62 4. RISULTATI
4.1 Influenza del pH sulle proprietà antibiofilm di hep20
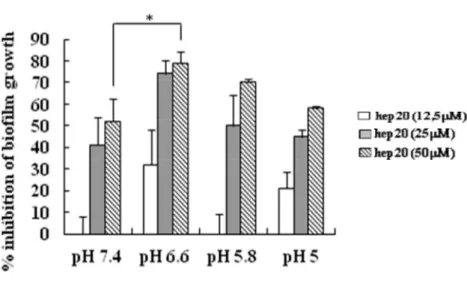
Dati precedenti ottenuti nel nostro laboratorio, hanno dimostrato che il pH è in grado di influenzare in larga misura le proprietà antimicrobiche di hep20 verso cellule planctoniche (Maisetta et al. 2010; Tavanti et al. 2011). Sono stati condotti, pertanto, esperimenti preliminari volti a valutare se il pH potesse anche influenzare l’attività antibiofilm di hep20, utilizzando un ceppo di S. epidermidis (SeBER) arbitrariamente scelto tra quelli oggetto della presente tesi. Come mostrato in Figura 17, quando l’attività antibiofilm di hep20 è stata valutata a differenti valori di pH come percentuale di riduzione della biomassa totale del biofilm rispetto ai controlli non trattati, è stato possibile evidenziare un optimum di attività del peptide a pH 6.6; a questo valore di pH moderatamente acido, corrispondente alle condizioni di pH dell’epidermide dell’uomo, è stata osservata una riduzione di quasi l’80% della biomassa totale del biofilm alla concentrazione più alta di peptide saggiata, riduzione che è risultata statisticamente significativa rispetto a quella osservata a pH 7.4. Sulla base di tali risultati tutti i successivi saggi di inibizione del biofilm sono stati condotti a pH 6.6.
Figure 17. Valutazione della formazione del biofilm di S. epidermidis SeBER in presenza di hep20 (50, 25, 12.5 M) a differenti valori di pH (7.4, 6.6, 5.8, 5) dopo 18 h d’incubazione. La figura indica la percentuale di riduzione della densità ottica (OD) a 570 nm dopo colorazione con cristal violetto, rispetto ai corrispondenti controlli non trattati. Analisi statistica ANOVA seguita dal test Tukey-Kramer, p<0.05:*.
63
4.2 Caratterizzazione fenotipica e genotipica dei ceppi batterici selezionati per lo studio
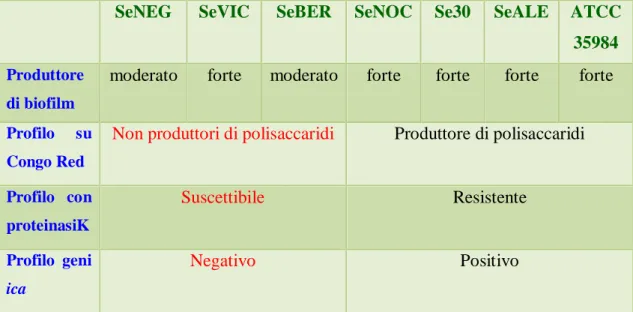
L’attività antimicrobica di hep20 è stata valutata verso sette ceppi di S. epidermidis, una specie batterica frequentemente coinvolta nella contaminazione di dispositivi medici in ambiente nosocomiale.
Sei di tali ceppi sono stati isolati da campioni clinici di pazienti ricoverati presso l’Azienda Ospedaliero-Universitaria Pisana e sono stati identificati con sistemi semi-automatici standard, presso l’Unità Operativa di Microbiologia Universitaria della stessa Azienda. In particolare, tre di tali ceppi (Se30, SeNOC e SeBER) sono stati isolati da segmenti di catetere venoso centrale, pervenuti al laboratorio di Microbiologia per la ricerca di batteri contaminanti, mentre gli altri tre ceppi (SeVIC, SeNEG, SeALE) sono stati isolati da sangue in seguito a positivizzazione di emocolture di pazienti ricoverati presso l’Ospedale sopra menzionato. Tutti i pazienti erano ricoverati in reparti diversi e non risultavano epidemiologicamente correlati gli uni agli altri.
Allo studio è stato aggiunto un ceppo standard di riferimento, S. epidermidis ATCC 35984, noto per la sua spiccata capacità di produrre biofilm (Williams e Bloebaum, 2010).
I ceppi selezionati sono stati sottoposti ad una caratterizzazione fenotipica e genotipica, che includeva la produzione e la natura della matrice extracellulare del biofilm.
4.2.1 Valutazione del profilo di suscettibilità agli antibiotici
Contemporaneamente all’identificazione dei sette ceppi selezionati per lo studio, è stata effettuata la valutazione del loro profilo di suscettibilità a farmaci antimicrobici convenzionali, utilizzati per la terapia delle infezioni da stafilococco. Come mostrato in Tabella 5, tutti i ceppi hanno mostrato resistenze multiple a classi diversi di antibiotici. Tali resistenze si estendevano ad almeno tre diverse categorie di antibiotici, che agiscono, cioè, con distinti meccanismi di azione, ed hanno, pertanto, permesso di definire tutti e sette i ceppi batterici multiresistenti (MDR) (Tabella 5).
64 Codice
identificativo dei ceppi
SeNEG SeVIC SeBER SeNOC Se30 SeALE ATCC 35984 Sito di isolamento C S* C S* C S* C Profilo di antibiotico suscettibilità Gentamicina R R S S R R R Levofloxacina I S R I R R S Tetraciclina S S S R S S S Tobramicina R R R S R NT NT Trimet/sulpham S S S S R I I Clindamicina S S R R NT R R Eritromicina R S R R R R R Acido Fusidico S S S S R R S Nitrofurantoina S S S S S NT NT Fosfomicina R S S S S NT NT Oxacillina R R R R R R R Penicillina G R R R R R R R Rifampicina S S S S S NT NT Teicoplanina NT S S S S S S Vancomicina S S S S S S S Linezolid S S S S S S S Tigecicline S S S S NT S S
4.2.2 Valutazione della capacità di formare biofilm
La capacità di formare biofilm dei sette ceppi batterici oggetto di studio è stata valutata utilizzando un saggio in vitro ampiamente descritto in letteratura (vedi Materiali e Metodi, sezione 3.4). In breve, il saggio prevede la distribuzione di batteri in fase Tabella 5. Profilo di suscettibilità agli antibiotici dei sette ceppi di S. epidermidis selezionati per lo studio.
