Capitolo 2
Biomeccanica dell’Octopus vulgaris
2.1 Introduzione
Dopo aver introdotto il concetto di biomimetica e le ragioni per cui molti studiosi guardano con crescente attenzione il polpo, è il momento di dedicare ampi spazi a questo curioso animale.
Il principale ostacolo ad uno studio sistematico e approfondito sul polpo è rappresentato in primis dall’ambiente in cui questi vive. Esistono numerose tecnologie e metodologie pensate e sviluppate per lo studio di animali terrestri e che, in linea di massima, potrebbero essere utilizzate anche per lo studio di animali acquatici. Andando su un piano più prettamente pratico, però, si incontrano notevoli difficoltà oggettive. Prendiamo, ad esempio, il caso dello studio sulla cinematica del tentacolo: la tecnica più semplice e per questo più utilizzata prevede l’uso di marker (attivi o passivi) che possono essere seguiti otticamente per la ricostruzione di traiettorie 2D o 3D. I problemi principali sono rappresentati dalla necessità di impermeabilizzazione degli strumenti, dal fissaggio dei marker e dalla rifrazione dell’acqua che può introdurre errori ottici non trascurabili. In aggiunta si deve osservare che una delle caratteristiche del polpo, che tanto abbiamo ammirato nel primo Capitolo, in questo caso gioca a nostro sfavore: la grande destrezza dei suoi tentacoli farebbe si che qualsiasi tipo di marker artificiale fissato alla cute avrebbe davvero vita breve... Ovviamente sono disponibili molte altre tipologie d’indagine, tra cui anche quelli che prevedono la stimolazione artificiale, ma queste, in generale, prevedono l’isolamento
del soggetto dal suo ambiente e quindi implicano tutta una serie di limitazioni all’attendibilità dei risultati. Per questa ragione i studi rivolti alla comprensione, ad esempio, della neurofisiologia sono ad un punto molto avanzato: i metodi utilizzabili sono gli stessi applicati al caso umano/terrestre. Al contrario, sui tentacoli (che non hanno un equivalente umano) non è ancora possibile uno studio sistematico e non se ne ha quindi una conoscenza profonda.
Nel presente Capitolo, dopo una breve, ma doverosa, descrizione anatomica, si passeranno al vaglio i risultati di diversi studi atti a caratterizzare la biomeccanica dei movimenti del tentacolo. Infine verrà posto l’accento sulle principali lacune esistenti in letteratura e verrà introdotto il filo conduttore dei successivi capitoli.
2.2 Anatomia del tentacolo
I polpi possiedono quattro paia di tentacoli che si proiettano dalla testa e che circondano la bocca. Essi sono affusolati ed attrezzati con una doppia fila di ventose (Octopus vulgaris) e sono usati per la locomozione, la cattura di prede e funzioni sensoriali [20]. Al loro interno, i tentacoli presentano una struttura densamente compatta e che si mantiene invariata per tutta la lunghezza del tentacolo (scalando in dimensione).
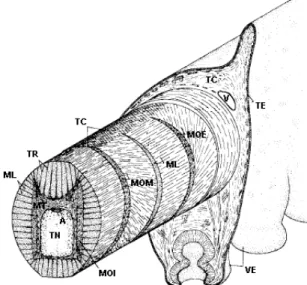
Fig. 2.1 Tentacolo di polpo: TN, tessuto nervoso; A, arteria; MT, muscoli trasversali; ML, muscoli longitudinali; MOI, muscoli obliqui interni; TR, trabeculae; TC tessuto connettivo;
MOM, muscoli obliqui mediani; MOE, muscoli obliqui esterni; V,vena; VE, ventose; TC, tessuto connettivo.
In Fig. 2.1 viene mostrato un diagramma schematico di un tentacolo di polpo [21]. Il tessuto nervoso centrale e la arteria principale ad esso associato sono disposti lungo l’asse centrale del tentacolo e sono circondati dai fasci muscolari trasversali. Queste masse sono molto spesse nella parte dorsale del tentacolo (diametralmente opposta alle ventose), intermedie ai lati e sottili in basso. Le fibre muscolari facenti parte della massa trasversale, sono estremamente fitte e orientate in maniera perpendicolare alla lunghezza del tentacolo. I fasci muscolari trasversali si estendono verso l’esterno inserendosi tra le fibre disposte longitudinalmente. Queste “trabeculae” sono disposte radialmente su tutta la sezione del tentacolo e si inseriscono in uno strato di tessuto connettivo fibroso che racchiude la maggior parte della muscolatura intrinseca ed è specialmente spesso nella parte che da alle ventose e quella diametralmente opposta (Fig. 2.2) [22]. Le trabeculae dei muscoli trasversali estendendosi lateralmente passano attraverso tre strati di muscoli obliqui (che si vedranno nel prosieguo) diramandosi [23] [24] [25] prima di inserirsi in un sottile strato laterale di tessuto connettivo.
