1.1 Introduzione
In questo capitolo verrà descritto brevemente il sistema nervoso che, come ha affermato Brodal nel 1969, è considerato come: “a multitude of minor units, each with is particular structural organization, specific as regards its finer intrinsic organization as well as its connections with other units”.
1.2 Caratteristiche del sistema nervoso
Il sistema nervoso costituisce il più perfetto meccanismo che regola la vita di relazione dell’animale: è un insieme di reti di comunicazione che consentono ad un organismo di interagire in modo appropriato con l’ambiente. [1, 2, 3]
Il sistema nervoso è provvisto di apparati sensoriali, atti a rilevare gli stimoli che si verificano nell’ambiente, di apparati di integrazione, che elaborano i dati sensoriali confrontandoli con le informazioni depositate nella memoria, e di apparati motori, che promuovono i movimenti e altri tipi di attività.
Il sistema nervoso viene diviso anatomicamente in due parti: il sistema nervoso centrale e il sistema nervoso periferico.
Il sistema nervoso centrale (SNC) è responsabile dell'integrazione, dell’analisi e della coordinazione dei dati sensoriali e dei comandi
motori. E' anche la sede di funzioni più importanti quali l'intelligenza, la memoria, l'apprendimento e le emozioni.
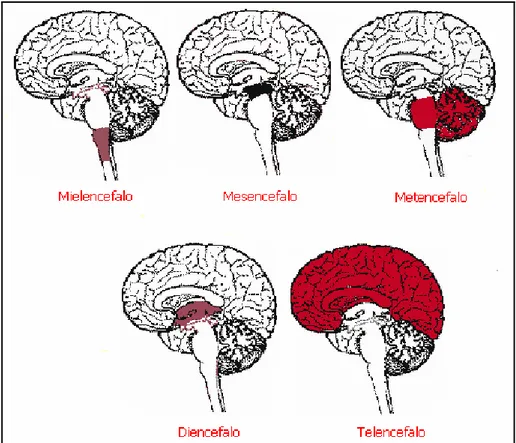
E’ costituito dall'encefalo, racchiuso nella scatola cranica, e dal midollo spinale, contenuto nel canale vertebrale. L’encefalo è ulteriormente divisibile, in base allo sviluppo embriologico, in cinque distretti o regioni: mielencefalo, mesencefalo, metencefalo, diencefalo e telencefalo (figura 1.1).
Figura 1.1 I 5 distretti dell’encefalo
Il sistema nervoso periferico (SNP) è costituito da tutto il tessuto nervoso al di fuori del SNC e costituisce una vera e propria interfaccia tra il SNC e l’ambiente, intendendo con il termine ambiente sia il mondo esterno sia le parti dell’organismo che sono
anatomicamente separate dal sistema nervoso. Svolge essenzialmente la funzione di trasmissione del segnale attraverso fasci di conduzione. I segnali, afferenti da un'unità periferica (organo) o in uscita (efferenti) verso un'unità periferica, decorrono in fibre separate (assoni) che generalmente sono raggruppate in un fascio di conduzione unitario (nervo).
Il sistema nervoso periferico si suddivide in due parti principali: il sistema nervoso somatico ed il sistema nervoso autonomo. Il primo innerva la muscolatura scheletrica ed è controllabile dalla nostra volontà; il secondo innerva visceri, vasi sanguigni e ghiandole e funziona in modo autonomo.
Il tessuto nervoso è composto da tre elementi fondamentali:
1. le cellule nervose o neuroni, che ne costituiscono le unità funzionali;
2. le cellule non nervose o gliali, i cui tipi principali sono gli astrociti, gli oligodendrociti e la microglia nel sistema nervoso centrale, e le cellule di Schwann nel sistema nervoso periferico; 3. il tessuto connettivo fibroso e i vasi venosi e arteriosi.
1.3 I neuroni
I neuroni sono cellule eccitabili altamente specializzate, in grado di ricevere, elaborare e trasmettere informazioni alle cellule adiacenti per mezzo di impulsi nervosi (potenziali d'azione o spike).