C: catetere; S*: sangue
65
stazionaria di crescita, opportunatamente diluiti, in piastre di polistirene da 96 pozzetti. Dopo un’incubazione per 24 ore a 37°C, necessaria ai batteri per formare il biofilm, si procede a ripetuti lavaggi ed alla colorazione del biofilm stesso con cristal violetto allo 0.1%. Segue un’estrazione con etanolo puro e la misurazione della densità ottica del sopranatante a 570nm. Sulla base dei valori di densità ottica ottenuti si è proceduto, quindi, ad una classificazione dei ceppi batterici in esame relativamente alla loro capacità di formare biofilm, seguendo i criteri descritti in letteratura da Stepanovic et al, 2000 (vedi Materiali e Metodi, sezione 3.6). Sulla base di tali criteri, i ceppi ATCC 35984, Se30, SeNOC e SeALE sono stati classificati come forti produttori di biofilm, mentre i ceppi SeBER, SeNEG e SeVIC, sono risultati moderati produttori (Tabella 6).
4.2.3 Valutazione della capacità di produrre polisaccaridi extracellulari
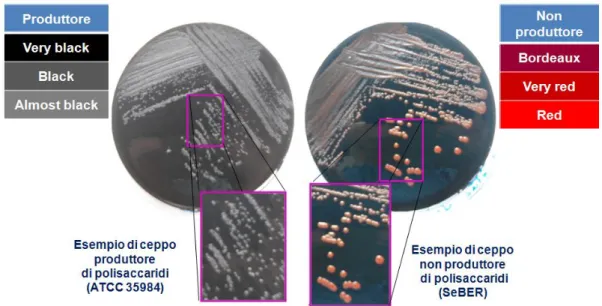
Dopo aver stabilito che tutti i ceppi selezionati erano produttori di biofilm, si è proceduto a valutare la capacità dei ceppi di produrre una matrice extracellulare di natura polisaccadirica. A tale scopo, i batteri sono stati seminati su terreno solido contenente Congo Red e, dopo incubazione per 24 ore a 37°C seguita da un’ulteriore incubazione di 24 ore a temperatura ambiente, è stata esaminata macroscopicamente la morfologia delle colonie. Seguendo una scala di tonalità cromatiche descritta da Arciola e colaboratori (Arciola et al., 2002), sono stati identificati i ceppi produttori di
Codice identificativo dei ceppi
SeNEG SeVIC SeBER SeNOC Se30 SeALE ATCC 35984 Capacità di produrre Biofilm a pH 6.6 OD570mn dopo colorazione con CV 0.76 0.93 0.64 1.66 0.94 1.47 2.21 Categorie in accord con Stepanovic et al. ++ MBP +++ SBP ++ MBP +++ SBP +++ SBP +++ SBP +++ SBP
Tabella 6. Valori di densità ottica a 570 nm, relativi ai sette ceppi di S. epidermidis utilizzati nello studio, dopo 24 ore di incubazione a 37°C in piastre di polistirene. Per ogni ceppo è riportata la classificazione in base alla quantità di biofilm prodotto secondo Stepanovic et al. 2000.
66
polisaccaridi, che presentano una colorazione delle colonie che va dal “quasi nero” al “molto nero” e quelli non produttori che danno origine a colonie con colorazione dal “rosso” al “bordeaux” (Figura 18). Secondo tali criteri, i ceppi ATCC 35984, SeALE, Se30 e SeNOC sono risultati produttori di polisaccaridi, mentre i ceppi SeBER, SeVIC e SeNEG sono risultati non produttori (Tabella 7).
SeNOC Se30 SeALE ATCC 35984 SeNEG SeVIC SeBER
Colonie nero-cristallino, secche Produttori di polisaccaridi (PIA+)
Colonie rosa, cremose Non produttori di polisaccaridi
(PIA-)
Figura 18. Aspetto delle colonie dei ceppi ATCC 35984 (produttore di polisaccaridi) e SeBER (non produttore di polisaccaridi) dopo semina e crescita su terreno solido contenente Congo Red.
Tabella 7. Risultati della valutazione della capacità di produrre polisaccaridi extracellulari da parte dei ceppi oggetto di studio, mediante analisi della morfologia delle colonie su Congo Red Agar.
67
4.2.4 Valutazione della suscettibilità dei biofilm alle proteasi
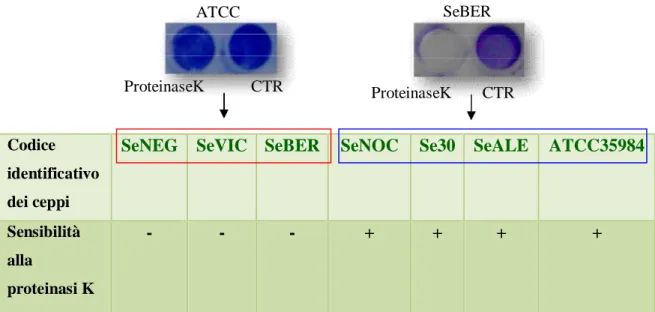
Essendo risultati non produttori di polisaccaridi, è stato ipotizzato che i ceppi SeNEG, SeBER e SeVIC potessero produrre, diversamente dai ceppi ATCC 35984, Se30, SeALE e SeNOC una matrice extracellulare di natura prevalentemente proteica. Per verificare tale ipotesi, è stata valutata la suscettibilità dei biofilm prodotti da tutti e sette i ceppi oggetto di studio, al trattamento per 2 ore con proteinasi K.
Mentre i biofilm prodotti dai ceppi ATCC 35984, Se30, SeALE e SeNOC, produttori di polisaccaridi, non risultavano sensibili all’azione della proteinasi K, i biofilm dei ceppi SeNEG, SeBER e SeVIC apparivano nettamente alterati dal trattamento enzimatico (Figura 19) confermando la natura proteica della loro matrice extracellulare.
