INTRODUZIONE --- 3
1.1 ASPETTI POSITIVI NELL'USO DI MACROTERME NELLE ZONE DI TRANSIZIONE --- 6
1.2 CRITICITÀ NELL'USO DELLE MACROTERME--- 9
1.3 CYNODON SPP. --- 11
1.4 ZOYSIA SPP. --- 15
1.5 RESISTENZA AL FREDDO DI CYNODON E ZOYSIA--- 17
1.6 CONGELAMENTO E STRESS CAUSATO DALLE BASSE TEMPERATURE --- 21
1.7 METABOLISMO E TOLLERANZA AL FREDDO--- 25
1.8 TECNICHE AGRONOMICHE PER AUMENTARE LA RESISTENZA AL FREDDO --- 28
1.8.1 IL RUOLO DEI FERTILIZZANTI --- 29
SCOPO DELLA TESI --- 33
MATERIALI E METODI--- 35
3.1 INSEDIAMENTO DELLE CULTIVARS--- 35
3.2 TRATTAMENTI --- 36
3.3 CAMPIONAMENTO--- 37
3.4 RECOVERY--- 38
3.5 ANALISI DEGLI ZUCCHERI SOLUBILI --- 39
3.5.1 ESTRAZIONE DEI METABOLITI --- 39
3.5.2 PROCEDURA PER LA DETERMINAZIONE ENZIMATICA DEGLI ZUCCHERI SOLUBILI--- 40
3.6 ANALISI DELL'AMIDO--- 43
3.6.1 ESTRAZIONE DELL'AMIDO --- 43
3.6.2 DETERMINAZIONE DELL'AMIDO CON IODIO--- 44
3.7 ANALISI DEI PIGMENTI FOGLIARI--- 45
3.7.1 ESTRAZIONE DEI PIGMENTI--- 45
3.7.2 QUANTIFICAZIONE DEI PIGMENTI--- 45
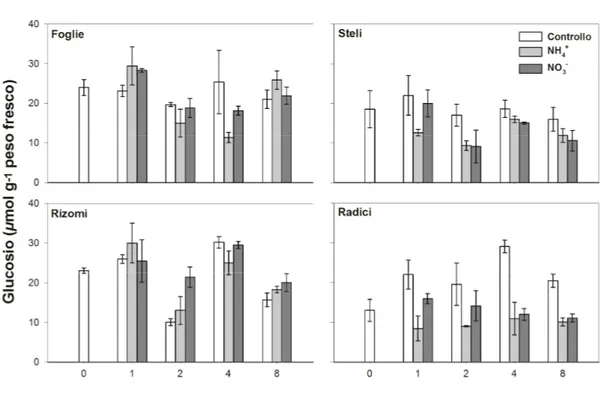
RISULTATI E DISCUSSIONE--- 47 4.1 TOLLERANZA AL FREDDO (LT50) --- 47 4.1.1 CYNODON--- 47 4.1.2 ZOYSIA--- 49 4.2 ZUCCHERI LIBERI --- 51 4.2.1 CYNODON--- 51 • SACCAROSIO --- 51 Foglie --- 52 Steli --- 52 Rizomi --- 53 Radici --- 53 • FRUTTOSIO--- 54 Foglie --- 55 Steli --- 55 Rizomi --- 56 Radici --- 56 • GLUCOSIO--- 57 Foglie --- 58
Introduzione
Steli --- 59
Rizomi --- 59
Radici --- 60
• ZUCCHERI LIBERI TOTALI--- 61
4.2.2 ZOYSIA--- 63 • SACCAROSIO --- 63 Foglie --- 64 Steli --- 64 Rizomi --- 65 Radici --- 65 • FRUTTOSIO--- 66 Foglie --- 67 Steli --- 67 Rizomi --- 68 Radici --- 68 • GLUCOSIO--- 69 Foglie --- 70 Steli --- 70 Rizomi --- 71 Radici --- 71
• ZUCCHERI LIBERI TOTALI--- 72
4.3 AMIDO --- 74 4.3.1 CYNODON--- 74 4.3.2 ZOYSIA--- 75 4.4 PIGMENTI FOGLIARI--- 77 4.4.1 CYNODON--- 77 4.4.2 ZOYSIA--- 80 CONCLUSIONI --- 83 BIBLIOGRAFIA --- 86
Introduzione
Capitolo 1
Introduzione
Ogni specie esprime al meglio il suo potenziale biologico e genetico quando è posta nel suo naturale areale di distribuzione. Infatti, quando è utilizzata in una zona in cui le caratteristiche pedo-climatiche sono differenti da quelle ottimali è più facilmente soggetta a danni da stress biotici e abiotici, richiedendo inoltre maggiori cure e inputs esterni
(Beard, 1973).