In rapporto a queste funzioni di comunicazione, nella cellula nervosa sono notevolmente sviluppate due proprietà: l’irritabilità e la conducibilità. La prima è definita come la capacità di reagire agli stimoli provenienti dall’ambiente esterno e interno trasformandoli
in impulsi nervosi. La seconda è la capacità di trasmettere i segnali ad altre parti della stessa cellula nervosa o ad altri neuroni o alle cellule epiteliali, muscolari, ecc.
1.3.1 Anatomia microscopica del neurone
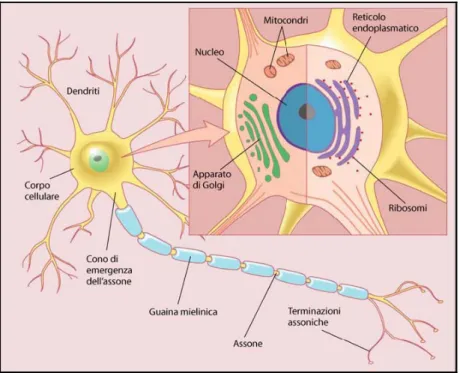
Ogni neurone è costituito da un corpo cellulare, detto pirenoforo o soma, e da lunghi prolungamenti di due tipi: i dendriti e l’assone o neurite (figura 1.2).
Figura 1.2 Principali componenti di un neurone
Una caratteristica comune di tutti i neuroni è una precisa polarizzazione nella conduzione dello stimolo, che viaggia sempre in una sola direzione:
I dendriti costituiscono il polo recettore del neurone e l’assone il polo trasmittente, mentre il pirenoforo costituisce il centro di elaborazione e integrazione dei messaggi in ingresso (input) provenienti da moltissime cellule, che concretizzerà in un messaggio in uscita (output).
Il soma di ogni neurone contiene il nucleo e il nucleolo. All’interno del nucleo, ricoperto da membrana, si trova il materiale genetico (cromosomi) che conserva le informazioni necessarie allo sviluppo della cellula ed alla sintesi proteica, indispensabile per il mantenimento e la sopravvivenza della cellula stessa. Il nucleolo produce i ribosomi, necessari per la translazione delle informazioni genetiche nelle proteine.
Il soma é fornito di un apparato biosintetico ben sviluppato che produce i costituenti di membrana, gli enzimi di sintesi e altre sostanze chimiche per le funzioni specializzate delle cellule nervose. L’apparato biosinetico include i corpi di Nissl, il reticolo endoplasmatico e l’apparato di Golgi. I corpi di Nissl sono gruppi di ribosomi usati per la sintesi proteica. Il reticolo endoplasmatico (RE) è un sistema di vescicole utilizzate per trasportare materiale nel citoplasma. Può essere dotato di ribosomi (RE rugoso) o meno (RE liscio). Quando vi sono i ribosomi, il RE è importante per la sintesi proteica.
L’Apparato di Golgi è una struttura costituita da membrane, importante per l'immagazzinamento di peptidi e proteine (compresi i neurotrasmettitori) in vescicole.
Il soma è inoltre dotato di numerosi mitocondri, che producono energia in forma di molecole di ATP, e di elementi del citosheletro: i neurofilamenti e i microtubuli.
I dendriti sono ramificazioni a breve decorso, che si legano al corpo del neurone in modo indistinto e possono avere più ramificazioni successive. Assieme al pirenoforo svolgono funzioni inerenti l’irritabilità, in quanto su di essi terminano i prolungamenti di altre cellule nervose in regioni di contatto specializzate, dette sinapsi. L’assone è un prolungamento in generale unico, di forma cilindrica, più lungo del dendrita. Il suo diametro, nell’uomo, può variare da un minimo di 1 μm ad un limite superiore di circa 20 μm, mentre raggiunge un massimo di 500 μm per l’assone gigante di calamaro. Esso si diparte dal corpo cellulare in corrispondenza di una protuberanza detta collo di emergenza dell’assone, prosegue restringendosi per pochi micron, formando il segmento iniziale, ed infine si estende con forma cilindrica quasi regolare fino alla ramificazione terminale, formando le sinapsi.