Codice identificativo dei ceppi
SeNEG SeVIC SeBER SeNOC Se30 SeALE ATCC35984
Sensibilità alla
proteinasi K
- - - + + + +
4.2.5 Caratterizzazione genotipica dell’operone ica
I sette ceppi di S. epidermidis sono stati caratterizzati anche genotipicamente per la presenza dell’operone ica che, come descritto nell’introduzione della tesi, codifica per prodotti genici responsabili della biosintesi dell’Adesina Polisaccaridica Intercellulare (PIA), un importante componente della matrice extracellulare, coinvolto nella fase di aggregazione del biofilm di S. epidermidis. A tale scopo, il DNA genomico è stato estratto da ogni ceppo batterico ed amplificato tramite PCR con primers specifici per ciascuno dei geni dell’operone ica (icaA, icaD, icaB, icaC) e per la sequenza
Figura 19. Risultati della suscettibilità dei biofilm formati dai ceppi oggetto di studio al trattamento con proteinasi K. (-): biofilm insensibile al trattamento; (+): biofilm sensibile al trattamento.
ProteinaseK CTR ProteinaseK CTR
SeBER ATCC
68
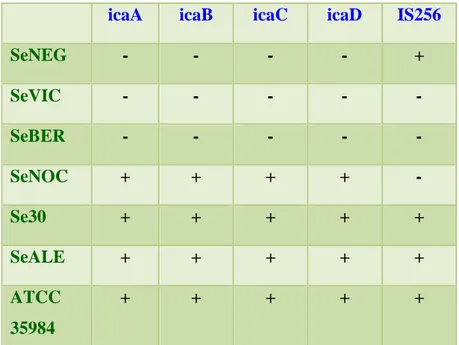
d’inserzione IS256, una sequenza trasponibile la cui inserzione nell’operone ica è stata associata con l’inattivazione dello stesso operone (Ziebuhr et al., 1999). L’analisi degli amplificati su gel d’agarosio (Figura 20) ha indicato per i ceppi ATCC 35984, SeALE, Se30 e SeNOC la presenza di quattro bande di amplificazione corrispondenti ai 4 geni ica; tre dei quattro ceppi (SeALE, Se30, ATCC 35984) erano positivi anche per la sequenza trasponibile IS256 mentre il ceppo SeNOC risultava IS256-negativo (Tabella 8).
Pur essendo presente in 3/4 dei genomi dei ceppi ica-positivi la sequenza IS256 non sembravava interrompere le ORF dei geni ica, dato che le singole bande di amplificazione risultavano delle dimensioni attese (dati non mostrati).
Nel genoma dei ceppi SeBER, SeVIC e SeNEG non è stata rilevata la presenza di alcuno dei geni dell’operone ica; tra questi ultimi ceppi la sequenza IS256 era presente solo nel genoma del ceppo SeNEG (Tabella 8).
Figura 20. Corsa elettroforetica su gel d’agarosio dei prodotti dell’amplificazione dei 7 geni dell’operone ica (ABCD) e della sequenza IS256 del genoma del ceppo ATCC 35984 rappresentato come esempio
69
Complessivamente i risultati della caratterizzazione genotipica dei sette ceppi hanno confermato quelli della caratterizzazione fenotipica e indicano che i ceppi ATCC35984, Se30, SeNOC e SeALE producono una matrice extracellulare di natura prevalentemente polisaccaridica e presentano nel loro genoma i geni ica; tali ceppi possono, pertanto, essere definiti PIA-positivi. Al contrario, la matrice dei ceppi SeBER, SeVIC e SeNEG è risultata di natura prevalentemente proteica; inoltre, poichè non è stata rilevata la presenza dell’operone ica nel genoma di tali ceppi, essi sono stati considerati PIA-negativi. La Tabella 9 riassume le caratteristiche distintive di tutti i ceppi.
icaA icaB icaC icaD IS256
SeNEG - - - - + SeVIC - - - - - SeBER - - - - - SeNOC + + + + - Se30 + + + + + SeALE + + + + + ATCC 35984 + + + + +
Tabella 8. Tabella riassuntiva dei risultati della caratterizzazione genotipica. (+): presenza del gene; (–): assenza del gene.
70
Prima di procedere a valutare l’attività antibiofim di hep20 verso i ceppi di S. epidermidis fino a qui caratterizzati si è voluto determinare la MIC del peptide verso cellule planctoniche di tutti e sette i ceppi in condizioni simili a quelle successivamente utilizzate per i saggi di inibizione della formazione di biofilm. A tale scopo le colture batteriche venivano fatte crescere fino alla fase stazionaria di crescita, una fase in cui lo stato fisiologico delle cellule batteriche è stato assimilato a quello di cellule che crescono in forma di biofilm. La sospensione batterica veniva quindi distribuita in aliquote ed incubata con diluizioni scalari del peptide. Dopo 24 ore di incubazione a 37°C veniva valutata la Minima Concentrazione Inibente (MIC) definita come la più bassa concentrazione di peptide in grado di inibire la crescita batterica e quindi di prevenire l’intorbidimento del mezzo di coltura.
Per tutti e sette i ceppi, la MIC di hep20 è risultata maggiore di 50 M.
4.3 Valutazione della capacità di hep20 di ridurre la biomassa totale del biofilm dei ceppi oggetto di studio
Nella fase successiva dello studio si è proceduto a valutare la capacità di hep20 di inibire la formazione di biofilm dei sette ceppi di S. epidermidis precedentemente caratterizzati. L’effetto del peptide sulla biomassa totale del biofilm, intesa come l’insieme delle cellule batteriche e della matrice extracellulare da esse stesse prodotta, è stato valutato dopo 18 h d’incubazione con diverse concentrazioni di peptide.
SeNEG SeVIC SeBER SeNOC Se30 SeALE ATCC
35984
Produttore di biofilm
moderato forte moderato forte forte forte forte
Profilo su Congo Red
Non produttori di polisaccaridi Produttore di polisaccaridi
Profilo con proteinasiK Suscettibile Resistente Profilo geni ica Negativo Positivo
Tabella 5. Tabella riassuntiva dei risultati della caratterizzazione fenotipica e genotipica dei sette ceppi di S. epidermidis oggetto di studio.
