorie del linguaggio (ma non solo). Il modello FLS considera, dunque, la produzione linguistica risultato dell’attività di molte aree cerebrali. Non viene negato, ovviamente, che le aree di Broca e di Wernicke sono coinvolte nella elaborazione linguistica, ma viene rigettata l’idea che queste aree sono moduli indipendenti ed esclusivi del “linguaggio”. In questa ottica
23
Ai soggetti erano mostrate fotografie di persone note, di animali e di oggetti e veniva chiesto loro di fornire la parola più specifica per definire ogni immagine.
anche le strutture subcorticali non sono più esclusivamente aree responsabili dell’attività motoria. E’ probabile che attraverso processi evolutivi si siano modificate fino ad essere impiegate anche in attività cognitive e linguistiche.
Secondo questa visione l’idea dell’innatismo biologico non si contrappone all’ipotesi evoluzionistica e filogenetica. Le strutture biologiche ci predispongono in maniera innata ad imparare e a rispondere al linguaggio.
Dunque, secondo Lieberman il linguaggio è innato nel senso che nel nostro cervello esistono strutture biologiche congenite che si occupano di elaborarlo. L’esperienza è lo stimolo esterno (interazione con gli altri membri di una comunità) per attivare queste strutture e rimodellarle. I bambini apprendono una lingua grazie ad un’interazione socio-linguistica che, sin dai primi mesi, si manifesta nel rapporto con la madre e che non si basa sull’ascolto.
La nostra esperienza, inoltre, plasma e modifica la corteccia. Come ha notato Evarts (1972:25-31), nell’uomo la corteccia motoria primaria ha la funzione di trattenere patterns motori e di rispecificare il compito dei circuiti neurali con i nuovi movimenti appresi da un’esperienza reiterata. I circuiti neurali non sono, quindi, specifici ed univoci, prendono parte a diverse funzioni, tanto che le funzioni motorie e cognitive possono condividere gli stessi circuiti neurali. La plasticità corticale e subcorticale fa in modo che i neuroni, presenti nei circuiti, si possano riformare in modo diverso secondo l’esperienza vissuta dall’individuo. Anche le simulazioni di reti neurali hanno mostrato che è possibile decodificare regole dall’esperienza. Un caso evidente di plasticità e adattamento cerebrale è dato dai sordi: la lingua dei segni fa attivare la corteccia uditiva (Nisimura et al., 1999:116), come se i loro segni manuali non fossero solo visti ma anche uditi.
III.5 Il ruolo nell’evoluzione delle strutture subcorticali
Dunque, è ormai evidente che le basi neurali del linguaggio non risiedono esclusivamente nelle classiche aree linguistiche di Broca e di Wernicke. Il linguaggio umano si colloca in un network di circuiti collegati a diverse strutture sia corticali che subcorticali.
I circuiti neurali cortico-striato-corticali sono coinvolti nella comprensione, in varie attività cognitive, nella creazione e decodificazione di sequenze motorie e linguistiche. Nello specifico, i gangli della base sono essenziali per il corretto funzionamento di questi circuiti e per il controllo delle funzioni cognitive e motorie. Le sequenze motorie riprodotte
grazie all’aiuto delle diverse strutture cerebrali sono apprese dall’esterno, dall’esperienza che modifica le strutture cerebrali biologicamente trasmesse.
Diversi studi comparativi hanno mostrato che il linguaggio umano condivide alcune caratteristiche con i sistemi comunicativi di altre specie. Ad esempio, la capacità di denominare gli oggetti, le azioni e tutto ciò che fa parte del mondo che ci circonda è una caratteristica presente anche negli scimpanzé esposti a forme non verbali (vedi ad esempio i segni dell’ASL). Infatti, queste scimmie possono apprendere fino a 150 segni e sono in grado anche di generalizzarli e decontestualizzarli. Secondo Lieberman, l’abilità lessicale poteva già essere presente agli inizi dell’evoluzione degli ominidi.