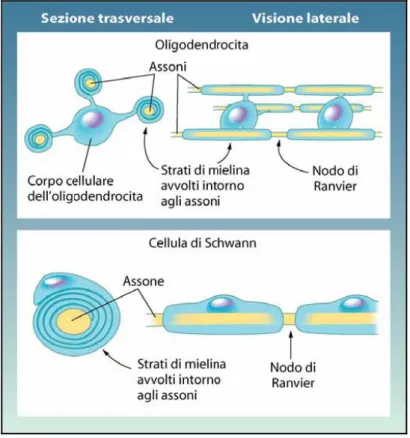
Gli assoni possono essere rivestiti da guaine, oppure esserne privi. Il rivestimento è originato dalle cellule di Schwann: il citoplasma di queste cellule produce una sostanza biancastra di natura fosfolipidica, detta mielina, che avvolge l’assone come una guaina dalla sua emergenza dal pirenoforo fino allo sfioccamento finale che resta nudo; la guaina è però interrotta lungo l’assone in vari punti detti nodi di Ranvier, che hanno molta importanza nella conduzione dell’impulso nervoso, e che rappresentano il punto di confine tra una cellula di Schwann e la successiva.
Per essere più precisi, le cellule di Schwann formano il rivestimento negli assoni del SNP; gli assoni che restano nell’ambito del SNC sono rivestiti in modo analogo dagli oligodendrociti. La differenza tra i due tipi di cellule consiste nel fatto che un oligodendrocita può contribuire contemporaneamente al rivestimento di più assoni.
Figura 1.3 Neuroni mielinati
1.3.2 Classificazione dei neuroni
I neuroni possono essere classificati in base a diversi criteri: la morfologia, la lunghezza e la funzione. [4]
I. La classificazione morfologica del neurone si basa sul numero di prolungamenti: si parla così di neuroni monopolari, bipolari o multipolari (figura 1.4).
Figura 1.4 Tipi di neuroni in base alla forma
I neuroni monopolari sono provvisti di un solo prolungamento e sono presenti nei mammiferi solo nella vita embrionale.
I neuroni dei gangli spinali sono chiamati pseudounipolari perché durante lo sviluppo i due prolungamenti di cui sono provvisti migrano lungo il pirenoforo e si avvicinano fino ad unirsi, costituendo pertano un unico prolungamento il quale, a distanza variabile dal pirenoforo, si divide a T ed emette due collaterali di cui una è diretta alla periferia e l’altra è diretta verso il sistema nervoso centrale.
I neuroni bipolari hanno due prolungamenti con funzione diversa: uno convoglia i segnali verso il soma (segnali di "input" alla cellula) e l'altro conduce gli impulsi generati dalla cellula stessa ("output" del neurone). Tali neuroni sono presenti nella retina e nell'epitelio olfattivo.
I neuroni multipolari sono i più rappresentati nel sistema nervoso e sono caratterizzati da un pirenoforo di forma variabile, da un singolo assone e da un gran numero di dendriti.
II. In base alla lunghezza dell’assone i neuroni possono essere divisi in neuroni del tipo I e II di Golgi .
I neuroni del I tipo di Golgi sono caratterizzati da un assone molto lungo che può decorrere per tratti variabili nel sistema nervoso centrale prima di mettersi in sinapsi con altri neuroni oppure prima di abbandonare il sistema nervoso centrale per portarsi alla periferia e mettersi in giunzione con cellule di tipo diverso.
I neuroni del II tipo di Golgi sono invece provvisti di un assone relativamente corto che non abbandona il sistema nervoso centrale e che, anche all’interno del sistema nervoso centrale, si mantiene all’interno dell’organo al quale il neurone stesso appartiene.
III. In base alla funzione si parla di neuroni sensoriali,
motoneuroni e interneuroni (figura 1.5).
I neuroni sensoriali rilevano i cambiamenti nell’ambiente interno o esterno e inviano questi segnali al sistema nervoso centrale.
