CAPITOLO 1
INTRODUZIONE
1.1
Il fenomeno delle invasioni: cause e vettori
La distribuzione di una specie è il risultato di un costante tentativo di colonizzazione di nuove regioni che può verificarsi attraverso dispersione marginale (espansione progressiva nelle aree adiacenti) o per dispersione remota (tra regioni con discontinuità spaziale) (Crisp 1958).
Per millenni l’uomo è intervenuto alterando la naturale distribuzione delle specie animali e vegetali, attraverso l’introduzione volontaria (connessa ad attività agricole o di allevamento) o accidentale. Il flusso di migrazione umana e le annesse attività commerciali hanno avuto un grosso impatto a partire dal XVI secolo, con un massiccio trasferimento di specie lungo le nuove rotte di navigazione (Di Castri 1989). L’alterazione della distribuzione naturale delle specie dovuta ad attività antropiche ha un’entità paragonabile all’effetto dei cicli di glaciazione e deglaciazione, nonostante si sia verificata in un arco di tempo significativamente minore (Mack et al. 2000).
Elton (1958) ha definito l’invasione biologica come il trasporto di organismi in nuove regioni – di solito distanti da quella d’origine – i cui discendenti proliferano, si diffondono e persistono. Una specie viene definita invasiva se: (i) colonizza una nuova regione in cui precedentemente non era presente; (ii) l’introduzione è legata, direttamente o indirettamente, all’attività umana; (iii) la regione nativa e il nuovo areale sono separati da una discontinuità biogeografia (dispersione remota) e (iv) le nuove generazioni della specie introdotta si riproducono nel nuovo sito senza assistenza da parte dell’uomo, costituendo così popolazioni che si autosostengono (naturalizzazione della specie) (Boudouresque & Verlaque 2002).
La proporzione di introduzioni volontarie o accidentali varia tra i diversi gruppi tassonomici: pochi microrganismi sono stati introdotti deliberatamente, per lo più lieviti per la fermentazione o mutualisti come micorrize. Le introduzioni volontarie sono rare tra gli insetti e gli invertebrati (questi ultimi introdotti prevalentemente in ambiente marino per l’acquacoltura) mentre rappresentano la maggior parte dei casi di introduzione di piante e vertebrati, per lo più pesci, mammiferi ed uccelli (Mack et al. 2000).
In ambiente acquatico i vettori, ovvero i mezzi attraverso cui le specie invasive vengono trasportate in nuove regioni, sono rappresentati dalla navigazione, dalle attività di pesca ed acquicoltura, dalla ricerca scientifica o dall’attività di singoli acquariofili. Meno numerosi sono i casi di introduzioni volontarie finalizzate al controllo biologico, oppure la connessione tra bacini isolati per eliminazione di barriere biogeografiche (ad esempio mediante costruzione di canali) (Ruiz et al. 2000, Bulleri & Airoldi, in stampa).
Le navi possono trasportare organismi che si sono insediati sullo scafo, nelle acque di zavorra, all’interno o al di sopra del carico, sul ponte o sulle ancore (Ruiz et al. 2000). Il progresso tecnologico dell’ultimo secolo ha portato ad un aumento della superficie degli scafi e del volume delle acque di zavorra rispetto al volume totale della nave, con conseguente aumento del numero di organismi trasportati. L’incremento della velocità di navigazione aumenta inoltre il numero e migliora le condizioni di vitalità dei propaguli nelle acque di zavorra, la cui sopravvivenza dipende spesso dalla durata del trasporto. Al contrario, la sostituzione di vascelli di legno con quelli in ferro e successivamente in acciaio, unitamente all’utilizzo di vernici anti-fouling, alla riduzione dei periodi di residenza in porto (e conseguente riduzione dell’insediamento degli organismi), nonché la maggiore velocità durante il viaggio in mare (che causa il distaccamento di un maggior numero di organismi dallo scafo) ha probabilmente causato il declino delle comunità fouling (organismi sessili). Inoltre si ritiene che alcune componenti del fouling attualmente ad ampia distribuzione (le cosiddette specie cosmopolite) rappresentino introduzioni avvenute in tempi antichi e non riconosciute come tali (Ribera & Boudouresque 1995).
Un assunto comune è che se un vettore ha operato sulla stessa rotta per un lungo periodo di tempo la maggior parte delle specie che possono essere da esso trasportate ed introdotte con successo si siano già insediate. In realtà finché il vettore opera l’invasione può continuare, poiché possono verificarsi modificazioni delle caratteristiche del vettore, dell’ambiente ricevente – per alterazione dell’habitat, eventi di inquinamento ed eutrofizzazione o cambiamenti climatici - nonché delle stesse specie invasive e delle loro interazioni (Carlton et
al. 2003).
