1. Introduzione
1.1. Serotonina
La storia della scoperta della serotonina origina a partire dagli anni ’30 con Vittorio Erspamer (Istituto di Anatomia Comparata e Fisiologia, Università di Pavia, Italia). Il Dott. Erspamer stava ricercando sostanze capaci di provocare la contrazione della muscolatura liscia e individuò una sostanza di questo tipo in un estratto di mucosa gastrica di coniglio: la chiamò enteramina (Erspamer e Vialli, 1937). Alla fine degli anni ‘40, il laboratorio di Irvine Page (Clinica di Cleveland, USA), isolò una sostanza vasocostrittrice a partire dal siero e la nominò serotonina (Rapport et al. 1948). Nel maggio del 1949, è stato dimostrato che la 5-idrossitriptamina (5-HT) coincide con la struttura descritta per la serotonina (Rapport, 1949). Intorno al 1952 venne confermato che l’enteramina e la serotonina erano la medesima sostanza. Nonostante la serotonina fosse stata isolata e sintetizzata nel 1951, la sua funzione restava sconosciuta. Nel 1952 Betty Mack Twarog trovò serotonina nel cervello di mammifero e pubblicò il dato nel 1953. Questa fu la scoperta che introdusse la serotonina nel campo delle Neuroscienze (Azmitia, 1999).
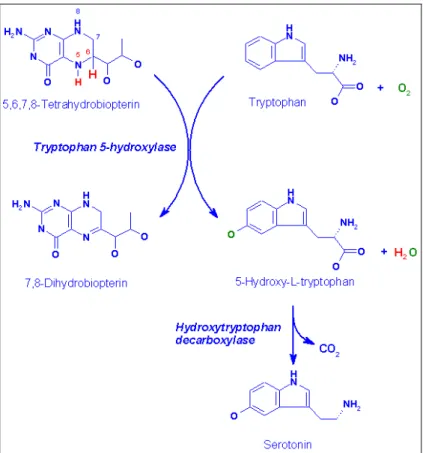
La serotonina (5-HT) è un’amina biogena a basso peso molecolare che appartiene ad un gruppo di composti aromatici (indoli) che presentano un anello a cinque atomi contenente azoto condensato con un anello benzenico. La biosintesi della 5-HT prevede come prima tappa l’idrossilazione del triptofano in posizione 5 a formare il 5-idrossi-triptofano (5-HTP) da parte dell’enzima 5-idrossi-triptofano idrossilasi (Tph). Sono note due isoforme di questo enzima; esse sono codificate da due geni (Tph1 e Tph2) espressi in cellule e tessuti differenti. Gli enzimi Tph1 e
Tph2 subiscono modificazioni post-trasduzionali mediante fosforilazione (Hasegawa and Ichiyama, 1987; Makita, 1990; Ehret, 1991; Walther et
al., 2003; Zhang et al., 2004):
- Tph1: è l’isoforma non-neuronale della Tph ed è localizzata prevalentemente nei tessuti periferici, nei fotorecettori dell’occhio e nella ghiandola pineale dove la sintesi di 5-HT è funzionalmente correlata alla produzione di melatonina (Walther et al., 2003). - Tph2: è l’isoforma neuronale della Tph ed è presente
esclusivamente nei neuroni serotoninergici del sistema nervoso centrale (SNC) (Walther et al., 2003; Patel et al., 2004).
Il 5-HTP viene quindi decarbossilato dalla triptofano-decarbossilasi a 5-HT. La 5-HT viene poi catabolizzata ad acido 5-idrossi-indolacetico (5-HIAA) per azione delle monoaminossidasi a (MAOa)
(Fig. 1.1).
Tutte le azioni che la 5-HT svolge nell’organismo sono mediate da una famiglia di recettori specifici di cui sono note almeno venti forme diverse (Hoyer et al., 1994).
Nel sistema nervoso centrale dei vertebrati, la maggior parte dei neuroni che producono 5-HT sono localizzati in una ristretta zona dell’encefalo, tra mesencefalo e rombencefalo dove sono raggruppati in nuclei noti come nuclei del raphe; esiste inoltre una piccola popolazione di questi neuroni anche a livello del nucleo dorsomediale ipotalamico.
Nei mammiferi, i nuclei del raphe (denominati B1-B9), compaiono a stadi molto precoci dello sviluppo e sono stati suddivisi in due gruppi in base alla posizione che assumono lungo l’asse rostro-caudale del tronco dell’encefalo (Lidov and Molliver, 1982; Wallace and Lauder, 1983); il gruppo rostrale comprende i nuclei B6-B9 (raphe dorsale, caudale,
ventrolaterale), mentre quello caudale comprende i nuclei B1-B5 (raphe magnus, obscurus, pallidus).
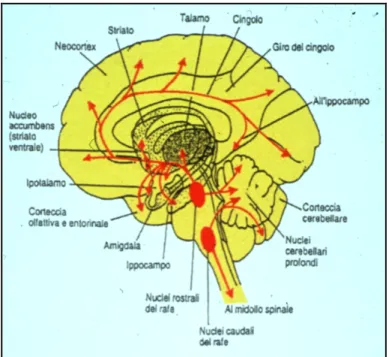
Nonostante il numero totale dei neuroni serotoninergici sia molto ridotto, il sistema che formano si sviluppa in un’ampia innervazione a tutte le aree dell’encefalo e al midollo spinale. I nuclei del raphe rostrale producono delle proiezioni assonali che principalmente attraverso il fascicolo prosencefalico mediale, raggiungono numerose strutture mesencefaliche e prosencefaliche come la corteccia cerebrale, l’ippocampo, il diencefalo, i nuclei della base e il cervelletto. Gran parte di queste fibre però non costituisce un contatto sinaptico diretto con i neuroni bersaglio ma piuttosto, rilascia 5-HT nell’ambiente extracellulare. I nuclei del raphe caudale invece proiettano al midollo spinale dove raggiungono i neuroni pregangliari simpatici, nella colonna intermedio laterale, i motoneuroni somatici e più tardivamente innervano le fibre delle corna dorsali (Fig. 1.2).
La 5-HT è sintetizzata anche nei fotorecettori della retina, nella ghiandola pineale, e a livello periferico nelle cellule enterocromaffini dell’intestino, nei corpi neuroepiteliali dei polmoni e nelle cellule parafollicolari della tiroide (Gaspar et al., 2003).
Fig. 1.2: Rappresentazione schematica delle proiezioni assonali dei neuroni serotoninergici dei
nuclei del raphe in una sezione medio sagittale di un encefalo umano.
Inoltre esistono altri tipi cellulari che grazie ad una transiente o meno espressione di un trasportatore serotoninergico, possono contenere 5-HT senza produrla. Per esempio nei mammiferi, molto precocemente nello sviluppo, è stata riscontrata la presenza di 5-HT a livello del cuore, del mesenchima craniale (Lauder et al., 1988; Shuey et al., 1992) e della notocorda (Wallace, 1982); si suppone però che la 5-HT riscontrata in questi distretti sia di origine materna in quanto negli embrioni non sono ancora comparsi i neuroni che la producono.
