notocorda e dei somiti e la soprastante piastra neurale posteriore si restringono (convergono) lungo la direzione mediolaterale (circumblastoporale), e si allungano (si estendono) in direzione antero-posteriore (frecce bianche, Fig. 5G-H, K-L). Questi movimenti accoppiati, spesso chiamati estensione
convergente, allungano l’asse corporeo e contribuiscono all’involuzione delle
cellule e alla chiusura del blastoporo (Keller, 1986; Keller et al., 2000). Durante tutti gli stadi di blastula e gastrula, la regione animale dell’embrione si espande per epibolia, estendendosi in direzione vegetativa nella regione lasciata vuota dalla IMZ quando quest’ultima si è internalizzata. L’epibolia si ha per assottigliamento e allargamento della porzione animale (Fig. 5A-C).
1.3.a Convergenza ed Estensione
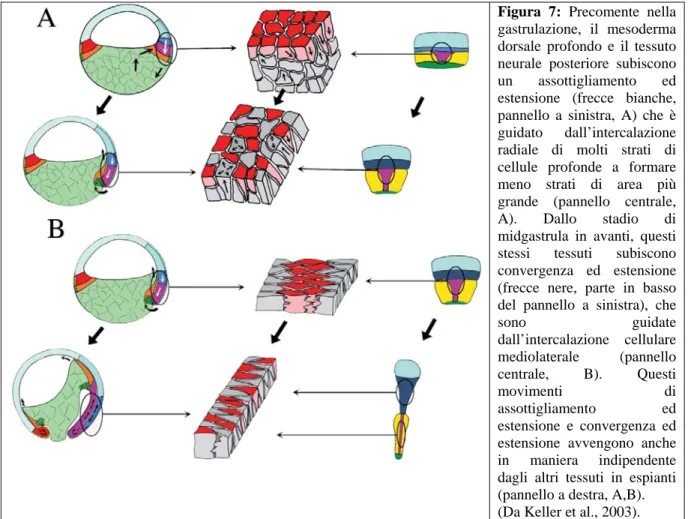
Durante la gastrulazione e la neurulazione, il mesoderma presuntivo della notocorda e dei somiti e la sovrastante piastra neurale posteriore, convergono (si restringono) nella loro direzione mediolaterale (circumblastoporale) e si estendono (si allungano) lungo la loro dimensione antero-posteriore (Keller, 1975; 1976). Questi movimenti allungano l’asse antero-posteriore, guidano l’involuzione, portano a una chiusura asimmetrica il blastoporo e risultano nella formazione di un archenteron allungato (Keller, 1986 e Keller et al., 2000; frecce bianche, pannelli a sinistra, Fig. 7A, B).
Figura 7: Precomente nella
gastrulazione, il mesoderma dorsale profondo e il tessuto neurale posteriore subiscono un assottigliamento ed estensione (frecce bianche, pannello a sinistra, A) che è guidato dall’intercalazione radiale di molti strati di cellule profonde a formare meno strati di area più grande (pannello centrale, A). Dallo stadio di midgastrula in avanti, questi stessi tessuti subiscono convergenza ed estensione (frecce nere, parte in basso del pannello a sinistra), che
sono guidate dall’intercalazione cellulare mediolaterale (pannello centrale, B). Questi movimenti di assottigliamento ed estensione e convergenza ed
estensione avvengono anche in maniera indipendente dagli altri tessuti in espianti (pannello a destra, A,B). (Da Keller et al., 2003).