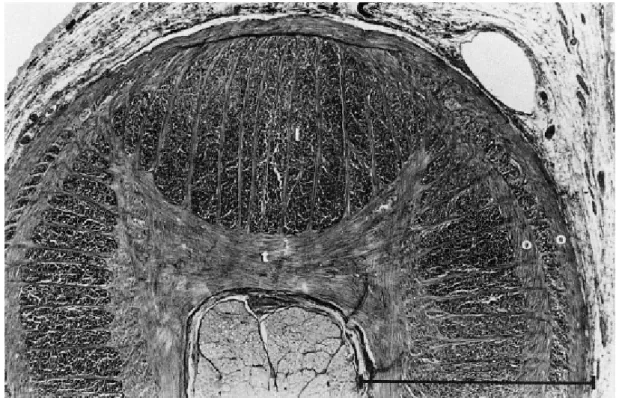
Fig. 2.2 Dettaglio della sezione di tentacolo con evidenziate le principali strutture muscolari: t, trasversali; l longitudinali; o, obliqui.
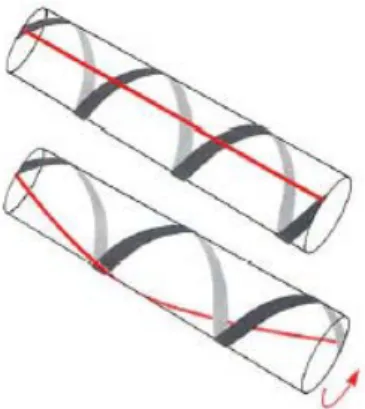
Il tentacolo di polpo è provvisto di tre set di strati di muscoli obliqui: interni; mediani; esterni. Questi fasci si sviluppano in lunghezza a forma di elica andando a circondare l’intero tentacolo. Il verso di avvolgimento di ogni elica è opposto a quello delle eliche che si trovano in posizione opposta. Per esempio, gli obliqui esterni ad un lato sono disposti in senso destrorso, quelli alla parte opposta si sviluppano in maniera sinistrorsa. In più le tre eliche dello stesso lato non hanno tutte la stessa disposizione: l’esterna ha la stessa dell’interna, ma l’opposta della mediana.
L’angolo che formano le fibre oblique rispetto all’asse centrale in un tentacolo rilassato è di ~60°. Un sottile strato di fibre muscolari disposte circonferenzialmente copre la superficie superiore della muscolatura intrinseca.
2.3 Biomeccanica dei movimenti base
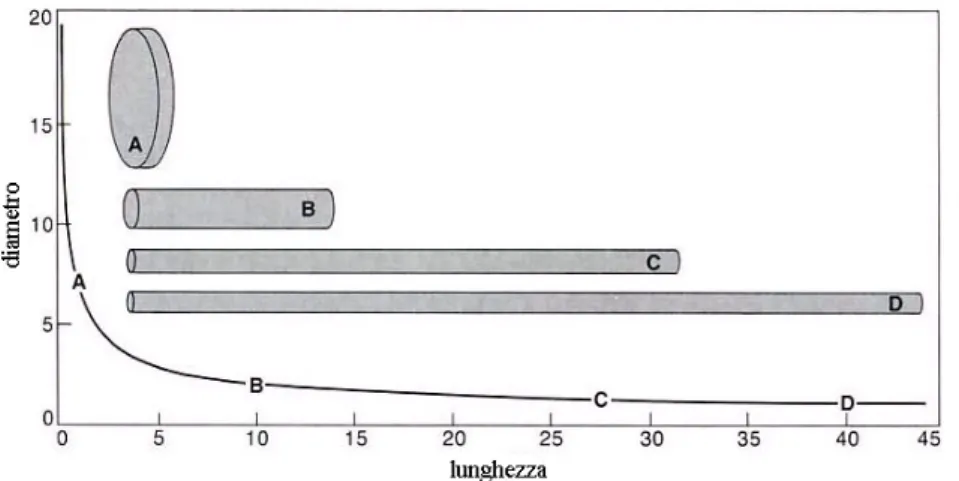
La struttura anatomica appena introdotta è di fondamentale importanza per capire come il polpo riesca a compiere tutta una serie di movimenti anche in mancanza di un supporto rigido. Come vedremo, la disposizione dei fasci muscolari è finalizzata all’ottimizzazione dei movimenti possibili, ma l’elemento fondamentale, che permette ad una struttura così fatta di emulare, e per certi versi superare, le funzioni usualmente prodotte dai vertebrati, risiede nell’incomprimibilità di tali muscoli. Questo tipo di muscoli (denominati Hydrostat [22] [26]) formano una struttura densamente riempita e mantengono un volume costante durante tutti i movimenti. Questo implica che la contrazione (e quindi l’accorciamento) di un fascio muscolare è sempre accompagnato da una sua dilatazione nelle altre due dimensioni (Fig. 2.3) e viceversa.
