4 RISULTATI
4.1 Marcatori sceltiScopo della tesi era studiare il ruolo di XBH1 nel patterning neurale durante gli stadi precoci di sviluppo del sistema nervoso di Xenopus laevis e la possibile dipendenza della sua funzione dalla sumoilazione.
Per fare ciò embrioni sono stati iniettati con mRNA di XBH1 wild type e mutanti, sono stati fissati in MEMFA a stadio di neurula ( st. 15-16) e sottoposti a whole mount ISH usando diverse sonde. In particolare, ho iniettato mRNA per i seguenti costrutti Xbh1-wt-HA , Xbh1-wt e Xbh1-DM, (vedere capitolo 2, paragrafo 1) nella regione dorso-animale, corrispondente alla piastra neurale presuntiva (v. oltre) insieme all’mRNA di LacZ codificante per la galattosidasi ( abbreviato gal), un enzima che fa assumere alle cellule che lo esprimono, se opportunamente trattate ( vedere capitolo 3 paragrafo 11) colorazione rossa.
Ai fini degli esperimenti sia di whole mount ISH che di saggio TUNEL whole mount sono stati considerati considerati nella casistica solo gli embrioni che presentavano il segnale della gal, definiti d’ora in poi nel testo gal positivi ( gal +)
I marcatori neurali scelti possono essere raggruppati in tre categorie: - geni del patterning antero-posteriore Xotx2 Xen2 Xbh2 Xkrox20 - geni espressi nel campo ottico Xrx1 Xpax6
- geni pan-neurali Xsox2 Xntub
I geni del primo gruppo sono già stati descritti nell’introduzione.
Questa catalogazione è una semplificazione infatti anche i geni espressi nel campo ottico hanno un ruolo nel patterning del SN, senza contare che gli occhi derivano dell’estroflessione di due vescicole del diencefalo quindi sono, a tutti gli effetti, una porzione di encefalo dislocata in periferia.
Sia Pax6 che Rx1 codificano per due fattori di trascrizione che appartengono ai EFTFs (Eye Field homeobox Transcription Factors ) .
Pax6 è un gene master per lo sviluppo dell’occhio e suoi omologhi sono espressi nell’occhio in sviluppo di tutti i phyla del regno animale: insetti (Drosophila), platelminti (Dugesia), nemertea (Lineus), anfibi (Xenopus) e mammiferi (Mus, Homo) (fonti Loosli et al., 1996; Mastick et al., 1997). La mutazione di questi ortologhi provoca la mancata formazione dell’occhio (Chow et al., 1999; Glaser et al., 1992).
Oltre ad essere necessario Pax6 è anche sufficiente alla formazione dell’occhio.
Quando, nel laboratorio di W. Gehring (Halder et al., 1995), si è fatto esprimere eyeless, l’omologo di Pax6 ,ectopicamente in drosofile transgeniche,si sono formati occhi sovrannumerari sulle antenne, zampe, ali.
Inoltre, anche nei Vertebrati in particolari condizioni Pax6 è sufficiente, come in Drosophila, a dare origine ad un occhio: quando sovraespresso nell'embrione di
Xenopus, causa la comparsa di occhi ectopici, piccoli ma ben formati, nella larva (Ali Hemmati-Brivanlou, 1999).
La funzione di questo gene è molto conservata nell’evoluzione: espressione di ortologhi di Pax6 di medusa, calamaro e topo in Drosophila porta a formazione di occhi ectopici (Tomarev et al 1997) .
Rx1 è richiesto per lo sviluppo dell’occhio e del proencefalo e nel controllo della proliferazione e della neurogenesi della piastra neurale anteriore.