I risultati di uno studio effettuato negli Stati Uniti suggeriscono che i principali vettori in ambiente marino siano la pesca e le attività associate alla navigazione (Ruiz et al. 2000). Nel bacino del Mediterraneo, biogeograficamente isolato e caratterizzato da un alto numero di endemismi (25% delle 8500 specie macroscopiche presenti; Occhipinti-Ambrogi & Savini 2003), si ritiene che la principale causa di invasioni sia rappresentata dalla costruzione del Canale di Lesseps (avvenuta nel 1869), che connette il Bacino Levantino con il Mar Rosso (Por 1978). Precedentemente alla realizzazione del canale i Laghi Amari costituivano una barriera al passaggio di specie ma la recente diminuzione della salinità ha reso possibile lo scambio di organismi tra i due bacini. La maggior parte delle specie introdotte attraverso questo canale sono rimaste però confinate al bacino orientale del Mediterraneo, che presenta caratteristiche chimico-fisiche più simili alla regione d’origine (Por 1990). Se si considerano flora e fauna nel loro complesso il numero di specie introdotte nel Mediterraneo è di circa 450
(di cui più di 90 macrofite) (Boudouresque & Verlaque 2002). La percentuale di specie introdotte (4-20% del totale, a seconda del gruppo tassonomico) è maggiore rispetto alle altre regioni del mondo. Questa proporzione si spiega osservando che le specie lessepsiane rappresentano circa la metà del totale e sono quindi la ragione di tale peculiarità (Ribera & Boudouresque 1995).
1.2
Le fasi dell’invasione
L’arrivo di una nuova specie in una nuova località non ne implica necessariamente la naturalizzazione in quanto le invasioni rappresentano il risultato di un processo complesso, costituito da più fasi (Ribera & Boudouresque 1995). La maggior parte degli organismi trasportati accidentalmente periscono durante il tragitto. Raggiunto il nuovo sito le probabilità di sopravvivenza sono basse ed è quindi impossibile quantificare la proporzione di insediamenti falliti; comunque alcune stime di organismi individuati una sola volta lontano dall’areale nativo suggeriscono che l’estinzione locale nelle fasi precoci sia enorme (Carlton & Geller 1993). In alcuni casi la specie è in grado di insediarsi, gli individui sopravvivono e si riproducono originando una nuova popolazione. Il rischio di estinzione per le prime generazioni di discendenti è molto alto e solo in alcuni casi la specie si naturalizza e persiste indipendentemente da eventi di reimmigrazione dal sito d’origine (Ribera & Boudouresque 1995, Di Castri 1989). Si ritiene che un’elevata frequenza di eventi di immissione (invasioni ripetute) aumenti la probabilità di insediamento permanente di una specie (Occhipinti-Ambrogi 1994).
Una volta naturalizzata la specie va incontro ad una fase di espansione ecologica e geografica, con progressione naturale o facilitata dall’azione umana (Ribera & Boudouresque 1995). L’espansione può essere preceduta da una fase di ritardo o “lag phase” di durata variabile – fino ad alcune decine di anni – in cui apparentemente l’invasione ha avuto esito negativo. La durata e le cause di tale fase sono probabilmente il risultato di diversi fattori che possono comprendere (i) i limiti nella capacità di rilevare una popolazione a basse densità; (ii) il processo di selezione naturale che agisce su genotipi rari o recentemente creati per ibridazione per adattarsi alle nuove condizioni ambientali e (iii) le fluttuazioni di fattori abiotici che si sommano ai normali effetti di stocasticità demografica caratteristici di popolazioni di piccole dimensioni (Mack et al. 2000, Crooks & Soule 1999). Dallo studio di numerosi episodi di invasione si deduce che la fase di crescita esponenziale può terminare con il raggiungimento di un plateau di massima densità e distribuzione o con un declino della
specie seguito da una stabilizzazione: il tasso di crescita si riduce e la specie, che ha raggiunto i limiti del nuovo areale, entra in una fase di persistenza (Mack et al. 2000, Ribera & Boudouresque 1995). Le cause di tale declino sono scarsamente note ma tra i possibili fattori vi sono (i) l’adattamento dei predatori indigeni al consumo della nuova specie; (ii) la proliferazione di potenziali predatori o parassiti autoctoni già presenti nella fase di insediamento ma con densità insufficienti per controllarne l’invasione; (iii) la difficoltà di adattamento genetico ai cambiamenti ambientali a causa dei bassi livelli di variabilità genetica e (iv) la depressione dovuta ad inbreeding (Boudouresque 1994). Talvolta può verificarsi un evento di collasso spontaneo per cui la specie invasiva, durante la fase di espansione o di persistenza, raggiunge localmente l’estinzione (Simberloff et al. 2004).