Fig. 2.3 Relazione tra lunghezza e diametro di una fibra muscolare cilindrica idrostatica La natura idrostatica dei muscoli permette un funzionamento simil-agonista/antagonista dei fasci muscolari che assumono questo specifico orientamento. Questo comportamento porta alla produzione di una varietà di movimenti base che includono: • Allungamento; • Accorciamento; • Stiffening; • Curvatura; • Torsione;
2.3.1 Allungamento
L’allungamento del tentacolo richiede la contrazione di quei fasci muscolari che diminuiscono il diametro del tentacolo; in particolare è decisivo il contributo dei muscoli trasversali. La contrazione di questi, infatti, esercita una pressione radiale sui muscoli longitudinali (che sono mantenuti rilassati) che quindi per le proprietà idrostatiche appena descritte subiscono un allungamento che si traduce in una strizione ed un incremento di lunghezza dell’intero tentacolo.
Da un punto di vista robotico, questo effetto amplificante è uno degli aspetti più interessanti da studiare ed è il punto cardine su cui si basa la progettazione del tentacolo robotico affrontata nel Capitolo 3.
2.3.2 Accorciamento
L’accorciamento avviene come risultato di una contrazione della muscolatura longitudinale e fornisce il mezzo attraverso cui i muscoli responsabili dell’allungamento possono essere antagonizzati. L’accorciamento segue gli stessi principi idrostatici visti per l’allungamento, anche se in questo caso l’effetto prodotto è di entità minore.
Vedremo infatti come in Natura l’accorciamento non è sfruttato quanto l’allungamento, poiché per avvicinare oggetti in prossimità della testa, il polpo usa altri tipi di strategie.
2.3.3 Stiffening
Un fenomeno che invece sfrutta sia i meccanismi visti nell’accorciamento che nell’allungamento del tentacolo è la variazione attiva di stiffness. Questo non è un vero è proprio movimento, ma è molto utilizzato dal polpo, in quanto gli permette di irrigidire volontariamente e selettivamente parti o l’intero tentacolo.
C’è da notare che in questo caso i due fasci muscolari coinvolti (muscoli longitudinali e trasversali) non hanno una funzione antagonista come nei due casi appena visti, ma agiscono sinergicamente a raggiungere lo scopo.
2.3.4 Curvatura
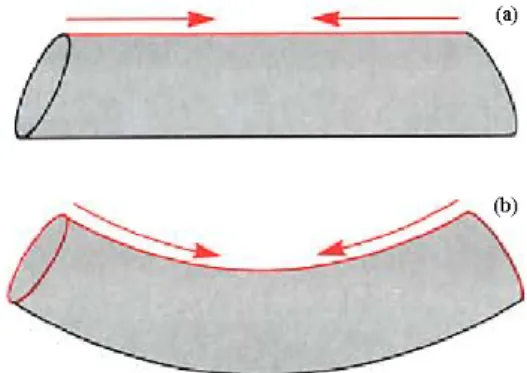
Per effettuare una curvatura il tentacolo viene sottoposto alla contrazione dei muscoli longitudinali lungo il lato in cui si deve formare l’interno della piega. Questo però non è sufficiente a garantire la formazione di una curva, in quanto il risultato che si avrebbe con la sola contrazione dei muscoli longitudinali sarebbe l’accorciamento di un lato e una conseguente variazione di diametro (Fig. 2.4 (a)). È necessario che qualche struttura impedisca attivamente questa variazione, dando modo di formare la curvatura richiesta in maniera proporzionale all’intensità di contrazione dei muscoli longitudinali. Da quello che si è visto fin’ora si può facilmente evincere come gli unici in grado di compiere questo lavoro resistente siano i muscoli trasversali, che grazie alla loro contrazione possono mantenere costante la sezione del tentacolo (Fig.2.4 (b)).