Xrx1 è espresso nella piastra neurale anteriore ed in seguito nelle strutture neurali dell’occhio in sviluppo (retina neurale ed epitelio pigmentato), ed in altre strutture del proencefalo che derivano dalla piastra neurale anteriore: nella ghiandola pineale, durante il suo sviluppo, nel diencefalo e nell’ipofisi. Il suo limite rostrale corrisponde alla cresta chiasmatica (chiasmatic ridge) che alcuni autori ritengono il limite più anteriore del tubo neurale ; quindi Xrx1 può rappresentare uno dei geni con omeodominio espressi più anteriormente. Inoltre la sua espressione in organi coinvolti nello stabilire i ritmi circadiani ( retina e ghiandola pineale) suggerisce per Xrx1 un ruolo nel controllo genico di questa funzione (Casarosa et al 1997).
Sox2 è un marcatore pan neurale precoce ed appartiene alla sottofamiglia di attivatori trascrizionali SoxB1 (Penzel et al., 1997). Sox2 inizia ad essere espresso a seguito dell’induzione neurale a livello del neuroectoderma (Nitta et al. 2006).
La regione regolatoria di Sox2 ha un modulo regolatorio conservato nelle diverse specie, e nel pollo questo modulo risponde a sia a FGF che a Wnt e porta alla sua accensione nella porzione posteriore della piastra neurale (Takemoto et al., 2006). Sox2 è espresso nella retina; il topo mutante Sox2 -/- presenta diminuzione della neurogenesi (Ferri et al., 2004), e la sua mutazione nell’essere umano porta ad anoftalmia (Fantes et al., 2003).
La n tubulina è una tubulina specifica del tessuto neurale. In Xenopus è espressa fin dallo stadio di blastula ( stadio 9) in tutto l’ectoderma salvo poi, a partire dallo stadio 12 (neurula), restringere il suo dominio all’ectoderma dorsale. Con il procedere dello sviluppo, la sua espressione segue il differenziamento delle cellule nervose; infatti a stadio di neurula avanzata il segnale è diffuso nella parte anteriore della piastra neurale, mentre posteriormente identifica sei cordoni di cellule nervose, tre per lato, che corrispondono, in senso mediale-laterale, ai progenitori primari dei motoneuroni, degli interneuroni e dei neuroni sensori (Oschwald et al. 1991).
4.2 Microiniezione del costrutto Xbh1-wt-HA
Per lo studio del ruolo di Xbh1 sul patterning del sistema nervoso, le microiniezioni sono state tutte effettuate, in embrioni a 4 cellule, a livello di uno dei blastomeri dorsali nella porzione animale. Questa è la regione destinata a dare il sistema nervoso.
In una prima fase del progetto si era deciso di usare i costrutti Xbh1 wild type e Xbh1 mutante per i due siti canonici di sumoilazione (doppio mutante) che al N-terminale della sequenza proteica del fattore di trascrizione recano l’epitopo
dell’emoagglutinina umana HA (rispettivamente denominati wt-HA e Xbh1-DM-HA); questa scelta era dovuta alla disponibilità immediata dei costrutti wild type e dei mutanti non sumoilabili preparati per i saggi biochimici, nell’idea che avessero identica attività biologica rispetto ai costrutti privi di epitopo. Gli effetti biologici ottenuti coi costrutti con epitopo sono essenzialmente gli stessi di quelli ottenuti coi costrutti senza epitopo e saranno descritti nel paragrafo che tratta dei risultati delle ISH fatte su embrioni iniettati con i costrutti senza epitopo.
Tuttavia confrontando i risultati delle ISH fatte su embrioni iniettati con i costrutti wild type con e senza epitopo HA, è stato rilevato come l’epitopo in qualche modo riducesse la funzione biologica della proteina consistente nella riduzione di vari geni marcatori La frequenza degli embrioni in cui l’espressione genica dei marcatori considerati è infatti inferiore quando vengono utilizzati costrutti con l’epitopo rispetto a quelli senza (Tabella 4.1 e 4.2 e grafico 4.1).
