CHAPTER 4
SYSTEMATIC PALAEONTOLOGY
4.1 INTRODUCTION
Within the past two decades a series of articles on the systematics of the Caribbean and West Atlantic sirenian species was published (Domning, 1988, 1989a, 1989b, 1990, 1997, 2005; Savage et al., 1994). More recently, new records have permitted review and description of the Neogene Euro-North African sirenians: Metaxytherium krahuletzi (Domning & Pervesler, 2001), Metaxytherium serresii (Carone & Domning, 2007) and Metaxytherium petersi (Domning & Pervesler, in prep.).
In this chapter, I review and describe the following species: Rytiodus capgrandi, Rytiodus sp., Miosiren kocki, Prohalicore dubaleni, Metaxytherium aquitaniae, Metaxytherium catalaunicum, Metaxytherium medium and Metaxytherium subapenninum, in order to complete the revisions of the Neogene Euro-North African sirenian species.
The Neogene Euro-North African sirenian species belong to three subfamilies: the Trichechidae Miosireninae; the Dugonginae Halitheriinae and the Dugongidae Dugonginae. Miosireninae and Halitheriinae are represented in the Palaeogene Euro-North African sirenian record too. In this chapter I include also the review of “Halitherium” bellunense, determined to be late Oligocene in age (see Chapter 3) and considered a dugongine (Domning, 1996), in order to demonstrate that the Dugonginae also lived along the Euro-North African coasts during the Palaeogene.
Moreover, I describe a new specimen from the Lower Miocene deposits of Manosque (southern France).
4.2 MATERIAL AND METHODS
The following descriptions are based on the main Neogene Euro-North African sirenian specimens listed in Appendix 2. Most of them were personally examined; the remainder were studied through literature and through pictures kindly provided by museum curators. Other sirenian specimens listed in Appendix 3 were personally observed for comparison.
The specimens that were personally examined were described; photographed in dorsal, ventral, lateral, medial, anterior and posterior views with a digital camera; and measured with calipers to the nearest millimetre (teeth to the nearest tenth of a millimetre). The descriptions supplement previous descriptions, if available, and are centred particularly on the cranial anatomy, whose characters have been used for the cladistic analysis presented in chapter 5.
Emended diagnoses and comparisons are made according to the results of the cladistic analysis.
The anatomical terminology and measurements follow Domning (1978; 1988), Bajpai and Domning (1997), and Domning and Pervesler (2001).
Measurements (in mm), ratios and selected character states of specimens are listed in Appendix 4.
Photos of sirenian specimens examined are shown in Appendix 5.
Abbreviations – Institutional abbreviations are listed in Appendix 1.
In reference to morphological character states: c. character state as described and numbered in Chapter 5 (e.g., c. 7(1) refers to state one of character seven).
In reference to anatomical features: AS, alisphenoid; BO, basioccipital; BS, basisphenoid; C, canine; dp, deciduous lower premolar; DP, deciduous upper premolar; E, ethmoid; EE, exethmoid; EO, exoccipital; ET, ethmoturbinalia; FIO, infraorbital foramen; FR, frontal; I, upper incisor; i, lower incisor; IN, incus; J, jugal; L, lacrimal; m, lower molar; M, upper molar; MA, malleus; ME, mesethmoid; MF, mastoid foramen; MN mandible; MX, maxilla; N, nasal; OC, occipital condyle; OS, orbitosphenoid; P, parietal; PE, periotic; PAL, palatine; PM, permanent upper premolar; pm, permanent lower premolar; PMX, premaxilla; PR, processus retroversus; PS, presphenoid; PT, pterygoid; RD, rostral deflection; S, stapes; SO, supraoccipital; SQ, squamosal; T, tympanic; TC, temporal condyle; V, vomer.
In reference to molar features: Upper molar: ac, anterior cingular cusp; hy, hypocone; mcl, metaconule; me, metacone; pa, paracone; pc, posterior cingular cusp; plc, posterolingual cingular cusp; pr, protocone; prl, protoconule. Lower molar: ac, anterior cingular cusp; end, entoconid; hld, hypoconulid lophule; hyd, hypoconid; med, metaconid; prd, protoconid.
4.3 DESCRIPTION AND COMPARISON
4.3.1 MIOSIREN KOCKI
Order SIRENIA Illiger, 1811 Family TRICHECHIDAE Gill, 1872 Subfamily MIOSIRENINAE Abel, 1919
Genus Miosiren Dollo, 1889
Halitherium Flower, 1874 Quart. Journ. Geol. Soc. London 30: 6.
Miosiren Dollo, 1889 Bull. (Proc.-verb.) Soc. Belge Géol. Pal. Hydrol. 3: 420.
Type species – Miosiren kocki Dollo, 1889
Included species – Only M. kocki Dollo, 1889; M. canhami (Flower, 1874) Sickenberg,
1934 is probaly a synonym.
Range – Known with confidence only from the Aquitanian of Belgium, and probably
from reworked deposits of England.
Emended diagnosis – The same as for the type species.
Miosiren kocki Dollo, 1889 (Plates 1-2; Figures 1-5)
Halitherium Canhami Flower, 1874 Quart. Journ. Geol. Soc. London 30: 6.
Miosiren Kocki Dollo, 1889 Bull. (Proc.-verb.) Soc. Belge Géol. Pal. Hydrol. 3: 420. Miosiren canhami (Flower) Sickenberg, 1934 Mém. Mus. Roy. Hist. Nat. Belgique 63:333 Dec. 31, 1934.
Holotype – RBINS M.136 (= no. 1682), cranium and partial skeleton of adult.
Type locality - Brickworks of Charles de Kock, “in den Hoek” near Boom, Belgium.
Sands of Edeghem Formation. Aquitanian in age (21.5 - 23.03 Ma).
Referred specimens - Includes Miosiren canhami (Flower, 1874) Sickenberg, 1934,
considered by Domning, 1996 as a possible synonym of M. kocki and represented by a partial skull (IM unnum.) reworked into the Red Crag Formation from earlier, possibly Miocene deposits of Foxhall, near Waldringfield, Suffolk, England.
Range – Known with confidence only from the lower Burdigalian of Belgium, and
probably from reworked deposits of England.
Emended diagnosis – Miosirenine trichechids characterized by a supraorbital process
of frontal dorsoventrally thickened [c. 36(1)]; a long paroccipital process [c. 65(0)]; a ventral extremity of jugal ventral to orbit [c. 85(2)]; a posterior border of palate not incised [c. 97(0)]; large tusks with alveoli extending much more than half length of the premaxillary symphysis [c. 140(2)]; also a posttympanic process of squamosal enlarged and clublike and a third molar very reduced, not molariform.
Remarks – The Miosiren kocki holotype (RBINS M.136) was found in 1889 in the
Miocene deposits of Boom (Anvers, Belgium) and given to the RBINS. After its extraction and restoration it was described by Dollo in a short note (1889). Dollo compared the specimen with the other sirenians known at the time and, on the basis of differences observed, coined the new binomial Miosiren kocki (“Miosiren” because it was a Miocene sirenian, and “kocki” in honour of M. Charles de Kock). In 1934 Sickenberg gave a complete description of the specimen in his review of sirenians of Belgium.
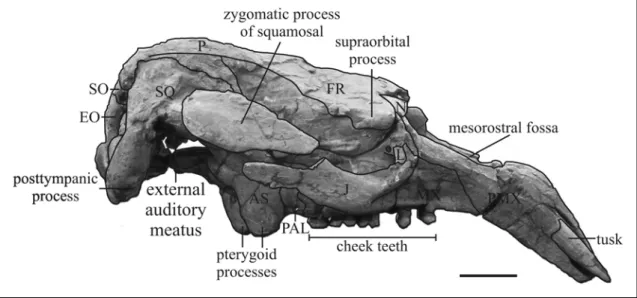
Description – The holotype of Miosiren kocki is a nearly complete skeleton housed in
RBINS, exhibited at a height of one metre, and mounted with metal studs; only the cranium was removed to be studied. Hence, I give a new description only of the cranial anatomy of the holotype, and I refer to Sickenberg (1934) for the postcranial description. The mandible is not preserved; the cranium is nearly complete and divided into three portions, which permits a good examination of the internal as well as the external anatomy. The body length is estimated to be about 4 m.
Premaxilla – The rostrum is small relative to the cranium [c. 3(0)]; it has a narrow dorsal keel and broadens posteriorly into an eminence that is posteromedially concave but with a boss in lateral view [c. 10(1)]. The lateral edges are nearly vertical, not flaring; the sides of the symphysis are convex surface is trapezoidal in outline; the palatal where swollen by the tusk alveoli, which extend nearly the entire length of the symphysis [c. 140(2)] (see the tusk description below). The symphysis is 123 mm long, not entirely fused. The nasopalatine canal is subtriangular in cross-section. The opening of the premaxillary canal lies posteroventral to the tusk alveolus. The masticating surface is smooth, nearly flat; its right and left halves form a wide angle about of 168° and are separated by a groove. The nasal process is long [c. 7(0)], contacts the frontal [c.