I motoneuroni controllano la contrazione muscolare e la secrezione ghiandolare.
Gli interneuroni sono localizzati completamente all’interno del SNC.
Gli interneuroni ad assone lungo (neuroni di proiezione) trasmettono informazioni su lunghe distanze, da una regione cerebrale all’altra; quelli ad assone corto elaborano le informazioni
a livello di circuiti locali. Circuiti composti da interneuroni sono responsabili di percezione, apprendimento, memoria e processi decisionali.
Figura 1.5 Tipi di neuroni in base alla funzione
1.4 Le sinapsi
La sinapsi (dal greco "συν - άπτειν " = "connettere") è la regione attraverso la quale un impulso nervoso si trasmette da una cellula elettricamente eccitabile ad un’altra.
I contatti sinaptici possono essere situati sul soma cellulare, sui dendriti o sull’assone e le denominazioni delle sinapsi identificano le zone di contatto dei neuroni presinaptici con quelli postsinaptici (figura 1.6).
Figura 1.6 Tipi di sinapsi
La sinapsi asso-dendritica identifica una sinapsi che unisce l'assone del neurone portatore del messaggio ad un dendrita di un altro neurone; la sinapsi asso-assonica mette a contatto tra di loro due assoni e la sinapsi asso-somatica è una sinapsi tra l'assone di un neurone e il soma di un altro.
Esistono due tipi di sinapsi: le sinapsi elettriche e le sinapsi chimiche. Nei vertebrati superiori prevalgono le sinapsi di tipo chimico.
Nella sinapsi elettrica due cellule eccitabili sono tra loro connesse mediante una giunzione comunicante o gap junction.
Le gap junction consentono la comunicazione tra cellule per passaggio diretto di correnti ioniche da una cellula all’altra, quindi non si verifica ritardo sinaptico. In genere le sinapsi elettriche, al contrario di quelle chimiche, consentono la conduzione in entrambe le direzioni, ma esistono anche sinapsi elettriche che conducono preferenzialmente in una direzione piuttosto che nell’altra: questa proprietà prende il nome di rettificazione.
Le membrane plasmatiche delle cellule accoppiate sono separate da uno spazio inferiore a 3 nanometri. Le particelle intermembranarie delle gap junction sono costituite da 6 subunità, ciascuna formata da una singola proteina, la connessina. Le 6 subunità circondano un canale centrale e sono disposte ad esagono, formando una struttura chiamata "connessone". Attraverso i connessoni passano molecole idrosolubili e quindi il passaggio di ioni determina un passaggio di corrente elettrica. Cellule che sono tra loro elettricamente accoppiate possono essere disaccoppiate per chiusura dei canali dei connessoni.
Le sinapsi elettriche sono particolarmente adatte quando serve rapidità nella trasmissione del segnale (ad esempio nei circuiti che presiedono ai sistemi di fuga dei pesci e dei molluschi o nel sistema visivo dei vertebrati) e quando è richiesta la sincronizzazione nell’attività di più cellule (ad esempio nel muscolo cardiaco nel quale le fibre devono contrarsi in sintonia).
Nelle sinapsi chimiche, affinché l’impulso possa passare dal neurone presinaptico a quello postsinaptico, è necessario che la membrana di quest’ultimo venga preparata a “riceverlo”; a questo scopo il primo neurone secerne nello spazio sinaptico i neurotrasmettitori.
Figura 1.7 Schema di un neurone con sinapsi multiple
Vista la difficoltà di studiare le sinapsi nel folto intrico di neuroni cerebrali, l’attenzione degli studiosi si è rivolta per prima alla trasmissione dell’impulso dai motoneuroni al muscolo. Dato che quest’ultimo è eccitabile elettricamente in modo simile al neurone, la giunzione neuromuscolare o placca motrice rappresenta un ottimo modello per spiegare il funzionamento delle sinapsi in generale.
In prossimità della giunzione neuromuscolare, la fibra motrice perde il suo rivestimento di mielina e si divide in sottili ramificazioni terminali che si adagiano lungo una doccia sinaptica.