Fig. 2.4 Principio di formazione della curvatura del tentacolo: contrazione dei longitudinali (a) e dei relativi trasversali (b).
Anche in questo caso i muscoli longitudinali e trasversali vengano attivati contemporaneamente in maniera non antagonista.
Una curvatura richiede quindi il controllo locale sia dei muscoli longitudinali che trasversali, il che implica un sistema neuromuscolare altamente partizionato. Variando la coordinazione di contrazione dei due fasci il punto di curvatura o la direzione possono essere variati. Il sistema quindi risulta estremamente flessibile, molto di più di altri sistemi muscolo-scheletrici.
2.3.5 Torsione
L’ultimo movimento di base implementabile è la torsione attorno all’asse centrale. I muscoli responsabili di questo movimento devono necessariamente essere disposti ad elica tutto intorno al tentacolo. Stiamo parlando ovviamente dei muscoli obliqui, che grazie alla loro struttura sono in grado di far ruotare parti più o meno lunghe di tentacolo a seconda del verso di rotazione dell’elica (destrorsa o sinistrorsa) (Fig. 2.5). Dei tre fasci menzionati nel paragrafo dedicato all’anatomia, i più influenti (da un punto di vista puramente meccanico) sono i fasci obliqui esterni, poiché hanno la possibilità di sviluppare un momento maggiore, poi nell’ordine vengono i mediani e gli interni.
Fig. 2.5 Meccanismo di torsione effettuato ad opera dei muscoli obliqui.
Un discorso molto importante va fatto a proposito dell’inclinazione delle fibre muscolari oblique rispetto all’asse longitudinale. Prendiamo un cilindro di volume costante avvolto da un singolo passo di elica (Fig. 2.6 A). Se tagliamo il cilindro longitudinalmente e lo rendiamo planare (Fig 2.6 B) possiamo individuare l’angolo θ con cui il fascio avvolge i tessuti sottostanti.
Fig. 2.6 Diagrammi funzionali allo studio dell’effetto dell’angolo di inclinazione delle fibre muscolari oblique.
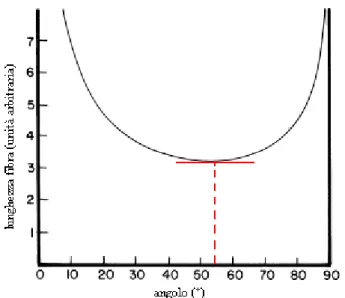
Ricordando che il volume, V,si mantiene costante e calcolando lunghezza, raggio del cilindro di partenza in funzione della lunghezza della diagonale D, si arriva a definire: 3 2 cos sin 4 θ θ π ⋅ = V D
che graficata ci mostra che esiste un punto di minimo esattamente nel punto in cui l’angolo di inclinazione è pari a 54°44’(Fig. 2.7)
Fig. 2.7 Relazione tra l’angolo d’inclinazione delle fibre muscolari oblique e lunghezza del cilindro che avvolge. In rosso il punto di minimo della funzione: 54°44’.
Questo significa che se i muscoli obliqui vengono contratti quando disposti ad un angolo maggiore di 54°44’ oltre ad una torsione si avrà un allungamento; se l’angolo di partenza è minore di 54°44’ allora si avrà torsione ed accorciamento.
Quindi, quando attivati, questi muscoli svolgono anche un’azione coadiuvante sia nell’allungamento (andando a comprimere i muscoli longitudinali insieme ai trasversali) che nel contenere l’azione dei muscoli longitudinali nella variazione di stiffness.