9(1)], the nasal and ventrolaterally the maxilla; but it contacts neither the lacrimal [c. 1(0)] nor the jugal. The portion of the nasal process in contact with nasal and frontal is thin and tapering [c. 6(0)], but extends less than half length of mesorostral fossa. The mesorostral fossa is long, reaching the level of the anterior margin of the orbit [c. 8(1)]; its anterior end is rounded and constricted by a bulge on medial side of premaxilla; its posterior end is incomplete, nevertheless the absence of a deep nasal incisure can be established [c. 37(0)]. The rostral deflection is 49°.
Fig. 1: Cranium of Miosiren kocki (RBINS M.136) in anterior view. Broken lines indicate the supposed nasal sutures. Scale bar: 5 cm.
Nasal – The nasals are large, thick, and massive, and meet in the midline [c. 31(0)]; the internasal suture is about 100 mm long. Their dorsal exposure is elliptic in shape, anterolaterally inclined at about 45°.
Ethmoidal region – The perpendicular plate reaches the roof of the narial passage, is 27 mm long ventrally, 15 mm thick in the centre, and narrows posteriorly, ventrally and
slightly dorsally. The anterior surface for the cartilaginous septum is wider dorsally than ventrally, and posterodorsally inclined at about 45°. Only a cylindrical dorsal longitudinal concha, anteriorly fractured and 100 mm long, is preserved.
Vomer – Not preserved.
Lacrimal – The lacrimals are small, irregular in shape, and without a nasolacrimal canal [c. 91(1) in contrast with Sickenberg, 1934: fig. 12]. The right one is better preserved; it abuts dorsally against supraorbital process of frontal; is sutured medially to maxilla, anteroventrally fits in socket on dorsal surface of jugal, and it is not in contact with the premaxilla. The jugal appears to overlap its lateral side.
Frontal – The frontals are dorsally fragmented and the interfrontal suture appears to be open. The supraorbital process is about 2 cm thick dorsoventrally with a small posterolateral corner [c. 36(1)], flattened in horizontal plane, with a dorsal surface inclined relatively gently ventrolaterad [c. 43(0)]; not divided on its lateral margin. A small orbicular apophysis is present. Some fragments of the lateral crests are preserved on the left side; they appear to be very low, not overhanging but with a sharp edge. The lamina orbitalis of frontal is about 4 cm thick [c. 38(1)], and forms the medial wall of the temporal fossa; the development of its falciform anterior edge is not determinable. The medial portion of frontal roof is convex [c. 42(0)], smooth, with a median boss.
Parietal – The parietals are shorter than the frontals in midline; they are dorsally fragmented and the interparietal suture appears to be open, while the occipito-parietal suture is closed and the fronto-parietal suture is not evident. The cranial vault is more or less trapezoidal in coronal section anteriorly; about 3 cm thick in anterior midline. The temporal crests are very low, nearly absent, and the posterolateral corners of roof are convex, smooth and indented by squamosal. The roof is broad, convex [c. 45(0)], without a sagittal crest [c. 51(1)]. The intertemporal region is as broad as the braincase [c. 46(2)]. The internal occipital protuberance is absent; the transverse sulcus is shallow with an anterior pit; the bony falx cerebri is low, and does not reach the frontoparietal suture [c. 47(1)]. A small concavity, lying 25 mm anterior to the external occipital protuberance, could represent a closed emissary foramen.
Fig. 2: Posterior portion of cranium of Miosiren kocki (RBINS M.136) in dorsal view. Broken lines indicate the supposed interparietal, interfrontal, internasal, fronto-parietal and fronto-nasal sutures. Scale bar: 5 cm.
Supraoccipital – The supraoccipital is hexagonal in outline with rounded dorsolateral corners. It forms with the after part of parietals an angle of 95°. The external occipital protuberance is low, and does not rise above the plane of parietal roof; the median ridge below it is distinct, developed into a rounded protuberance in its central part; and long, reaching the ventral border of the supraoccipital. The nuchal crest is low, rounded, and indistinct at its lateral end near the squamosal. The areas of insertion for the semispinalis capitis muscles are flattish, oval, rugose, and face posterodorsad, extending more than halfway to ventral end of supraoccipital. The lateral borders are thick and rounded, sloping outward at bottom, without overhanging upper corners. The suture with the exooccipitals is not clearly visible. The ratio of width to height of supraoccipital is 1.9.
Exoccipitals – The exoccipitals seem to be united in midline, but with a dorsally peaked foramen magnum [c. 66(0)]. The dorsolateral border is about 2 cm thick and overhanging posteriorly, forming a rugose surface, but not a flange [c. 70(0)], at level of the top of the supracondylar fossa. The fossa is deep, extending across entire width of occipital condyle [c. 67(1)]. The arc of condylar articular surface subtends an angle of 122°. The condyloid foramen is single. The paroccipital process is well developed, reaching slightly below the level of the tip of the condyle [c. 65(0)]. It has a rugose posterior surface and a posteroventrally curved flange.
Fig. 3: Line drawing of cranium of Miosiren kocki (RBINS M.136) in posterior view. Broken linese indicate the supposed interexoccipital and supraoccipital-exoccipital sutures. Grey lines indicate the widening of supracondylar fossae. Scale bar: 5 cm.
Basioccipital – The basioccipital bears a single convex rugosity on the ventral side for the longus capitis muscles. The posterior slope is steeper than the anterior.
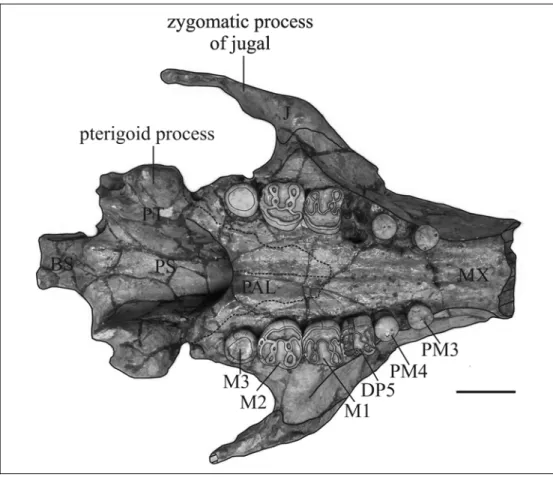
Basisphenoid – The ventral side is fragmented; it appears to be flat, about 63 mm wide, with two posterolateral longitudinal protuberances. The sella turcica is shallow and the tuberculum sellae is high, anteroventrally inclined. The medial walls of the pterygoid processes are nearly vertical.
Presphenoid – It is fused with the surrounding bones. Only the base of the orbitosphenoidal crest is preserved; it appears to be long and well developed.
Orbitosphenoid – The optic foramen seems to lie at the level of dorsal side of sphenorbital foramen. The bony wall lateral to the optic foramen bears a distinct pointed process.
Alisphenoid – The posterolateral side of the pterygoid process is smooth, dorsoventrally concave with medial and lateral crests. A slight convexity continues forward edge of the zygomatic root anteriorly. An alisphenoid canal is absent [c. 101(1)]. The suture with squamosal includes an anteroventral large sphenorbital foramen. The foramen ovale is open.
Pterygoid – Fused with the surrounding bones; forms the posteromedial part of the pterygoid process. The pterygoid processes are relatively short, extending just 17 mm below the level of the palate. The pterygoid fossa is broad and well-developed, extending above level of roof of internal nares [c. 102(1)]. The ventral tips of alisphenoid and pterygoid enclose end of palatine in slot between them anteromedially: The ventral tip of the pterygoid process is concave, not enlarged.
Palatine – The palatine forms the anteromedial side and (together with alisphenoid) the smooth and rounded anterolateral extremity of the pterygoid process. The medial sutural contact between palatines extends from forward of the posterior edge of the zygomatic-orbital bridge [c. 99(0)] to behind the level of M3; therefore the palate is long and thick, being 34 mm thick at the level of M2 [c. 16(1)], and its posterior border is slightly concave, not incised [c. 97(0)].
Maxilla – The alveolar portion of maxilla is heavy and massive; its dorsal side is slightly convex mediolaterally with a longitudinal groove for the vomer and the mesethmoid attachments. The vertical plate of maxilla helps form the wall of temporal fossa and contacts the lamina orbitalis of frontal; moreover it forms the wall of the orbital fossa probably with ethmoids. The infraorbital foramen is round, moderately large, with a diametre of about 13 mm [c. 13(1)], and not obstructed [c. 20(0)]. On the right side, on the anterodorsal edge of the orbital fossa, above the infraorbital foramen, a small obturated foramen is present.
The edges of the palatal surface are lyriform; the surface narrows forward of the molars, becomes narrowest forward of the anterior edge of the zygomatic-orbital bridge, then widens at the posterior end of the rostral masticating surface. The palatal and rostral
surfaces are flat. The zygomatic-orbital bridge is elevated above palate [c. 11(1)]; it is about 36 mm thick, medially 87 mm long and narrows distally. The ventral side is triangular and flat; the anterior side is wide, slightly concave, posteroventrally inclined and overlapped by the anterior crest of jugal; the posterior side is concave.