Nei terminali assonici sono presenti molte vescicole sinaptiche contenenti acetilcolina (ACh), il mediatore chimico della placca motrice, sintetizzata in periferia del neurone.
Quando il potenziale d'azione che si propaga lungo la fibra motrice giunge al terminale presinaptico, la depolarizzazione della membrana provoca l’apertura dei canali di calcio, voltaggio-dipendenti. La concentrazione extracellulare di Ca2+ è maggiore di quella interna e perciò il Ca2+ entra nella cellula seguendo il gradiente di concentrazione. Inoltre è attratto nello spazio intracellulare anche dalla polarità negativa della membrana: è spinto ad entrare da un doppio gradiente.
L’aumentata concentrazione di Ca2+ nel terminale provoca la fusione delle vescicole sinaptiche con la membrana plasmatica ed il conseguente svuotamento per esocitosi del loro contenuto in acetilcolina nel vallo sinaptico (figura 1.8).
Sulla membrana della cellula muscolare sono presenti recettori per ACh: si tratta di canali attivi che si aprono in seguito al legame. A differenza dei canali voltaggio-dipendenti questi canali sono aspecifici, cioè consentono il passaggio di ogni tipo di ione. All’apertura di questi recettori-canale il sodio (Na+) entra all’interno del muscolo, spinto sia dalla forza chimica che da quella elettrostatica, e il potassio (K+) esce fuori dal muscolo, spinto dalla forza chimica. Questo passaggio di ioni avviene contemporaneamente. La forza che spinge il Na+ ad entrare è maggiore di quella che spinge il K+ fuori dalla cellula e si ha perciò una depolarizzazione. Tale depolarizzazione è definita potenziale di placca o EPP e si tratta di un potenziale transitorio in quanto l’acetilcolina viene presto idrolizzata, ovvero scissa in colina ed acetato, dall’acetilcolinaesterasi, presente sulla membrana postgiunzionale.
Il potenziale elettrico di membrana (Em) a riposo del muscolo è –90
mV; dopo l’apertura dei recettori-canale ACh-dipendenti e il flusso di ioni, l'Em del muscolo diventa –40 mV. Questo valore è maggiore
del valore soglia, ma non scatena un potenziale d’azione (è un elettrotono), perché lungo la doccia sinaptica sulla superficie del sarcolemma non ci sono canali voltaggio-dipendenti per il Na+. Il potenziale di placca dunque, come tutti gli elettrotoni, è un potenziale locale, ma per potersi propagare lungo tutta la fibra muscolare deve essere trasformato in potenziale d’azione.
Le regioni del sarcolemma adiacenti alla doccia sinaptica hanno canali voltaggio-dipendenti per il Na+; tra la regione della placca (in cui Em = – 40 mV) e la regione contigua (in cui Em = – 90 mV) c’è
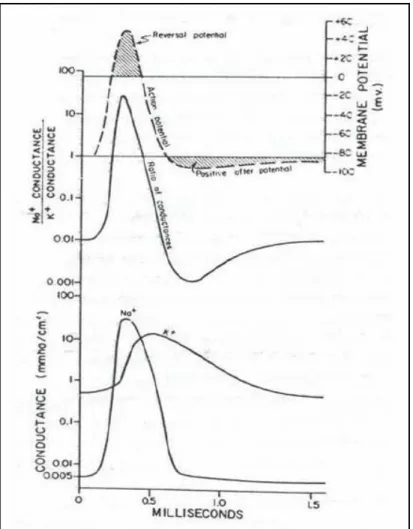
una differenza di potenziale, perciò si verifica uno spostamento di carica che forma un circuito di corrente capace di depolarizzare la regione di membrana su cui sono presenti i canali voltaggio-dipendenti per il Na+,generando un potenziale d’azione (figura 1.9).
Figura 1.9 Generazione dello spike
Il potenziale d’azione si propaga lungo tutto il muscolo provocandone la contrazione.