71
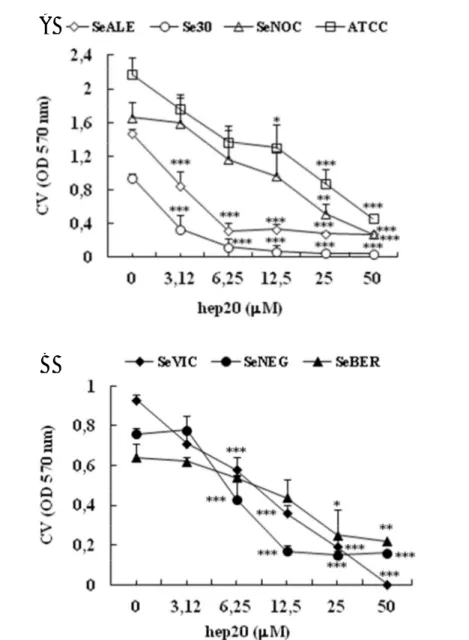
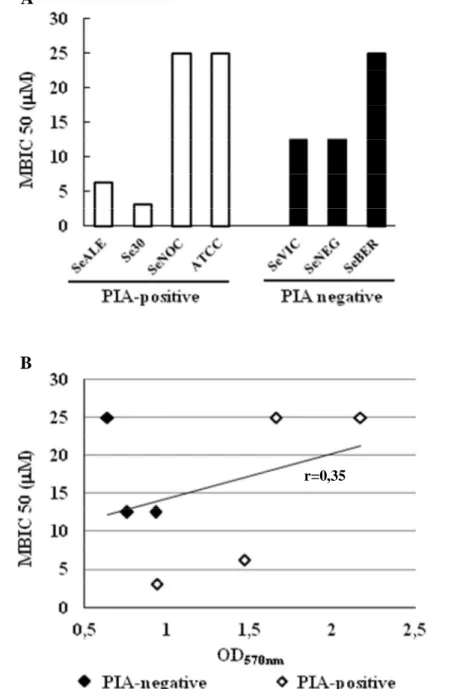
Come mostrato in Figura 21 hep20, usata a concentrazioni al di sotto della MIC, riduceva la biomassa del biofilm sia dei ceppi positivi (Figura 21 A) che PIA-negativi (Figura 21 B) in maniera dose-dipendente. La suscettibilità dei ceppi al peptide è stata valutata analizzando la MBIC50, definita come la concentrazione minima di hep20 in grado di ridurre la biomassa del biofilm del 50% o più. Come mostrato in Figura 22 A, i diversi ceppi hanno dimostrato una certa variabilità nella suscettibilità al peptide con una MBIC50 compresa tra 3.12 M (per il ceppo Se30) e 25 M (per i ceppi SeNOC, SeBER e ATCC 35984). Nonostante ciò, non sono state osservate evidenti differenze di suscettibilità al peptide tra i ceppi positivi e quelli PIA-negativi analizzati; ad esempio, i ceppi più suscettibili al peptide (Se30 e SeVIC) erano rispettivamente un PIA-positivo ed un PIA-negativo (Tabella 7). Non è stata osservata, inoltre, alcuna correlazione tra la suscettibilità dei ceppi batterici a hep20 in termini di MBIC50 e la loro capacità di formare biofilm valutata come OD570nm dopo colorazione con CV (Figura 22 B). Infatti, i ceppi ATCC 35984 e SeBER che sono risultati i meno suscettibili ad hep20, erano anche quelli che producevano rispettivamente la quantità massima (ATCC 35984) e quella minima (SeBER) di biofilm tra i ceppi analizzati.
72
A
B
Figura 21. Riduzione della biomassa del biofilm dei ceppi di S. epidermidis PIA-positivi (A) e PIA-negativi (B) dopo trattamento con hep20. I ceppi batterici distribuiti in piastre da 96 pozzetti venivano trattati con diverse concentrazioni di hep20. La biomassa del biofilm veniva valutata dopo 18h d’incubazione mediante colorazione con CV e misura della OD570nm. Analisi statistica: test ANOVA seguito da
73
Figura 22. Suscettibilità dei diversi ceppi di S. epidermidis a hep20 valutata come Minima Concentrazione Inibente il Biofilm (MBIC) 50 (A) e relazione (B) tra MBIC 50 e la capacità dei ceppi batterici di formare biofilm (OD570nm dopo
colorazione con CV). r: coefficiente di correlazione di Pearson; non significativo. A
B
74
4.4 Cinetica dell’attività antibiofilm di hep20 verso ceppi di S. epidermidis PIA-positivi e PIA-negativi.
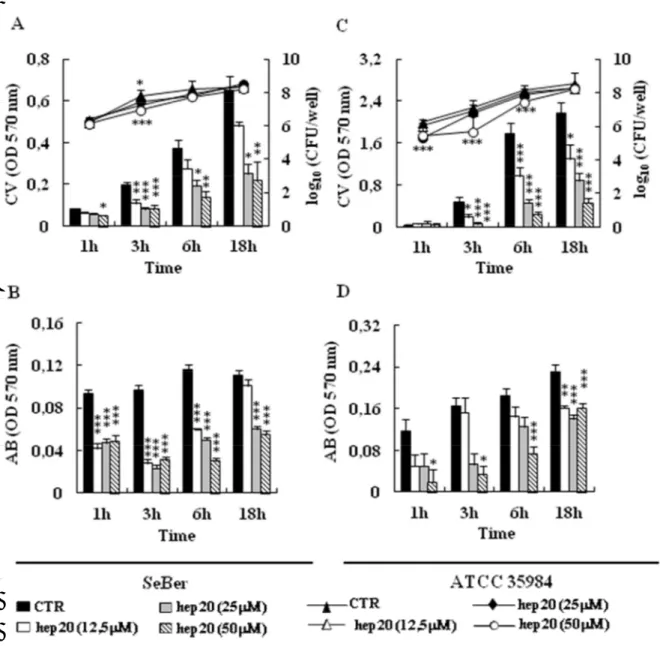
La cinetica dell’effetto antibiofilm di hep20 è stata valutata verso due ceppi di S. epidermidis arbitrariamente scelti, SeBER e ATCC 35984, rappresentativi rispettivamente del ceppi PIA-negativi e PIA-positivi. A tale scopo, si è proceduto a valutare, a vari tempi dall’inoculo (1h, 3h, 6h, 18h), la quantità di biomassa totale, il metabolismo cellulare ed il numero di batteri vitali associati al biofilm. Per entrambi i ceppi batterici si osservava una riduzione statisticamente significativa della biomassa del biofilm già dopo 3-6 h d’incubazione con il peptide (Figura 23 A, C); tale riduzione si manteneva, alle concentrazioni più alte di peptide, fino alle 18h. Parallelamente alla riduzione della biomassa si osservava una netta riduzione anche dell’attività metabolica cellulare valutata mediante saggio con alamar blue (Figura 23 B, D). L’analisi delle cellule vitali associate al biofilm evidenziava, inoltre, una riduzione statisticamente significativa anche nel numero delle CFU, che però, per entrambi i ceppi analizzati, era limitata ai tempi precoci (3-6h) di incubazione (Figura 23 A, C) ed alle concentrazioni più elevate del peptide (25 e/o 50 ).