Squamosal – The squamosal indents the posterolateral corner of the parietal roof and barely extends to the temporal crest [c. 76(1)]. The posterior edge along mastoid foramen is rounded. The prominent sigmoid ridge extends from dorsal end of mastoid foramen to ventral end of posttympanic process, forming a strong laterad-projecting flange with its posteroventral edge broad and rugose. The surface of cranial portion dorsal to zygomatic root is not inflated or bulging. The postglenoid process and postarticular fossa are distinct and well developed. A posttympanic process is present [c. 73(1)], complete on the right side, enlarged and clublike. The external auditory meatus is very broad and shallow, 48 mm wide anteroposteriorly and 33 mm high [c. 82(2)], and about 8 mm long mediolaterally. The zygomatic process is roughly lozenge-shaped in lateral view; broader posteriorly than anteriorly [c. 83(1)]; its medial side is relatively concave, slightly inclined inward dorsally; its posterodorsal edge is convex laterad, enlarged and rugose; its anterior tip is prolonged, but it does not reach the level of posterior extremity of the supraorbital process. A processus retroversus is absent [c. 77(0)]. The zygomatic bridge of the squamosal is 42 mm long anteroposteriorly, with a maximum thickness of 18 mm at the centre; it lies more dorsally than the zygomatic bridge of maxilla and it is anteroventrally inclined; on its ventral side the temporal condyle is oval and shallow.
Jugal – The right jugal is nearly complete, while the left one lacks the posterior portion. The ventralmost point lies ventral to orbit [c. 85(2)]. The ventral tip is thickened and rounded, while the border behind it is narrower. The preorbital process does not contact the premaxilla [c. 87(0)]; it is relatively flat and thin [c. 88(1)], but with a longitudinal crest on the anterolateral border. A postorbital process is present [c. 86(1)]. The ventral margin of orbit is broad, concave but not significantly overhanging [c. 90(0)]. The zygomatic process is anteroventrally inclined, tapered in outline, with a posterior longitudinal process [c. 89(0)]; its lateral side is flat, its medial side is concave.
Fig. 4: Cranium of Miosiren kocki (RBINS M.136) in right lateral view.Some suture lines are not discernable. Scale bar: 5 cm.
Periotic – The lateral portion of periotic is partially preserved on the cranium, better preserved on the right side than on the left. The periotic sits in a closely fitting socket in the squamosal.
DENTITION – The whole upper dentition is preserved with the teeth fixed in the alveoli; only the right DP5 is absent. The upper lifetime dental formula appears to be I 1, C 0, PM 3-4, DP 5, M 1-3 [c. 139(0); c. 143(1); c. 144(1); c. 150(0); c. 155(2)]. The molars are well developed. The molar enamel is smooth and thick, with a maximum thickness of about 2.2 mm on the lingual side of the M2.
I – Both tusks are preserved fixed in the alveoli; about 14 mm protruded. They are large, subelliptic in cross-section [c. 141(0)], nearly straight, and slightly divergent anteriorly. The crown is entirely covered by thin (less than 1 mm thick) enamel, with thin longitudinal lines [c. 136(0)]; it is about 84 mm long and is distinct from the root [c. 137(0) in contrast with Domning, 1994], but the exact boundary is not definable. The roots are closed, indicating the completed development of the tusks.
PM3 and PM4 – They are rounded, with a prominent neck, and they seem to be one-rooted [c. 157(1)]. The crown is entirely covered by enamel. PM3 is constituted by a main centrolabial cusp, slightly worn, with a smaller posterolingual accessory cuspule. PM4 is bigger than PM3, and is constituted by a main centrolabial cusp, slightly worn, with two smaller accessory cuspules; the posterolingual one is unworn, while the anterolabial is slightly worn.
DP5 – Only the left DP5 is preserved, while on the right side just the DP5alveolus is preserved with the loci for three roots: a large root on the lingual side and two smaller roots on the labial side.
The tooth is fully molariform and much bigger than PM3-4. The crown is flattened by wear, but the cusp pattern is visible: two transverse main lophs of three cusps and narrower pre- and postcingula connected lingually to the main lophs. The transverse valley, the anterior and the posterior cingular valleys are labially open
M1 – It is moderately worn; it differs from DP5 in its larger size and in that the precingulum is shorter and limited to the lingual side, but this state could be due to wear.
M2 – It is less worn than the DP5 and M1; it differs from them primarily in its larger size. As in the M1 the precingulum is short, limited to the lingual side.
M3 – It is reduced, smaller than the other molars [c. 159(1)], not molariform; the crown is rounded, with a worn, flat occlusal surface covered by enamel. The roots are not visible.
Fig. 5: Posterior portion of cranium of Miosiren kocki (RBINS M.136) in ventral view.Broken lines indicate the supposed limites of the palatines. Scale bar: 5 cm.
Comparison – Miosiren kocki belongs to the family Trichechidae in having a rostrum
small relative to cranium [c. 3(0)]; a processus retroversus of squamosal absent [c. 77(0)], and an external auditory meatus of squamosal very broad and shallow, wider anteroposteriorly than high [c. 82(2)]; moreover a nasal process of premaxilla not in contact with the lacrimal [c. 1(0)],
Miosiren kocki is close to Anomotherium langewieschei, forming the subfamily Miosireninae, by having primitively large nasals meeting in midline [c. 31(0)]; a thick lamina orbitalis of the frontal [c. 38(1)]; and a supracondylar fossa of the exoccipital deep, extending across entire width of condyle [c. 67(1)]. 38(1), 45(0)r.
Miosiren kocki differs from Anomotherium langewieschei and from the other trichechids in having a supraorbital process of frontal dorsoventrally thickened [c. 36(1)], a long paroccipital process [c. 65(0)]; and a posttympanic process of squamosal enlarged and clublike. Moreover Miosiren kocki differs from A. langewieschei in having a third molar very reduced, not molariform.
Miosiren kocki is characterized by a palate > 1 cm thick at level of penultimate cheek tooth [c. 16(1)], as in the trichechine trichechids Ribodon limbatus and Trichechus spp.; a ventral extremity of jugal ventral to orbit [c. 85(2)]; a posterior border of palate not incised [c. 97(0)]; and large tusks with alveoli extending much than half length of the premaxillary symphysis [c. 140(2)]. These characters can not be scored for Anomotherium langewieschei; therefore I can not establish whether they are diagnostic only for Miosiren kocki or for the whole Miosireninae.
4.3.2 PROHALICORE DUBALENI
Order SIRENIA Illiger, 1811 Family TRICHECHIDAE Gill, 1872 Subfamily MIOSIRENINAE Abel, 1919
Genus Prohalicore Flot, 1887 Prohalicore dubaleni Flot, 1887
(Figure 6)
Prohalicore Dubaleni Flot, 1887, Bull. Soc. Géol. France (3)15: 135.
Holotype – MMM unnum. partial mandible.
Type locality – Quarry at Audon, near Tartas, Landes, SW France. Sables Fauves
Formation; Matilon Member. Serravallian in age.
Referred specimens – Only the holotype is known.
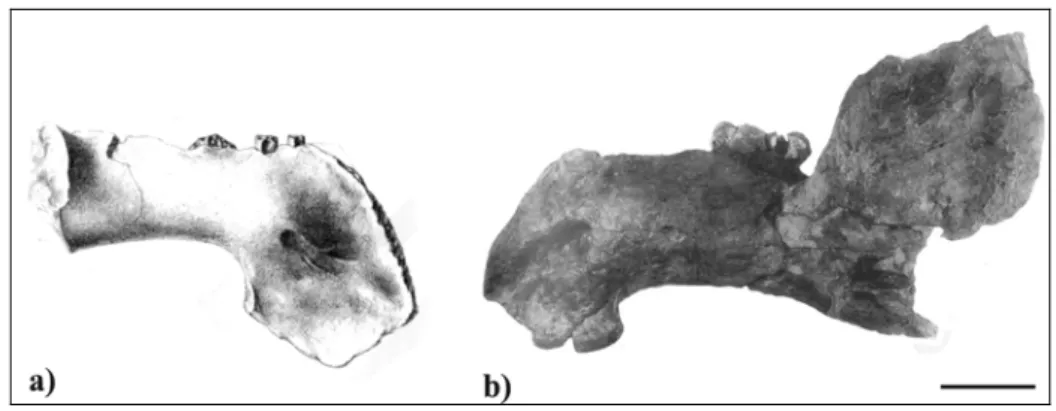
Range – Known only from the Serravallian deposit of Audon, Landes, SW France. Remarks – The status and affinities of this species, known only from a partial mandible
(MMM unnum.), are considered uncertain by Domning (1996). I did not have the opportunity to examine this specimen, but following the description and the figures of Flot (1887), I made some observations.
The mandible is Middle Miocene (Serravallian) in age and not Early Miocene as previously supposed (see Chapter 3). The only recognized Middle Miocene sirenian species considered to live along the western Euro-North African coasts is the halitheriine Metaxytherium medium. MMM unnum. differs from Metaxytherium in having a horizontal ramus relatively slender [c. 128(0)], with a ventral border moderately concave, sharply downturned anteriorly [c. 122(1)], and permanent premolars present and single-rooted [c. 157(1)]. These characters also exclude MMM unnum. from the Dugonginae.
