2. PROTEINE CHE LEGANO LA SEQUENZA CCAAT: IL FATTORE
NUCLEARE NF-Y
Diverse proteine regolatrici della trascrizione che legano il DNA nella regione CCAAT sono state isolate e caratterizzate (Mantovani et al., 1992, Zorbas et al., 1992; Aufiero et al., 1994; Andres et al., 1994; Harada et al., 1995; Osada et al., 1996, Liberati et al., 1998, Mantovani 1998, 1999), ma tra queste il fattore nucleare NF-Y è il più direttamente e strettamente dipendente dall’integrità del pentanucleotide. Il fattore NF-Y (“Nuclear Factor-NF-Y”, chiamato anche CBF, α-CP1 o CPP1) è stato identificato per la prima volta come proteina che riconosce la sequenza Y (CCAAT) del promotore di classe II Ea del maggiore complesso di istocompatibilità, e nel lievito S. cerevisiae attraverso lo studio di mutanti con ridotti livelli di espressione del gene CYC1.
Nelle piante, rispetto a vertebrati e lieviti, il meccanismo funzionale del complesso NF-Y è poco conosciuto, ed è complicato dal fatto che le singole subunità sono codificate da una famiglia genica mentre in vertebrati e lieviti ogni subunità è codificata da un gene a singola copia. (Edwards et al., 1998, Gusmaroli et al., 2001, 2002).
Molte delle informazioni sulle proteine che attivano il CCAAT box provengono, infatti, da studi su lieviti: è dimostrato che NF-Y (chiamato anche CBF e in lievito HAP2/3/4/5) è un trimero composto da distinte subunità:
• NF-YA (anche detta CBF-B, HAP2 nel lievito), • NF-YB (anche detta CBF-A, HAP3 nel lievito ) e
• NF-YC (anche detta CBF-C, HAP5 nel lievito) (Fig. 2.1).
Tutte e tre le subunità contengono una parte evolutivamente conservata, che raggiunge una identità di sequenza aminoacidica superiore al 70% e una similarità superiore all’80% (Bellorini et al., 1997) e considerate specie più distanti. La regione conservata è differentemente
dislocata nelle diverse subunità: si ritrova nell’estremità C dell’NF-YA, nella parte centrale di NF-YB e nell’N-terminale di NF-YC. Nelle regioni adiacenti ai domini conservati, NF-YA e NF-YB hanno larghi domini ricchi in glutammina e residui idrofobici che si ritrovano nei mammiferi nel riccio di mare, nello Schisostoma mansoni ed in alcune piante (Mantovani, 1999). In accordo con gli schemi classici di struttura dei fattori di trascrizione, le regioni conservate sono richieste per le interazioni con il DNA o con altri fattori proteici, mentre le funzioni d’attivazione sono da ricondurre alle regioni ricche in glutammina. Queste ultime, anche se meno strettamente conservate, devono avere residui aminoacidici funzionalmente equivalenti mantenuti in posizioni strategiche tanto che il dominio di attivazione della subunità NF-YA dello Schisostoma mansonii attiva le cellule di mammifero. Nei lieviti S. cerevisiae e S. pombe non si hanno dominii ricchi in Q conservati, nelle subunità HAP2 e HAP5, corrispondenti rispettivamente alle subunità NF-YA e NF-YC, però si ha una quinta subunità, HAP4, con un dominio C-terminale fortemente acido, per la quale è supposto il ruolo di attivazione del complesso quadrimerico (Mantovani, 1999).
Fig. 2.1. Rappresentazione schematica delle subunità NF-Y. I domini omologhi alle subunità HAP2, HAP3 e HAP5 del lievito sono indicate. I domini in bianco delle NF-YB e NF-YC indicano le posizioni delle α-eliche del motivo di ripiegamento istonico (HFM). Nelle cornici sotto le subunità sono mostrate le sequenze del dominio conservato dell’uomo NF-YC, dell’Arabidopsis thaliana NF-YCb, S. cerevisiae HAP5, NC2α, e
Xenopus H2A (cornice inferiore) e le sequenze corrispondenti dell’uomo NF-YB, di Arabidopsis thaliana NF-YBb, S. cerevisiae HAP3, NC2β e Xenopus H2B. I simboli +
indicano la posizione delle α-eliche nel HFM; L1 e L2 indicano le regioni delle anse (Figura tratta da Mantovani, 1999).