Più tardivamente nell’embriogenesi, quando i neuroni serotoninergici sono differenziati, si può riscontrare la presenza di 5-HT nel talamo, nella corteccia limbica, nell’ipotalamo, nella retina e nel nucleo olivare superiore (Lebrand et al., 1996, Cases et al., 1998; Upton et al., 1999). Tra le fibre che contengono 5-HT senza essere in grado di produrla meritano particolare attenzione quelle che innervano la corteccia sensoriale (D’Amato et al., 1987); infatti queste fibre non originano dai nuclei del
raphe ma sono assoni talamo-corticali che contengono 5-HT e che molto
probabilmente la internalizzano mediante il trasportatore e la rilasciano usandola come neurotrasmettitore “in prestito” (Lebrand et al., 1996).
Infine nell’organismo adulto si può riscontrare la presenza di 5-HT anche nelle piastrine, che la ricaptano dal circolo sanguigno e la rilasciano quando sono attivate.
Il sistema serotoninergico dei vertebrati ha una vastissima distribuzione nel sistema nervoso centrale e influenza quasi tutte le funzioni fisiologiche dei mammiferi, dalla regolazione del sistema cardiovascolare (Myata et al., 2000; Nebigil et al., 2000; Thorin et al., 1990), della respirazione, del sistema gastrointestinale (Kato et al., 1999) alla termoregolazione e molte altre. Inoltre è coinvolto nel mantenimento del ritmo circadiano e nel controllo dell’appetito, dell’aggressività, del comportamento sessuale, dell’umore, dell’apprendimento e della memoria.
Un’alterata regolazione della trasmissione serotoninergica incide su numerosi aspetti comportamentali e può risultare alla base di patologie neuropsichiatriche come ad esempio la schizofrenia, l’ansia, lo stress, la depressione, i disordini alimentari. Per questo motivo, molte sostanze che modulano il sistema serotoninergico sono la base di trattamenti farmacologici per queste malattie.
Inoltre stanno emergendo evidenze di un coinvolgimento di questo neurotrasmettitore in patologie come la sindrome di Down, l’autismo, la malattia di Alzheimer e l’epilessia (Sodhi and Sanders-Bush, 2004).
Le connessioni neuronali del SNC presentano un’enorme plasticità e vengono riarrangiate in base agli stimoli che i neuroni ricevono dall’ambiente che li circonda e questo le rende suscettibili anche a fattori come lo stress, la malnutrizione, i farmaci e gli ormoni. Negli ultimi anni stanno emergendo sempre più elementi che sostengono l’ipotesi che la 5-HT, prima di agire come un neurotrasmettitore importante per l’encefalo adulto, agisca come fattore chiave nella modulazione della plasticità sinaptica e dello sviluppo, sia del sistema nervoso centrale che di altri tessuti.
1.2. Ruolo della serotonina nello sviluppo embrionale
La 5-HT è una molecola filogeneticamente antica e numerosi studi suggeriscono l'idea che, come altri neurotrasmettitori, svolga durante lo sviluppo embrionale un ruolo diverso da quello svolto nel sistema nervoso maturo: essa funzionerebbe come segnale di regolazione nei processi di differenziamento e/o di morfogenesi (Turlejsky, 1996).La 5-HT è infatti presente all’interno degli ovociti e degli embrioni precoci di vertebrati ed invertebrati, dal riccio di mare ai mammiferi. Essa si ritrova ancor prima della comparsa delle strutture nervose (Buznikov et
al., 2005; Levin et al., 2006). E’ prodotta dalle cellule staminali embrionali
(Walther et al., 1999) e probabilmente gioca un ruolo importante nella maturazione dell’uovo (Buznikov et al., 1993), nel controllo della segmentazione e della frequenza con cui le cellule si dividono, nel patterning delle strutture citoscheletriche (Lee et al., 1991), nell’adesione tra blastomeri, nei movimenti di gastrulazione (Hamalainem and Kohonen, 1989; Colas et al., 1999), nel controllo della comunicazione mediante giunzioni gap (Moore and Burt, 1995; Rorig and Sutor, 1996), nello stabilirsi dell’asimmetria sinistra-destra (Levin et al., 2006), nella specificazione dell’identità neurale e delle connessioni all’interno del sistema nervoso in via di sviluppo (Lauder et al., 1983; Whitaker-Azmitia, 2001; Fukumoto et al., 2005).
Tra i sistemi monoaminergici, quello serotoninergico è il primo a comparire nell’embriogenesi precoce (Hamon and Bourgoin, 1979).
Molti fattori di crescita che influenzano lo sviluppo del sistema serotoninergico sono anche importanti negli eventi di plasticità sinaptica. Tra questi, quello più studiato è il fattore di crescita astrogliale, S100β, i cui livelli sono aumentati dalla 5-HT, a prova del fatto che i neuroni serotoninergici possono regolare la loro stessa crescita oltre a quella dei loro “targets” (Whitaker-Azmitia, 2001).
Il sistema serotoninergico è anche in grado di autoregolare il proprio differenziamento attraverso l’attivazione del recettore 5-HT1A, del
“brain-derived neurotrophic factor” (BDNF) e il suo recettore trkB (Sodhi and Sanders-Bush, 2004).
Il rilascio di 5-HT dai terminali sinaptici può influenzare i processi di neurogenesi (Lauder and Krebs, 1976), di apoptosi, di rifinimento dendritico, di migrazione cellulare, e plasticità sinaptica (Chubakov et al., 1986; Lauder, 1990). La combinazione di questi eventi dà origine alle sofisticate organizzazioni dell’ippocampo e delle mappe della corteccia somatosensitiva.
Più tardivamente nello sviluppo, invece, la 5-HT regola la crescita dendritica, la formazione delle spine dendritiche e le arborizzazioni (Faber and Haring, 1999).
Il ruolo della 5-HT nello sviluppo è stato studiato con approcci diversi. Uno di questi è l’inibizione della sintesi di 5-HT somministrando paraclorofenilalanina (PCPA), un composto che agisce bloccando la triptofano idrossilasi. In seguito a questo trattamento si osserva un aumento della sintesi dell’ossido nitrico nel corpo calloso, nello striato e nell’ippocampo che mette in evidenza l’esistenza di una stretta relazione tra il sistema serotoninergico e quello nitrergico durante lo sviluppo (Ramos et al., 2002). Questo dato risulta interessante alla luce del fatto che anche per l’ossido nitrico è stato ipotizzato un ruolo nella regolazione della plasticità sinaptica. Inoltre da questi studi emerge che la corretta densità di spine dendritiche dell’ippocampo e del raphe dipende strettamente dalle proiezioni serotoninergiche in queste aree (Alves et al., 2002).
E’ stato ipotizzato che parte dell’azione di questa amina nella plasticità neuronale si esplichi attraverso il controllo dell’espressione di molecole di adesione cellulare quali le PSA-NCAM, che negli organismi presi in considerazione risultano notevolmente ridotte (Brezun and Daszuta, 2000).
In ratti a cui la PCPA viene somministrata subito dopo la nascita, mediante iniezione nella regione lombare, è possibile osservare come la deplezione di 5-HT influisca sulla maturazione dei neuroni motori; infatti
questi animali riportano disfunzioni posturali probabilmente dovute ad un arresto dello sviluppo di questi motoneuroni (Pflieger et al., 2002).