On the contrary, all the characters scored for MMM unnum. are consistent with those of the miosirenine trichechid Anomotherium langewieschei: a ventral border of horizontal mandibular ramus moderately concave, sharply downturned anteriorly [c. 122(1)]; accessory mental foramina absent [c. 123(1)]; a horizontal ramus relatively slender [c. 128(0)]; canines absent [c. 144(1)]; supernumerary molars absent [c. 150(0)]; a postcanine dental formula reduced from P/p1-5, M/m1-3, by loss of anterior premolars [c.155(2)], permanent premolars single-rooted [c. 157(1)]; a masticating surface
rectangular in shape and a mandibular deflection of about 60° (about 50° for Anomotherium langewieschei).
The only other miosirenine trichechid known is the Early Miocene Miosiren kocki. Since the mandible of this latter species is unknown, only the dental characters can be compared with MMM unnum.: [c. 144(1)] [c. 150(0)] [c.155(2)] [c. 157(1)]. All of these characters are consistent with MMM unnum; but the latter differs from Miosiren kocki in having a greater rostral deflection (about 60° for the MMM unnum. mandible versus 49° for the rostrum of M. kocki) and a third molar molariform and not reduced or rounded.
In conclusion, MMM unnum. seems substantially similar to Anomotherium langewieschei (Fig. 6); therefore it could be referred to the Trichechidae and in particular to the Miosireninae, a subfamily of trichechids with a fossil record confined to the European North Atlantic coast from the Late Oligocene to the Early Miocene. According to this interpretation, the miosirenine record extends to the Middle Miocene (Serravallian). This datum could also be supported by the partial skull (IM unnum.) described as Miosiren canhami (Flower, 1874) and considered by Domning (1996) as a possible synonym of M. kocki, possibly reworked from Miocene deposits into the Red Crag Formation of England.
Concerning the validity of the species Prohalicore dubaleni, there are no unambiguous differences between Prohalicore dubaleni and Anomotherium langewieschei; but Prohalicore is considered to be Serravallian in age, while Anomotherium is Chattian. This wide difference in age and the fragmentary nature of MMM unnum. do not permit the exact systematic position of Prohalicore dubaleni to be established.
Fig. 6: Mandibles in lateral view of a) Prohalicore dubaleni (MMM unnum) (modified from Flot, 1886) and b) Anomotherium langewieschei (modified from Siegfried, 1965). Scale bar: 5 cm.
4.3.3 “HALITHERIUM” BELLUNENSE
Order SIRENIA Illiger, 1811 Family DUGONGIDAE Gray, 1821
Subfamily DUGONGINAE (Gray, 1821) Simpson, 1932 Genus “Halitherium” Kaup, 1838
“Halitherium” bellunense Zigno, 1875 (Plates 3-4; Figures 7-11)
Halitherium bellunense Zigno, 1875, Mem. Ist. Veneto Sci. Lett. Arti. 18: 438.
Metaxytherium bellunense (Zigno) Lepsius, 1882, Abh. Mittelrheinischen Geol. Ver. 1: 180.
Holotype – The following bones belonging to the same individual: MGPD-18Z
skullcap; MGPD-19Z left premaxilla with fragment of tusk; MGPD-20/1Z fragment of left maxilla with DP4-M2; 22Z left zygomatic process of squamosal; 23Z right zygomatic process of squamosal; 7358/9Z rib fragment; MGPD-7362Z vertebral corpus; MGPD-7363Z rib fragment; MGPD-7364Z rib fragment; MGPD-7366Z rib fragment; MGPD-7367Z vertebral corpus; MGPD-7368Z vertebral corpus; MGPD-7369Z vertebral corpus; MGPD-7373Z mandibular fragment with a molar fragment; MGPD-7374Z vertebral fragment; MGPD-7375Z vertebral fragment; MGPD-7376Z vertebral fragment; MGPD-7381Z rib fragment; MGPD-7383Z vertebral corpus; MGPD-7384Z fragment of left jugal; MGPD-7385/6Z fragment of right maxilla with DP5-M1; MGPD-7387Z fragment of tooth.
Type locality – Cavarzana, Valle delle Guglie, near Belluno, northern Italy. Glauconitic
sandstones (“Glauconia”), basal portion of Belluno Glauconitic Sandstone Formation. Chattian in age.
Referred specimens – Only the holotype is known.
Range – Known only from Chattian deposits of Belluno (northern Italy).
Emended diagnosis –Dugongine dugongids retaining the following primitive features:
a frontal roof flat between temporal crests [c. 42(0)]; a ventral rim of orbit not distinctly overhanging the lateral surface of jugal [c. 90(0)]; a tusk with enamel on all sides [c. 136(0)]; and a tusk alveolus about half the length of the premaxillary symphysis [c. 140(1)]; but having the following derived features, common with derived dugongines: a nasal process of premaxilla short [c. 7(1)], with posterior end broadened and bulbous [c.
6(2)]; a ventral extremity of jugal ventral to orbit [c. 85(2)]; and a postorbital process of jugal present [c. 86(1)].
Remarks – The specimens were described by Zigno (1875), who founded the new
species Halitherium bellunense on them. In 1882 Lepsius revised this species and considered it referable to Metaxytherium, but Zigno (1887) maintained the binomial Halitherium bellunense. In 1905 Abel redescribed its molars in detail, but left unresolved the matter of its taxonomic position. In 1989 Domning proposed that Halitherium bellunense could be a rytiodontine instead of a halitheriine; in 1994 he included the Rytiodontinae in the subfamily Dugonginae and in 1996 he considered “Halitherium” bellunense as Dugonginae incertae sedis.
Description - The following observations supplement the descriptions of Zigno (1875)
and Abel (1905).
Premaxilla – (MGPD-19Z) Only the dorsal posterior portion of the left premaxilla is preserved. The fragment is 178 mm long. A small tusk is present (see below for the description of the tusk and its alveolus). The nasal process is 25 mm in width mediolaterally, just 84 mm long [c. 7(1)], and dorsally flat, but its posterior end is broadened and bulbous, about 50 mm in width, with a postero-lateral oblique joint surface [c. 6(2)] partially encrusted by sediment. The symphysis is laterally compressed, with a low middorsal ridge anteriorly and a low boss posteriorly, visible in lateral view [c. 10(1)].
Fig. 7: Premaxilla of “Halitherium” bellunense (MGPD 19Z) in dorsal view with broken tusk. Scale bar: 5 cm.
Frontal – (MGPD-18Z) Only the posterior portion of the frontals is preserved. The roof is flat and anteriorly slightly convex in the centre [c. 42(0)]. The lateral sides are partially eroded and the temporal crests seem to be limited to the parietals. The interfrontal suture is not visible.
Parietal – (MGPD-18Z) The parietals bear massive, rounded, lyriform temporal crests with lateral edges that are thin posteriorly. The crests show their maximum development in the centre with a width of 29 mm; they diverge anteriorly and posteriorly and become progressively lower and thinner (TYPE B). The apexes of the crests are separated by a minimum of 35 mm across a median valley about 20 mm wide. An anteromedially direct groove separates the posteromedial side of each crest from the nuchal crest. The posterior portion is partially eroded and the nuchal crest appears to be indistinguishable from the external occipital protuberance.
The parietal roof is moderately concave posteriorly and flat anteriorly [c. 45(1)]. A sagittal crest is absent [c. 51(1)]. The parietals are 73 mm long in the median line, 75 mm wide posteriorly, narrower anteriorly at 73 mm in the centre and 68 mm at the anterior end. A intertemporal constriction seems to be absent [c. 46(2)]. The suture with the frontal forms an angle of about 94°. No interparietal suture is visible. The cranial portion of the squamosal is absent but the lateral sutures of the parietals indicate that the squamosal does not reach the level of the roof and does not indent the parietal [c. 76(0)].
Supraoccipital - (MGPD-18Z) Only the dorsal portion of the supraoccipital is preserved. It meets the parietal roof at an angle of 115°. The dorsolateral corner is moderately rounded. The external occipital protuberance is partially eroded but well developed, and rises above the plane of the parietal roof. The median crest below it is distinct, but low. The areas of insertion for semispinalis capitis muscles are flattish and oval.
Maxilla – (MGPD-20/1Z left; MGPD-7385/6Z right) Only two maxillary fragments are preserved with molars associated. On the left side the base of the zygomatic-orbital bridge is preserved: 42 mm long, thicker posteriorly (17 mm) than anteriorly (12 mm), and elevated about 1 cm above the alveolar margin; while on the right side just a posterior fragment of the base of the zygomatic-orbital bridge is preserved.
Fig. 8: Skullcap of “Halitherium” bellunense (MGPD 18Z) in dorsal view. The broken line indicates the supposed frontoparietal suture. Scale bar: 5 cm.