2.4 Biomeccanica dei movimenti complessi
I movimenti di base descritti possono essere usati singolarmente o in combinazione e formare movimenti molto più complessi. Per l’esecuzione di questi ultimi è lecito porsi il quesito di come possa un animale con un così relativamente piccolo sistema nervoso centrale riuscire a far fronte a questa complessità (senza contare che il polpo è in grado di muovere contemporaneamente tutti e 8 i tentacoli...). La risposta è da cercarsi nelle strategie di controllo che esso adotta nell’eseguire tali movimenti [27]. In particolare ve ne sono due particolarmente interessanti:
• STEREOTIPIZZAZIONE DEL MOVIMENTO, che gli permette di ridurre drasticamente il numero di gradi di libertà da controllare;
• DIVISIONE DEI LAVORI, tra sistema nervoso centrale e periferico (quest’ultimo estremamente evoluto e complesso)
2.4.1 Reaching
Il movimento di reaching viene utilizzato per raggiungere ed afferrare gli oggetti che entrano nel campo visivo del polpo [28]. È uno dei movimenti più utilizzati ed è riassumibile in due passi fondamentali (Fig. 2.8):
1. Viene creata una curva in un punto qualunque lungo il tentacolo; la curvatura avviene sempre dorsalmente in modo da presentare il lato ventrale e le ventose nella direzione di estensione del tentacolo.
2. La piega si propaga lungo il tentacolo fino all’apice mentre la parte prossimale, già investita dallo stimolo, rimane rigida ed estesa.
Il polpo non ha strumenti dedicati alla manipolazione (il cui equivalente potrebbero essere le mani umane), per questo esso proietta i tentacoli in avanti ponendo le ventose frontalmente. Le ventose sono lo strumento più efficace che esso possiede per l’ancoraggio e la presa di oggetti ed in queste sono anche presenti gli organi di senso. Dunque non c’è una zona preferenziale adibita alla funzione tattile o di presa, queste possono avvenire su tutta la lunghezza del tentacolo.
Quello che sembra invece essere un elemento molto importante è il punto di curvatura del tentacolo durante il movimento.
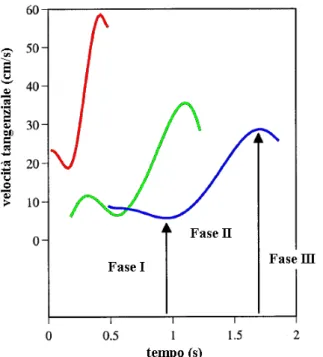
Evidenze hanno dimostrato che il movimento è sempre molto simile, ripetitivo, limitato su un solo piano e che la velocità tangenziale del punto di curvatura assume sempre una forma a campana: da una prima lenta fase di esecuzione, si passa ad una fase più veloce, per poi finire tornando ad una velocità bassa indipendentemente dalla durata e dalla direzione del movimento (Fig. 2.9) [13].
Fig. 2.9 Andamento invariabilmente a campana della velocità tangenziale e distinzione delle tre fasi componenti il movimento.
Considerando il caso di un braccio articolato, per qualsiasi movimento, ogni giunto del manipolatore dovrà assumere una determinata posizione al fine di far giungere l’end effector all’obiettivo. Questo passaggio va sotto il nome di inversione cinematica
e nel caso di manipolatori ridondanti (cioè ad alto numero di gradi di libertà) è un problema di non facile soluzione in virtù della non unicità della soluzione.
Quest’ultima considerazione, insieme alle informazioni sperimentali riportate hanno fatto teorizzare che il polpo potrebbe utilizzare un movimento stereotipato per compiere il reaching, riducendo drasticamente il numero di gradi di libertà da controllare direttamente. In questo modo, infatti, sono sufficienti due variabili per fissare la posizione della base del tentacolo e un’altra per dettare la velocità tangenziale di distensione, limitando il calcolo dell’inversione cinematica a tre sole variabili. Questa teoria è stata confermata da esperimenti di neurofisiologia, in cui tentacoli recisi o denervati sono stati eccitati elettricamente. Tutte le prove hanno portato allo stesso risultato: una distensione del tentacolo sempre nella stessa direzione e sempre alla stessa velocità, a conferma del fatto che queste variabili siano di competenza centrale e che il programma motorio basato su un meccanismo feed-forward sia intrinseco al sistema neuromuscolare stesso. D’altra parte anche i numeri parlano piuttosto chiaro: ~ cellule nervose presenti all’interno del tentacolo contro solo ~30000 fibre nervose che portano informazioni sensoriali e motorie da e verso il sistema nervoso centrale, lasciano intendere che gran parte dei task motori viene deputata ad un controllo periferico.