Un altro approccio sperimentale molto utilizzato è la generazione di topi “knock-out” (KO) per alcuni dei geni coinvolti nello sviluppo e/o nella funzionalità del sistema serotoninergico. Dallo studio del topo “knock-out” per la monoaminoossidasi A (MaoA), l’enzima responsabile della degradazione della 5-HT, sono emersi molti dati interessanti. Innanzitutto l’inattivazione di questo enzima porta ad un aumento, pari a nove volte, dei livelli di 5-HT nell’encefalo durante la prima settimana dopo la nascita. In questo periodo si riscontra un forte accumulo di 5-HT in tutti i neuroni che esprimono in modo transiente il trasportatore serotoninergico (SERT). La corteccia somatosensoriale di questi topi non presenta più la sua tipica divisione in colonne, che sono la base morfologica delle mappe corticali, a dimostrazione del fatto che l’aumento di 5-HT compromette la corretta aggregazione e segregazione delle fibre talamocorticali (Cases et al., 1996, 1998). Simili effetti si ritrovano nel sistema visivo dove viene alterata la normale segregazione degli assoni retinici (Upton et al., 1999). Altre alterazioni riscontrate in questo modello riguardano il controllo locomotorio e respiratorio nei neonati (Bou-Flores et al., 2000).
Nel topo ”knock out” per il trasportatore serotoninergico (SERT), la 5-HT continua ad essere secreta ma non viene rimossa dal vallo sinaptico. Questo comporta un ristagno sinaptico della 5-HT ed un aumento dei livelli basali di 5-HT extracellulare a carico del telencefalo (Mathews et al., 2004) quindi uno sviluppo anormale degli assoni talamocorticali e retinici, molto simile a quello osservato nei topo “knock out” per MaoA. Questo indica che un eccesso di 5-HT a livello dello spazio extracellulare, causa modificazioni a carico dello sviluppo (Upton et al. 1999; Persico et al., 2001). I topi Sert-/- mostrano inoltre un aumento del comportamento legato ad ansia e depressione e una riduzione del comportamento aggressivo (Holmes et al.,2002; Holmes et al., 2003) anche se ciò sembra un paradosso se si considera che proprio l’inibizione del trasportatore è alla base della maggior parte delle cure antidepressive (Bengel et al.,
1998; Lesch et al., 2003). Altri neuroni potrebbero invece beneficiare dell’aumentata stimolazione dei recettori serotoninergici come indicato da una diminuzione del fenomeno di morte cellulare a carico del telencefalo dei topi “knock out” per SERT (Persico et al., 2003).
La produzione di topi “knock-out” per la Tph1, ha messo in evidenza che questa è fondamentale per lo sviluppo corretto del cuore. I topi KO per Tph1 hanno un cuore di dimensioni maggiori rispetto al cuore dei “wild type” e sono affetti da disfunzioni cardiovascolari progressive che comportano l’insorgenza di difetti cardiaci (Cote et al., 2003). La distruzione del gene Tph1, induce una perdita drammatica di 5-HT a livello periferico ed è proprio per questo che i topi KO per Tph1 rappresentano uno strumento assai utile per indagare il ruolo della 5-HT materna durante lo sviluppo. In uno studio del 2007, femmine KO per Tph1 vengono fatte accoppiare sia con maschi “wild type” che con maschi eterozigoti o KO per Tph1. Cote e colleghi hanno dimostrato che le anomalie a carico dello sviluppo cerebrale e di altri tessuti riportate dalle cucciolate, erano strettamente correlate al genotipo materno. La 5-HT di origine materna è quindi necessaria per un corretto sviluppo embrionale (Cote et al., 2007).
Altri studi condotti sul topo mostrano un coinvolgimento della 5-HT materna nella migrazione delle cellule delle creste neurali facciali (Yavarone et al., 1993; Moiseiwitsch and Lauder, 1995), nella morfogenesi delle strutture craniofacciali (Shuey et al., 1992, 1993; Bhasin et al., 2004) e nella proliferazione delle cellule del miocardio (Nebigil and Maroteaux, 2001). Infatti, embrioni in coltura trattati con antagonisti serotoninergici hanno uno sviluppo embrionale anormale caratterizzato da una deficienza del mesenchima della testa, da archi mandibolari ipoplastici, da occhi e tubo neurale anormali e da difetti cardiovascolari che includono la mancanza di trabecole e la riduzione dello spessore della parete ventricolare (Choi et al., 1997).
Più recentemente, sono state generate linee di topi “knock out” per il gene Tph2 (Gutknecht et al., 2008; Savelieva et al., 2008). Savelieva e colleghi hanno generato sia una linea di topi “knock out” per Tph2 che una
linea di topi “doppio knock out” per Tph1/2. Sorprendentemente, nessuna di queste mutazioni ha dato origine ad un fenotipo palese. Sia i topi KO per Tph2 che quelli KO per Tph1/2, erano più piccoli e meno pesanti rispetto ai topi “wild type” ma risultavano pressoché normali se sottoposti a test comportamentali. Nonostante ciò, sono state registrate evidenti differenze comportamentali in seguito alla somministrazione di sostanze come gli SSRI o preparati che agivano a livello dei recettori serotoninergici. Alenina e colleghi (2009) hanno inoltre asserito che la mancanza di 5-HT nel cervello di topi Tph2-/-, conferma che Tph2 è il principale enzima adibito alla sintesi della 5-HT cerebrale. In questi topi, la mancanza di 5-HT a livello centrale, comporta una compromissione a carico delle fasi precoci della crescita postnatale e un’alterazione del controllo del sonno, della respirazione, della termoregolazione, del battito cardiaco e della pressione sanguigna. I topi sembrano inoltre più aggressivi e le femmine trascurano la prole. Nonostante ciò, i topi che sono quasi totalmente privi di 5-HT a livello cerebrale, sono vitali, morfologicamente normali e fertili. E’ quindi possibile concludere che la 5-HT sintetizzata dall’enzima Tph2 è coinvolta nella regolazione del comportamento e nel controllo delle funzioni autonome ma non è essenziale per la mera sopravvivenza dell’adulto (Alenina et al., 2009).
In conclusione, tutti i dati che sono noti sino ad oggi dimostrano come il sistema serotoninergico sia sicuramente coinvolto in processi di formazione dei circuiti neuronali e di sinaptogenesi. Rimangono ancora da chiarire, però, sia il ruolo di questo sistema nello sviluppo precoce, cioè in processi quali la proliferazione, differenziamento, migrazione e i processi morfogenetici, sia i meccanismi molecolari che sono alla base di questa sua azione.
1.3. I recettori della serotonina
La grande varietà di funzioni in cui abbiamo visto essere implicata la 5-HT si riflette nella complessità farmacologica dei suoi recettori; infatti finora sono stati clonati quindici geni che codificano per recettori serotoninergici che danno origine a venti forme diverse mediante processi di “splicing” alternativi o di mRNA “editing”. Tutti questi recettori sono stati classificati, in base alle caratteristiche farmacologiche, strutturali e trasduzionali, in sette famiglie (Hoyer et al., 1994). Le classi HT1, 5-HT2, 5-HT4, 5-HT5, 5-HT6 e 5-HT7 sono tutte accoppiate a proteine G; di queste, 5-HT1 è accoppiata a Gi , 5-HT2 a Gq, 5-HT4/6/7 a Gs e 5-HT5
ancora non è noto. La classe 5-HT3, invece, è un recettore canale ionico. Tra tutti questi recettori quelli per cui è noto un coinvolgimento nello sviluppo embrionale sono i recettori 5-HT1A, 5-HT1B, 5-HT2A, 5-HT2B e 5-HT2C.