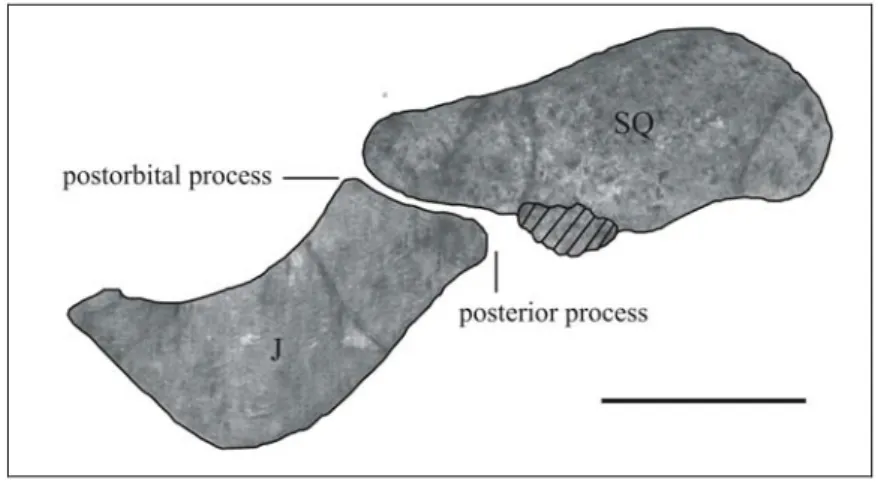
Squamosal – Only the zygomatic processes of the squamosals are preserved (MGPD-22Z left and MGPD-23Z right). The left bone is 101 mm long, the right one 108 mm. The zygomatic process is lozenge-shaped, much broader posteriorly (38 mm) than anteriorly (18 mm) [c. 83(1)]. The dorsal and ventral margins are narrow and rounded. The medial side is not swollen and is slightly concave, and is moderately inclined inward dorsally. The posterodorsal edge is slightly convex in outline. The processus retroversus is moderately developed, moderately inflected, not projected below line of suture with jugal [c. 77(1)]; its posterior end is smooth, without protuberances. The base of the zygomatic root is 50 mm long and 17 mm high.
Jugal – (MGPD-7384Z) Only the left jugal is preserved, lacking the anterior and posterior extremities; it is 97 mm long and 38 mm high. The ventralmost point seems to lie under the centre of the orbit [c. 85(2?)]. The ventral tip is rounded but not thickened; the bone thickens dorsally and the ventral margin of the orbit is concave and thick, but not overhanging [c. 90(0)]. The anterior portion is narrow. The postorbital process in front of the tip of the squamosal is 1 cm high and prominent [c. 86(1)]. The zygomatic process is incomplete; the preserved suture with the squamosal is 36 mm long; observing the corresponding suture on the squamosal, it is estimated to have been on the whole about 50 mm long; therefore with a posterior process about 14 mm long [c. 89(0)].
Fig. 9: Left zygomatic arch of “Halitherium” bellunense (7384Z and MGPD-22Z) in lateral view. The stripped area indicates a possible portion of the left posterior process of the jugal. Scale bar: 5 cm.
Mandible - (MGPD-7373Z) Only a mandibular fragment with a molar fragment is preserved. On the ventral side the dorsal surface of the mandibular canal is visible.
Dentition – The upper dentition is represented by a fragment of the left tusk in the premaxilla (MGPD-19Z); and by the following upper molars that I interpret as the left DP4-M2(MGPD-20/1Z); and the right DP5-M1(MGPD-7385/6Z); in contrast to Zigno (1875) and Abel (1905), who interpreted them as the left DP5-M3(MGPD-20/1Z); and the right M1-M2(MGPD-7385/6Z). The upper dental formula is interpreted to be I 1, C 0, DP 3?-5, M 1-3 [c. 139(0); c. 143(1); c. 144(1); c. 150(0); c. 155(2); c. 157(2)]. The lower dentition is represented only by a molar fragment in the mandibular fragment (MGPD-7383Z). An isolated molar fragment is also preserved (MGPD-7387Z).
Only the base of the left tusk is preserved at about 44 mm from the anterior extremity of the mesorostral fossa [c. 139(0)]. It is anterolaterally inclined about 20º with respect to the medial plane; not strongly curved, lens-shaped in cross-section, with sharp anterior and posterior edges [c. 141(1)] and diametres of 18x8 mm; the length of the exposed portion is about 20 mm. The enamel is dark, shiny, thin and present on the entire tooth surface [c. 136(0)]. In Zigno’s description (1875) the tusk is complete, small, about 30 mm long, not worn and probably not erupted (Fig. 10). The tusk alveolus appears to be at least about half the length of the symphysis [c. 140(1 or 2?) scored as 1].
Fig. 10: Premaxilla of “Halitherium” bellunense (MGPD-19Z) in lateral view with the complete tusk from Zigno (1875).
The cheek-tooth enamel is smooth, and about 1 mm thick.
The description of the cheek teeth is based on the left molars, which are better preserved than the right ones.
A small tooth fragment on the lingual side anterior to the DP5could represent the fourth left deciduous premolar.
The left DP5 is incomplete and heavily worn. The roots are not visible.
The left M1 is completely erupted. The roots are not visible. On the whole, it does not differ from the M2.
The left M2 is unworn and not completely erupted. The roots are not visible. The precingulum is arched, low and narrow. The anterior cingular valley is lingually blocked by the precingulum that meets the protoloph; it is anterolabially open, but it does not reach the labial side because of the anterior accessory cuspule of the paracone. The protoloph bears three aligned lingually-inclined cusps: the paracone on the labial side, the protoconule in the centre, and the protocone on the lingual side. The transverse valley is deep and open on the both sides. The metaloph also bears three aligned cusps: the metacone and the central metaconule are lingually inclined, but on the contrary, the hypocone is labially inclined and nearly meets the metaconule. The postcingulum is confluent with the hypocone lingually and diminishes in height labially. The posterior cingular valley is open labially.
This tooth is interpreted as M2 instead of M3, in contrast with Zigno (1875) and Abel (1905), because of its position relative to the posterior margin of the zygomatic-orbital bridge of the maxilla, and its rounded shape in contrast with the typical elongated shape of M3. Moreover, considering the tooth eruption sequence, the presence of an unerupted tusk is inconsistent with a partially erupted M3.
Fig. 11: “Halitherium” bellunense (MGPD-20/1Z) a) left M1 and b) left M2 in occlusal view; c) portion of left maxilla with M1 and M2 in lingual view. Scale bar: 3 cm.
Vertebrae – Three thoracic vertebral centra (7367Z, 7368Z, MGPD-7369Z), a lumbar vertebral centrum (MGPD-7383Z) and four vertebral fragments, (MGPD-7362Z, MGPD-7374Z, MGPD-7375Z, MGPD-7376Z) are preserved.
The centra have flat anterior and posterior faces. The transverse processes are not preserved.
Ribs – Four fragments of small anterior ribs (7363Z, 7364Z, MGPD-7366Z, MGPD-7381Z) and a fragment of a larger rib (MGPD-7358/9Z) are preserved. MGPD-7358/9Z is a distal fragment of a large rib about 240 mm long, elliptic in cross-section (50 x 29 mm), growing thinner distally, with a cortical surface about 5 mm thick. It is covered by cemented gravels and shells. The lateral surface is slightly convex, the medial one concave and the anterior and posterior ones rounded.
MGPD-7366Z is a proximal fragment, about 128 mm long, of an anterior left rib, probably the first.
It is elliptic in cross-section. Its capitulum is anteroposteriorly flattish, the tuberculum is moderately prominent and an anteromedial flange is present at about 45 mm from the tuberculum on the underside of the neck.
MGPD-7363Z is a central fragment of an anterior right rib, 135 mm long, elliptic in cross section.
MGPD-7364Z is a proximal fragment of an anterior left rib, 147 mm long, elliptic in cross-section, smaller than MGPD-7366Z with the capitulum narrower.
MGPD-7381Z is a proximal fragment of an anterior left rib, 66 mm long, elliptic in cross-section, with a small protuberance for the iliocostal muscle insertion.
Comparison - Comparison with other sirenian species is difficult because the holotype
belongs to a very juvenile individual, as shown by the dentition (the tusk is unworn and probably not erupted, and the M2 is unworn and not completely erupted), and because it is incomplete and lacking data for several characters.
Nonetheless we can observe that:
“H.” bellunense differs from Trichechidae in having a nasal process of premaxilla broadened and bulbous at the posterior end [c. 6(2)], a squamosal that does not indent the posterolateral corner of parietal [c. 76(0)], a tusk lens-shaped in cross section [c. 141(1)], and in particular a processus retroversus of squamosal [c. 77(1)], contrary to the primitive state of this character in the Trichechidae.
“H.” bellunense has a more or less flat frontal roof [c. 42(0)]; and tusks with enamel on the entire tooth surface [142(0)], in common with the other Oligocene dugongids.
“H.” bellunense differs from the contemporaneous dugongine Crenatosiren olseni in having a short nasal process of premaxilla [c. 7(1)], with posterior end broadened and bulbous [c. 6(2)]; a squamosal that does not indent the posterolateral corner of parietal [c. 76(0)]; a ventral extremity of jugal ventral to orbit [c. 85(2)]; a postorbital process of jugal present [c. 86(1)]; an orbital rim of jugal not overhanging [c. 90(0)]; and tusks lens-shaped in cross-section [c. 141(1)].
“H.” bellunense differs from the genus Halitherium and all other Halitheriinae in having a short [c. 7(1)], nasal process of premaxilla with posterior end broadened and bulbous with a postero-lateral oblique joint surface [c. 6(2)]; and tusk lens-shaped in cross-section, with sharp anterior and posterior edges [c. 141(1)]. The short and broadened nasal process of the premaxilla [c. 6(2) and c. 7(1)] is present in the most derived Dugonginae, including the Early Miocene genus Rytiodus, the only dugongine
genus known along the Euro-North African coasts (see Bajpai & Domning, 1997); while modified (flattened) tusks are observed only in the dugongines.