2.1 Il motivo di ripiegamento istonico
Nei nuclei interfasici, il DNA è impacchettato in una struttura nucleoproteica complessa detta cromatina. Il primo livello di impacchettamento del DNA è costituito dai nucleosomi, che rappresentano l’unità fondamentale della cromatina. Il nucleosoma è costituito da un “core” proteico avvolto da 145-147 bp di elica sinistrorsa
di DNA che lo avvolge per 1.65 giri. “Core” adiacenti sono separati dalle regioni di DNA “linker” di 20-80 bp. La componente proteica della particella “core” è costituita da un ottamero proteico in cui sono rappresentate due copie per ciascun istone H2A, H2B, H3 e H4, mentre l’istone H1 si associa al DNA “linker”.
Gli istoni sono le proteine più conservate dell’evoluzione e contengono una regione di 65 aminoacidi “histone fold motif” (HFM), comune a tutti loro, con un’identità di sequenza aminoacidica che è mediamente del 14-18%, ma con un’elevata somiglianza strutturale. Il dominio HFM, è organizzato in tre o quattro α-eliche: α1, α2, α3 e αC o αN , separate da corte regioni dette anse: L1, L2, L3, che intervengono nell’interazione con gli altri istoni e con il DNA. L’allineamento delle sequenze aminoacidiche di istoni e subunità NF-YB e NF-YC suggeriscono che questo ultime contengono il motivo strutturale di HFM (Baxevanis et al., 1995) con un’elevata conservazione aminoacidica dall’uomo, alla A. thaliana (YBb/YCb) e al lievito (HAP3/HAP5). Nelle porzioni di sequenza corrispondenti alle eliche α1, α2 e L2 , e anche all’ansa L1 delle subunità NF-YC la similarità è più elevata, mentre la terza elica, α3, risulta essere poco conservata. Sono mantenuti in particolare molti aminoacidi caratteristici del HFM: alanine in posizioni critiche nelle seconde metà delle due eliche, α2 e α3; residui basici in α1, alternanza di aminoacidi carichi e idrofobici in α2; residui basici e una treonina nell’ansa L2. Il confronto con gli istoni evidenza la presenza di una quarta elica, αC, per le proteine NF-YB, come per gli istoni H2B, e αN per le proteine NF-YC, come per gli istoni H2A. La somiglianza con gli istoni è tale che è stato ipotizzato che la subunità NF-YB appartenga alla famiglia dell’istone H2B, avendo il 30% di identità aminoacidica con esso; analogamente, la NF-YC e l’istone H2A farebbero parte della stessa famiglia avendo una identità del 21% ed una similarità del 47% (Baxevanis et al., 1995). La subunità NF-YA, a differenza di NF-YB e NF-YC, non ha dominii omologhi a nessun motivo noto di legame al DNA, ma ha comunque una regione conservata che può essere divisa in due distinte metà, ciascuna lunga
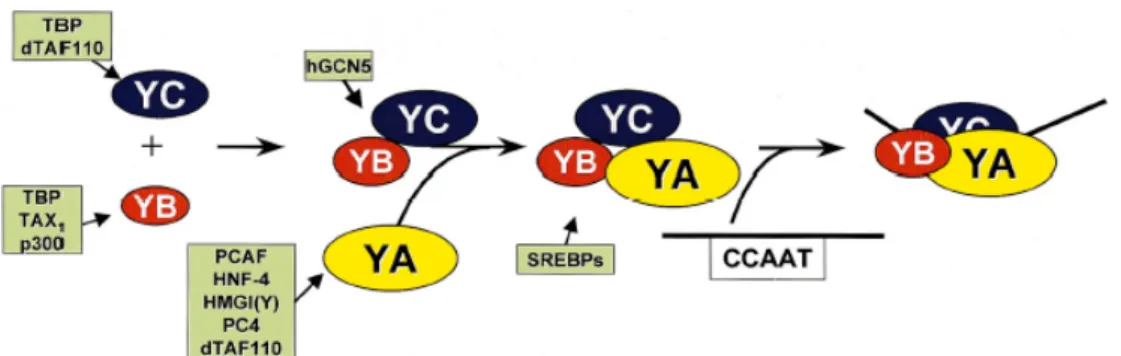
20 aminoacidi, che formano α eliche anfipatiche. Studi di mutagenesi, in ratto e in lievito, dimostrano che l’interezza del motivo di ripiegamento istonico è necessaria per la corretta formazione del trimero e per la interazione con la sequenza CCAAT. Per prime le subunità, YB e NF-YC, formano uno stretto dimero che offre una superficie di legame per la terza subunità NF-YA. Il trimero risultante solo ora può legarsi al DNA con un’elevata specificità e affinità e per certi siti la costante di dissociazione, di questo fattore di trascrizione, raggiunge perfino valori tra 10-10 e 10-11. Tutte le subunità del fattore NF-Y sono, in ogni caso, necessarie per il legame al DNA (McNabb