1.3
Gli effetti dell’invasione biologica
Il fenomeno delle invasioni biologiche è in grado di alterare la biodiversità del sistema ricevente a livello genetico, di popolazione, di popolamenti e di ecosistema (Grosholz 2002).
Da un punto di vista genetico possono verificarsi alterazioni indirette dei meccanismi genetici a seguito di una forte riduzione numerica della popolazione, oppure dirette, attraverso ibridazione e immissione di nuovi geni nel pool della specie autoctona (Parker et al. 1999). L’ibridazione altera il flusso genico della specie nativa e, su grandi scale spazio-temporali, provoca perdita di biodiversità a livello genetico (Carroll & Dingle 1996, Crooks 2002). Tale meccanismo può generare una nuova specie o ridurre il successo della specie autoctona per formazione di ibridi sterili che competono con la specie nativa, fino a causarne talvolta l’estinzione (Thompson 1991, Trenham et al. 1998).
La specie alloctona può determinare un declino dell’abbondanza della specie nativa, fino a provocarne l’estinzione. Ciò avviene tramite alterazione di importanti processi ecologici, quali la predazione, il pascolo, la competizione oppure per introduzione di agenti patogeni o parassitari (Mack et al. 2000, Parker et al. 1999).
A livello di popolamenti gli effetti vengono spesso valutati in termini di riduzione della biodiversità (numero di specie), anche se molte invasioni nelle fasi iniziali causano un aumento della ricchezza specifica qualora non vi sia l’eliminazione di specie autoctone (Parker
et al. 1999). L’alterazione della struttura dei popolamenti nativi può avvenire anche attraverso
modificazioni del comportamento di alcune specie residenti nell’utilizzo dell’habitat o delle risorse trofiche (Brown & Moyle 1991).
Il funzionamento di un ecosistema può essere drasticamente alterato, in quanto le specie introdotte possono influenzare la quantità delle risorse disponibili per le specie native ed interferire nei meccanismi di utilizzo (Mack et al. 2000, Vitousek et al. 1987, 1990). Le specie introdotte possono modificare il flusso di energia o di biomassa alterando la struttura della rete trofica ed il tasso di acquisizione delle risorse da parte degli organismi: si ritiene che le specie con maggiore impatto ecologico siano quelle che svolgono una funzione completamente nuova nei popolamenti riceventi (Crooks 2002, Parker et al. 1999). Infine, le specie introdotte possono modificare il regime naturale di disturbo (Crooks 2002, Mack & D’Antonio 1998) o alterare la struttura fisica dell’ecosistema, a varie scale spaziali (Crooks 2002). L’alterazione della biodiversità e dei processi ecologici può avere drastiche ripercussioni a livello economico. Le invasioni biologiche possono portare alla diminuzione della resa economica dello sfruttamento delle risorse naturali, nonché ad elevati costi per la loro lotta, con forme di controllo, eradicazione e quarantena (Mack et al. 2000).
1.4
Le modalità di invasione
Lo studio del fenomeno delle invasioni riveste una notevole importanza in ecologia perché permette la comprensione di processi fondamentali a livello di popolazione, popolamenti ed ecosistema (Ruiz et al. 2000, Crooks 2002). Vari aspetti che regolano il fenomeno sono stati analizzati, primo fra questi l’esistenza di caratteristiche ricorrenti tra le specie invasive. È stato ipotizzato che alcuni attributi della specie possano determinarne la capacità di invadere nuove regioni (Rejmanek & Richardson 1996). Ad esempio, le macrofite invasive presenti nell’area Mediterranea sono per lo più di grandi dimensioni, perenni, mancano di stadi di resistenza, presentano efficiente riproduzione vegetativa, possiedono metaboliti tossici e sono raramente predate da specie erbivore autoctone. L’alga verde Caulerpa racemosa presenta la maggior parte di tali caratteristiche (Boudouresque & Verlaque 2002).
Un altro aspetto importante delle invasioni biologiche riguarda le dinamiche di apporto della specie invasiva. Secondo il modello della disponibilità di propaguli, la probabilità di successo di un’introduzione è fortemente correlata alle dinamiche di apporto dei propaguli nel nuovo ambiente (MacArthur & Wilson 1967, Williamson 1996). Queste sono caratterizzate da: (i) quantità totale dei propaguli; (ii) densità, frequenza e durata dell’inoculo; (iii) esistenza di diverse regioni donatrici e apporto di diversi genotipi e (iv) dalle condizioni dei propaguli al loro arrivo nella regione ricevente (Ruiz et al. 2000).