75
Figura 23. Effetto di hep20 sulla biomassa totale del biofilm, sull’attività metabolica cellulare e sul numero delle cellule vitali associate al biofilm dei ceppi di S. epidermidis SeBER (A, B) e ATCC 35984 (C, D) a diversi tempi d’incubazione.
Gli istogrammi rappresentano i valori di densità ottica dopo colorazione con cristal violetto (A, B) o con alamar blu (B, D), mentre le linee continue (A,C) indicano il numero delle CFU. Analisi statistica: test ANOVA seguito da Tukey-Kramer. p<0.05:*; p<0.01:**; p<0.001:***.
76
4.5 Valutazione dell’attività antibiofilm di hep20 mediante microscopia confocale Gli effetti di hep20 sul biofilm di un ceppo di S. epidermidis PIA-positivo (ATCC 35984) e di un ceppo PIA-negativo (SeBER) sono stati valutati mediante microscopia confocale. A tale scopo, dopo 18 h di trattamento con il peptide, i biofilm di entrambi i ceppi sono stati sottoposti a marcatura con i traccianti fluorescenti SYPRO Ruby (fluorescenza rossa) e Oregon Green 488, coniugato all’agglutinina di germe di grano (Wheat Germ Agglutinin, WGA) (fluorescenza verde) (Figura 24). Il primo di tali traccianti colora la componente proteica della matrice extracellulare, mentre il secondo si lega all’N-acetilglucosammina e rileva, pertanto, la presenza di PIA nella matrice extracellulare del biofilm, anche se può riconoscere anche altri componenti della stessa matrice quale il peptidoglicano o gli acidi teicoici. In accordo con i dati ottenuti dalla caratterizzazione fenotipica e genotipica dei ceppi batterici, il biofilm del ceppo ATCC 35984 mostrava una prevalente ed intensa colorazione verde dovuta al legame di WGA all’N-acetilglucosamina, il costituente prevalentemente del PIA (Figura 24 A, C). Il trattamento del biofilm del ceppo ATCC 35984 con hep20 causava una chiara riduzione dell’intensità di fluorescenza verde in maniera dose-dipendente ed una distruzione evidente dell’architettura del biofilm con solo un modesto effetto sul contenuto proteico del biofilm (Figura 24 A, B, C, D). Al contrario, il biofilm del ceppo SeBER veniva colorato in maniera evidente con il colorante delle proteine SYPRO Ruby (Figura 24 B, D) indicando che la matrice di tale ceppo era costituita prevalentemente da proteine. L’incubazione con hep20 causava, in questo caso, una marcata riduzione dell’intensità della fluorescenza rossa senza effetti evidenti sul contenuto di N-acetilglucosamina (Figura 24 A, B, C, D). In accordo con i dati ottenuti valutando il numero delle cellule vitali associate al biofilm (CFU) a 18h, quando i biofilm di entrambi i ceppi batterici venivano colorati con il dead/live BacLight Bacterial viability kit, nessun effetto evidente di hep20 è stato osservato sulla vitalità delle cellule associate al biofilm (Figura 25).
77
Figura 24. Microscopia confocale dei biofilm di S. epidermidis ATCC e SeBER trattati o meno con hep 20 per 18h. (A) colorazione dei biofilm con il coniugato Oregon green-WGA che si lega alla componente polisaccaridica; (B) colorazione dei biofilm con SYPRO Ruby che si lega alla componente proteica della matrice; (C) e (D) intensità media di fluorescenza dopo colorazione con Oregon green-WGA e con SYPRO Ruby, rispettivamente.
0 M 12,5M 25M 50M
C
78
Figura 25. Immagini al microscopio confocale del biobilm di S. epidermidis ATCC 35984 colorato con il dead/live BacLight Bacterial viability kit, dopo 18 h di trattamento con hep20. L’intensa colorazione verde indica le cellule vitali associate al biofilm
0 M
50 M 25 M 12,5 M
79
4.6 Durata dell’effetto inibitorio di hep20 su biofilm di S. epidermidis
In vista dell’applicazione terapeutica di peptidi antimicrobici, in particolare nel contesto dello sviluppo di sistemi di rilascio controllato, può essere importante determinare l’effetto post-antimicrobico del peptide (Post antimicrobial effect, PAE) definito come la durata dell’effetto inibitorio dopo la rimozione del peptide. A questo scopo, le cellule di un ceppo rappresentativo di S. epidermidis (SeBER) venivano fatte aderire a piastre di polistirene per 3h in presenza di hep20 (25 e 50 ); il peptide e i batteri non adesi venivano quindi rimossi e l’attività residua di hep20 veniva valutata come biomassa totale, attività metabolica e numero di CFU a 3, 6, e 18 h dopo il trattamento (Figura 26). L’attività inibitoria di hep20 sulla biomassa del biofilm si manteneva fino a 18 h dalla rimozione del peptide (Figura 26 A), mentre l’inibizione dell’attività metabolica durava fino a 6 h dopo il trattamento (Figura 26 B). Nessun effetto evidente veniva osservato sul numero di CFU, a nessuno dei tempi analizzati (Figura 26 A).