Per quanto riguarda la sintassi, si possono considerare nuovamente i dati raccolti dagli esperimenti sugli scimpanzé, che riescono a comprendere ed usare frasi semplici in ASL. In altre specie, come la scimmia Diana, Zuberbuhler (2002:293-299) mostra che tali scimmie rispondono diversamente all’allarme di richiamo di un’altra specie, le scimmie di Campbell, a seconda della sequenza di suoni emessa da queste ultime.
E’ probabile che nei primi ominidi i comandi sintattici fossero simili a quelli degli scimpanzé e che solamente con l’evoluzione dei circuiti cortico-striato-corticali si sia creata una sintassi più completa. L’abilità lessicale e sintattica sono proprietà primitive e, secondo Deacon (1997), si sono co-sviluppate con l’aumento del cervello, in particolare della corteccia prefrontale, del cervelletto e dei gangli delle base, strutture più ampie nell’uomo rispetto agli scimpanzé. I gangli delle base e la corteccia prefrontale sono di estrema importanza, come ho già più volte ricordato, proprio per l’apprendimento di programmi e sequenze motorie e cognitive.
Vi sono, invece, molte difficoltà nell’individuare le aree di Broca e di Wernicke negli ominidi. Negli ultimi anni, oltre ai ricordati studi di Kimura, che rilevano il coinvolgimento dell’area di Broca nella programmazione e nel controllo motorio manuale, è stata individuata da studi di Rizzolatti e Arbib e di altri nell’area F5, contenente neuroni specchio implicati nell’imitazione e nel controllo motorio manuale24.
24
IV I Neuroni Specchio
Come ricordato nel capitolo precedente, Lieberman nel suo Reptilian Brain ci ricorda che i gangli della base e le strutture subcorticali in genere non solo partecipano alla sequenziazione degli atti motori, i nostri “gesti”, ma anche alla coordinazione articolatoria del linguaggio. Inoltre, gli studi sui Parkinsoniani, già ricordati, ci dicono che un danno alle strutture subcorticali ha effetti sul linguaggio simili a quelli presenti negli afasici di Broca. Esiste, quindi, un rapporto tra azione e parola che va oltre la semplice coordinazione e sequenziazione dei movimenti. Ad esempio, secondo Nicolai (2006), il rapporto tra le parole di azione (verbi) e le azioni è molto profondo, si può dire che l’azione stessa soggiace al verbo, “(I)l verbo è un’azione o delle azioni della struttura
corporea e del sistema neuromuscolare. Le lingue gestuali risvegliano così in ogni soggetto la coscienza dei fondamenti metaforici del linguaggio, spesso seppelliti sotto espressioni verbali disincarnate del gestuale che le ha fatte nascere” (Nicolai, 2006:79).
In effetti, i gesti contengono in sé tutte le caratteristiche contenute nel loro corrispettivo verbale e descritte tramite una successione di azioni, sono, in altre parole, rappresentazioni animate che, come il linguaggio, creano relazioni tra elementi diversi.
La teoria dei neuroni specchio proposta da Arbib cerca di spiegare la stretta relazione fra gesto e linguaggio, considerando il linguaggio il punto di arrivo della comunicazione linguistica, iniziata molto probabilmente attraverso le “azioni”.
Nel 1998 Rizzolatti e Arbib pubblicano uno studio in cui si dimostra la presenza nei macachi di neuroni specchio, cioè neuroni che si attivano nell’area cerebrale F5 sia quando il macaco compie un’azione di presa, come afferrare una noce di cocco, sia quando il
Gli studi di neuroimmagine funzionale condotti sui macachi hanno rilevato la presenza nell’area F5 di un gruppo di neuroni che si attivano sia quando la scimmia compie una presa sia quando osserva un suo simile o un uomo afferrare un oggetto. E’ stato, perciò, ipotizzato che questi neuroni, denominati neuroni specchio, sono fondamentali per comprendere le azioni e i loro programmi motori. Un dato importante è giunto da un’analisi comparativa del sistema cerebrale umano con quello dei primati, da cui sembra emergere che l’area F5 corrisponda all’area di Broca. Questi dati hanno suggerito a un gruppo di ricercatori guidati da Arbib e Rizzolatti (1998) che i neuroni specchio abbiano posto le basi biologiche per l’evoluzione del linguaggio umano.