Questi movimenti sono attivi, generatori di forze e creati localmente, come mostrato dal fatto che espianti di tessuti mesodermici (Holtfreter, 1944; Keller e Danilchik, 1988) e neurali (Keller e Danilchik, 1988; Keller et al., 1992c; Elul e Keller, 2000) convergono e si estendono in coltura anche se non attaccati a substrati esterni o ad altre parti dell’embrione (pannelli a destra, Fig. 7A, B). Tecniche di “cell tracing” (Keller e Tibbets, 1989) e di “timelapse recording” (videoregistrazione nel tempo) (Keller et al., 1985; Wilson et al., 1989; Wilson e Keller, 1991) o di cellule marcate con la fluorescenza (Keller et al., 1989; Shih e Keller, 1992a, b) hanno mostrato che la convergenza e l’estensione coinvolgono due tipi di intercalazione cellulare. Per prima cosa molti strati di cellule profonde si intercalano lungo il raggio dell’embrione (intercalazione radiale) per produrre un numero minore di strati
(assottigliamento) di maggiore lunghezza (estensione; pannello centrale, Fig. 7A) e successivamente le cellule profonde si intercalano mediolateralmente (intercalazione mediolaterale) per produrre una fila più stretta (convergenza) e più lunga (estensione) (pannello centrale, Fig. 7B). L’intercalazione radiale predomina nella prima metà della gastrulazione (Fig. 7A); l’intercalazione mediolaterale predomina nella seconda metà della gastrulazione e nella neurulazione (Fig. 6B), sia nel tessuto mesodermico dorsale che nel tessuto neurale posteriore presuntivo (midollo spinale e rombencefalo).
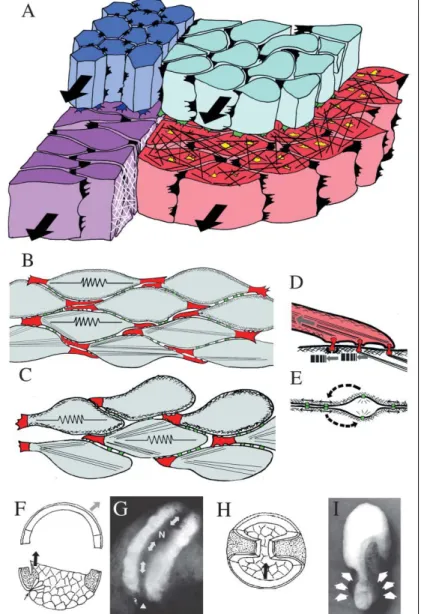
Figura 8: Il diagramma illustra il
comportamento delle cellule durante la convergenza ed estensione (A): le cellule neurali (blu chiaro) mostrano attività protrusiva monopolare in direzione mediale; le cellule del mesoderma somitico (rosso) e le cellule della notocorda (magenta) mostrano attività protrusiva bipolare in direzione mediolaterale; le cellule della piastra neurale (blu scuro) mostrano attività protrusiva multipolare. Si pensa che queste attività protrusive siano la causa dell’estensione convergente e dell’allungamento dei tessuti (frecce nere). L’intercalazione cellulare è guidata dalle protrusioni bipolari nel caso del mesoderma (B) o da quelle monopolari nel caso del neurectoderma (C). Le protrusioni sono agganciate saldamente alle cellule adiacenti per impedire lo scivolamento (D). Tra le cellule sono inoltre presenti degli agganci molto deboli ma molto numerosi che si rompono e riformano continuamente (E). Se si rimuove il tetto del blastocele (F), l’embrione chiude comunque il blastoporo grazie alla convergenza ed estensione (G). Se si fornisce un substrato fuori asse (H), il substrato viene tirato sopra all’asse in estensione, ma non fa divergere l’asse di estensione (I). Diagrammi e foto da Keller et al., 2003.