80 * * * * * * * * * * * * * * * * * * * * * * * * * * * * A B
Figura 26. Durata dell’effetto inibitorio di hep20 sul biofilm di S. epidermidis SeBER dopo rimozione del peptide valutata come: biomassa totale (istogrammi pannello A), metabolismo cellulare (istogrammi, pannello B) e numero di CFU associate al biofilm (linee continue pannello A). Analisi statistica: test ANOVA seguito dal test Tukey-Kramer; p<0,05:*; p<0,001***.
81
4.7 Effetto combinato del pre-trattamento con hep20 seguito dal trattamento con vancomicina sulla sopravvivenza dei batteri associati al biofilm di S. epidermidis L’effetto del pre-trattamento con hep20 sull’attività battericida della vancomicina verso biofilm di un ceppo PIA-negativo (SeBER) ed uno PIA-positivo (ATCC 35984) veniva valutato dopo l’esposizione sequenziale del biofilm ad hep20 (50 M) per 18 h e successivamente a 24 h con concentrazioni di vancomicina (32 g/ml) sub-battericide per 24 h. Quando confrontata all’effetto di hep20 o vancomicina usate singolarmente, veniva osservata una riduzione statisticamente significativa nel numero di cellule vitali, per entrambi i ceppi batterici (Figura 27). Tale riduzione nel numero di CFU era particolarmente evidente per il ceppo SeBER per il quale l’effetto combinatorio di hep20 e vancomicina causava una riduzione nel numero di cellule vitali di almeno 2 logaritmi rispetto ai controlli rappresentati da batteri trattati con sola hep20 o batteri trattati con sola vancomicina (P<0.001, ANOVA, seguita dal test Tukey Kramer).
Figura 27. Aumento dell’attività battericida della vancomicina (32 g/ml) dopo pre-trattamento dei biofilm dei ceppi di S. epidermidis SeBER e ATCC 35984 con hep20 (50 ) per 18 h. Analisi statistica: test ANOVA seguito dal test Tukey-Kramer, p<0.05:*; p<0.01:**; p<0.001:***.
82 5. DISCUSSIONE
Il problema dell’esistenza di microrganismi patogeni antibiotico-resistenti risulta particolarmente aggravato dalla loro capacità di formare biofilm. I biofilm sono complesse comunità microbiche aderenti a superfici solide con proprietà che spesso differiscono largamente da quelle delle corrispondenti cellule in forma planctonica. Nel corso degli ultimi anni, è risultato evidente che la formazione di biofilm, contribuisce in modo significativo alla resistenza batterica verso i chemioantibiotici ed è coinvolta in una grande proporzione (fino all’80%) di infezioni ospedaliere nei paesi industrializzati, molte delle quali sono correlate all’impiego di dispositivi medici. Nonostante la rilevanza clinica delle infezioni associate a biofilm, ad oggi non esiste ancora una terapia clinicamente efficace per la prevenzione e/o l’eradicazione di tali complesse comunità microbiche. Uno degli ostacoli maggiori all’eradicazione del biofilm è rappresentato dalla matrice extracellulare, una complessa struttura formata da un insieme di molecole di diversa natura chimica che, a seconda della specie batterica e alle condizioni ambientali, può includere esopolisaccaridi, proteine, acidi teicoici, lipidi e DNA extracellulare (Flemming and Wingender 2010). I componenti della matrice possono ostacolare l’attività dei farmaci antimicrobici con vari meccanismi che includono: il sequestro o la repulsione elettrostatica del farmaco, la sua inattivazione da parte di proteine ad attività enzimatica, il rallentamento della sua diffusione attraverso gli strati della matrice, l’impedimento della sua interazione con la superficie batterica. Tra i microrganismi produttori di biofilm, una delle specie più frequentemente coinvolta nella contaminazione di dispositivi medici è S. epidermidis e la capacità di formare biofilm è considerata il principale fattore di virulenza di tale microrganismo. Dato che la matrice extracellulare costituisce una delle componenti principali del biofilm degli stafilococchi, recentemente sono state proposte molteplici strategie che hanno come target tale componente e che includono l’inibizione della sua sintesi o la sua degradazione enzimatica (Jabbouri and Sadovskaya, 2010).
Scopo del presente lavoro di dottorato è stato quello di valutare l’attività antibiofilm del peptide di derivazione umana hep20
,
verso isolati clinici di S. epidermidis multi-farmaco-resistenti caratterizzati in dettaglio relativamente alla loro capacità di produrre biofilm e alla natura della matrice extracellulare prodotta.83
Dati ottenuti precedentemente nel nostro laboratorio (Maisetta et al., 2010; Tavanti et al 2011) hanno chiaramente dimostrato che l’attività antimicrobica di hep20 verso cellule planktoniche viene marcatamente potenziata a valori di pH decisamente acidi (pH 5.0) (Maisetta et al., 2010; Tavanti et al 2011). E’ noto che nello spessore di un biofilm possono originarsi, con larga probabilità, nicchie ambientali caratterizzare da bassi valori di pH, causate dall’accumulo locale di metaboliti acidi prodotti dalla fermentazione dei batteri. S. epidermidis, inoltre, rappresenta un tipico colonizzatore della cute dell’uomo, caratterizzata da valori di pH debolmente acido. Tale considerazioni ci hanno fornito delle basi razionali per saggiare hep20 verso biofilm di tale specie batterica.
In una prima fase dello studio, sono stati effettuati esperimenti preliminari per valutare se, analogamente a quanto osservato con cellule in fase planktonica, anche l’eventuale attività antibiofilm di hep20 fosse influenzata dal pH del mezzo di coltura. I risultati ottenuti con un ceppo rappresentativo, hanno dimostrato che l’attività migliore si aveva quando il biofilm veniva allestito ad un pH debolmente acido (6.6) ottimale anche per la crescita di S. epidermidis, mentre a valori di pH decisamente acidi (pH 5.0) il peptide perdeva parte della sua attività antibiofilm. Tale osservazione suggerisce che le condizioni ottimali per l’attività di un peptide antimicrobico verso cellule planktoniche non necessariamente coincidono con quelle che ottimizzano la sua attività verso le stesse cellule in forma di biofilm e che, quindi, i meccanismi dell’azione antimicrobica possono differire a seconda che le cellule siano in sospensione od in forma sessile. Sulla base di questi esperimenti preliminari tutti gli esperimenti successivi sono stati effettuati a pH 6.6.