Anche le caratteristiche dei popolamenti riceventi giocano un ruolo importante nel determinare la resistenza alle invasioni (Chapin III et al. 1998). Elton (1958) osservò che i popolamenti con minor numero di specie sono soggetti a fluttuazioni naturali maggiori e risultano quindi più vulnerabili alle invasioni (McCann 2000). Il modello della ricchezza di specie suggerisce che la resistenza alle invasioni aumenti proporzionalmente al numero di specie presenti nell’ambiente ricevente (Kennedy et al. 2002). Una variante di tale modello descrive la capacità di resistenza dei popolamenti legata non genericamente al numero di specie bensì alla presenza di specie tassonomicamente simili all’invasore, poichè specie alloctone simili a quelle native hanno maggiori probabilità di trovare nel popolamento predatori, parassiti o competitori che ne limitino l’abbondanza (Ricciardi et al. 2004). Tale modello si fonda sulla maggiore probabilità di trovare specie affini all’invasore in popolamenti con maggiore ricchezza di specie. Sulla base della teoria di Elton sono stati sviluppati numerosi modelli allo scopo di identificare i meccanismi attraverso cui la biodiversità può influenzare le modalità di invasione. Di seguito vengono brevemente descritte alcune delle teorie esistenti.
Secondo “il modello delle nicchie vacanti, inutilizzate o sottosfruttate” i popolamenti delle isole e di altri ambienti che contengono un numero ridotto di specie native non possono opporre resistenza biotica alle specie invasive. Tuttavia i potenziali invasori, non trovando tra le poche specie native impollinatori, simbionti o altre specie associate necessarie, non sono in grado di invadere i popolamenti (Simberloff 1995). L’ipotesi della “fuga dalle costrizioni biotiche” suggerisce invece che la specie invasiva sia favorita dalla mancanza degli abituali competitori, predatori, pascolatori e parassiti presenti nel luogo d’origine. Secondo questa ipotesi il successo di una specie invasiva è imputabile quindi non a caratteristiche straordinarie delle specie, bensì al vantaggio tratto dall’assenza di specie virulente o debilitanti associate (Strong
et al. 1984, De Bach & Rosen 1991). È stato inoltre proposto “il modello del disturbo” prima o
durante l’invasione: eventi di disturbo causati dall’uomo o da specie invasive da esso introdotte alterano le condizioni ambientali e possono facilitare l’arrivo di specie invasive, soprattutto se le specie native non sono state in grado di acclimatarsi o adattarsi al nuovo regime di disturbo (Harper 1965). Secondo “il modello dell’invasione progressiva” le invasioni ripetute possono produrre effetti sinergici positivi e favorire ulteriori introduzioni, poichè si istaurano meccanismi di facilitazione tra più specie invasive (Simberloff & Von Holle 1999). Al contrario il “modello della resistenza biotica” descrive come l’introduzione di specie invasive nei popolamenti ne aumenti la resistenza e riduca l’impatto di future invasioni. Ciò si spiega ammettendo che le invasioni che hanno successo introducono specie con caratteristiche
particolari, ad esempio forti competitori, in grado di respingere invasioni future (Moulton & Pimm 1983).
1.5
Studi sul modello di resistenza
Il fenomeno delle introduzioni è stato ampiamente studiato nei popolamenti vegetali terrestri, dove è stato dimostrato che l’alterazione della composizione dei popolamenti può alterare il funzionamento dell’ecosistema (Tilman et al. 1997). Generalmente, a piccole scale spaziali, si evidenzia una relazione positiva tra ricchezza di specie e resistenza all’invasione, che agisce sia sulla probabilità di insediamento (numero di invasori) che sul successo dell’invasore (proporzione di individui di grosse dimensioni) (Kennedy et al. 2002).
Una serie di studi di tipo osservazionale condotti a scale spaziali maggiori suggeriscono invece che popolamenti con maggiori livelli di diversità contengono un maggior numero di specie alloctone, mostrando alta suscettibilità all’invasione (Levine et al. 2004, Lonsdale 1999, Bruno et al. 2004). Questa apparente contraddizione sottolinea l’importanza della scala spaziale a cui viene analizzato il processo: infatti in esperimenti a piccola scala spaziale le variabili ambientali possono essere mantenute costanti manipolando nel contempo il numero di specie native. Su scale spaziali maggiori invece gli studi includono variazioni significative di fattori ambientali associati alla biodiversità quali ad esempio il disturbo, la disponibilità di risorse, la pressione dei propaguli, l’abbondanza di competitori e predatori. Si ritiene che tali fattori covarino con la diversità nei sistemi naturali e che siano in gradi di influenzarne l’invasibilità (Byers et al. 2003, Kennedy et al. 2002).