Moreover, “H” bellunense has a postorbital process of jugal [c. 86(1)] like the dugongines Rytiodus sp. and Xenosiren yucateca.
Nevertheless, of the three synapomorphies that characterize the Dugonginae (Bajpai & Domning, 1997), just the orbital rim of jugal is scored for H. bellunense, but it appears to be not overhanging [c. 90(0)] in contrast with those of the dugongine species. Due to the very young age of the holotype and the loss of the anterior portion of its premaxilla, no observation can be made on the exact tusk size and alveolus length; I can only observe that the tusk alveolus appears to be at least about half the length of the symphysis [c. 140(1 or 2)].
In conclusion, I agree with Domning (1989b, 1996) in considering “H.” bellunense a primitive dugongine instead of a halitheriine, because of its short nasal process of premaxilla with posterior end broadened and bulbous with a postero-lateral oblique joint surface [c. 6(2), c. 7(1)] and its tusk lens-shaped in cross-section, with sharp anterior and posterior edges [c. 141(1)].
The attribution of “H”. bellunense to the subfamily Dugonginae provides new information about the palaeobiogeography and relationships of the basal Dugonginae.
4.3.4. THE GENUS RYTIODUS
Order SIRENIA Illiger, 1811 Family DUGONGIDAE Gray, 1821
Subfamily DUGONGINAE (Gray, 1821) Simpson, 1932 Genus Rytiodus Lartet, 1866
Rytiodus Lartet, 1866, Bull. Soc. Géol. France (2)23: 282.
H[alitherium]. (Lartet) Cope, 1883, Proc. Acad. Nat. Sci. Philadelphia 1883: 52. Rytiodus (Lartet) Woodward, 1885, Quart. Jour. Geol. Soc. London 41: 470. Thelriope (Lartet) Pilleri, 1987, Sirenia of the Swiss Molasse: 65.
Type – Rytiodus capgrandi Lartet, 1866
Included species – Rytiodus capgrandi Lartet, 1866; Rytiodus sp. Heal, 1973. Range – Aquitanian-Burdigalian, SW France and Libya.
Emended diagnosis – Dugongine dugongids with large tusks broad and extremely
flattened mediolaterally [c. 141(2)], with enamel mainly on medial side [c. 136(2)], characterized by exoccipitals which do not meet in a suture [c. 66(1)], nasals relatively large, separated in midline by nasal incisure [c. 31(1)], and parietal roof constricted at the centre [c. 46(1)].
Rytiodus capgrandi Lartet, 1866 (Plates 5-6)
Rytiodus Capgrandi Lartet, 1866, Bull. Soc. Géol. France (2)23: 282.
H[alitherium]. capgrandi (Lartet) Cope, 1883, Proc. Acad. Nat. Sci. Philadelphia 1883: 52.
Rytiodus Capgrandi (Lartet) Woodward, 1885, Quart. Jour. Geol. Soc. London 41: 470. Thelriope capgrandi (Lartet) Pilleri, 1987, Sirenia of the Swiss Molasse: 65.
Syntypes – MNHN MBA 1-9, two pairs of tusks of different individuals, plus two rib
fragments and three cranial fragments.
Type locality – Bournic, Lot-et-Garonne, southwestern France. “Calcaire marin
coquillier (étage de Bazas)”. Aquitanian – early Burdigalian in age.
Referred specimens – Specimens listed in Appendix 2.
Range - Known only from Aquitanian – lower Burdigalian deposits of the Aquitaine
Basin (SW France).
Emended diagnosis – Rytiodus in which the contact between premaxilla and frontal is
preserved [c. 9(1)], and the infraorbital foramen is moderately large [c. 13(1)].
Remarks – The new genus and new species Rytiodus capgrandi were founded by Lartet
(1866) on the basis of two pairs of tusks and some rib and cranial fragments found at Bournic (Lot-et-Garonne, SW France).
In 1880 Delfortrie referred to Rytiodus capgrandi a skeleton, about 5 m long, found in 1861 at Saint-Morillon (Gironde, SW France). This specimen is now missing and I have to base my observations on illustrations and descriptions made by Delfortrie (1880); following Domning (1978b), who remarked that the rostrum of this specimen, described by Delfortrie (1880) as undeflected, was erroneously restored.
After 1880, just three other tusks have been referred to Rytiodus capgrandi; they were also found in the Aquitanian deposits of Gironde (SW France).
Sintype description – Of the specimens described by Lartet (1866) just the two pairs of
restored tusks (MNHN MBA 6-9), two rib fragments (MNHN MBA 4-5) and three cranial fragments (MNHN MBA 1-3) are preserved.
MNHN MBA 1 is a parietal fragment 38 mm long in the midline, 85 mm long on the left side, 41 mm wide at the frontoparietal suture and about 68 mm wide posteriorly [c. 46(1)]. The cranial vault is nearly rectangular in coronal section anteriorly, about 27
mm thick in midline and with the lateral sides nearly vertical. The temporal crests are incomplete, but they appear to be lyriform, low, smoothly rounded, not overhanging laterally, and separated by about 30 mm (TYPE C from Domning, 1988), with a flat roof, just slightly convex posteriorly [c. 45(2)]. A sagittal crest is absent [c. 51(1)]. On the ventral side a low bony falx is present [c. 47(1)].
MNHN MBA 2 is a basisphenoid-basioccipital fragment, 63 mm long, 75 mm wide at the level of the pterygoid processes, 26 mm thick in the midline anteriorly and 34 mm posteriorly; with the basisphenoid-basioccipital suture perfectly fused. The ventral side is posteriorly convex, bearing a rugosity for the longus capitis muscles, and anteriorly flat and 31 mm wide between the pterygoid processes. Only the bases of these processes are preserved, 31 mm long anteroposteriorly and 17 mm wide mediolaterally, with a longitudinal groove on the posterior side. The dorsal side is slightly inclined anteroventrally with two longitudinal grooves separated by a narrow median crest. MNHN MBA 3 is a right exoccipital fragment, 67 mm high. The sutural surface for the supraoccipital is rugose, 36 mm long, 27 mm thick medially and 18 mm thick laterally. On the dorsomedial side there is no sutural surface for the left exoccipital, so they could be considered separated in midline [c. 66(1?)]. The dorsolateral border has a smoothly rounded and slightly overhanging posterior edge [c. 70(0)]. The dorsal portion of the right condyle is preserved, with a deep supracondylar fossa extending across the entire width of the occipital condyle [c. 67(1)].
MNHN MBA 4-5 are two rib fragments, lens-shaped in cross section. They seem not to differ from other dugongid ribs.
MNHN MBA 6-7 are a pair of nearly complete large tusks, about 255 mm long, mediolaterally flattened, about 77 mm wide anteroposteriorly, but only 24 mm in mediolateral thickness [c. 141(2)]. They are curved outward, with the medial surface convex and the lateral surface concave, with a longitudinal shallow groove. The tips are restored. The tusks are characterized by about forty white transverse sulci on the lateral and on the medial surfaces, slightly arched with the concavity directed proximally, which become more closely spaced posteriorly. Longitudinal lines are also present. A thin layer of enamel, less than 1mm thick, is present on the whole medial side of the tusk [c. 136(2)]. The crowns are not distinct from the roots [c. 137(1)]. The roots seem to be closed.
MNHN MBA 8-9 are another pair of tusks, lacking the tips. They are slightly smaller than MNHN MBA 6-7, but they show the same features.
The transverse sulci, which characterize the tusks, are interpreted as external growth layers found also in some tusks of Dugong dugon (e.g. Marsh, 1980: 188 fig 9B).
Description of referred specimens – Of the three specimens referred to Rytiodus
capgrandi, I personally examined the tusk (NHMB O.G.916) from Courtebotte studied by Pilleri (1987) and a cast (MNHN unnum.) of a tusk from L’Ariey.
NHMB O.G.916 is a large tusk lacking the distal portion; it is 180 mm in length, 68 mm in anteroposterior width, but only 25 mm in mediolateral width [c. 141(2)]. It is slightly curved outward, with the medial surface convex and the lateral surface concave. The tip is incomplete, with a large worn surface on the lateral side, 80 mm wide, smooth, and more concave than the remainder of the lateral side.
The tusk is characterized by ten white transverse sulci on the lateral and on the medial surfaces, slightly arched with the concavity directed proximally. A thin layer of enamel, less than 1mm thick, is present on the medial side of the whole tusk [c. 136(2)]. In cross-section the dentine does not seem to have accretion lines, but this could be due to diagenetic processes. Longitudinal lines are also present.
MNHN unnum. is a cast of an incomplete tusk with an open root. It shows the same shape as the tusks described above, but apparently without the transverse sulci; only the longitudinal lines are present.