Il recettore 5-HT1A, clonato nei mammiferi (Raymond et al., 2001) e in
Xenopus laevis (Marracci et al., 1997), è espresso precocemente nella vita
embrionale ed è localizzato principalmente nei nuclei del raphe, nell’ippocampo e nell’occhio ma si ritrova transientemente anche nei motoneuroni spinali e nel cervelletto (Hillion et al., 1994; Miquel et al., 1994). Studi in vivo mostrano che il suo ruolo si esplica soprattutto nel controllo della neurogenesi nell’adulto e della maturazione dendritica dell’ippocampo. Se attivato, infatti, stimola neurogenesi nel giro dentato e nella zona subventricolare, aree che sono sede di produzione di neuroblasti a lunga vita (Brezun et al., 2000; Gould, 1999; Santarelli et
al., 2003). Inoltre è stato visto che una deplezione postnatale di 5-HT
riduce la lunghezza dei dendriti e il numero delle spine dendritiche dei neuroni ippocampali e questo effetto dipende dal recettore 5-HT1A. Per questo recettore sono state generate tre linee di topi “knock-out” (Parks
et al.,1998; Ramboz et al., 1998; Heisler et al., 1998). Tutte e tre le linee mostrano un aumento dei comportamenti legati all’ansia ma una diminuzione di quelli legati alla depressione. Questa situazione può essere
spiegata perché l’assenza del recettore, provocando un aumento della trasmissione serotoninergica, ha un effetto antidepressivo ma ansiogeno (Blier & de Montigny, 1994; Griebel, 1995). Da questi studi emerge che la 5-HT è necessaria perché i topi adulti abbiano un normale comportamento legato all’ansia e che questo suo ruolo è mediato dal recettore 5-HT1A (Gross et al., 2002).
Il recettore 5-HT1B ha un profilo di espressione precoce e dinamico; è localizzato nei nuclei del raphe, nello striato, nel cervelletto, e nelle cellule gangliari della retina (Boschert et al., 1994). Inoltre, transientemente lo si ritrova nei nuclei sensoriali talamici nel periodo immediatamente dopo la nascita. In tutti questi neuroni, il recettore è sempre espresso sui terminali assonali presinaptici dove modula il rilascio di glutammato (Laurent et al., 2002).
Studi effettuati sul topo “knock-out” per MaoA e per il trasportatore della serotonina SERT mettono in evidenza che i difetti di segregazione assonale tipici in questi due modelli sono recuperabili mediante l’inattivazione di questo recettore. Ciò dimostra che il recettore 5-HT1B è in grado di influenzare selettivamente lo sviluppo degli assoni dei neuroni talamici (Salichon et al., 2001; Rebsam et al., 2002; Upton et al., 2002). Questo recettore agisce attraverso l’inibizione della produzione di AMP ciclico e dell’entrata di calcio nei terminali assonici (Raymond et al., 2001).
Il recettore 5-HT2A è espresso tardivamente nello sviluppo e per questo motivo si pensa sia implicato in processi di maturazione e differenziamento neuronale (Azmitia, 2001). Esso è ampiamente espresso a livello cerebrale (corteccia, nucleo caudato, tubercolo olfattivo, nucleo accumbens e ippocampo) (Pazos et al., 1985), nei muscoli scheletrici (Guillet-Deniau et al., 1997; Hajduch et al., 1999) e lisci (Kuemmerle et
al., 1995; Ellwood and Curtis, 1997), nei reni (Garnovskaya et al., 1995)
e nelle piastrine (Cook et al., 1994). Nel cervello, questo recettore rappresenta il sito d’azione di sostanze allucinogene come LSD
(Aghajanian and Marek, 1999) e di anti-psicotici atipici come clozapina e risperidone (Adell et al., 2005).
Il topo “Knock-out” per questo recettore però non presenta sostanziali anomalie a livello del sistema nervoso centrale. Sembra invece che siano riconducibili ad una sua eccessiva attivazione le disfunzioni motorie che si notano nel topo privo di MaoA perché si possono recuperare somministrando un suo antagonista (Bou-Flores, et al., 2000). Il recettore 5-HT2A è coinvolto nella modulazione dell’espressione del “brain-derived neurotrophic factor” (BDNF) nella neocorteccia e nell’ippocampo attraverso cui influenza i processi tardivi dello sviluppo di queste aree (Vaidya et al., 1997).
Il recettore 5-HT2B e il suo ruolo nello sviluppo saranno discussi nel paragrafo successivo.
Il cDNA del recettore 5-HT2C è stato clonato nei mammiferi (Raymond et al., 2001) e in Xenopus laevis (De Lucchini et al., 2003). Nei gatti neonati, ha un’espressione che rispecchia la caratteristica struttura colonnare del IV strato della corteccia visiva durante il periodo critico per la formazione della dominanza oculare. La sua deplezione provoca l’alterazione della formazione della dominanza oculare. Questo supporta l’ipotesi che questo recettore sia implicato nello sviluppo sinaptico della corteccia visiva (Wang et al., 1997; Kojic et al., 1997). Inoltre i topi che mancano di 5-HT2C riportano dei deficit nel potenziamento a lungo termine dell’ippocampo, a ulteriore conferma del suo ruolo nella plasticità sinaptica (Edagawa, 2001).
L’attivazione di questo recettore inibisce l’attività dei neuroni dopaminergici (Gobert et al., 2000; Ji et al., 2006) infatti i farmaci anti- psicotici atipici bloccano l’inibizione garantendo così l’attivazione del sistema dopaminergico (Gobert et al., 2000; De Deurwaerdere et al., 2004; Berg et al., 2005). Questa attivazione potrebbe essere importante ai fini del trattamento contro depressione e sintomi negativi della schizofrenia.
1.4. Il recettore 5-HT2B
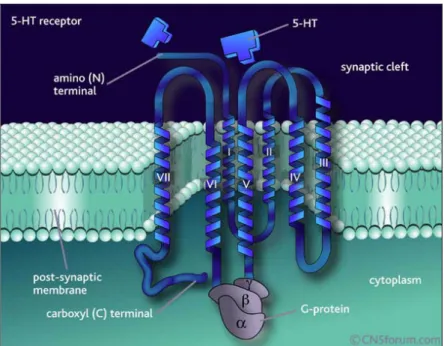
Il cDNA del recettore 5-HT2B è stato clonato nel ratto e nel topo (Foguet et al., 1992), nell’uomo (Kursar et al., 1994) e in Xenopus laevis (De Lucchini et al., 2003). Nella struttura è molto simile a tutti i recettori accoppiati a proteine G; è una proteina integrale di membrana, di 450-500 aminoacidi (il numero varia a seconda delle specie considerate) e con sette domini idrofobici transmenbrana, connessi da tre domini idrofilici intracellulari e tre extracellulari (Fig. 1.3).