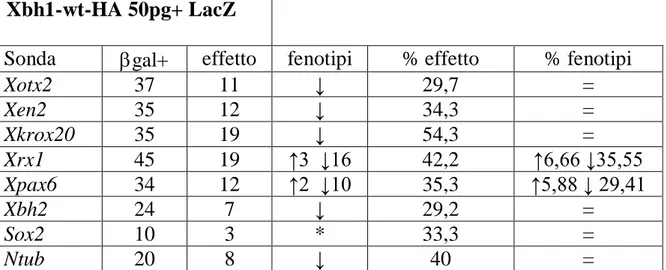
Xbh1-wt-HA 50pg+ LacZ
Sonda gal+ effetto fenotipi % effetto % fenotipi
Xotx2 37 11 ↓ 29,7 = Xen2 35 12 ↓ 34,3 = Xkrox20 35 19 ↓ 54,3 = Xrx1 45 19 ↑3 ↓16 42,2 ↑6,66 ↓35,55 Xpax6 34 12 ↑2 ↓10 35,3 ↑5,88 ↓ 29,41 Xbh2 24 7 ↓ 29,2 = Sox2 10 3 * 33,3 = Ntub 20 8 ↓ 40 =
Tabella 4.1- Risultati delle ibridazioni in situ per gli embrioni iniettati con 50 pg di Xbh1-wt-HA +
LacZ. Con effetto si intende una variazione dell’espressione del gene rispetto al lato non iniettato
che è il controllo interno dell’esperimento. Le frecce nella quarta e nella sesta colonna indicano diminuzione del segnale fino alla scomparsa ( ↓ ) oppure aumento del segnale ( ↑ ) del marcatore dal lato iniettato. Il simbolo * sta ad indicare che Sox2 presenta un variazione dell’espressione dal lato iniettato diversa da quella degli altri marcatori e che sarà descritta in seguito.
Grafico 4.1- Visualizzazione per istogammi delle percentuali di embrioni iniettati col costrutto wt senza epitopo (Xbh1-wt) o con epitopo (Xbh1-wt-HA) che hanno variazioni del pattern di espressione dei geni indicati dal lato iniettato.
Per non avere dei falsi negativi dovuti alla minore attività biologica conseguente all’epitopo HA piuttosto che alla alterazione dei siti di sumoilazione (soprattutto per quanto riguarda l’iniezione dei costrutti mutanti) si è ricorsi a costrutti privi di questo epitopo (Xbh1-wt e Xbh1-DM).
Durante gli esperimenti sono stati iniettati i seguenti RNA messaggeri: -300 pg LacZ
-250 pg LacZ + 50 pg Xbh1-wt -250 pg LacZ + 50 pg Xbh1-DM -250 pg LacZ + 5 pg Xbh1-wt
4.3 Mutagenesi sito diretta
Poiché i costrutti con l’epitopo HA mostravano una attività sensibilmente inferiore a quelli senza epitopo, è stato necessario ottenere i costrutti mutanti non sumoilabili senza epitopo.
Per questo scopo, a partire dal doppio mutante già impiegato negli esperimenti di lipofezione in piastra neurale (vedi capitolo1 paragrafo 5), ho prodotto il triplo mutante (K→ R 148, 161, 172) e poi il quadruplo mutante ( K→R 148, 161, 172,
189). Entrambi sono i costrutti sono stati trascritti e saranno utilizzati per microiniezioni.