Per lo studio sono stati selezionati 7 ceppi, di cui 6 isolati clinici ed un ceppo di riferimento spesso usato in studi che abbiano come oggetto biofilm di S. epidermidis. I risultati ottenuti dal saggio su CRA (Congo Red Agar), dal saggio di suscettibilità del biofilm alle proteasi e dall’analisi genetica dell’operone ica (i cui prodotti genici sono responsabili della sintesi del PIA), hanno indicato che 4 di tali ceppi (ATCC 35984, Se30, SeAle, SeNOC) possedevano una matrice extracellulare di natura prevalentemente polisaccaridica, mentre 3 ceppi (SeBER, SeVIC, SeNEG) possedevano una matrice extracellulare di natura prevalentemente proteica. Tutti e sette i ceppi hanno dimostrato la capacità di produrre abbondante biofilm in vitro. Tale osservazione è a sostegno di quanto sta recentemente emergendo in letteratura (Kogan et al., 2006) che,
84
diversamente da quanto suggerito in precedenza, il PIA non è assolutamente indispensabile alla formazione di biofilm da parte di S. epidermidis e che nel processo di aggregazione possono essere coinvolte anche altre molecole, tra cui le proteine. Per alcune proteine è di fatto già stato dimostrato un ruolo importante nella formazione di biofilm di stafilococchi come per la Bap (Biofilm Associated Protein) (Tormo et al., 2005a) o la Aap (Accumulation Associated Protein) (Hussain et al. 1997).
L’analisi della capacità di hep20 di inibire la formazione dei biofilm da parte dei ceppi caratterizzati ha rivelato che il peptide è, in generale dotato di una discreta capacità di provocare una riduzione dose-dipendente della biomassa totale del biofilm di S. epidermidis, valutata dopo colorazione con cristal violetto. Tale attività si aveva, nella maggior parte dei casi, a concentrazioni ben al disotto dei valori di MIC valutati su cellule planctoniche, suggerendo che l’effetto inibitorio non fosse legato ad una attività antimicrobica diretta, ma verosimilmente ad un meccanismo biofilm-specifico. Sebbene si sia osservato un certo grado di variabilità nella suscettibilità dei ceppi a hep20 non si avevano evidenti differenze tra ceppi PIA-positivi e PIA-negativi. Questa osservazione potrebbe essere rilevante per un futuro uso di hep20 come nuova molecola antibiofilm poichè, come già sottolineato in precedenza ceppi PIA-positivi e PIA-negativi possono essere entrambi coinvolti in infezioni nosocomiali associate a biofilm. L’inibizione della biomassa valutata come decremento della colorazione di CV, correlava con il decremento dell’attività metabolica del biofilm saggiata mediante colorazione con Alamar blue. Al contrario, il numero dei batteri vitali associati al biofilm veniva solo parzialmente ridotto dalla presenza del peptide, in accordo con il fatto che l’effetto inibitorio era ottenuto a concentrazioni di hep20 sub-MIC. Tali risultati suggeriscono, innanzi tutto, delle considerazioni metodologiche. La colorazione con CV viene considerata un indice della biomassa totale del biofilm in quanto mette complessivamente in evidenza la componente cellulere adesa alla superficie e la matrice extracellulare (insieme di polisaccaridi, proteine, DNA etc), non discriminando tra cellule vive e morte. La vitalità cellulare è, ovviamente, un aspetto molto importante quando si valuta l’attività di un composto ad attività antimicrobica verso biofilm microbici ed il saggio con AB, che misura l’attività metabolica delle cellule associate al biofilm, è largamente utilizzato in letteratura come indice indiretto della vitalità cellulare. Bisogna considerare, tuttavia, che nel caso dei biofilm la riduzione dell’attività metabolica misurata con AB potrebbe non necessariamente essere dovuta a
85
morte cellulare poiché è noto che nel biofilm molte cellule possono entrare in uno stato di dormienza metabolica come risposta adattativa alle condizioni di stress. I risultati ottenuti nella presente tesi di valutazione dell’attività antibiofilm di hep20 saggiando in parallelo, la riduzione della biomassa totale con cristal violetto, la riduzione dell’attività metabolica con AB e la riduzione del numero di batteri vitali mediante semina su piastra sottolineano che l’attività metabolica e la vitalità cellulare non necessariamente sono due parametri correlati e che, per una completa caratterizzazione in vitro di un agente antibiofilm, essi devono essere determinati entrambi. Come sottolineato anche da altri autori (Welch et al., 2012), a causa della complessità strutturale dei biofilm microbici e dei numerosi potenziali target dell’attività antibiofilm dei peptidi antimicrobici è sempre più sentita l’esigenza di sviluppare procedure standardizzate e condivise tra diversi laboratori per i saggi antibiofilm che possano consentire di studiare accuratamente e confrontare gli effetti delle molecole in studio, analogamente a quanto fatto da tempo per i saggi di suscettibilità in fase liquida.
La capacità di hep20 di ridurre marcatamente la biomassa del biofilm e la sua attività metabolica con solo trascurabili effetti sul numero di CFU, suggerisce l’ipotesi che il peptide possa interferire con la produzione della matrice extracellulare da parte delle cellule associate al biofilm. Un analogo meccanismo antibiofilm è stato suggerito per altri peptidi antimicrobici ed anche per antibiotici convenzionali. Ad esempio, Totè e collaboratori hanno valutato l’attività antibiofilm di antibiotici convenzionali valutandone separatamente l’effetto sulla matrice extracellulare e sulla vitalità delle cellule biofilm-associate di S. aureus e P. aeruginosa (Totè et al., 2009) . Sebbene, ad elevate concentrazioni, la maggior parte degli antibiotici saggiati aveva effetto antimicrobico sulle cellule del biofilm ma non sulla componente di matrice, ve ne erano anche alcuni che agivano con il meccanismo inverso riducendo cioè la matrice con trascurabili effetti sulla vitalità cellulare. Tra gli antibiotici saggiati solo uno, la rifampicina, agiva causando contemporaneamente la riduzione della matrice extracellulare e delle CFU dei biofilm di entrambi i ceppi batterici testati.