Per comprendere la relazione tra diversità e resistenza a livello di intero ecosistema è, però, indispensabile estendere lo studio a più livelli trofici ed includere nell’esperimento l’ecosistema nel suo complesso (McCann 2000). Coerentemente a tale affermazione sono stati condotti esperimenti in mesocosmi manipolando simultaneamente produttori, erbivori, carnivori ed organismi decompositori. L’aumento del numero di specie per ognuno dei livelli trofici ha rivelato una correlazione positiva tra diversità e resistenza all’invasione. Questi esperimenti sono stati però effettuati in sistemi semplificati rispetto a quelli naturali (Levine et
al. 2004, Chapin III et al. 1998).
Infine è importante notare che gli studi sperimentali sulle invasioni sono generalmente di breve durata anche se il fenomeno si verifica nell’arco di molti anni: questo può rappresentare un limite alla comprensione delle dinamiche che regolano le invasioni biologiche (Meiners et
1.6
Studi in ambiente marino
Il fenomeno delle invasioni biologiche ha ricevuto una grande attenzione in ambienti terrestri ma non in ambiente marino (Stachowicz et al. 2002) dove, a causa del ridotto riconoscimento storico del fenomeno delle estinzioni, si tende a sottostimare l’impatto delle invasioni (Carlton et al. 1999, Occhipinti-Ambrogi 2001).
Studi osservazionali che confrontano il tasso di espansione geografica di specie introdotte in regioni costiere con dinamiche di invasione in ambienti terrestri mostrano che le invasioni in ambiente marino sono caratterizzate da maggiore variabilità nel tasso di diffusione e frequentemente si verificano crescite demografiche esponenziali e grande dispersione nello spazio in periodi molto brevi (Grosholz 1996, 2002).
Per testare gli effetti della ricchezza di specie dei popolamenti residenti sulla capacità d’invasione sono stati effettuati alcuni esperimenti simili a quelli condotti nei popolamenti di piante terrestri. Questi prevedono l’introduzione di reclute di specie invasive in popolamenti con diverso numero di specie native assemblati dallo sperimentatore (Stachowicz et al. 1999). Si è dimostrato che una grande diversità specifica può ridurre la sopravvivenza e la percentuale di copertura degli invasori poiché popolamenti a maggiore diversità mostrano minore variabilità temporale nello sfruttamento delle risorse, riducendone la disponibilità per le specie invasive (Stachowicz et al. 2002). Studi condotti utilizzando specie native appartenenti allo stesso gruppo trofico e funzionale suggeriscono una particolare resistenza ai fenomeni di invasione nei popolamenti ricchi di specie funzionalmente ridondanti (Stachowicz et al. 1999, Tilman et al. 1997, 2000).
1.7
Caulerpa racemosa
Caulerpa racemosa (Forsskål) J. Agardh è una Chlorophyta ampiamente diffusa in acque
temperate e tropicali. Nel Mediterraneo popola principalmente acque poco profonde ma anche habitat subtidali fino ai 50 m dove colonizza substrati duri e molli (o costituito da matte morta di Posidonia oceanica) in cui sono presenti feltro (un complesso di alghe filamentose), alghe erette e fanerogame marine (Piazzi et al. 2005). C. racemosa mostra una rapida crescita, riproduzione asessuata predominante, alta dispersione e ampia tolleranza all’inquinamento; la monopolizzazione del substrato può alterare profondamente i popolamenti indigeni in vari tipi di habitat (Ceccherelli & Piazzi 2001).
La prima segnalazione nel Mediterraneo è avvenuta nel porto di Sousse in Tunisia (Hamel 1926, Djellouli et al. 1998). Nella prima metà del secolo la specie è rimasta confinata al Bacino Levantino, senza mostrare tendenza invasiva (Verlaque et al. 2000); pertanto si riteneva che C.
racemosa fosse una specie di origine lessepsiana (Por 1978; Verlaque 1994; Ribera &
Boudouresque 1995). All’inizio degli anni ’90 tuttavia si è rapidamente diffusa in tutto il Mediterraneo, colonizzando anche le acque relativamente fredde del bacino occidentale e raggiungendo anche l’Italia (Alongi et al. 1993, Piazzi et al. 1994), la Francia (Verlaque et al. 2000) e le isole Baleari (Verlaque et al., 2000).
Verlaque et al. (2003) hanno suggerito che nel Mediterraneo coesistano tre taxa: (i) C.
racemosa var. turbinata (J. Agardh) Eubank ± var. uvifera (C. Agardh) J. Agardh, nota dal 1926 in
Tunisia (Hamel 1926) e dal 1931 nel bacino Levantino (Hamel 1931); (ii) C. racemosa var.
lamourouxii (Turner) Weber-van Bosse f. requienii (Montagne) Weber-van Bosse, nota dagli
anni ’50 nel bacino Levantino (Huvé 1957); e (iii) C. racemosa (Forsskål) J. Agardh var.
cylindracea (Sonder) Verlaque, Huisman et Boudouresque, la cosiddetta “varietà invasiva” che
si è diffusa nel Mediterraneo a partire dagli anni ’90. La varietà cylindracea mostra differenze morfologiche rispetto alle altre due presenti nel Mediterraneo (Verlaque et al., 2000); inoltre studi genetici confermano che si tratti di un taxon originario dell’Australia sud-occidentale recentemente introdotto nel Mar Mediterraneo (Verlaque et al.. 2003).