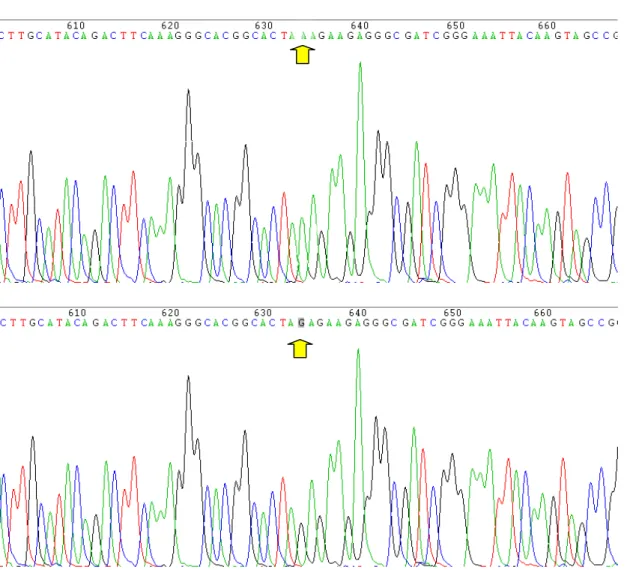
Nella figura 4.1 è mostrato l’elettroferogramma della sequenza di Xbh1 prima e dopo la mutagenesi sito diretta.
Fig. 4.1- Elettroferogrammi della Sequenza di Xbh1 prima, in alto, e dopo, in basso, la mutagenesi sito diretta che ha trasformato la Lisina 189 in Arginina. Le frecce indicano la base mutagenizzata.
4.4 Risultato delle ibridazioni in situ whole mount sugli embrioni iniettati con costrutti Xbh1-wt e Xbh1-DM
Dagli esperimenti di ISH è emerso un effetto generalmente di repressione di Xbh1 su geni marcatori presi in esame, in accordo con la sua funzione di repressore trascrizionale, o di spostamento del dominio di espressione in senso posteriore, con l’eccezione di Sox2 e, in misura minore, di Xrx1 e Xpax6.
I dati relativi agli esperimenti di ISH effettuati su embrioni iniettati con il costrutto Xbh1-wt sono raccolti nella tabella 4.2
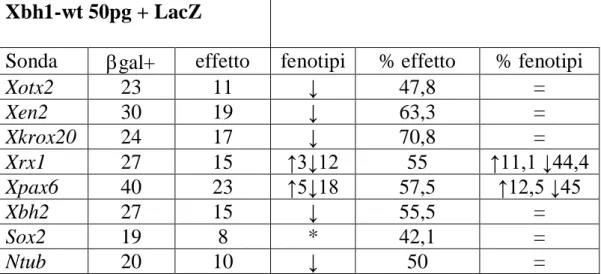
Xbh1-wt 50pg + LacZ
Sonda gal+ effetto fenotipi % effetto % fenotipi
Xotx2 23 11 ↓ 47,8 = Xen2 30 19 ↓ 63,3 = Xkrox20 24 17 ↓ 70,8 = Xrx1 27 15 ↑3↓12 55 ↑11,1 ↓44,4 Xpax6 40 23 ↑5↓18 57,5 ↑12,5 ↓45 Xbh2 27 15 ↓ 55,5 = Sox2 19 8 * 42,1 = Ntub 20 10 ↓ 50 =
Tabella 4.2- Risultati delle ibridazioni in situ per gli embrioni iniettati con 50 pg di Xbh1-wt +
LacZ. Le frecce e il simbolo * hanno il significato descritto in tabella 4.1
Un problema è sorto con i marcatori pan-neurali, in quanto il segnale della sonda spesso maschera il segnale sottostante della galattosidasi portando a scartare dalla casistica embrioni che mostrano un effetto su un solo lato della piastra neurale, ma di cui non si poteva accertare che fossero stati iniettati. Per quanto riguarda gli embrioni iniettati con il costrutto mutante sui due siti canonici di sumoilazione (Xbh1-DM), i risultati delle ISH sono stati diversi dall’atteso.
Visti i risultati della lipofezione dei progenitori retinici col doppio mutante (che perde l’effetto biologico di promozione del destino gangliare tipico di Xbh1-wt), ci si aspettava che anche l’effetto sui geni marcatori neurali fosse minore. In realtà è stato assai simile all’effetto del costrutto Xbh1-wt (vedi tabella 4.3).