et al., 1995; Sinha et al., 1995). Per la formazione del dimero sono richiesti aminoacidi specifici nell’elica α2, nell’ansa L2 e nell’elica α3 delle subunità NF-YC e NF-YB mentre per l’associazione con la subunità NF-YA sono necessari due aminoacidi conservati nell’elica α2 in NF-YB e alcuni residui nelle eliche, α1 e α2 e nella parte C-terminale dell’elica α3 dell’ NF-YC. Il legame col DNA richiede che siano intatte le eliche α1 e α2 del peptide NF-YB e dell’elica α1 del peptide NF-YC. Per dare origine al legame con il DNA sono indispensabili probabilmente anche aminoacidi con carica positiva nell’ansa L2 (Fig. 2.2).
Fig. 2.2. Associazione delle subunità NF-Y, legame al DNA ed interazioni con vari attivatori trascrizionali (Figura tratta da Mantovani, 1999).
2.2 Interazioni con altri fattori proteici associati alla cromatina
Molti sono i peptidi associati alla cromatina con i quali il fattore nucleare NF-Y deve interagire per esercitare la sua funzione. Un ruolo molto importante e’ svolto dalla porzione delle NF-YB e NF-YC in cui è presente l’HFM. Questo motivo si ritrova anche in altre proteine che intervengono nella regolazione della trascrizione: (I) le due subunità, NC2α e NC2β, di NC2 (chiamato anche Dr1/DRAP1) che lega TBP (proteina di legame al motivo TATA) con effetto inibitorio sulla trascrizione. In particolare NF-YC è molto simile a NC2α soprattutto nell’elica α1 e nella parte C terminale dell’elica α3, mentre per NF-YB e NC2α, la similarità è ristretta all’elica α2, all’ansa L2 e alla porzione C-terminale dell’elica α3. La similarità tra l’istone H2A, NF-YC e NC2α ha fatto ipotizzare che questi tre peptidi facciano parte di un’unica famiglia istonica, così come l’istone H2B, NF-YB e NC2α. (II) alcuni TAFII (dTAFII60 / hTAFII80, dTAFII40 / hTAFII31 e hTAFII20 / dTAFII30α). Ι fattori TAFII interagiscono strettamente con TBP e mediano l’attivazione della trascrizione come parte del complesso TFIID. La subunità NF-YB sembra essere la più direttamente coinvolta nell’interazione tra NF-Y e il complesso TFIID o con TBP. L’interazione con il complesso TFIID è stato individuato tramite esperimenti di immunoprecipitazione che hanno indicato essere implicato circa il 20% di NF-YB, da solo o con NF-YC, ma mai con NF-YA. In particolare, i fattori hTAFII80, hTAFII31 e hTAFII20i, componenti del complesso TFIID, sono coinvolti. Deve essere ricordato che hTAFII80 e hTAFII31 sono molto simili agli istoni H4 e H3 rispettivamente, anche se non nella misura in cui NF-YB e NF-YC somigliano agli istoni H2B e H2A. Sicuramente sono conservati i residui aminoacidici necessari per le interazioni di tipo istonico, che sottintendono la possibilità di formazioni nucleosoma-simili tra i fattori TAFII e il dimero NF-YB/NF-YC. Le subunità NF-YB e NF-YC sono coinvolte anche in una interazione diretta con TBP in una porzione compresa rispettivamente tra gli aminoacidi 100 e 112 per la NF-YB di Z. mais, e gli aminoacidi 198 e 210 di S. cerevisiae per la NF-YC (Bellorini et al., 1997).Queste regioni sono molto conservate, in termini di identità o similarità aminoacidica, anche nel fattore hFTAFII20 e in NC2α tanto che questi fattori proteici sono stati proposti come membri di una sub-famiglia delle proteine HFM che ha acquisito la capacità di legare TBP prima della radiazione funghi/piante.