Il potenziale di placca ha sempre un valore sufficiente per scatenare il potenziale d’azione. Quindi questa è una sinapsi "1:1", cioè una sinapsi in cui il potenziale di placca scatena il potenziale d’azione.
Fra neuroni le sinapsi sono del tipo "molti:1", cioè occorrono più potenziali d’azione pre-sinaptici per scatenare un potenziale d’azione nel neurone post-sinaptico.
Infatti, in ogni istante, la cellula postsinaptica integra le varie afferenze, alcune delle quali possono anche essere iperpolarizzanti, ovvero inibitorie. Se la somma di tutti gli effetti di segno opposto provocati dalle varie afferenze è tale da depolarizzare la cellula fino al raggiungimento della soglia, essa genera il potenziale d’azione.
1.5 Lo sviluppo neurale
La sfida centrale dello sviluppo neurale è la spiegazione del modo in cui assoni e dendriti crescono, trovano i loro partner corretti e formano sinapsi selettivamente con essi per creare una rete funzionale. Il problema è formidabile: il cervello umano contiene più di 1011 neuroni, ciascuno dei quali, in media, stabilisce connessioni con un migliaio di altri, secondo un piano di collegamenti regolare e predicibile. [5]
Lo sviluppo del sistema nervoso procede in tre fasi, illustrate in figura 1.10.
Figura 1.10 Le tre fasi dello sviluppo neurale
La prima fase è lo sviluppo dei neuroni. I neuroni (e le cellule gliali che li accompagnano sempre) sono generati da precursori ectodermici e prima di emettere assoni e dendriti, spesso migrano dal punto in cui sono nati e si stabiliscono in altre posizioni. Neuroni nati in tempi e luoghi diversi, sotto il controllo di segnali induttivi o morfogeni, esprimono serie diversi di geni e le differenze di espressione genica ne modulano i caratteri e li aiutano a prendere connessioni con partner diversi.
La fase successiva comporta un tipo di morfogenesi che è una caratteristica unica del sistema nervoso: assoni e dendriti crescono lungo vie specifiche, stabilendo una rete di connessioni provvisoria ma ordinata tra le parti separate del sistema.
Strutture come la piastra del pavimento del midollo embrionale secernono sia chemioattraenti che chemiorepellenti, ai quali coni di crescita di classi diverse di neuroni rispondono in modo diverso.
Quando raggiungono la loro area bersaglio, gli assoni terminano selettivamente su una sottopopolazione delle cellule accessibili e in molte parti del sistema nervoso si formano mappe neurali, proiezioni ordinate di una serie di neuroni su un’altra.
L’ultima fase, che continua nella vita adulta, è la rifinitura delle connessioni sinaptiche.
Si verificano due tipi principali di aggiustamenti: eliminazione del surplus di neuroni mediante morte cellulare e eliminazione di sinapsi.
Più del 50% dei neuroni muoiono poco dopo aver raggiunto il bersaglio. Questa morte su larga scala è il risultato di una competizione. Ciascun tipo di cellula bersaglio rilascia una quantità limitata di un fattore neurotrofico, quale il fattore di crescita dei nervi o NGF, che i neuroni innervanti quel bersaglio richiedono per sopravvivere. I neuroni apparentemente competono per quel fattore e quelli che non ne ottengono abbastanza muoiono per morte cellulare programmata. Tale morte regola la quantità di innervazione secondo le dimensioni del bersaglio.
Dopo che il periodo di morte neuronale normale è terminato, inizia l’eliminazione delle sinapsi: singole sinapsi sono “potate” in alcuni punti, rafforzate in altri, in modo da creare uno schema di connessioni ordinato più precisamente. Tale processo dipende da attività elettrica: sinapsi che sono frequentemente attive sono rinforzate e neuroni diversi che contattano la stessa cellula bersaglio tendono a mantenere le loro sinapsi sul bersaglio in comune soltanto se di frequente attivi contemporaneamente (figura 1.11).
Figura 1.11 La modificazione delle sinapsi e la sua dipendenza da
attività elettrica
Tale fenomeno può essere riassunto nella frase “neuroni che sparano insieme si connettono insieme”.