Tabella 4.3- Risultati delle ibridazioni in situ per gli embrioni iniettati con 50 pg di Xbh1 DM senza epitopo + LacZ. Le frecce e il simbolo * hanno il significato descritto in tabella 4.1
Come controllo, embrioni iniettati solo con LacZ, la cui espressione è ininfluente sulle cellule e sullo sviluppo embrionale, sono stati usati i esperimenti simili di ISH (vedi tabella 4.4)
Xbh1-DM 50 pg + LacZ
Sonda gal+ Effetto fenotipi % effetto % fenotipi
Xotx2 30 19 ↓ 66,6 = Xen2 25 15 ↓ 60 = Xkrox20 25 19 ↓ 76 = Xrx1 30 22 ↑3 ↓19 73,3 ↑10 ↓63,3 Xpax6 32 25 ↑5 ↓20 78,1 ↑15,65 ↓62,5 Xbh2 30 18 ↓ 60 = Sox2 16 9 * 56,25 = Ntub 23 15 ↓ 65,2 =
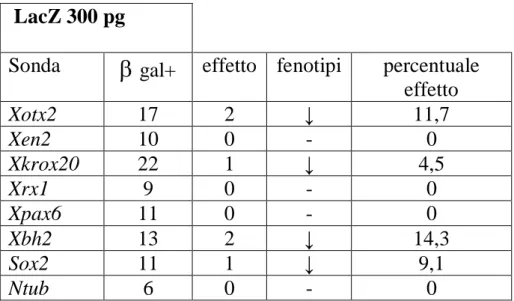
LacZ 300 pg
Sonda
gal+ effetto fenotipi percentuale effetto Xotx2 17 2 ↓ 11,7 Xen2 10 0 - 0 Xkrox20 22 1 ↓ 4,5 Xrx1 9 0 - 0 Xpax6 11 0 - 0 Xbh2 13 2 ↓ 14,3 Sox2 11 1 ↓ 9,1 Ntub 6 0 - 0Tabella 4.4- Risultati delle ibridazioni in situ per gli embrioni iniettati con 300 pg di LacZ. Le frecce hanno il significato descritto in tabella 4.1
Grafico 4.2- Visualizzazione per istogammi delle percentuali di embrioni iniettati col costrutto
Xbh1-wt, Xbh1-DM e solo con LacZ che hanno variazioni del pattern dal lato iniettato.
Di seguito sono inserite due figure che offrono una panoramica generale sugli effetti della sovraespressione di Xbh1-wt e Xbh1-DM sui marcatori presi in esame.
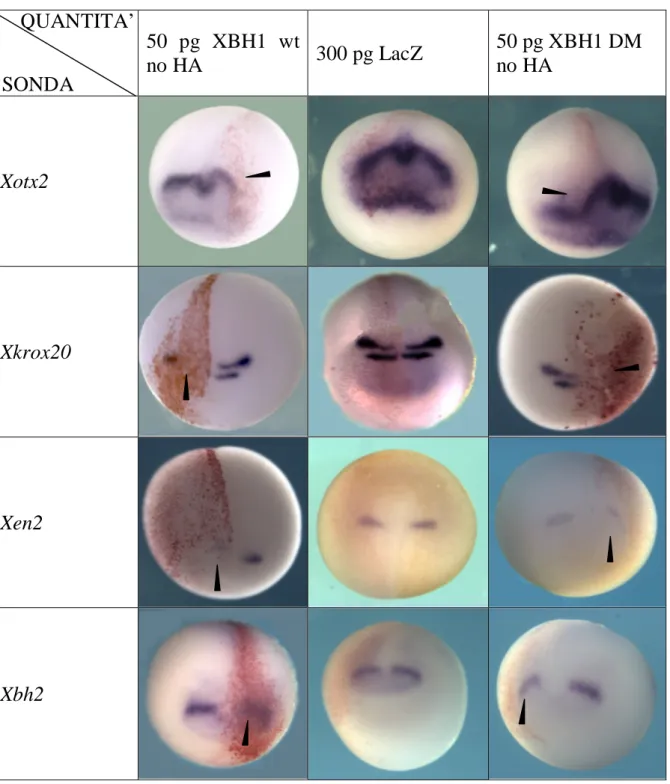
50 pg XBH1 wt no HA 300 pg LacZ 50 pg XBH1 DM no HA Xotx2 Xkrox20 Xen2 Xbh2