Su tali presupposti hanno, infine, creato diversi modelli (Fagg-Arbib-Rizzolatti-Sakata-FARS e hand-state) che spiegano il ruolo dei neuroni specchio e la loro specializzazione, sottolineandone l’importanza nell’apprendimento di un repertorio di azioni tramite l’imitazione sia di azioni innate sia di azioni apprese.
I neuroni specchio, secondo Arbib (2006) costituiscono l’anello ‘neurale’ mancante fra il linguaggio orale dell’uomo moderno e le abilità motorie e di comunicazione gestuale dei nostri antenati ominidi. Arbib ipotizza, allora, che nel corso dell’evoluzione l’uomo abbia adattato il sistema dei neuroni specchio per la comunicazione, passando dalla mera imitazione, alla pantomima fino alla creazione di gesti simbolici che hanno fornito le basi per la creazione di un lessico prima gestuale e poi orale.
macaco osserva un altro macaco, o uomo, compiere un’azione di presa. Inizialmente la presenza dei neuroni specchio era stata osservata, anche se non ancora nella sua forma completa, dal gruppo di ricerca guidato dal gruppo di Rizzolatti che si occupava della corteccia premotoria ed in cui si indicava che nel solco intraparietale anteriore e nell’area F5 delle scimmie si formano degli elementi chiave facenti parte di un circuito corticale che trasformano informazioni visive su proprietà intrinseche di un oggetto target in movimenti manuali che permettono all’animale di afferrare correttamente l’oggetto. In altre parole si riconosceva in queste aree la presenza di neuroni allenati a riconoscere le caratteristiche di un oggetto in modo da poter compiere la presa adeguata all’oggetto. Secondo Rizzolatti et
al. (1988:491-507) l’area F5 contiene un vocabolario di schemi motori in cui si scelgono le
azioni appropriate a rispondere a determinate caratteristiche fisiche dell’oggetto.
Nel 1998 Rizzolatti ed Arbib classificano i neuroni dell’area F5 in diverse categorie corrispondenti all’azione da compiere, ad esempio i neuroni “della presa con la mano”, i neuroni del “sorreggere”, i neuroni specchio, invece, sono quelli che si attivano anche quando l’azione è osservata. Bisogna notare, però, che i neuroni specchio non si attivano in risposta alla presenza di un oggetto, ma solo se si compie un’azione. Inoltre, fra i neuroni specchio ve ne sono alcuni altamente specializzati che si attivano, ad esempio, solo se l’azione è compiuta con il pollice, o quando l’oggetto è afferrato con la bocca (Ferrari et
al., 2003:1703-1714).
Si può dedurre che i neuroni specchio ed i neuroni canonici dell’area F5 differiscono perché questi ultimi si attivano solo quando l’animale compie un’azione in risposta ad un oggetto target, mentre i primi si attivano anche quando l’azione è osservata. Il compito dei neuroni specchio, quindi, secondo Rizzolatti ed Arbib, è principalmente quello di comprendere un’azione che un altro individuo sta compiendo e differenziarla dalle altre25.