Lungo il litorale livornese l’alga ha mostrato una rapidissima espansione tra il 1993 e il 1997 (Piazzi et al. 1997). L’introduzione di C. racemosa nel mar Tirreno è probabilmente attribuibile ad attività umane di tipo ricreativo (acquari) o alla navigazione (Boudouresque & Ribera 1994, Verlaque et al.. 2003). Questi meccanismi potrebbero essere stati favoriti da altri fattori quali le correnti e i cambiamenti climatici, come ipotizzato per altre specie termofile (Astraldi 1995, Modena et al. 2000). La rapida espansione della specie deve essere correlata alla strategia riproduttiva altamente efficiente, associata al trasporto naturale da parte delle correnti o antropico, tramite ancore o reti (Renencourt et al. 2002).
La riproduzione sessuale della varietà invasiva è stata osservata nel Mediterraneo (Panayotidis & Zuljevic, 2001) anche se non sembra essere il meccanismo principale per la diffusione dell’alga (Piazzi et al. 2003). La riproduzione vegetativa avviene attraverso lo sviluppo di stoloni indipendenti (lo stolone si ramifica e dopo alcuni mesi muore nel punto di ramificazione, generando due talli separati) o per dispersione naturale di frammenti dell’alga prodotti dall’azione di forze idrodinamiche o di erbivori marini (ricci o granchi). Un’altra strategia di riproduzione è rappresentata da un processo di frammentazione che si verifica in assenza di disturbo e riguarda una specifica porzione del tallo, i ramuli. I ramuli, inseriti
radialmente sull’asse della fronda (rachis), si separano spontaneamente costituendo propaguli piuttosto che frammenti accidentali. Circa cinque giorni dopo la loro formazione i propaguli globulari, cilindrici o clavati originano filamenti contenenti clorofilla che si sviluppano nella forma del tipico tallo. I propaguli possono essere risucchiati nelle operazioni di carico delle acque di zavorra nei porti il cui fondale è colonizzato da C. racemosa: ciò spiegherebbe la comparsa dell’alga all’imboccatura o in prossimità di porti come Marsiglia, Palma di Majorca, Genova, Livorno, Salerno, Siracusa Cagliari e Famagusta (Renencourt et al. 2002).
Un’altra caratteristica del genere Caulerpa è la produzione di metaboliti tossici. Il principale metabolita secondario prodotto è un sesquiterpenoide citotossico, la caulerpenina, la cui concentrazione varia tra diverse porzioni dell’alga (Meyer & Paul 1992), tra specie, località e stagioni (Jung et al. 2002). Questa sostanza è un importante meccanismo di difesa dagli erbivori e può aumentare le capacità di competizione dell’alga (effetto antifouling) (Lemée et
al. 1993, 1997, Dumay et al. 2002).
C. racemosa è un’alga rizofitica, capace cioè di assumere nutrienti sia dal sedimento che
dalla colonna d’acqua, caratteristica comune alle angiosperme marine. L’assorbimento rizoidale può influenzare l’esito delle interazioni competitive per i nutrienti con le fanerogame marine (Ceccherelli & Cinelli 1997). C. racemosa è stata osservata all’interno delle praterie di
Cymodocea nodosa e di Posidonia oceanica, dove mostra di avere successo quando le piante sono
già in declino (Ceccherelli et al. 2000, Occhipinti-Ambrogi & Savini 2003). Studi effettuati sull’isola di Cipro, dove la specie si trova ad una batimetrica di circa 60 m, suggeriscono che
C. racemosa sia responsabile della modificazione del popolamento della macrofauna in
abbondanza e composizione. Prima dell’invasione di C. racemosa il taxa dominante era rappresentato dai gasteropodi (44%) mentre dopo l’espansione dell’alga risultano essere i policheti (38%) (Argyrou et al. 1999).
Gli studi sperimentali condotti su fondale roccioso sottolineano la grande capacità invasiva e competitiva dell’alga. Dopo che gli stoloni hanno occupato il substrato disponibile, a scapito delle specie native, la crescita verticale del tallo continua creando una struttura multistratificata (Piazzi et al. 2001). Il conseguente intrappolamento del sedimento danneggia le specie native, causando impoverimento dei popolamenti, ovvero una drastica riduzione delle specie macroalgali ed alterazione delle abbondanze relative (Piazzi et al. 2001).