Fig. 4.2- Fotografie di embrioni sottoposti a ISH whole mount; in colonna il costrutto con cui sono stati iniettati in orizzontale il gene di cui è stata analizzata l’espressione. Le punte di freccia indicano la zona dell’embrione in cui è visibile una variazione del dominio di espressione del gene in esame
SONDA
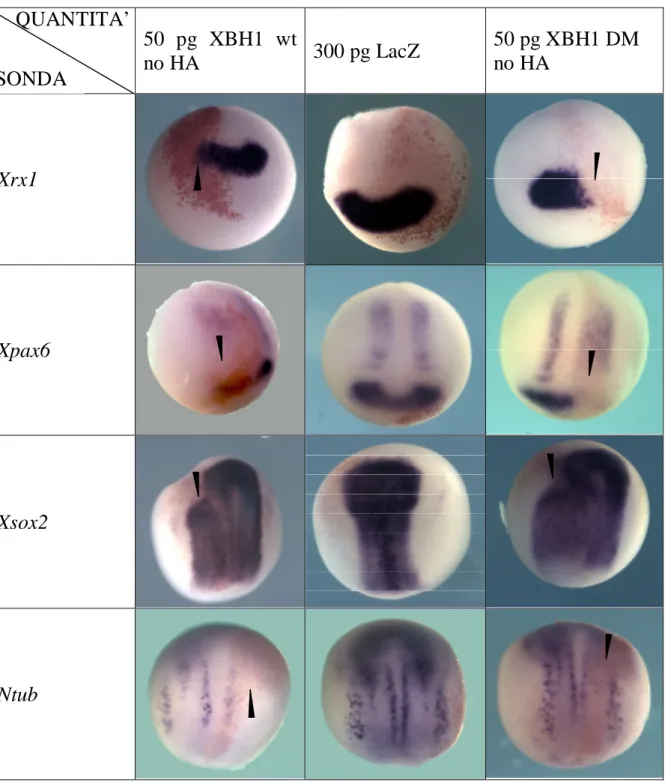
50 pg XBH1 wt no HA 300 pg LacZ 50 pg XBH1 DM no HA Xrx1 Xpax6 Xsox2 Ntub
Fig 4.3- Fotografie di embrioni sottoposti a ISH whole mount; in colonna il costrutto con cui sono stati iniettati in orizzontale il gene di cui è stata analizzata l’espressione. Le punte di freccia indicano la zona dell’embrione in cui è visibile una variazione del dominio di espressione del gene in esame.
Per quanto riguarda i geni marcatori del campo ottico, nelle tabelle sono state inserite immagini relative alla soppressione di questi geni nel lato iniettato, che è l’effetto più frequentemente osservato.
Esaminerò adesso i casi in cui si è riscontrato aumento del loro dominio di espressione e le possibili cause di esso nel capitolo successivo.
SONDA
Le modificazioni del pattern dei marcatori del campo ottico presentano una variabilità che non è stata registrata per gli altri marcatori.
Una percentuale che oscilla tra il 10 e il 20% degli embrioni gal positivi iniettati tanto col costrutto wild type quanto col doppio mutante, mostra un aumento del segnale in corrispondenza del lato iniettato ( vedi Fig 4.4) .