Durante l’intercalazione cellulare mediolaterale del mesoderma, l’attività protrusiva delle cellule diventa polarizzata, con grandi protrusioni lamelliformi alle estremità mediali e laterali delle cellule e piccole protrusioni filiformi sulle loro superfici anteriore e posteriore (cellule rosse, Fig. 8A; Fig. 8B). Le protrusioni mediali e laterali sembrano esercitare una trazione sulle cellule adiacenti e generare una tensione sull’asse mediolaterale. Le cellule diventano allungate mediolateralmente, orientate parallelamente l’una all’altra e si muovono tra loro lungo l’asse mediolaterale, formando una fila più lunga e più stretta (cellule rosse, Fig. 8B; Keller et al., 1992a). L’intercalazione mediolaterale avviene nel tessuto neurale posteriore in maniera simile, con una grande differenza. Invece di essere bipolari, le cellule neurali hanno la loro attività protrusiva fortemente polarizzata verso le estremità mediali (Elul e Keller, 2000; cellule blu, Fig. 8A,C). Questa attività protrusiva “monopolare” diretta medialmente è dipendente dalla presenza di una notocorda/notopiastra nella linea mediana (Elul et al., 1997 e Elul e Keller, 2000) e si pensa che guidi l’intercalazione cellulare tramite lo stesso meccanismo di trazione cellula-cellula del modo bipolare (Fig. 8C).
In Drosophila la polarità delle cellule è regolata da due vie: la via della Polarità Cellulare Planare (PCP), o quella della polarità tissutale planare (Adler, 2002). Queste vie di segnale sono composte da numerose molecole regolatrici i cui omologhi sono implicati nella convergenza ed estensione in
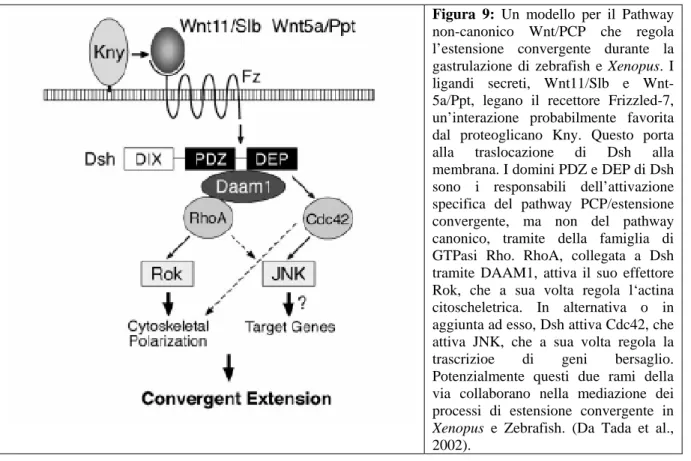
Figura 9: Un modello per il Pathway
non-canonico Wnt/PCP che regola l’estensione convergente durante la gastrulazione di zebrafish e Xenopus. I ligandi secreti, Wnt11/Slb e Wnt-5a/Ppt, legano il recettore Frizzled-7, un’interazione probabilmente favorita dal proteoglicano Kny. Questo porta alla traslocazione di Dsh alla membrana. I domini PDZ e DEP di Dsh sono i responsabili dell’attivazione specifica del pathway PCP/estensione convergente, ma non del pathway canonico, tramite della famiglia di GTPasi Rho. RhoA, collegata a Dsh tramite DAAM1, attiva il suo effettore Rok, che a sua volta regola l‘actina citoscheletrica. In alternativa o in aggiunta ad esso, Dsh attiva Cdc42, che attiva JNK, che a sua volta regola la trascrizioe di geni bersaglio. Potenzialmente questi due rami della via collaborano nella mediazione dei processi di estensione convergente in Xenopus e Zebrafish. (Da Tada et al., 2002).