C. racemosa segue le stesse fluttuazioni temporali descritte per altre specie di Caulerpa nel
Mediterraneo: essa ha un periodo di crescita vegetativa tra giugno e novembre alternato a riposo vegetativo da dicembre a maggio. Durante il riposo vegetativo diminuiscono la copertura e la biomassa ma alcuni stoloni restano sul fondo e, all’inizio dell’estate successiva,
permettono la rapida invasione del substrato. Il periodo di riposo vegetativo non permette ai popolamenti macroalgali di restaurare la struttura originaria; le alterazioni persistono nel popolamento e aumentano nel corso della stagione successiva (Piazzi et al. 2001).
Un esperimento di rimozione di C. racemosa ha suggerito che la variazione temporale del disturbo può essere di estrema importanza nel controllare la diffusione della specie. L’effetto del disturbo sulle dinamiche di colonizzazione di C. racemosa è risultato dipendente dal periodo dell’anno in cui si ha la disponibilità di spazio: le aree disturbate durante l’inverno e la primavera non vengono ripopolate fino all’inizio dell’estate successiva. Durante la stagione calda la capacità di occupare lo spazio disponibile per propagazione laterale degli stoloni permette all’alga di ripopolare rapidamente le aree disturbate (Piazzi et al. 2003).
Un altro studio sperimentale, condotto manipolando le componenti del popolamento macroalgale (alghe incrostanti, alghe che formano un feltro ed alghe erette con tallo laminare o carnoso) e trapiantando C. racemosa, ne ha valutato le modalità di espansione (Ceccherelli et
al. 2002). I risultati dell’esperimento indicano che la diffusione dell’alga dipende dal tipo di
habitat in cui essa si insedia e non dalla sua complessità: il feltro risulta favorirne l’insediamento più delle sole alghe incrostanti o dell’eliminazione dell’intero popolamento macroalgale. Il disegno sperimentale utilizzato non permette tuttavia di distinguere l’effetto della densità del popolamenti algale da quello della ricchezza di specie.
1.8
Scopo dell’esperimento
Questo studio si basa sull’approccio ipotetico deduttivo, secondo il quale viene elaborato un modello esplicativo del fenomeno osservato e successivamente si procede al vaglio sperimentale delle ipotesi da esso derivate (Underwood 1990).
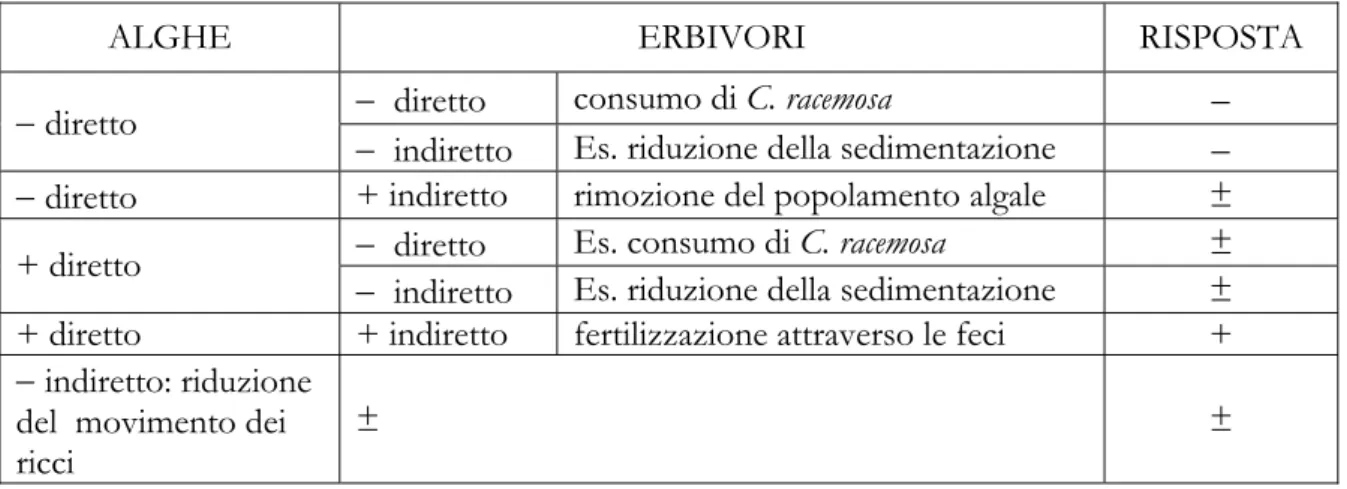
L’esperimento ha permesso di analizzare le modalità di invasione di C. racemosa in relazione alle caratteristiche dei popolamenti naturali. In particolare è stato testato il modello secondo cui popolamenti caratterizzati da una maggiore biodiversità offrono maggiore resistenza all’invasione da parte di specie esotiche. È stata quindi testata l’ipotesi secondo la quale la capacità invasiva di C. racemosa sia ridotta da una maggiore diversità dei popolamenti di produttori primari (alghe) e degli erbivori (echinoidei). Previsioni relative alle possibili risposte di C. racemosa alla manipolazione della biodiversità possono essere derivate da modelli basati su reti di interazioni tra i due livelli trofici che includono possibili effetti diretti ed indiretti, positivi e negativi (Tab. 1.1). La diversità dei popolamenti algali può avere effetti
negativi diretti su C. racemosa. Questi effetti possono essere esacerbati dalla presenza degli erbivori qualora questi abbiano effetti negativi diretti (consumo di C. racemosa) o indiretti (es. attraverso la diminuzione della sedimentazione, assumendo che il sedimento abbia effetti positivi su C. racemosa). Al contrario gli erbivori possono controbilanciare gli effetti negativi diretti del popolamento algale su C. racemosa, se la loro azione di pascolo si esercita principalmente sul popolamento algale anziché su C. racemosa.