2.3 Modalità d’azione di NF-Y
Sono state individuate varie forme d’interazione tra NF-Y e altri fattori proteici, ma in cosa esse si traducono e in che modo si esplica l’effetto sulla trascrizione è poco noto. In alcuni casi, come nel rapporto con le SREBP (proteine che si legano agli elementi di regolazione SRE, presenti a monte di geni coinvolti nel metabolismo del colesterolo) (Mantovani, 1999), NF-Y determina un aumento dell’affinità delle stesse SREBP agli elementi SRE; viceversa nel caso dell’interazione NF-Y/Sp1 viene reciprocamente stabilizzato il legame con le rispettive sequenze bersaglio del DNA. In questi esempi sono probabilmente le regioni ricche in glutammina delle subunità NF-YA e NF-YC ad interagire con regioni analoghe degli altri fattori, e molto importante è sicuramente la distanza reciproca tra le rispettive sequenze nucleotidiche bersaglio. La subunità NF-YB sembra essere invece la più direttamente coinvolta nella formazione del complesso di pre-inizio della trascrizione ed è richiesta anche per la re-iniziazione. In particolare, sulla base delle strette somiglianze dei dimeri NFY-B/NF-YC e NC2 è ipotizzato un meccanismo d’azione che ritiene il fattore NF-Y, capace di esercitare un’azione positiva sulla trascrizione in modo indiretto, cioè impedendo l’accesso di un regolatore negativo, come NC2, e comportandosi quindi come un contro repressore (Bellorini et al., 1997). Per quanto riguarda l’interazione NF-Y/TFIIID, in esperimenti di immunopurificazione e immunoprecipitazione è stato valutato che circa il 20% di NF-YB è associato con TFIID. L’interazione può ancora essere mediata dal motivo HFM e interessare all’interno del complesso TFIID i componenti che
presentano lo stesso motivo (dTAFII60 / hTAFII80, dTAFII40 / hTAFII31 e hTAFII20 / dTAFII30α). Un’importante classe di proteine con le quali il fattore NF-Y deve interagire è rappresentata dagli istoni. Non sono disponibili molte informazioni, ma alcuni dati interessanti sono stati acquisiti. Gli istoni sono associati al DNA nella struttura ripetitiva, rappresentata dai nucleosomi, generalmente inibitoria per il legame delle proteine trascrizionali alla sequenza del promotore. Una domanda chiave è, quindi, come, nell’attivazione fisiologica dell’espressione genica, gli attivatori gene specifici e i fattori generali di trascrizione possano raggiungere i rispettivi siti-bersaglio, se questi sono organizzati in strutture cromatiniche.