La stessa regola dello sparo che correla mantenimento delle sinapsi e attività neurale aiuta a organizzare lo sviluppo del nostro cervello alla luce dell’esperienza.
1.6 Classificazione dei circuiti nervosi
Attraverso la rete di connessioni che si costituisce fluisce l’informazione, secondo circuiti che possono essere così classificati: [6]
¾ circuiti locali (Rakic, 1976), che connettono neuroni appartenenti alla medesima regione. Essi possono comprendere moduli elementari che agiscono come unità di integrazione, definiti da Shepherd nel 1978 microcircuiti;
¾ circuiti interregionali (Shepherd, 1988), che connettono tra loro regioni a diverso significato funzionale;
¾ sistemi, che connettono più circuiti interregionali in organizzazioni complesse individuate in rapporto a grandi funzioni. Esempi tipici sono rappresentati da sistemi che comprendono vie ascendenti (dalla periferia al centro) e discendenti (dal centro alla periferia), deputati a garantire risposte finalizzate complesse;
¾ sistemi distribuiti (Shepherd, 1988), che coordinano l’attività di molti circuiti interregionali e dei sistemi durante l’attuazione di comportamenti che coinvolgono l’intero organismo.
Caratteristica comune a tutti i tipi di circuito è la presenza di meccanismi di retroazione.
Il flusso dell’informazione lungo un determinato circuito avviene secondo una direzione (ortodromica), determinata dalle proprietà delle sinapsi (Cajal, 1933). Benché il neurite sia in grado di condurre un impulso nervoso indifferentemente dal soma alle terminazioni e dalle terminazioni al soma, quest’ultima direzione (antidromica) ha scarsa importanza fisiologica in quanto il segnale una volta giunto al soma non è comunque in grado di essere trasmesso a ritroso alle terminazioni presinaptiche delle fibre afferenti.
1.7 Le cellule gliali
Le cellule della glia sono state scoperte nel 1891 da Santiago Ramón y Cajal e il loro numero nel cervello supera di nove volte quello dei neuroni.
Nel SNC esistono 4 classi principali di cellule gliali: astrociti, oligodendrociti, cellule ependimali e microglia, ognuna con funzioni differenti. Nel SNP, invece, esistono le cellule di Schwann e le cellule satelliti (figura 1.12).
Figura 1.12 Alcuni tipi di cellule gliali
Gli astrociti sono piccole cellule stellate ubiquitarie che svolgono importanti funzioni e sono tutt'altro che semplici elementi di sostegno.
Sono forniti di numerose estroflessioni che ancorano i neuroni al loro rifornimento di sangue.
Regolano l'ambiente chimico esterno dei neuroni rimuovendo gli ioni, in particolare il potassio, e riciclano i neurotrasmettitori rilasciati durante la trasmissione sinaptica.
La teoria corrente sostiene che gli astrociti siano i "blocchi di costruzione" della barriera emato-encefalica e che siano in grado di
regolare la vasocostrizione e la vasodilatazione producendo sostanze come l'acido arachidonico, i cui metaboliti sono vasoattivi. Le ricerche degli ultimi anni hanno dimostrato che gli astrociti “ascoltano” le comunicazioni neuronali e “rispondono”, modulando così le loro funzioni. [7, 8, 9]
Nel 1996 Porter e McCarthy scoprirono che gli astrociti rispondono al potenziale d’azione dei neuroni per mezzo di recettori metabotropici di glutammato. Quando avviene la sinapsi, il glutammato, primo messaggero, è rilasciato dal neurone pre-sinaptico all’interno della fessura sinaptica che è limitata in parte dai recettori metabotropici di glutammato (mGluR) dell’astrocita sinaptico. Il glutammato si lega, in parte, a tali recettori e l’astrocita sinaptico produce e rilascia l’IP3, che agisce come un secondo messaggero ed è la chiave delle segnalazioni intracellulari di calcio. L’IP3 si lega ai recettori di IP3 situati sul reticolo endoplasmatico. Tali recettori sono dei canali ionici e rilasciano grandi quantità di calcio nel RE.