Fig. 4.4- Risultato di ISH whole mount su embrioni iniettati con Xbh1-DM per i geni Xrx1 ( A) e
Xpax6 ( B)
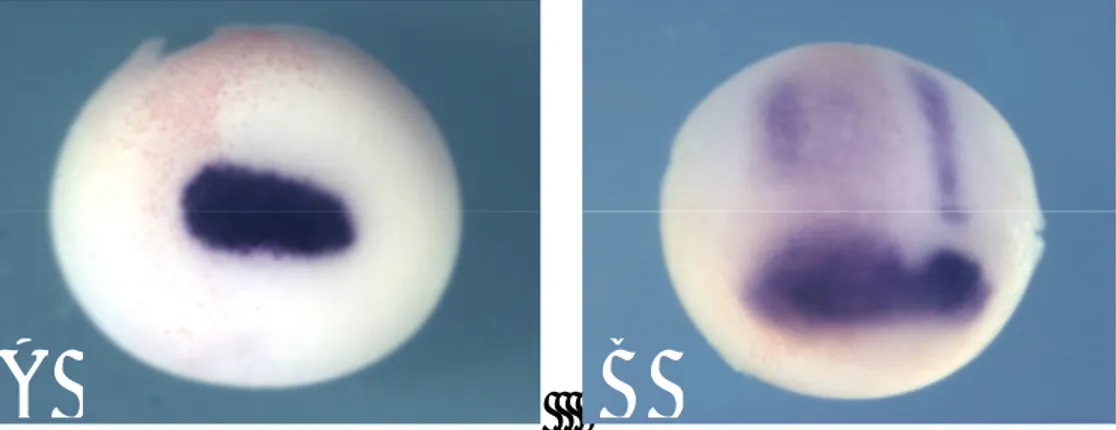
Per quanto riguarda Xsox2 dal lato iniettato si riscontra un aumento del dominio di espressione posteriore ( vedi Fig. 4.5) l’opposto del risultato ottenuto da Offner et al. ( 2005) per un altro gene appartenente al gruppo dei fattori SoxB1 Xsox3. Dei motivi di questa modificazione del pattern tratterò nel prossimo capitolo.
Fig. 4.5- A Embrione iniettato con 50 pg Xbh1-wt e sottoposto a ISH whole mount per il gene
Xsox2. La punta della freccia indica il punto dove c’è soppressione del segnale, la linea tratteggiata
indica il solco mediano della piastra neurale e le parentesi graffe sottolineano l’ampiezza dei domini di espressione del gene dal lato iniettato , dx, e dal lato di controllo, sin. B Risultato di whole mount
ISH per il gene Xsox3 su embrione di X. laevis iniettato monolateralmente con 50 pg di Mbarhl2 (
omologo murino di Xbh1) risultati simili erano stati ottenuti anche con l’iniezione del gene di
Xenopus. Da Offner et al. 2005.
B
B
A
B
4.5 Risultati del saggio di TUNEL whole mount
Visto il ruolo che ha Xbh1 nell’indurre apoptosi nelle cellule mesodermiche e della piastra neurale (Offner et al. 2005)si sono effettuati saggi TUNEL su embrioni iniettati con LacZ , per controllo, con LacZ + Xbh1-wt e con LacZ + Xbh1-DM per verificare se la sumoilazione regolasse questa funzione della proteina.
Così come non si sono riscontrate differenze sostanziali per i risultati delle ISH tra gli embrioni iniettati con il costrutto wild type e quello mutato, così nessuna differenza è stata rilevata con il saggio di TUNEL.
Costrutto Dose gal+ Effetto Percentuale effetto
LacZ 300 pg 15 2 13,3 %
Xbh1 wt+LacZ 50pg + 250 pg 25 10 40% Xbh1 DM+LacZ 50pg + 250 pg 30 15 50%
Tabella 4.7- Risultati del saggio di TUNEL sugli embrioni iniettati solo con LacZ, con Xbh1-wt e con Xbh1-DM. In questo caso con “effetto” si intende un numero maggiore , di cellule andate incontro ad apoptosi, nel lato iniettato rispetto a quello non iniettato.