Gli attivatori della trascrizione possono essere suddivisi in tre classi sulla base della loro affinità per il nucleosoma, che è mediamente bassa. Una delle tre classi, però, alla quale appartengono i recettori ormonali, per glucocorticoidi e per il progesterone, è capace di associarsi efficientemente ai nucleosomi: NF-Y sembra rientrare in questa categoria. Esperimenti di ricostruzione in vitro della cromatina dimostrano, infatti, che NF-Y, come trimero, si associa indifferentemente con DNA nudo o impegnato nella struttura nucleosomale, senza richiedere l’intervento d’altri elementi (fattori di trascrizione o proteine rimodellanti la cromatina), che ne facilitano il processo. E’ stato ipotizzato che NF-Y di per se non possegga una reale forte potenzialità d’attivazione della trascrizione, ma piuttosto giochi un ruolo dominante nella determinazione dell’architettura del promotore. In effetti, NF-Y compete efficientemente con la formazione dei nucleosomi anche se non è chiaro come. Due sono i meccanismi ipotizzati: NF-Y si lega al DNA e previene l’associazione ad esso del tetramero istonico H3-H4; alternativamente il dimero NF-YB/NF-YC, sulla base della somiglianza con il dimero istonico H2B/H2A, può competere con questo nella formazione del nucleosoma. In quest’ultima ipotesi l’intervento della subunità NF-YA stabilizzerebbe successivamente il legame alla sequenza CCAAT. Il dimero NF-YB/NF-YC è, in effetti, stato identificato (Bellorini et al., 1997;
Motta et al., 1999). Un ruolo sicuramente importante è svolto da enzimi che modificano post-traduzionalmente gli istoni e che possono agire direttamente in loco sulla cromatina. Questo è il caso ad es. degli enzimi acetilanti/deacetilanti. In effetti, sono stati individuati dimeri di NF-YB/NF-YC associati ad attività acetilasica ed inoltre TSA, inibitore di HDAC (istone deacetilasi), induce in vivo promotori NF-Y dipendenti, viceversa, mutazioni nella sequenza CCAAT bloccano l’induzione da parte di HAT (istone acetil-trasferasi). NF-Y probabilmente media l’azione di HAT ed in particolare NF-YA è la subunità che lega l’enzima, tanto da coprecipitare con esso in esperimenti di immunoprecipitazione (Caretti et al., 2003). Il reclutamento dell’acetil-trasferasi per gli istoni, da parte di NF-Y, potrebbe spiegare come siano preservate le caratteristiche epigenetiche del promotore antisenso del gene Kcnq ICR del topo. E’ stato dimostrato che, in condizioni in cui il gene Kcnq ICR deve essere silente, NF-Y è importante per mantenere il promotore antisenso in uno stato trascrizionalmente competente, ma anche per impedire la metilazione de novo delle isole CpG dello stesso promotore. E’ stato in effetti osservato che la lisina in posizione 9 dell’istone H3 risulta meno acetilata e più metilata in mutanti in cui sostituzioni nel pentanucleotide si associano a aumento della metilazione e perdita delle capacità trascrizionali del promotore antisenso (Pandey et al., 2004). NF-YB è stato invece identificato come substrato esso stesso di acetilazione da parte di p300 sul promotore di HSP70 in oociti di X. laevis. Un altro esempio: l’associazione tra NF-Y e p300 sui promotori della ciclina B2 in vivo è risultata regolata durante il ciclo cellulare e positivamente correlata con la funzione del promotore (Salsi et al., 2003). Possiamo ipotizzare, anche se in modo molto riduttivo, che ordini superiori di struttura della cromatina impieghino NF-Y in complessi contenenti attività acetilasiche, la cui azione modifichi le interazioni con e tra le code istoniche dei nucleosomi vicini, facilitando l’accesso ad altri fattori trascrizionali. La fosforilazione è un altro tipo di modificazione post-traduzionale che interessa gli istoni, ma anche la NF-YA è risultata un substrato fisiologico
della fosforilasi cdk2-ciclinaA dipendente. La fosforilazione avviene sia in vitro sia in vivo, in parallelo con l’attivazione, correlata al ciclo cellulare, della fosforilasi cdk2-ciclinaA dipendente ed interessa due serine vicine al dominio di legame con il DNA. La localizzazione dei residui aminoacidici fosforilati spiega probabilmente perché la fosforilazione della NF-YA non ha effetto sulla formazione dell’eterotrimero, ma è necessaria per l’efficiente legame dell’NF-Y al DNA.
2.4 Regolazione del fattore NF-Y
Inizialmente era stato supposto che NF-Y fosse un fattore trimerico costante, non inducibile, osservazioni più recenti hanno invece modificato questa convinzione. Infatti, La capacità del fattore nucleare di legare il pentanucleotide CCAAT cambia in stadi diversi di sviluppo o differenziamento (cellule B, cellule muscolari) e nella senescenza cellulare (Yun et al., 2003). La regolazione di NF-Y può avvenire a vari livelli, nei paragrafi successivi sono descritti alcuni esempi.
2.5 Regolazione post trascrizionale
Le quantità degli mRNA relativi alle tre subunità rimangano sostanzialmente costanti, in diversi sistemi è stata rilevata una regolazione post trascrizionale che riguarda le subunità NF-YA e NF-YB. Si hanno infatti variazioni della disponibilità della subunità NF-YA, in differenti tipi cellulari e/o condizioni di crescita (Mantovani, 1999) ma non è chiaro se questo tipo di controllo si instaura sulla base della traduzione o della degradazione del peptide. Sicuramente la NF-YA costituisce la subunità regolatrice del complesso trimerico. La subunità NF-YB invece, sebbene rimanga costante in quantità, potrebbe essere regolata attraverso la fosforilazione e l’acetilazione ricordate nel paragrafo precedente (vedi Modalità d’azione di NF-Y).