7
10 5⋅
Inoltre, ad ulteriore testimonianza è stato dimostrato che quando il tentacolo è già esteso e l’obiettivo si trova vicino alla parte distale dell’arto, il polpo non è in grado di raggiungerlo con un movimento traslatorio laterale del tentacolo, ma deve ripercorre necessariamente tutte le fasi di reaching ripartendo dalla porzione prossimale [29]. La spiegazione è implicita nella definizione di movimento stereotipato e in più in questo caso è avvalorata anche da ragioni fisiche: traslare un tentacolo disteso di 30-40 cm in acqua richiede una forza piuttosto alta per vincere la resistenza del mezzo; il movimento che compie il polpo, sebbene apparentemente più macchinoso, presenta una sola piccola porzione di tentacolo alla volta alla forza resistente dell’acqua.
2.4.2 Fetching
Un altro movimento complesso che il polpo è in grado di attuare è quello di fetching, cioè avvicinare alla bocca ciò che è stato raggiunto e afferrato in un qualsiasi punto del tentacolo durante il movimento di reaching.
È stato dimostrato che in questo caso la strategia adottata è completamente diversa e davvero sorprendente: esso sfrutta lo stesso meccanismo dei vertebrati, configurando il suo arto in una struttura rigida e quasi-articolata [30]. Evidentemente la Natura ha decretato che la strategia migliore per spostare un oggetto da un punto ad un altro è la stessa anche in ambiente marino.
Quest’altro tipo di movimento stereotipico è stato studiato ed analizzato attraverso sequenze di immagini in cui è stata ripresa la reazione dei soggetti alla deposizione di cibo su una qualsiasi porzione del tentacolo. Le ventose dell’animale afferrano rapidamente il cibo e vengono generate due curvature: una a livello distale e una a livello mediale, mentre la base del tentacolo funge da terzo “giunto” (prossimale). La posizione del cibo ed i giunti formati definiscono, quindi, una struttura quasi-articolata e (Fig. 2.10 e 2.11) che ruotando principalmente attorno all’asse mediale avvicina la parte distale alla testa.
In particolare è stato riscontrato che i tre giunti dinamici si comportano in maniera molto diversa tra di loro: il giunto prossimale (θ1) compie piccole rotazioni, prima diminuendo di valore, poi aumentando; il giunto mediale (θ2) aumenta per tutta la durata del movimento; infine il giunto distale (θ3) è altamente variabile e gioca un ruolo chiave solo nell’ultima fase del movimento in cui il cibo viene portato alla bocca.
Fig.2.11 Struttura quasi-articolata che il polpo genera per avvicinare alla bocca gli oggetti afferrati.
Anche in questo caso, quindi, viene operata una profonda riduzione di ridondanza al fine di semplificare il calcolo di inversione cinematica. Creando una struttura rigida, ma dinamicamente variabile, il numero di gradi di libertà viene ridotta da infiniti a tre.
L’analisi cinematica del movimento di fetching rivela l’uso di una strategia di controllo particolare che basa la generazione della struttura dinamica quasi-articolata sulla posizione dell’oggetto da avvicinare. Sembra, infatti, che il polpo, a differenza dell’uomo, non abbia la possibilità di scegliere se operare nello spazio dei giunti o nello spazio operativo, poiché avrebbe a disposizione una sola strategia di controllo motorio che prevede l’uso di un unico spazio, chiamato spazio di configurazione dell’arto. Il polpo utilizza questo spazio di lavoro per aggiustare dinamicamente la lunghezza dei segmenti della struttura quasi-articolata in funzione della posizione dell’oggetto sul tentacolo.
Esperimenti eseguiti su tentacoli lesionati o recisi non hanno dato gli stessi risultati avuti nel reaching, il che lascia pensare che dietro questo tipo di movimento ci sia un controllo diretto da parte del sistema nervoso centrale.
Riassumendo: uno stimolo, prodotto dalla presa dell’oggetto, attiva un meccanismo periferico presumibilmente dettato da un centro motorio del sistema nervoso centrale.
Questo meccanismo consiste nella generazione simultanea di due onde di contrazione muscolare: una proveniente dalla base del tentacolo, una dalla posizione dell’oggetto. Queste due onde, viaggiando alla stessa velocità, collideranno in un punto che sarà presumibilmente a metà della distanza che separa l’oggetto dalla base del tentacolo. In quel punto viene formata l’ansa mediale.