Fig. 1.3: modello tridimensionale di un recettore serotoninergico accoppiato a proteine G. E’ una
proteina integrale di membrana, con sette domini idrofobioci transmembrana e ha l’estremità amminica nel versante extracellulare e quella carbossilica nel versante citosolico.
Il terminale amminico della sequenza è orientato verso lo spazio extracellulare mentre il terminale carbossilico verso quello intracellulare. Lungo i domini extracellulari sono presenti dei siti canonici di glicosilazione, segno di possibili modificazioni post-traduzionali, e dei residui di cisteina che permettono la formazione di ponti disolfuro e conseguenti modificazioni conformazionali della proteina. A livello dei
domini intracellulari, invece, si trovano dei siti per l’interazione con proteine Gq/11 e altre proteine regolatorie, e dei siti di fosforilazione,
bersaglio per diverse serina/treonina chinasi. Inoltre a livello del terminale carbossilico c’è un motivo molto conservato chiamato S/TXV che interagisce con il dominio PDZ delle proteine che lo contengono; questo dominio è coinvolto nelle vie di trasduzione del segnale attivate da cNOS (sintasi dell’ossido nitrico costitutiva) e iNOS (sintasi dell’ossido nitrico inducibile) (Raymond et al., 2001). Sempre a questo terminale sono presenti dei siti di palmitoilazione e fosforilazione che sono indispensabili per l’internalizzazione del recettore (Parker et al., 2003).
Nell’uomo, l’RNA messaggero per il 5-HT2B è espresso abbondantemente nel fegato e nel rene e a livelli più bassi nel pancreas, nella milza, nel cervelletto, nell’ipotalamo dorsale, nel setto laterale e nell’amigdala mediale (Bonhaus et al., 1995; Duxon et al., 1997).
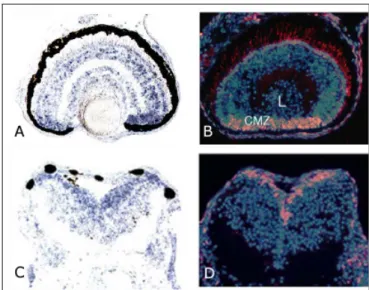
Inoltre studi condotti in Xenopus laevis hanno messo in evidenza che l’RNA messaggero di X5-HT2B, durante lo sviluppo, è presente nei precursori neuronali in proliferazione del tubo neurale e della retina (De Lucchini et al., 2003).
Figura 1.4: in A e C, espressione dell’mRNA di 5-HT2B negli strati nucleare interno ed esterno e
nella zona del margine ciliare della retina e nel tubo neurale di embrioni di Xenopus. In B e D, localizzazione dei precursori neuronali in proliferazione della retina e del tubo neurale di embrioni di
Xenopus, mediante un saggio per l’incorporazione della BrdU. Dal confronto delle quattro immagini
La sua espressione nell’embriogenesi lo rende un ottimo candidato per la mediazione degli effetti morfogenetici serotonina-dipendenti (Fig.
1.4).
Studi effettuati sul topo hanno messo in evidenza la presenza dell’RNA messaggero per questo recettore durante l’embriogenesi precoce anche nelle cellule delle creste neurali, nel tubo neurale, nel cuore, nell’intestino, nei somiti, nelle vescicole ottiche e otiche, negli archi faringei e nel mesenchima craniofacciale (Choi et al.,1997).
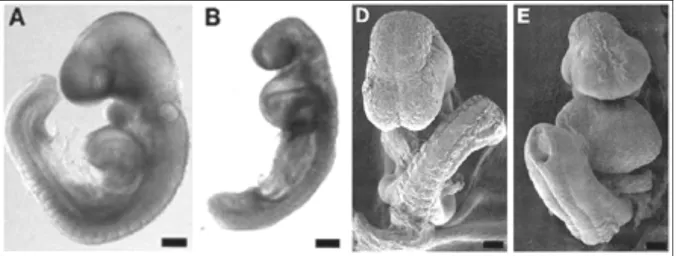
Le prime evidenze, in vivo, del ruolo di questo recettore nello sviluppo derivano da uno studio fatto su embrioni di topo trattati con la ritanserina, antagonista dei recettori 5-HT2 (Fig. 1.5).
Fig. 1.5: In A e D, embrioni di topo “wild type” a 9 giorni di gestazione, in visione laterale (A) e
frontale (D). In B e E, embrioni di topo, trattati per 24 ore con ritanserina, a 9 giorni di gestazione. Dal confronto emergono il forte ritardo dello sviluppo e le gravi alterazioni a carico delle regioni cefaliche e del cuore dei topi trattati rispetto ai controlli. Da: Choi et al., 1997.
Questi embrioni mostrano un forte ritardo della crescita, un’anomalia della piega cefalica, prosencefalo e rombencefalo sottosviluppati, archi branchiali ipoplastici, difetti del numero e della forma dei somiti, della chiusura e della forma del tubo neurale e malformazioni cardiache tra cui edema pericardico e assenza delle trabecole miocardiche. Gli embrioni trattati presentano anche un aumento del numero delle cellule delle creste neurali craniali che vanno in apoptosi. Da questo studio
è emersa l’ipotesi che il recettore 5-HT2B possa avere un ruolo importante nei processi di migrazione, proliferazione cellulare e sopravvivenza delle cellule delle creste neurali e nella migrazione e/o nell’inibizione del differenziamento cellulare delle cellule del miocardio (Choi et al., 1997).
Il fenotipo dei topi trattati con ritanserina è paragonabile a quello di topi trattati con acido retinoico, fattore che regola negativamente la proliferazione e la migrazione delle cellule creste neurali. Questa osservazione, aggiunta alla presenza di un sito di legame per l’acido retinoico nella sequenza del promotore di 5-HT2B, aveva fatto ipotizzare che l’acido retinoico regolasse negativamente il recettore 5-HT2B ma uno studio condotto sul topo smentisce questa ipotesi suggerendo che più che avere un’azione diretta l’uno sull’altro, i due fattori possano agire su elementi comuni (Bhasin et al., 2004).
Il topo “knock-out” per 5-HT2B ha un fenotipo letale, con diversi gradi di penetranza, embrionale o neonatale, causato da malformazioni cardiache. I topi che sopravvivono alla nascita mostrano dilatazione cardiaca con una penetranza del 100% e un’anomalia nell’organizzazione miofibrillare e nelle giunzioni cellulari cardiache. Il fenotipo di questo topo sembra dovuto a difetti della proliferazione e della crescita delle cellule miocardiche nello sviluppo. Ciò supporta l’ipotesi che la 5-HT, attraverso il suo recettore 2B agisca come fattore mitogenico sui cardiomiociti. Il confronto di questi topi con quelli “knock-out” per ErbB2, recettore tirosina chinasico di una neuroregulina, suggerisce che i due recettori 5-HT2B e ErbB2 sfruttino la medesima via di trasduzione del segnale durante lo sviluppo del cuore (Nebigil and Maroteaux, 2001). Gli stessi autori che hanno studiato il topo “knock-out” per 5-HT2B hanno proseguito le ricerche su cardiomiociti in coltura dimostrando che in condizioni di stress, la 5-HT protegge le cellule da morte apoptotica e che questo effetto è mediato dal recettore 5-HT2B (Nebigil et al., 2003).