Questi includono la molecola secreta Wnt11 (Tada e Smith, 2000) e il recettore transmembrana di Wnt, Frizzled (Djiane et al., 2000). La via di segnale Wnt/Frizzled sembra regolare la convergenza ed estensione tramite la regolazione della famiglia di GTPasi Rho che controllano l’organizzazione citoscheletrica, la polarità cellulare e l’attività protrusiva (Settleman, 2001). Frizzled, tramite Dishevelled (Sokol, 1996; Wallingford et al., 2000; Wallingford e Harland, 2001; Habas et al., 2001) interagisce a valle con DAAM1 (Dishevelled Associated Activator of Morphogenesis) che a sua volta regola RhoA (Habas et al., 2001) e la Rho chinasi (Marlow et al., 2002; Fig. 9). Wnt11 agisce anche (sempre tramite Frizzled e Dishevelled) attivando Rac, in maniera indipendente dall’attivazione di Rho, in un contesto necessario per la convergenza ed estensione (Habas et al., 2003). La via di segnale
Wnt11/Frizzled regola anche l’attività di Cdc42 tramite l’attivazione di proteine G, secondo una via di segnale detta via Wnt/Ca. Cdc42 sembra essere importante sia nella convergenza e estensione (Choi e Han, 2002) che nella separazione dei tessuti (Winklbauer et al., 2001). Un’altro componente della via è una proteina a 4 passi transmembrana, Strabismus (Darken et al., 2002; Goto e Keller, 2002; Park e Moon, 2002). E’ stato mostrato che questa via regola l’attività protrusiva (Wallingford et al., 2000), ma non è noto se questa via agisca durante l’intercalazione radiale con i suoi effetti a cascata attraverso il successivo processo di intercalazione mediolaterale, o se agisca direttamente sull’intercalazione mediolaterale.
È stato dimostrato (Andreazzoli et al., 1997; Morgan et al., 1999a; 1999b) che Xotx2 è in grado di bloccare i movimenti di estensione convergente. Esperimenti di sovraespressione hanno infatti mostrato che sia
Xotx1 che Xotx2 sono in grado di indurre forti riduzioni nelle strutture
posteriori. I movimenti di estensione convergente possono essere studiati in esogastrule, dove sono direttamente rilevabili in quanto avvengono senza l’involuzione della zona marginale (Keller, 1991). A differenza degli embrioni di controllo, la maggior parte delle esogastrule ottenute da embrioni iniettati con gli mRNA di Xotx1 o Xotx2 sono più corte e, se si osserva la zona marginale, essa appare essere scarsamente allungata (Andreazzoli et al., 1997). Questo è stato dapprima spiegato come un’azione di repressione sulla formazione di strutture posteriori da parte di questi due geni “anteriorizzanti”. Successivamente, Morgan et al., (1999b) hanno dimostrato che è dovuto ad un effetto diretto di Xotx2 sull’attivazione di XclpH3. Il prodotto di questo gene ha un’identità amminoacidica dell’85% con la calponina H2 di topo, un proteina che lega l’actina che può inibire l’attività Mg-ATPasica della miosina e di conseguenza impedire lo scorrimento dei filamenti di actina sul substrato di miosina (Shirinsky et al., 1992). Sebbene molto simile alla calponina H2 per
gran parte della sua lunghezza, i 21 amminoacidi carbossi-terminali di XclpH3 sono altamente divergenti dalle isoforme H2 di altre specie; per questo la proteina è stata denominata Xenopus calponin H3 (XclpH3). Il fenotipo ottenuto sovraesprimendo XclpH3 è molto simile alla sovraespressione di
Xotx2, con forti riduzioni posteriori e spine bifide molto numerose, ma senza
induzione di ghiandole ectopiche del cemento. Inoltre anche i “patterns” d’espressione sono comparabili, dal momento che anche XclpH3 è materno e che si restringe durante lo sviluppo alle regioni anteriori dell’embrione. L’espressione di entrambi i geni è localizzata in tutte e tre gli strati (ectoderma, mesoderma e endoderma dorsale), ad eccezione dell’endoderma ventrale, che formerà il pavimento dell’archenteron, che invece non li esprime. Allo stadio di bottone caudale l’espressione di entrambi è ristretta agli occhi e alla regione del prosencefalo. La sovraespressione di Xotx2 in espianti induce rapidamente l’espressione di XclpH3 e questa induzione è insensibile all’inibizione della sintesi proteica con cicloesimmide (Morgan et al., 1999b) indicando che
XclpH3 è un bersaglio diretto di Xotx2. L’iniezione di un antisenso di XclpH3
inoltre, è in grado di recuperare gli effetti dell’espressione ectopica di Xotx2 (Morgan et al., 1999b).
Tutto ciò suggerisce quindi che Xotx2 possa inibire l’estensione convergente attivando il gene XclpH3.