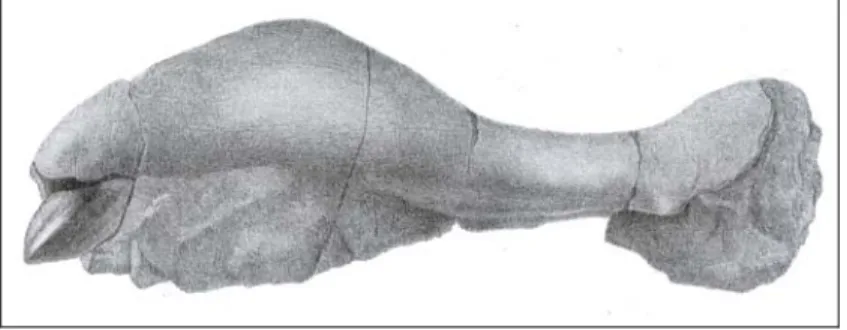
Rytiodus sp. Heal, 1973 (Plates 7-10; Figures 12-16)
Locality – Gebel Zelten, Libya. Formation – Marada Formation. Age – Aquitanian - Burdigalian.
Referred specimens – NHM M45674a-c skull, endocranial cast, associated vertebrae
and ribs; BRSUG 20853a-e skull, endocranial cast, associated vertebrae and ribs; BRSUG 20854 maxillary fragment; BRSUG 20855a-b two right tympanics, BRSUG 20856 left mandibular ramus with m3; BRSUG 20857 three posterior thoracic vertebrae.
Range – Known only from the Aquitanian - Burdigalian deposits of Gebel Zelten,
Libya.
Emended diagnosis – Rytiodus with a large infraorbital foramen [c. 13(2)], a preorbital
process of jugal relatively flat and thin [c. 88(1)], and without the contact between premaxilla and frontal [c. 9(0)], because of the lacrimal inserted between the posterior extremity of the nasal process of premaxilla and the anterior extremity of the supraorbital process of frontal.
Remarks – These specimens were described as a new species of Rytiodus by Heal
(1973) in his unpublished PhD thesis. The cladistic analysis of the Dugonginae carried out by Bajpai and Domning (1997) included this species in a clade that also comprised Rytiodus capgrandi and Corystosiren varguezi.
Description – The following description of the cranial anatomy is based on the NHM
specimens personally examined, on photographs of BRSUG 20853a made by Simon Powell (University of Bristol), and on Heal’s illustrations. Both crania preserved are incomplete and damaged, with ventral side worn . Some features absent from one of the cranium tend to be present on the other. NHM M45674a is the most complete posteriorly and BRSUG 20853a is the most complete anteriorly; even if the premaxilla is incomplete and damaged in both crania. BRSUG 20853a is best preserved and most sutures are evidently identifiable.
Premaxilla –The rostrum is incomplete; the anterior and palatal surfaces are not preserved. The exact length of the premaxillary symphysis can not be established, but the rostrum is enlarged relative to the cranium [c. 3(1)]. NHM M45674a preserves both
lateral sides, but these are incomplete: the left one is broken and the right one is separated and enclosed in matrix, so that the alveoli are exposed and the posterior portions of the tusks are preserved and visible on both sides. BRSUG 20853a preserves only the incomplete left lateral side, smooth, slightly convex and ventrally overhanging. In anterior view the posterior portions of both alveoli with tusk fragments are visible. The large incisor alveoli extend posteriorly along the symphysis [c.140(2)] (see below for description of the tusks and their alveoli).
The premaxillary symphysis is laterally compressed, bearing a middorsal ridge anteriorly and with its posterior end upraised to form a boss in lateral view [c. 10(1)]. The nasal process is short [c. 7(1)], in contact with maxilla and lacrimal [c. 1(1)], but not with jugal; it is broadened and bulbous at posterior end, with a nearly vertical posterior joint surface [c. 6(2)]. This posterior joint surface is exposed in both crania and asymmetric in the holotype. It contacts on the medial side the anterior extremity of nasal and on the lateral side the lacrimal; this last intrudes between the posterior extremity of the nasal process of premaxilla and the anterior extremity of the supraorbital process of frontal, so that the dorsal contact between premaxilla and frontal is lost [c. 9(0)].
The anterior end of the mesorostral fossa is rounded and broad, without a constriction. The rostral deflection is estimated to be around 62°.
Nasal – The nasals are long and narrow, separated in the midline [c. 31(1)] by the long nasal incisure; the surfaces of contact with the supraorbital processes of the frontals are nearly vertical.
Ethmoidal Region and Vomer – Not visible, because the mesorostral fossa is completely filled by matrix.
Lacrimal – The lacrimals are relatively large, dorsally subtriangular in shape, and without a nasolacrimal canal [c. 91(1)]. The best preserved is the left lacrimal of BRSUG 20853a. It rests in a socket at the upper end of the maxilla and is dorsally exposed between premaxilla, frontal and maxilla; its narrow posterolateral corner extends ventrad between the maxilla and the frontal, reaching the anterodorsal surface of the orbital ring.
Frontal – The mesorostral fossa is about 95 mm wide in the centre and more than 200 mm long, being prolonged backward by a nasal incisure about 30 mm wide and 45 mm long [c. 37(2)]. The posterior end of the mesorostral fossa (including the nasal incisure) lies about 90 mm behind the anterior ends of the supraorbital processes, so the external nares are retracted and enlarged, reaching beyond the level of the anterior margin of the orbit [c. 8(2)]. The supraorbital process is long, massive, dorsoventrally about 40 mm thick [c. 36(1)], with the anteromedial border curving downward and outward and the dorsal surface sloping ventrolaterad at an angle of about 60° to the horizontal plane [c. 43(1)]. In the holotype the right posterolateral corner is broken and the internal cancellous bone is exposed. On the lateral side the lamina orbitalis is clearly discernible [c. 38(0)]. The dorsal surface bears a prominent, thick ridge continuous with the temporal crest; these ridges diverge anteriorly and reach the anterior margin of the supraorbital process. An internasal process is absent; the nasals are separated in midline by the long nasal incisure, and just two slender anterior laminae of the frontals are extended on the medial side of the nasals. The frontoparietal suture is W-shaped. The frontal roof is concave, sloping steadily ventrad to the anterior margin [c. 42(1)], and about 63 mm long in the midline.
Fig. 12: Cranium of Rytiodus sp. from Libya (BRSUG 20853a) in dorsal view. Frontoparietal suture not discernable. Scale bar: 5 cm.
Parietal – The parietals are well preserved only in the holotype. They bear massive, rounded, lyriform, prominent but not laterally overhanging temporal crests with lateral sides concave. The crests are anteriorly about 12 mm wide, and separated by a minimum of 4 mm across a flat median valley. Anteriorly they are in continuity with the
prominent frontal ridges, while posteriorly they diverge and seem to be separated from the nuchal crest by a narrow and shallow anteromedially-directed groove visible only on the right side. The intertemporal region is slightly constricted at the centre of the braincase [c. 46(1)]. The parietal roof is flat [c. 45(1)], and about 75 mm long in the midline. A sagittal crest is absent [c. 51(1)]. The interparietal suture is fused.
Internally, the internal occipital protuberance is low and the bony falx cerebri is low and short [c. 47(1)]; it does not reach the frontoparietal suture. A medial boss is present at the joint between the internal occipital protuberance and the bony falx cerebri.
Fig. 13: Posterior portion of cranium of Rytiodus sp. from Libya (NHM M45674a) in dorsal view. The broken lines indicate supposed sutures. Scale bar: 5 cm.
Supraoccipital – The supraoccipital is preserved only in the holotype. It is subrectangular in shape, about 100 mm wide dorsally, 81 mm wide ventrally, and 66 mm high (ratio of width to height is about 1.5). It meets the parietal roof at an angle of about 115°. The rugose insertions for the semispinalis capitis muscles are well developed, concave and nearly round. The nuchal crest is distinct, sharp and concave posterad above the muscle insertions. The external occipital protuberance is well developed and posteriorly overhanging with a prominent median ridge extending below. The sutural surfaces for the exoccipitals meet at an angle of about 144°, but they are separated in midline by a slight median constriction.
Exoccipital – Only the dorsal portion of the right exoccipital is preserved in the holotype, 42 mm wide, 23 mm high and 27 mm thick. The dorsolateral border is rounded, smooth, not flangelike [c. 70(0)]. As observable from the supraoccipital-exoccipital sutures, the complete medial surface of the right supraoccipital-exoccipital and the shape of the endocranial cast, the foramen magnum is peaked and the exoccipitals are separated in midline [c. 66(1)].
Fig. 14: Cranium of Rytiodus sp. from Libya (NHM M45674a) in posterior view. The broken area indicates the sutural surface for the left exoccipital. Scale bar: 5 cm.
Basioccipital – Not preserved.
Basisphenoid, Presphenoid, Alisphenoid, Pterygoid – They are preserved only in the holotype. The occipitosphenoid eminence bears the usual bilateral rugosity. The basisphenoid is robust, with the dorsal side flat and the ventral side convex without median tip. The sella turcica is broad and shallow. No distinct tuberculum sellae or chiasmatic grooves seem to be present. The pterygoid process is preserved only on the left side; its ventral end is eroded and so its total length is not determinable. The posterior side is slightly concave and the lateral and medial sides are smooth, without crests. A pterygoid fossa is present [c. 102(1)]. An alisphenoid canal is absent [c. 101(1)].
Palatine – The palatines extend anteriorly beyond posterior edge of zygomatic-orbital bridge [c. 99(0)]. Their median suture is about 50 mm long and their posterior border is about 1 mm thick and incised, to as far forward as level of M1[c. 97(1)], so that the palate appears incomplete at the level of M2 [c. 16(0)]. The bones continue laterally back towards the pterygoid processes, constituting their anteromedial walls.