Un altro approccio per lo studio del ruolo di 5-HT2B è stato quello di costruire un topo transgenico che sovraesprimesse questo gene. Questi topi mostrano ipertrofia cardiaca, dovuta ad un aumento sia del numero
che della crescita dei cardiomiociti, accompagnata da proliferazione e attività enzimatica mitocondriale anormali (Nebigil et al., 2003).
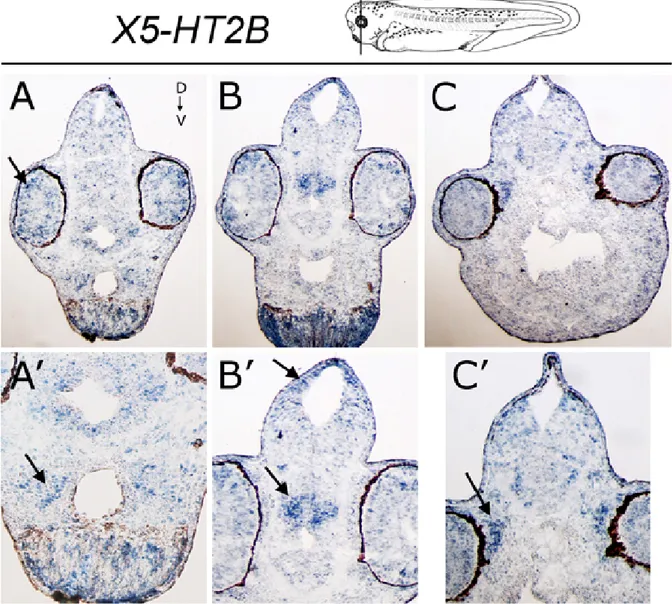
Nel nostro laboratorio è stato analizzato il “pattern” di espressione del recettore 5-HT2B in Xenopus, mettendo in evidenza la sua espressione a livello della retina, degli archi branchiali, in cellule sparse del tubo neurale e del diencefalo e a livello del mesenchima perioculare (Figura
1.6).
Figura 1.6 Embrioni di Xenopus a stadio 37 processati per ibridazione in situ con la sonda complementare all’mRNA del recettore 5-HT2B. La marcatura è presente nella retina (freccia in A), nel primo arco branchiale (freccia in A’), in cellule sparse del tubo neurale e del diencefalo (freccia in B’) e nel mesenchima perioculare (freccia in C’)
maniera “cell autonomous” il comportamento delle cellule post migratorie scheletogeniche delle creste neurali craniali del primo arco branchiale, partecipando alla formazione dell’articolazione mascella-mandibola. (De Lucchini et al., 2005; Reisoli et al., 2008; Reisoli et al., 2010).
L’azione del recettore 5-HT2B si esplica attraverso diverse cascate di trasduzione che sono state studiate mediante esperimenti in vitro.
Come tutti i recettori della 5-HT di tipo 2 è accoppiato, sia ad una proteina Gq/11 che attiva la fosfolipasi Cβ (PLCβ) e porta alla produzione di
IP3 e diacilglicerolo (DAG), sia ad un’altra proteina G che attiva la
fosfolipasi A2 (PLA2), che a sua volta stimola la produzione di acido arachidonico (AA) (Raymond et al., 2001).
Questo recettore interviene nella regolazione del rilascio di Ca2+ dalle riserve intracellulari sia attraverso il coinvolgimento dei recettori rianodinici (Ullmer et al., 1996) sia attraverso una via IP3- dipendente che
porta all’apertura di un canale Cl-, Ca2+ -dipendente (Foguet et al., 1992;
Parekh et al., 1993).
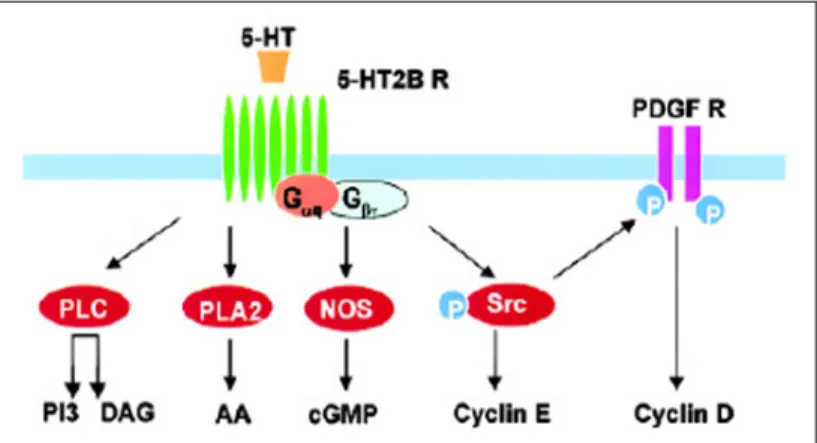
Il recettore 5-HT2B inoltre, può stimolare la progressione del ciclo cellulare attraverso due vie indipendenti e parallele che portano all’attivazione dei complessi chinasici ciclina D1/cdk4 e ciclina E/cdk2. L’attivazione della ciclina D1 sembra coinvolgere la transattivazione della via di trasduzione del segnale del recettore chinasico del fattore di crescita derivato dalle piastrine (PDGF); la ciclina E, invece non richiede l’intervento di questo recettore. A monte di entrambe queste vie si ritrova un elemento comune che è la chinasi Src (Nebigil et al., 2000b; Raymond
et al., 2001).
Il recettore 5-HT2B è in grado di stimolare anche la produzione intracellulare di cGMP attraverso l’attivazione delle sintasi dell’ossido nitrico costitutiva e inducibile (cNOS e iNOS). L’interazione con questi due enzimi avviene a livello dei domini PDZ presenti al terminale carbossilico del recettore, ma dato che solo la via di iNOS coinvolge l’attivazione di una proteina G13 si pensa che anche in questo caso il recettore possa agire
Fig. 1.7: schema delle principali vie di trasduzione del segnale attivate dal legame della serotonina
al recettore 5-HT2B. Da: Nebigil et al., 2000
Esiste anche un’altra via di azione del recettore 5-HT2B: quella antiapoptotica. E’ stato visto come l’accoppiamento di 5-HT2B con una proteina Gq porti all’attivazione, in caso di stress, sia della via di
ERK1/ERK2 (extracellular signal-regulated kinase), che di quella di PI3K/Akt (fosfaditilinositolo 3 chinasi), che rispettivamente inibiscono, indipendentemente, Bax e ANT-1, fattori proapoptotici; ciò impedisce il rilascio del citocromo c dal mitocondrio e, di conseguenza, l’attivazione della caspasi 9 e l’entrata in apoptosi della cellula. La via PI3/Akt oltre ad inibire il rilascio di ANT-1, stimola la degradazione di IкB-α e permette la traslocazione nel nucleo del fattore di trascrizione NF-кB (Fig. 1.8).