L’ipotesi da noi formulata che hep20 possa agire sul biofilm di S. epidermidis interferendo con l’accumulo della matrice extracellulare, con solo un minimo effetto sul numero delle cellule vitali è stata pienamente supportata dagli esperimenti di microscopia confocale. Le tecniche di microscopia (ad esempio Scanning Electron Microscopy, Confocal Laser Scanning Microscopy ed altre) sono sempre state
86
ampiamente utilizzate per studiare gli effetti di molecole con potenziale antimicrobico sulle cellule batteriche. Tali tecniche, tuttavia, hanno assunto un’importanza fondamentale nello studio degli agenti antibiofilm. Le informazioni ottenute dalle immagini microscopiche, infatti, se combinate con tecniche di immuno-marcatura o di marcatura con sostanze fluorescenti specifiche per le varie componenti del biofilm, possono dare delle indicazioni cruciali nel chiarire il meccanismo dell'azione antibiofilm ed i possibili traget molecolari delle molecole attive. I risultati ottenuti in microscopia confocale, marcando la componente polisaccaridica e quella proteica della matrice extracellulare di due ceppi rappresentativi di S. epidermidis, hanno chiaramente confermato che hep20 provocava una netta alterazione, dose-dipendente dell’architettura del biofilm ed una riduzione della matrice sia del ceppo PIA-positivo che di quello PIA-negativo, indipendentemente cioè dalla natura della matrice stessa. I dati hanno anche confermato che a 18h le cellule associate al biofilm trattato con hep20 sono in gran parte vitali, nonostante la marcata riduzione dei componenti di matrice per entrambi i ceppi saggiati.
Il meccanismo di inibizione della matrice extracellulare da parte di hep20 è al momento completamente sconosciuto e sarà oggetto di studi futuri. L’osservazione che l’effetto inibitorio vada di pari passo alla forte riduzione dell’attività metabolica suggerisce che l’inibizione della produzione della matrice extracellulare possa essere il risultato di un’inibizione generale, peptide-mediata, di processi attivi quali la biosintesi di proteine o di polisaccaridi. Alernativamente, in virtù della sua natura cationica, il peptide potrebbe interporsi tra le cellule batteriche caricate negativamente ed ostacolare la deposizione di matrice extracellulare. Ciò potrebbe, a sua volta, riflettersi in una ridotta aggregazione cellulare come supportato dalla riduzione del numero delle CFU osservata nelle fasi più precoci (3 e 6 ore).
Tra le strategie mirate a colpire la matrice extracellulare oggi allo studio, c’è il trattamento dei biofilm degli stafilococchi con enzimi ad attività idrolitica verso il PIA (per esempio la dispersina B) o verso le proteine di matrice (per esempio la proteinasi K o la tripsina) (Chaignon et al 2007; Jabbouri and Sadovskaya, 2010). Per l’eterogeneità della matrice tra isolati clinici Chaignon e collaboratori suggeriscono che un pre-trattamento con dispersina B seguito da quello con proteinasi K possa essere necessario per facilitare l’eradicazione del biofilm di una grande varietà di ceppi di stafilococco. L’uso di AMPs che in modo simile a hep20 abbiano la capacità di inibire la matrice sia
87
dei ceppi PIA-positivi che PIA-negativi di S. epidermidis potrebbe eliminare la necessità di due o più trattamenti consecutivi e, nello stesso tempo potrebbe evitare il problema dei possibili effetti collaterali del trattamento con proteinasi K sui tessuti circostanti la protesi infetta del paziente trattato.
Il primo parametro per misurare l’attività antimicrobica di un antibiotico è quello di determinare la minima concentrazione inibente (MIC), definita come la concentrazione più bassa di un composto in grado di inibire completamente la crescita di un microrganismo. Essa è un ottimo indicatore della potenza antimicrobica diretta di un composto ma dà poche indicazioni sulla sua attività nel corso del tempo dopo la somministrazione. La persistenza degli effetti antimicrobici di una molecola nel tempo viene valutata come effetto post-antibiotico (Post-Antibiotic Effect, PAE) (Frimodt-Moller). Tale parametro potrebbe essere particolarmente importante nel pianificare terapie combinate o l’impiego di nuovi sistemi di delivery di farmaci a rilascio controllato. Quando si è proceduto a valutare la persistenza degli effetti inibitori di hep20 su biofilm di S. epidermidis è emerso un dato interessante. Appena 3 ore di esposizione al peptide erano sufficienti per far durare l’inibizione della matrice fino a 18 ore dalla rimozione del peptide stesso. Tale osservazione indica che gli effetti inibitori del peptide sono pittosto stabili nel tempo e suggerisce possibili cinetiche di somministrazione del peptide in terapie combinate con antibiotici che agiscano riducendo il numero delle CFU.
Al fine di valutare l’effettivo potenziale applicativo di hep20 in terapie combinate è stata valutata la capacità del peptide di potenziare l’attività antibatterica della vancomicina verso biofilm di S. epidermidis PIA-positivi e PIA-negativi. I risultati ottenuti hanno permesso di osservare che il pre-trattamento del biofilm con hep20 favorisce l’attività antimicrobica della vancomicina abbassandone notevolmente la concentrazione attiva e causando una riduzione statisticamente significativa nel numero delle CFU rispetto ai biofilm non pre-trattati con hep20. L’aumento dell’attività antibiofilm della vancomicina dopo pre-trattamento con hep20 supporta l’ipotesi che l’inibizione della produzione della matrice extracellulare da parte del peptide possa favorire la diffusione dell’antibiotico attraverso gli strati del biofilm e, quindi, la sua interazione con le cellule batteriche fino al raggiungimento dei propri bersagli molecolari.
88
In conclusione, lo sviluppo di nuovi approcci terapeutici e nuove strategie mirate all’inibizione e/o alla distruzione dei biofilm microbici in ambito medico rappresenta ad oggi un importante campo di ricerca. Negli anni futuri i biofilm microbici rappresenteranno una seria sfida per diverse figure professionali (medici, microbiologi, chimici) e solo un approccio multidisciplinare ed una profonda conoscenza delle proprietà fisiologiche e strutturali celle cellule batteriche in forma di biofilm potrà permettere la messa a punto di strategie idonee per la loro eradicazione.