50 pg XBH1 wt
no HA 300 pg LacZ
50 pg XBH1 DM no HA
TUNEL
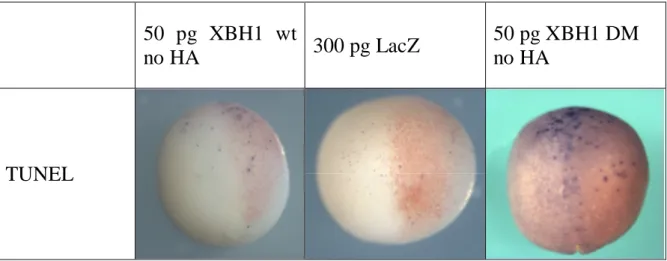
Fig. 4.6 - Fotografie di embrioni iniettati con diversi costrutti e sottoposti a saggio TUNEL.
4.6 Analisi delle conseguenze dell’overespressione di Xbh1-wt a stadi di larva natante
Per verificare, se l’effetto di modulazione del patterning della piastra neurale avesse ripercussioni sullo sviluppo tardivo degli embrioni, una parte degli embrioni iniettati unilateralmente con il costrutto Xbh1-wt nella dose maggiore è stata fatta sviluppare fino a stadio 37/38.
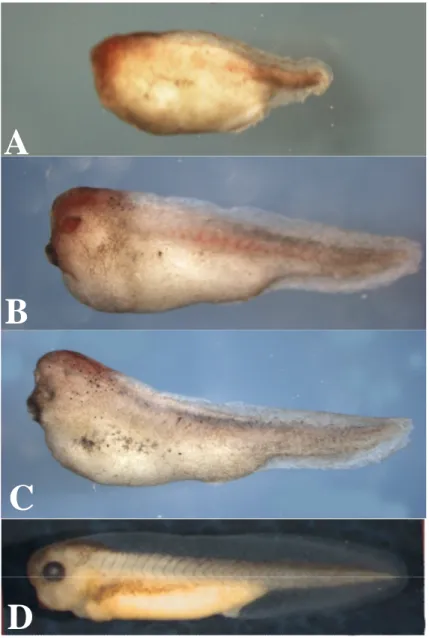
L’effetto della sovrespressione del gene ha portato nel 30 % dei casi ( n=40) a ventralizzazione (vedi Fig. 4.7 A,B,C). Questo effetto era già stato osservato da Offner et al. ( 2005), anche se nel loro esperimento l’iniezione era bilaterale e quindi
il fenotipo era più frequente; esso è collegato al fatto che le cellule che subiscono apoptosi esprimono chordin e quindi una sovrespressione di Xbh1 comporta un eccesso di BMP e di conseguenza ventralizzazione (Gamse e Sive, 2000), questo fenotipo anche in embrioni iniettati monolateralmente è probabilmente dovuto a una diffusione dell’mRNA su entrambi i lati dell’embrione.
Fig. 4.7-A, B e C Fotografie di tre embrioni di X. laevis a stadio 37/38 ventralizzati in seguito all’iniezione di 50 pg di Xbh1 wt + LacZ . D Embrione di X. laevis a stadio 37/38 non iniettato.
Nel resto degli embrioni gli effetti sul patterning e sullo sviluppo dell’occhio erano minimi ( vedi Fig. 4.8 e 4.9) segno che probabilmente col progredire dello sviluppo si instaura un meccanismo di recupero del fenotipo.
A
B
C
Fig. 4.8- fotografia di entrambi i lati di un embrione iniettato con 50 pg Xbh1-wt . A: lato iniettato che presenta un occhio di dimensioni lievemente minori. B: lato non iniettato.
Fig. 4.9- fotografia di entrambi i lati di un embrione iniettato con 50 pg Xbh1-wt . A: lato iniettato che presenta un occhio di dimensioni lievemente maggiori. B: lato non iniettato.