1.5. Morfogenesi oculare
Lo sviluppo dell’occhio nei vertebrati è un complesso processo che coinvolge molteplici interazioni induttive tra il neuroectoderma del prosencefalo (che dà origine alla retina e al nervo ottico), l’ectoderma superficiale (che dà origine alla lente, all’epitelio della cornea, alla congiuntiva e alle palpebre), e le cellule delle creste neurali da cui deriva parte del mesenchima perioculare (che darà origine alla coroide, alla sclera, allo stroma della cornea, all’iride e alla camera anteriore) (Johnston et al., 1979; Trainor and Tamm 1995; Cvekl and Tamm, 2004)
(Fig. 1.9). Molte strutture del segmento anteriore originano dalla unione
di due differenti tessuti embrionali come l’ectoderma e il mesenchima. Nella cornea, ad esempio, l’epitelio corneale è di derivazione ectodermica, mentre lo stroma della cornea e l’endotelio sono di derivazione mesenchimatica (Gage et al.2005).
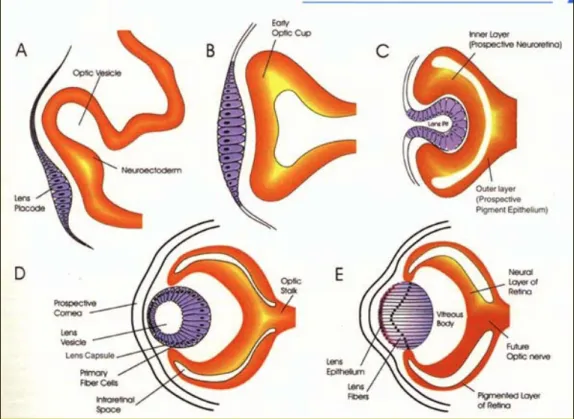
Figura 1.9: schema dello sviluppo dell’occhio nei vertebrati. La vescicola ottica evagina dal diencefalo e
promuove la formazione del placode della lente (A,B), poi si invagina diventando una coppa ottica bi-stratificata (C,D). E: anatomia dell’occhio completamente sviluppato.
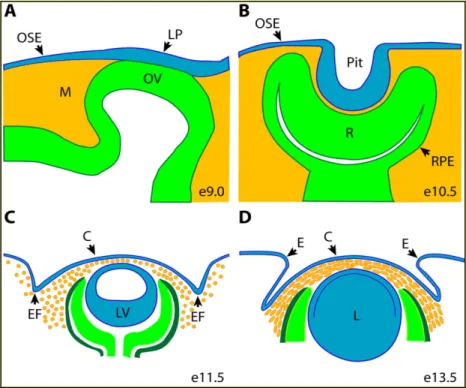
Studi condotti sul topo hanno mostrato i diversi eventi richiesti per un corretto sviluppo oculare e per la normale morfogenesi del segmento anteriore dell’occhio. La “optic pits”, prima struttura morfologica oculare a formarsi, verso lo stadio embrionale E9 del topo evagina lateralmente dal neuroepitelio del diencefalo ventrale a formare la vescicola ottica (Pei e Rhodin 1970; Kaufman, 1992), fino ad incontrare la superficie oculare dell’ectoderma (OSE). La OSE è uno strato multipotente che quando opportunamente stimolato contribuisce alla formazione della lente, della cornea e della congiuntiva.
Il segmento anteriore dell’occhio inizia la sua formazione quando la vescicola ottica distale prende contatto con l’ OSE sovrastante. Nei topi questo contatto avviene allo stadio embrionale E9.5 e risulta nell’induzione del placode del cristallino all’interno dell’OSE (Pei e Rhodin 1970; Kaufman, 1992). Il placode del cristallino si invagina verso la linea mediana della testa formando il precursore della lente, in contemporanea la vescicola ottica si invagina anch’essa formando il calice ottico costituito da due strati che si differenziano diversamente: lo strato esterno forma la retina pigmentata, mentre lo strato interno forma la retina nervosa.
Durante la fase (E10) si ha anche la formazione della cornea e di altre strutture del segmento anteriore, la cui morfogenesi ha inizio con la migrazione del mesenchima in gran parte di derivazione delle creste neurali, che migra nel territorio tra l’abbozzo della lente e l’OSE (Pei e Rhodin, 1970; Kaufman, 1992; Cvekl and Tamm, 2004). Durante questa fase una sottopopolazione di cellule delle creste neurali che migrano nel primo arco faringeo darà origine, insieme alle cellule mesenchimatiche della testa di origine mesodermica, al mesenchima perioculare (POM). Il POM è perciò una struttura formata da cellule mesenchimali che circondano l’occhio nelle prime fasi di sviluppo. (Fig. 1.10)
In particolare la sottopopolazione di cellule del POM che origina dalle creste e si porta sotto l’ectoderma dell’occhio in formazione darà origine alle strutture del segmento anteriore. Nella periferia dell’occhio invece,
queste cellule formano le cartilagini e le ossa orbitali, vasi sanguigni, tessuti connettivi associati ai muscoli extraoculari.
Fig. 1.10: Schematizzazione della morfogenesi del segmento anteriore dell’occhio. Ectoderma
(blu), ectoderma neurale (verde), e cellule della cresta neurale e cellule mesenchimatiche di origine mesodermica (arancione). (Gage and Zacharias, 2009).
Esperimenti classici suggeriscono che segnali provenienti da tessuti circondanti l’occhio, sono essenziali durante il suo sviluppo precoce. Holtfreter (1939) osservò in espianti in coltura, derivati dalla piastra neurale anteriore di anfibio, che in assenza di contatto con l’epidermide e con il mesenchima di derivazione dalle creste neurali, lo sviluppo dell’occhio si arrestava allo stadio di vescicola ottica (Fuhrmann et
al.,2000).
La funzione principale del POM è quella di fornire multipli “lineages” cellulari maturi che sono necessari per il corretto sviluppo del segmento anteriore dell’occhio (che comprendono lo stroma e l’endotelio corneale, la camera anteriore e lo stroma dell’iride). Inoltre è necessario che le cellule originate dall’epitelio superficiale interagiscano con il POM per il corretto sviluppo della palpebra (Le Lievre and Le Douarin, 1975). Infine, il POM
fornisce segnali essenziali per il patterning dell’ectoderma oculare primordiale, includendo la specificazione dell’epitelio pigmentato della retina e la differenziazione del nervo ottico a partire dall’ectoderma neurale (Evans and Gage 2005; Fuhrmann et al., 2000, Matt et al., 2008). Per quanto riguarda i meccanismi molecolari coinvolti nelle diverse funzioni del POM, studi condotti sul topo e sul pollo hanno fornito l’evidenza del legame tra il segnale dell’acido retinoico proveniente dalla coppa ottica, il fattore di trascrizione con omeodominio Pitx2 che si trova espresso all’interno delle creste neurali, e il segnale canonico Wnt/β-catenina all’interno dell’ectoderma superficiale oculare. Pitx2 è un nodo critico di integrazione che lega le due vie di segnalazione. Questa rete di interazioni molecolari è necessaria durante lo sviluppo dell’occhio precoce e fornisce una buona base per comprendere a fondo i meccanismi alla base dello sviluppo del segmento anteriore dell’occhio. (Gage and Zacharias, 2009).