Le specie da tappeto erboso si possono distinguere in due grandi categorie che determinano la loro distribuzione nel pianeta: macroterme (C4 - Hatch-Slack) e microterme (C3). La
distinzione è fatta sulla base del diverso metabolismo fotosintetico, adattamento a diversi optimum di temperatura. La fotosintesi C3 viene così definita perché il primo prodotto
ottenuto dalla fissazione della CO2 è formato da tre atomi di
carbonio (acido trifosfoglicerico). Le macroterme hanno una fotosintesi più efficiente ad alte temperature in quanto hanno scarsa o nulla fotorespirazione (punto di compensazione vicino allo zero).
La CO2 che entra nel ciclo di Calvin è ottenuta
decarbossilando un composto a 4 atomi di carbonio (acido ossalacetico, OAA), da qui il nome C4. Le due caratteristiche
fondamentali che differenziano le specie con il ciclo Hatch-Slack da quelle C3 sono l'anatomia Kranz (a corona) e le due
Introduzione
carbossilazioni successive. La prima carbossilazione avviene nelle cellule del mesofillo fogliare ad opera dell'enzima PEP carbossilasi, la seconda nelle cellule a corona della guaina del fascio (provviste di cloroplasti) ad opera della Rubisco.
Nelle cellule del mesofillo avviene la formazione di un acido a quattro atomi di carbonio (OAA), trasportato e decarbossilato nelle cellule della guaina del fascio. Con la decarbossilazione si ottiene un composto a tre atomi di carbonio e CO2. Il composto a tre atomi di carbonio ottenuto
torna nella prima cellula per iniziare nuovamente il ciclo mentre la CO2 entra nel ciclo di Calvin. Grazie a questo nelle
cellule del mesofillo (impermeabili ai gas) la concentrazione di CO2 si mantiene superiore a quella dell'atmosfera.
In questo tipo di ciclo non c'è possibilità di fotorespirazione, in quanto si annulla la possibilità che la Rubisco, trovandosi ad alte concentrazioni di CO2, possa
reagire con l'ossigeno. C'è inoltre una divisione di compiti fra le cellule del fascio e quelle del mesofillo che impedisce il trasporto elettronico e quindi la formazione di O2 nelle
cellule della guaina del fascio. D'altra parte, dato che nelle cellule del mesofillo c'è assenza di Rubisco e di enzimi del ciclo di Calvin, questo impedisce ulteriormente la fotorespirazione. La fotorespirazione è favorita nelle microterme dalle alte temperature perché queste diminuiscono la specificità della Rubisco per la CO2 e diminuiscono
maggiormente la solubilità della CO2 rispetto quella dell'O2.
La produttività del ciclo Hatch-Slack è maggiore di 25-30 μg CO2 m-2s-1 rispetto a quello C3 15-25 μg CO2 m-2s-1. La
fotosintesi C4 è quindi un adattamento alle alte temperature
in quanto fa risparmiare acqua nella fase di fissazione del carbonio, gli stomi stanno quindi più chiusi e risulta più
Introduzione
efficiente ad alte temperature. Oltre ad un miglior uso dell'acqua queste piante hanno un minor bisogno di azoto grazie alla maggior efficienza della Rubisco.
Le specie microterme hanno come range di temperature ideali per la fotosintesi (massimo tasso di fotosintesi per unità di area fogliare) circa 15-24°C, mentre le macroterme 27-35°C.
I due areali non sono nettamente distinti, ma è presente una fascia detta zona di transizione dove è possibile utilizzare sia macroterme che microterme, ma entrambe ai limiti dei loro range climatici.