Tale rilascio di Ca2+ si può manifestare nella forma di onde intracellulari di Ca2+ e l’onda di calcio si propaga quindi attraverso la membrana cellulare nello spazio extracellulare agli astrociti adiacenti. In essi aumenta la concentrazione di calcio e ciò determina il rilascio di glutammato nello spazio extracellulare.
Il glutammato si propaga con l’onda di calcio extracellulare, e può modulare sinapsi vicine e lontane con meccanismi eccitatori o inibitori.
In figura 1.13 sono illustrati i processi sinaptici locali che coinvolgono il neurone pre-sinaptico, il neurone post-sinaptico e l’astrocita sinaptico. Si parla di sinapsi tripartita.
Figura 1.13 Sinapsi tripartita
Gli oligodendrociti sono responsabili della mielinizzazione dei neuroni del SNC. Ogni oligodendrocita fornisce la guaina mielinica a 3-50 fibre e ne circonda molte altre, senza però formare mielina. Le cellule ependimali, chiamate anche ependimociti delimitano le cavità del sistema nervoso centrale e, col battito delle ciglia, favoriscono la circolazione del liquido cerebrospinale. Esse costituiscono i "muri" che delimitano le varie sezioni. Sono privi di una membrana basale e continuano in prolungamenti e connessioni in continuità con gli astrociti. È stata ipotizzata una loro possibile
funzione di cellule staminali del tessuto nervoso e comunque sembra siano coinvolte nella rigenerazione del medesimo.
Le microglia sono macrofagi specializzati in grado di effettuare la fagocitosi e quindi di proteggere i neuroni del sistema nervoso centrale. Benché tecnicamente non siano cellule della glia in quanto derivano dai monociti e non dal tessuto ectodermico, sono categorizzate in questo modo per via del loro ruolo di supporto ai neuroni. Le microglia sono cellule relativamente piccole rispetto alle macroglia, hanno forma diversa e un nucleo oblungo. Si spostano all'interno del cervello, normalmente sono presenti in piccolo numero ma si moltiplicano in caso di danni al cervello.
Le cellule di Schwann formano la guaina mielinica degli assoni del sistema nervoso periferico. Hanno inoltre un'attività fagocitaria e ripuliscono dai residui cellulari permettendo la ricrescita dei neuroni del sistema nervoso periferico.
Le cellule satellite sono piccole cellule che delimitano la superficie esterna dei neuroni del sistema nervoso periferico ed aiutano a regolare l'ambiente chimico esterno.
Bibliografia
1. G.C. Baldoni, E. Brizzi; L. Comparini, “Anatomia umana”, vol.
3, ed. Emes
2. Robert M.Berne, Matthew N. Levy, “Principi di fisiologia”,
seconda edizione, casa editrice Ambrosiana
3. M. Pusceddu Nardella, G.Testoni, “Biologica”, Trevisini Editore
Milano
4. Prof. Danilo De Rossi, Dispense del corso di Modelli di sistemi
biologici, 2004-2005
5. Bruce Alberts, Alexander Johson, Julian Lewis, Martin Raff,
Keith Roberts, Peter Walter, “Biologia molecolare della cellula”, Quarta edizione, Zanichelli
6. Emanuele Biondi, Pietro Marasso; Vincenzo Tagliasco,
“Neuroscienze e scienze dell’artificiale: dal neurone all’intelligenza”, Patron editore
7. A. Araque, V. Parpura, R.P. Sanzgiri, P.G. Haydon, - Glutamate
dependent astrocytic modulation of synaptic transmission between cultured hippocampal neurons, Eur. Neurosi. 10 (1998), p.p. 229-2142
8. J.T. Porter, K.D. McCarthy, - Hippocampal astrocytes in situ
respond to glutamate released from synaptic terminals. The Journal of Neuroscience 16 (1996), p.p. 5073:5081
9. Suhita Nadkarni, Peter Jung, - Dressed neurons: modelling