Maxilla – The maxilla bears a very wide infraorbital foramen, 36 mm in diametre [c. 13(2) in contrast with Baipaj & Domning, 1997: 13(1)] not obstructed [c. 20(0)]; and filled by matrix; medial to it on the palatal surface there is a prominent palatine foramen. An ill-defined ridge extends from the internal border of the palatine foramen to as far back as the external edge of the internal nares. A small dorsal extension of maxilla contacts the nasal process of premaxilla and overlaps the dorsal surface of the preorbital process of jugal. The ventral surface is partially eroded on the holotype, so that the cheek teeth and their alveoli are poorly preserved; while on BRSUG 20853a it is totally eroded, so that the cheek teeth and the alveoli are not preserved, except for some anterior alveolar remnants. The ventral surface bears a shallow medial palatal gutter that widens anterad at the level of the infraorbital foramen. The palatal premaxilla-maxilla suture is not discernible. The row of tooth alveoli is about 120 mm long and the maximum width between labial edges of left and right alveoli is about 100 mm at the level of M2.
The zygomatic-orbital bridge is about 50 mm long anteroposteriorly and 20 mm thick; its ventral surface is flat and lies about 7 mm above the alveolar margin [c. 11(0)].
Squamosal – Only the dorsal portion of the squamosal is preserved on the left side of the holotype, with its zygomatic process nearly complete. The squamosal reaches the level of the roof of the parietal and slightly indents its outline posterolaterally, but does not reach the summit of the temporal crest [c. 76(0)]. The sigmoid ridge is not preserved. The root of the zygomatic process is 66 mm long anteroposteriorly, inclined anterad and dorsally concave. The zygomatic process is 133 mm long and lozenge-shaped [c. 83(1)], with the medial side relatively concave and the dorsal edge medially inclined. A processus retroversus is present, simple and slightly inflected [c. 77(1)]. A posttympanic process is present [c. 73(1)]. The glenoid surface is broad laterally, but narrows medially, and it is bordered posteriorly by a conspicuous postglenoid process.
Fig. 15: Posterior portion of cranium of Rytiodus sp. from Libya (NHM M45674a) in left lateral view, indicating the zygomatic process of squamosal and a small portion of zygomatic process of jugal. Scale bar: 5 cm.
Jugal – An incomplete right jugal and an anterior portion of the left jugal are preserved on the holotype, while on the BRSUG 20853a only an incomplete left jugal is preserved. The ventral extremity of jugal is eroded, so that the exact position of the ventralmost point can not be established, but it was located below the orbit [c. 85(2)], anterior to the well-developed postorbital process [c. 86(1)]. The ventral rim of orbit distinctly overhangs the lateral surface of jugal [c. 90(1)]. The preorbital process is relatively thick and robust, but anyway the posteromedial-anterolateral breadth of the portion lateral to the maxillojugal suture is wider than its anteromedial-posterolateral thickness [c. 88(1)], and it is dorsally overlapped by the maxilla, so it does not contact the premaxilla [c. 87(0)]. Its lateral surface is smooth with a rounded, anteriorly-projecting flange. The posterior portion is incomplete, but the zygomatic process is estimated to be about 96 mm long, so longer than the diametre of the orbit, estimated to be 69 mm [c. 89(0)].
Fig. 16: Cranium of Rytiodus sp. from Libya (BRSUG 20853a) in left lateral view. Scale bar: 5 cm.
Ear region – Associated with the holotype there are a left malleus, a left incus and an anterior fragment of the left tegmen tympani; moreover two right tympanics (pars temporalis) are preserved in the Bristol collection (BRSUG 20855a-b).
The preserved portion of tegmen tympani bears a well-developed ridge on the internal surface and the attachment for the tympanic.
The distal border of the tympanic is U-shaped, rounded in outline with a blunt tip. The malleus is narrow. The area of orbicular apophysis on posterior end is convex. The processus muscularis is massive, swollen and elliptic in outline. A sharp horizontal ridge runs forward on lateral side from dorsal end of manubrium. The external edge of manubrium is convex. The articulation with the incus is constituted by two facets: the anterior one is larger and flat, the posterior is smaller and saddle-shaped.
The incus bears, on the ventral side, the two articular surfaces for the malleus. The anterolateral edge of crus breve is very slightly convex in outline, the crus longum (processus lenticularis) is more developed, cylindrical, and curves sharply inward.
Mandible – The mandible is lacking on the holotype, but a left mandibular ramus with m3 and incomplete m1-m2 is preserved in the Bristol collection (BRSUG 20856) (see Heal, 1973).
The horizontal ramus is dorsoventrally broad with a minimum height of 86.5 mm [c. 128(1)]. Its ventral border, even if incomplete, appears to be strongly concave [c. 122(3)]. The posterior border of the mental foramen is preserved at the level of
deflection of dorsal edge of ramus. No accessory foramina are present behind it [c. 123(1)]. Remnants of the three molars are preserved, but the anterior portion of the alveolar row is lacking. Just the anterior base of the vertical ramus is preserved, and the anterior border seems to extend anterior to the base [c. 126(1?)].
Endocranial cast – Natural endocranial casts were removed from both the crania. The following description is based on that from the holotype, it being the more complete one.
The endocranial cast is rounded, slightly narrower in the centre, with the cerebral hemispheres bean-shaped in dorsal view and separated by a narrow and low longitudinal sulcus, posteriorly enlarged into a fossa. Anteriorly the olfactory lobes are large and ovoid, Ventrally there are the two roots of cranial nerve II and a slight concavity in the centre with the marks of the complex of nerves III, IV, V and VI. The cerebellar region is anteroposteriorly compressed, and not evidently separated from the cerebral region. Posteriorly there are two lateral concavities (impressions of the exoccipital bones), and the cast of the anterior portion of the spinal cord, subtriangular in transverse section with a dorsal apex.
DENTITION - Besides the pair of tusks, the upper dentition seems to be represented by 2 premolars and 3 molars: I 1, C 0, DP4-5, M 1-3 [c. 139(0); c. 143(1); c. 144(1); c. 150(0); c. 155(2); c. 157(2)]. The molars are well developed.
I - On the holotype, only a posterior medial fragment of the tusk, 32 mm long, is preserved on the left side and a posterior portion of the tusk, 72 mm long, is preserved on the right side; while on BRSUG 20853a just the posterior portion of the right tusk and the posterior ventral portion of the left tusk are preserved in the alveoli.
The tusks are estimated to be more than 78 mm long, about 25 mm thick mediolaterally, and about 54 mm wide anteroposteriorly [c. 141(2)]; they are medially convex and laterally concave. They diverge laterally both anteriorly and posteriorly. The roots are open and the tusks appear to be divided into two laminae mediolaterally compressed, with the labial one thicker than the lingual, on either side of the pulp cavity. A thin layer of enamel seems to be present only on the medial side [c. 136(2)], extending entire length of tusk [c. 137(1)]. Some transverse sulci are present on the surface.
The upper cheek dentition is lacking on BRSUG 20853a, while on the holotype, it is better preserved on the left side. The alveoli and some tooth fragments are preserved and the DP4-M2 are three-rooted, with a longer lingual root and two smaller labial roots.
M3 – It is not completely erupted and not worn. It is rectangular in shape, longer than M2. Its roots are not visible. The enamel is about 1 mm thick. The anterior portion of the crown is broken. The protoloph is confluent with the anterior cingulum lingually. The transverse valley is closed labially. The metaloph is formed by three cusps, with the central metaconule higher than the metacone and hypocone, and lingually inclined. The posterior basin is open posteriorly, with the small posterior cuspules connected to the metaloph both lingually and labially.
m3 – It is preserved in the left mandibular ramus (BRSUG 20856). The occlusal surface is rectangular in outline. The protolophid and hypolophid are worn and appear flat, without remaining cusp pattern; the transverse valley is blocked by contact of a protolophid spur and the crista obliqua. Posteriorly just a hypoconulid cusp is visible on the labial side.
Comparison - Almost all of the characters scored for Rytiodus sp. are consistent with
those scored for Rytiodus capgrandi, except for the larger infraorbital foramen [c. 13(2)] and the loss of contact between premaxilla and frontal [c. 9(0)]. Apart from these two characters the two Rytiodus species can be considered together in comparison with the other sirenian species.
According to Domning (1994) and Bajpai and Domning (1997), the genus Rytiodus belongs to the Dugongidae Dugonginae in having a nasal incisure extending posterior to supraorbital process [c. 37(2)]; a supraorbital process of frontal turned markedly downward, with dorsal surface inclined strongly ventrolaterad and posterolateral corner projecting posteriorly [c. 43(1)]; and the orbital rim of the jugal overhanging [c. 90(1); not scored for R. capgrandi].
Rytiodus is close to the dugongines “Halitherium” bellunense, Corystosiren varguezi, Dioplotherium spp. and Xenosiren yucateca in having: a nasal process of premaxilla broadened and bulbous at the posterior end, having a more or less vertical joint surface in contact with frontal [c. 6(2)], a squamosal not indenting the posterolateral corner of parietal [c. 76(0)], and tusks specialized, not oval in cross section [c. 141(1 or 2)].