25
La scoperta dei neuroni specchio fornisce un supporto parziale alla Teoria Motoria della Percezione Linguistica, secondo la quale il link fra emittente e destinatario non è il suono, ma un meccanismo neurale condiviso che riconosce i “gesti fonetici” e ne permette la produzione. Nello specifico questa teoria, elaborata da Alvin Liberman negli anni ’60, sostiene che l’informazione uditiva aziona nell’ascoltatore un piano dei movimenti in grado di ricreare quell’informazione. Dallo schema uditivo si produce uno schema motorio e l’unione di questi due schemi è ricreata quando l’ascoltatore percepisce una sequenza acustica di suoni che automaticamente e istantaneamente si collega allo schema motorio dell’ascoltatore. Per creare uno schema motorio l’ascoltatore adotta il sistema imitativo dei gesti, intesi in questa ottica come il punto di partenza di qualsiasi tipo di informazione.
IV.1 L’ipotesi del sistema specchio
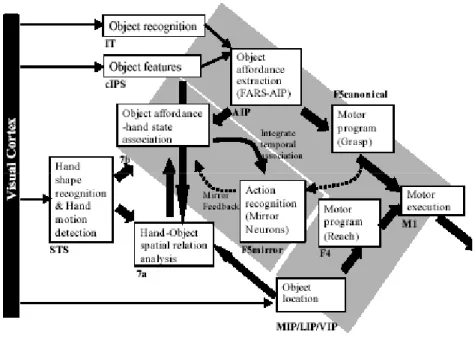
Taira et al. (1990:29-36) hanno mostrato che nel solco intraparietale anteriore (AIP) si attivano dei neuroni alla vista di un oggetto manipolabile. Nel modello FARS (Fagg - Arbib - Rizzolatti - Sakata) si mostra la collaborazione fra l’AIP e l’area F5: entrambe agiscono all’interno di un circuito di trasformazione visivo-motoria che trasforma nel cervello la vista di un oggetto nell’esecuzione motoria di una presa. Si può altresì dire che l’AIP fornisce una presa su un oggetto e la F5 seleziona e guida l’esecuzione di una presa.
Per essere più precisi l’AIP estrae le caratteristiche di un oggetto e la F5 seleziona la presa adeguata dal menù dell’AIP. L’AIP, pertanto, gioca un ruolo molto importante non solo nel processo che sviluppa un’azione in risposta ad un oggetto, ma anche come “magazzino delle prese”.
Negli anni ’90 Iberall e Arbib proposero un modello alternativo al FARS, l’ipotesi
hand-state.
Per comprendere l’ipotesi hand-state è opportuno prima introdurre il concetto di
virtual fingers, le entità fisiche costituite da una o più dita, o dal palmo della mano, usate
tramite l’applicazione di una forza al fine di compiere una presa su un oggetto. I controlli motori partecipano predisponendo la mano alla presa di un oggetto, e sono le sue caratteristiche fisiche che determinano il tipo di presa. Successivamente si ha la manifestazione motoria della presa con l’estensione appropriata del braccio, calcolata in base alla distanza dall’oggetto. La funzione dei neuroni specchio in questa ipotesi è l’elaborazione dell’automatismo adatto, l’hand-state. Così “(G)iven this functionality, the
social role of the F5 mirror system in understanding the actions of others may be seen as an exaptation gained by generalizing from self-hand to other’s-hand” (Oztop &Arbib,
http://www.hbp.usc.edu/).
Il Sistema del neurone specchio si fonda sulle sopraccitate argomentazioni e pone l’accento sulle connessioni esistenti fra le diverse aree cerebrali. Prima di spiegare questo sistema, bisogna sottolineare che nell’area F5, oltre ai neuroni specchio, sono presenti anche i neuroni canonici che si attivano solo durante l’azione.
Il sistema dei neuroni specchio opera in due modi: tramite la comprensione ed il riconoscimento dell’azione. Nel primo modo si estraggono le informazioni sull’oggetto al fine di eseguire il programma motorio di presa più adatto. Nel secondo modo si estrapolano le caratteristiche dell’oggetto così da definire la traiettoria e la forma della mano in movimento anche quando è osservata. I neuroni specchio, quindi, rispondono alla