2.6 Regolazione della localizzazione nucleare
Durante l’embriogenesi di X. laevis si instaura un controllo della localizzazione nucleare dell’attività di legame al pentanucleotide CCAAT: nelle uova non fertilizzate e nei primi stadi di sviluppo, questa è localizzata nel citosol e nell’estratto nucleare non associato alla cromatina, viceversa aumenta notevolmente la frazione associata alla cromatina allo stadio di blastula intermedia, quando si verifica un aumento considerevole del numero di geni trascritti.
- “Splicing” Alternativo
La frazione NF-YA presenta lo “splicing” alternativo di un esone relativo al dominio di attivazione ricco in glutammina (Li et al., 1992), ed anche alcune NF-Y di A. thaliana hanno uno “splicing” tessuto specifico (Gusmaroli et al., 2001).
2.7 Regolazione nelle piante
Un tipo di regolazione peculiare deve essere supposto per le piante che presentano famiglie multigeniche per tutte e tre le subunità del NF-Y, a differenza degli altri eucarioti che presentano un gene per subunità. In A. thaliana ad es. per tutte e tre le subunità esistono membri ubiquitari e altri strettamente tessuto specifici (Gusmaroli et al., 2002). I geni LEAFY COTYLEDON1 e LEAFY COTYLEDON1-LIKE, che appartengono alle NY-B e sono espressi specificamente negli embrioni, saranno trattati nel paragrafo successivo.
2.8 Le proteine LEAFY COTYLEDON1 e LEAFY COTYLEDON1-LIKE
sono subunità altamente specializzate del fattore NF-Y
Alcuni studi hanno dimostrato che i geni LEAFY COTYLEDON1 (LEC1) e LEAFY COTYLEDON1-LIKE (L1L) codificano per presunti fattori di trascrizione. Il gene LEC1 codifica una proteina con sequenza simile alla subunità HAP3; una subunità del fattore di trascrizione NF-Y (Nuclear Factor-Y), conosciuto anche come CBF, che lega il pentanucleotide CCAAT-box-binding-factor (Lotan et al., 1998) (Fig. 2.3).
Fig. 2.3. Allineamento delle sequenze aminioacidiche del dominio B delle subunità HAP3 di Arabidopsis, lievito e ratto. In giallo sono mostrati i residui identici. In rosso sono evidenziati i residui aminoacidici specifici di LEC1 e L1L. Le linee continue e tratteggiate in azzurro indicano rispettivamente le posizioni delle α-eliche e le anse nel motivo di ripiegamento istonico. I numeri indicano le posizioni degli aminoacidi all’interno di LEC1.(Lee et al., 2003).
In particolare all’interno del dominio B, è evidenziabile un motivo di ripiegamento istonico (HFM) costituito da tre/quattro α-eliche, α1, α2, α3 e αC, intervallate da corte regioni dette anse: L1, L2 e L3 e la comparazione con gli istoni evidenzia che le subunità HAP3
appartengono alla famiglia istonica H2B, avendo il 30% di identità di sequenza. Sulla base della conservazione e sull’identità di sequenza del domino centrale B, in Arabidopsis thaliana, la famiglia genica che codifica per le subunità HAP3 è suddivisa in due distinte classi: una di “tipo non-LEC1” (NB) e l’altra di “tipo-LEC1” (LB). Il dominio LB è stato trovato in Phaseolus coccineus, Daucus carota, Triticum aestivum e Oryza sativa. In A. thaliana, esso è presente nelle proteine codificate dai geni LEC1 e L1L, e differisce dal dominio B di tipo “non-LEC1” per 16 residui amminoacidici conservati. Il dominio di “tipo-LEC1” è richiesto per il corretto funzionamento del fattore NF-Y, è sufficiente ad indurre lo sviluppo embrionale da cellule vegetali già differenziate ed è in grado di regolare l’embriogenesi zigotica e somatica (Lotan et al., 1998; Harada 2001).