1.6. Il fattore di trascrizione Pitx2
Il gene Pitx2 è un gene “homeobox” che codifica per un fattore di trascrizione fondamentale nel corso del differenziamento embrionale ed in particolare svolge un ruolo importante nello sviluppo del segmento anteriore dell’occhio (Gage and Zacharias, 2009).
I dati oggi noti che riguardano la funzione di questo gene nello sviluppo embrionale derivano quasi esclusivamente da studi effettuati nel topo. L’espressione genica di Pitx2 durante lo sviluppo del topo è stata riscontrata a livello di occhi, denti, ombelico, adenoipofisi, cuore, intestino e arti. (Hjalt et al.,2000).
Nell’uomo, mutazioni a carico di questo gene provocano la sindrome di Axenfeld Rieger (ARS), patologia genetica autosomica dominante che comprende una serie di malformazioni congenite che affliggono
principalmente il segmento anteriore dell’occhio. Gli individui affetti presentano multiple malformazioni oculari ed extra-oculari quali anomala morfogenesi del segmento anteriore dell’occhio (provocando cecità da glaucoma nel 50% dei soggetti), malformazioni sistemiche con incompleta penetranza ed espressività variabile come malformazioni degli abbozzi dentari, dismorfismo craniofacciale (ipoplasia mascellare o ipertelorismo), difetti cardiaci congeniti (Tümer and Bach-Holm,2009).
Il fattore di trascrizione Pitx2 può essere modificato a livello post-trascrizionale tramite fosforilazione da parte di una PKC. L’attività trascrizionale di Pitx2 viene regolata negativamente se la fosforilazione avviene all’ N-terminale e regolata positivamente se la fosforilazione avvine al C-terminale. Recenti studi hanno messo in evidenza che mutazioni a livello del C-terminale sono associate con la ARS (Espinoza et
al., 2005).
Topi modificati geneticamente, in cui è stato inattivato un allele di
Pitx2, presentano fenotipi molto simili alle caratteristiche della sindrome
(Lu et al., 1999; Gage et al., 1999), topi omozigoti invece muoiono prima della nascita a causa di difetti dello sviluppo del cuore (Evans and Gage, 2005). Pitx2 inoltre è coinvolto nella via di segnalazione Nodal/Sonic hedgehog che determina la polarità destra/sinistra (Campione et al.,1999).
Per determinare lo specifico ruolo di Pitx2 nello sviluppo dell’occhio, è stato generato un topo “knock out” condizionale, in cui il gene Pitx2 è stato inattivato solo a livello delle creste neurali. In questo caso gli animali riescono a sopravvivere, e hanno permesso di analizzare la funzione di
Pitx2 durante lo sviluppo tardivo dell’occhio (Evans and Gage, 2005).
Questi topi oltre ad avere difetti nel segmento anteriore dell’occhio, mostrano un fenotipo in cui gli occhi sono progressivamente disposti lungo la linea mediana, fino al loro diretto attacco all’ipotalamo ventrale.
Ulteriori studi hanno dimostrato che fenotipi simili a quelli riscontrati nella ARS si ottengono nel topo sia facendo sovraesprimere Pitx2 nel mesenchima corneale e nell’iride, sia abrogandone la funzione. Questo ha
messo in evidenza che l’azione di Pitx2 è correlata ad un “effetto dose dipendente”. (Holmberg et al.,2004).
1.7. Xenopus laevis come sistema modello
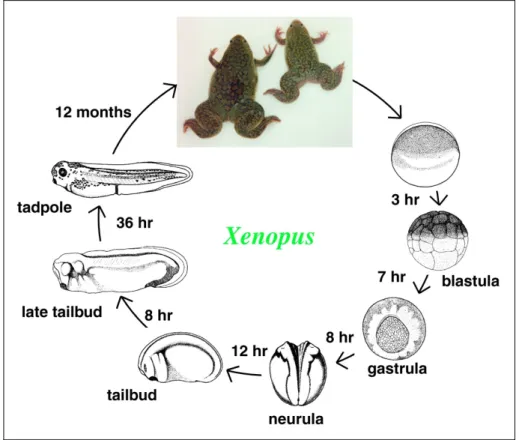
Fig. 1.11: schema dei principali stadi di sviluppo di Xenopus laevis. Da Wolpert et al.,1998.
Fin dalla fine dell’ottocento, per diversi motivi sperimentali molti degli studi volti a capire i meccanismi alla base dello sviluppo embrionale dei vertebrati sono stati svolti negli anfibi. La scelta di queste specie animali come modello sperimentale si è basata sul fatto che le modalità di sviluppo di tutti i vertebrati sono alquanto simili e perciò i meccanismi di base dell’embriogenesi degli anfibi sono validi per tutti i vertebrati, dai pesci all’uomo. Come sistema modello è stato scelto nel nostro laboratorio l’anuro africano Xenopus laevis.
Questo anfibio costituisce un modello di elezione per gli studi sullo sviluppo perché, in qualsiasi periodo dell’anno, la femmina può essere indotta, mediante iniezione dell’ormone umano gonadotropina corionica, a deporre nell’ambiente esterno 1000/1500 uova, che possono essere fecondate con facilità in vitro. Le grandi dimensioni delle uova degli anfibi, circa 1mm di diametro, le rendono particolarmente adatte alla manipolazione sperimentale. Un’altra caratteristica di questo anfibio è rappresentata dalla rapidità con cui si sviluppano gli embrioni: una volta fecondato, l’uovo si divide in due blastomeri in circa 90 minuti e le divisioni successive avvengono in maniera sincrona ogni 20 minuti, fino allo stadio di blastula. L’embriogenesi di Xenopus è stata studiata e descritta da Nieuwkoop e Faber (1967) e quindi mediante la consultazione delle tavole compilate da questi autori è possibile capire a che stadio si trovi l’embrione. Esse individuano infatti le principali modificazioni che distinguono i diversi stadi dello sviluppo. Lo stesso lavoro fornisce un’importante indicazione sui tempi di sviluppo dell’embrione a diverse temperature, rendendo possibile una manipolazione dei tempi stessi che possono arrivare a ridursi di un terzo se gli embrioni sono stabulati a 14°C; questo aspetto ha un evidente vantaggio pratico in quanto permette, se necessario, di rallentare notevolmente la crescita dell’embrione, adattandola alle proprie esigenze sperimentali.
Grazie alle caratteristiche che ho elencato è possibile effettuare sugli embrioni, sin dagli stadi più precoci dello sviluppo, sia trattamenti farmacologici, sia esperimenti di perdita e guadagno di funzione mediante la tecnica della microiniezione in embrioni allo stadio di due o quattro cellule .
Per fare esperimenti di guadagno di funzione si microinietta l’ mRNA del gene di interesse, sovraesprimendolo, mentre per gli esperimenti di perdita di funzione si microiniettano o oligonucleotidi antisenso modificati (morpholino) o mRNA antisenso o mRNA codificanti per proteine mutate o delete. Inoltre, insieme ai costrutti di interesse è possibile microiniettare il
trascritto di un gene reporter come EGFP o βgalattosidasi che permette di visualizzare la zona iniettata.