Tabella 1.1 Range ottimali di temperatura delle specie macroterme e microterme per l'apparato epigeo e ipogeo.
Le temperature massime e minime sono le prime ad indirizzare nella scelta della specie adatta.
L'Italia si definisce una zona di transizione in quanto le specie microterme e macroterme incontrano i limiti di crescita rispettivamente in estate e in inverno. In entrambi i casi si potrebbe avere un disseccamento della parte aerea.
Apparato epigeo Apparato ipogeo
Macroterme 27-35°C 24-32°C
Introduzione
1.1 Aspetti positivi nell'uso di Macroterme
nelle zone di transizione
• Consumi idrici
Le microterme hanno valori di evapotraspirazione potenziale (ETP) molto più alti delle macroterme (Casnoff,
1989).
In caso di forte richiamo dell'acqua dalla pianta all'atmosfera la foglia chiude gli stomi per ovviare alla perdita d'acqua. Le microterme soffrono di più in queste condizioni in quanto il ciclo C3 perde efficienza a basse
concentrazioni di CO2. La richiesta d'acqua delle microterme
per produrre un grammo di sostanza secca è circa tre volte maggiore di quella richiesta dalle macroterme (Hull, 1996).
Molto bassa < 6 Buchloe dactyloides
Bassa 6-7 Cynodon spp.
Eremochloa ophiuroides
Zoysia spp.
Media 7-8.5 Festuche fini
Paspalum spp.
Alta 8.5-10 Lolium perenne
Molto alta >10 Festuca
arundinacea Agrostis stolonifera
Introduzione
Lolium multiflorum
Tabella 1.2 Valori di evapotraspirazione potenziale delle principali specie da tappeto erboso (Panella et al., 2000).
ETP mm giorno-1 SPECIE
L'ETP di una specie influenza direttamente la sua resistenza allo stress idrico. Le macroterme resistono meglio a questo tipo di stress e manifestano segnali di sofferenza (perdite di colore) solo se questo è prolungato nel tempo
(Turgeon, 1980).
Oltre al tipo di metabolismo c'è un altro fattore che influenza la resistenza alla siccità ed è quello della lunghezza dell'apparato radicale. Le macroterme hanno apparati radicali più profondi ed estesi e anche per questo aspetto sono adatte a resistere a periodi di siccità.
Le radici delle macroterme possono spingersi anche ben oltre un metro di profondità, mentre quelle delle microterme in genere si concentrano nei primi 30-40 cm, anche se le radici singole possono superare questa profondità (Christians, 1998). L’apparato radicale, l’angolo di inserzione fogliare, la presenza di peli sulla lamina fogliare e la cuticola inspessita favoriscono le specie macroterme in situazioni di prolungata carenza idrica associate ad alte temperature, riducendo le perdite di acqua e permettendo il recupero della poca acqua disponibile. Inoltre permettono di avere consumi idrici inferiori fino al 45% rispetto a quelli necessari alla sopravvivenza alle microterme (Biran et al., 1981).
Introduzione
Questo aspetto è da tenere in forte considerazione in quanto il risparmio delle risorse idriche nella situazione attuale di carenza idrica che investe il pianeta è di estrema importanza.
• Resistenza alle avversità
Viene definita avversità una qualsiasi modificazione, causata da un organismo vivente o dall’ambiente, che interferisce con il metabolismo della pianta causandone un’alterazione a livello esterno o interno (Beard, 1973).
Come avversità possiamo identificare sia agenti biotici che abiotici. La resistenza cambia molto da specie a specie, ma in generale sappiamo che le macroterme risultano più resistenti agli attacchi fungini e meno a quelli provocati da insetti (Pannella et al., 2000)
• Esigenze nutrizionali
Con apporto esterno si forniscono gli elementi principali della nutrizione N,P,K. Con modalità diverse in relazione alle diverse esigenze delle specie e alla funzione del tappeto erboso. Proprio in base alla funzione definiamo uno standard di qualità in base al quale viene impostato il piano di concimazione. L'elemento principalmente fornito è l'azoto. Le macroterme in genere hanno esigenze inferiori di azoto rispetto alle microterme.