Gli effetti dei popolamenti algali sull’invasione da parte di C. racemosa possono, alternativamente, essere positivi (ad esempio attraverso l’intrappolamento di sedimento o permettendo un migliore ancoraggio degli stoloni). Gli effetti positivi dei popolamenti algali su C. racemosa potrebbero essere esacerbati da effetti positivi indiretti degli erbivori (fertilizzazione attraverso le feci). Alternativamente, gli erbivori potrebbero controbilanciare tali effetti sia direttamente, attraverso il consumo di C. racemosa, che indirettamente, attraverso il consumo dei popolamenti algali. Infine, i popolamenti algali potrebbero influenzare indirettamente C. racemosa agendo sulle modalità di movimento dei ricci qualora questi avessero un effetto positivo su C. racemosa.
Gli studi sulle invasioni biotiche condotti in ambiente marino risultano scarsi in letteratura, inoltre sono poco frequenti gli esperimenti che analizzano il fenomeno a più livelli trofici. Generalmente l’effetto della densità delle specie residenti sulle modalità di invasione non viene distinto dall’effetto della ricchezza di specie (Benedetti-Cecchi 2004). Ciò avviene perchè lo sperimentatore crea un gradiente di diversità assemblando popolamenti con crescente numero di specie autoctone all’interno di unità sperimentali di pari dimensioni, con conseguente riduzione proporzionale della densità delle stesse. Anche gli esperimenti che manipolano il numero di specie per rimozione dei popolamenti mancano di tale distinzione. Nel nostro caso, invece, diversi gruppi morfologici sono stati manipolati in maniera da permettere l’analisi distinta dei due fattori. Anche la manipolazione delle specie di erbivori secondo un disegno fattoriale permette di rilevare distintamente l’effetto della densità a cui essi sono presenti dall’effetto “specie”.
Sono stati presi in esame gli effetti dei seguenti fattori sull’espansione di C. racemosa:
1- L’effetto del pascolo delle specie degli echinoidei Paracentrotus lividus e Arbacia lixula, separatamente e in combinazione.
2- L’effetto della pressione di pascolo (determinata da una diversa densità di erbivori) da parte di Paracentrotus lividus e Arbacia lixula.
3- L’effetto della diversità dei popolamenti algali naturali costituiti da feltro ed alghe incrostanti, separatamente e in combinazione.
4- L’effetto della densità dei popolamenti algali.
È da notare che questo esperimento, pur permettendo di distinguere tra effetti diretti ed indiretti tra i due livelli trofici, non permette di identificare i meccanismi alla base del secondo tipo di effetti. Pertanto i meccanismi indiretti indicati in Tab. 1.1 sono solo a titolo di esempio
e non escludono la possibilità che altri meccanismi possano generare effetti indiretti.
Tab. 1.1 Risposta di C. racemosa prevista rispetto agli effetti diretti ed indiretti degli erbivori e del popolamento
algale. ± indica una risposta alternativamente negativa o positiva a seconda del bilancio degli effetti ai due livelli trofici. Gli effetti indiretti assumono che C. racemosa sia influenzata positivamente dalla sedimentazione, come suggerito da altri studi (es. Ceccherelli & Cinelli 1997).
ALGHE ERBIVORI RISPOSTA
− diretto consumo di C. racemosa −
− diretto
− indiretto Es. riduzione della sedimentazione −
− diretto + indiretto rimozione del popolamento algale ±
− diretto Es. consumo di C. racemosa ±
+ diretto
− indiretto Es. riduzione della sedimentazione ±
+ diretto + indiretto fertilizzazione attraverso le feci +
− indiretto: riduzione del movimento dei ricci