Introduzione
• Resistenza al calpestamento
Le specie macroterme offrono in generale una buona resistenza all'usura, anche se a volte associata a un lento tasso di crescita come nel caso del genere Zoysia. Questo è dovuto alla durezza delle foglie ma, principalmente, alla presenza di lignina negli stoloni e nei rizomi (Lulli et al.,
2011).
Per tutti questi motivi risulta interessante studiare come ampliare l'areale di utilizzo delle macroterme e utilizzarle più diffusamente anche nelle zone di transizione cercando di limitare al massimo i danni invernali. In generale risulta vantaggioso espandere l'areale di specie macroterme da tappeto erboso, per avere tappeti erbosi più sostenibili, a bassi inputs e ecocompatibili.
1.2 Criticità nell'uso delle Macroterme
• Formazione di feltro
Tendenzialmente le macroterme, in particolare Cynodon e Zoysia sono più inclini alla formazione del feltro, uno strato organico compreso tra il suolo e i tessuti epigei del tappeto costituito sia da sostanza organica, sia da parti vive come corone, radici e germogli (Hurto e Turgeon, 1978). La formazione del feltro può essere amplificata da tutte quelle pratiche che riducono l'attività dei microrganismi nel terreno. Questo è comune nei tappeti erbosi sportivi ad alta manutenzione.
Introduzione
L'eccessiva presenza di feltro impedisce alle radici di approfondirsi nel terreno, causa uno strato idrofobico rendendo difficile l'infiltrazione dell'acqua. Il feltro viene controllato tramite pratiche agronomiche: correzione del pH in caso sia il pH a impedire la demolizione della sostanza organica, riduzione degli apporti di azoto e drenaggio. Viene inoltre rimosso con metodi meccanici come la carotatura o il
verticutting. La prima operazione viene effettuata con
macchine carotatrici che asportano carote di terreno, la seconda con macchine provviste di lame che effettuano tagli verticali. Si tratta di azioni che favoriscono l'ossigenazione del terreno. A seguito viene distribuito un sottile strato di sabbia (fino a un massimo di 5-6mm) per diluire il feltro, questa pratica viene chiamata topdressing.
• Stress termico
Il maggior limite nell'uso delle macroterme è costituito dall'andamento climatico del luogo in cui intendiamo insediarle. La latitudine è il principale fattore che influenza l'andamento delle temperature. Le temperature minime, in particolare, sono quelle che determinano la diffusione delle macroterme.
Le basse temperature influiscono negativamente sul ciclo vitale delle macroterme a seconda di quanto a Nord ci spingiamo:
- in primo luogo abbiamo l'inconveniente della dormienza invernale (stop della crescita e perdita del colore con temperature sotto gli 0°C).
Introduzione
- in secondo luogo con temperature ancora più limitanti è possibile incorrere in una certa percentuale di mortalità nel tappeto erboso.
La causa della mortalità durante l'inverno è la temperatura del terreno che danneggia le corone (Beard, 1984).
Il metabolismo delle piante è influenzato principalmente dalle temperature massima e minima e dalla durata del giorno.
Le microterme hanno due picchi di crescita il primo e più grande in primavera quando escono dalla dormienza. In estate la crescita subisce un rallentamento. In autunno l'attività vegetativa inizia a riprendere fino a rallentare nuovamente quando le temperature diminuiscono in Inverno. Le microterme in Inverno entrano in uno stato di "semidormienza" e il tappeto erboso mantiene il colore. In questo periodo vengono accumulati negli organi di riserva i carboidrati che saranno utilizzati in Inverno. D'altra parte le macroterme entrano in dormienza in Inverno alla prima gelata e le parti aeree possono disseccare. Solitamente quando la temperatura dell'aria scende sotto i 10°C si assiste a una perdita di colore del tappeto erboso. Escono dalla dormienza verso Aprile e raggiungono il massimo della crescita in estate.
1.3 Cynodon spp.
Cynodon spp. (Bermudagrass) (fam.: Poaceae, sottofam.:
Eragrostideae, tribù: Chlorideae) è un genere originario del bacino dell'Oceano Indiano, del Corno d'Africa. È stato introdotto in America verso la metà del 1700 (Hanson et al., 1969) ed è ormai naturalizzata negli USA meridionali.
Introduzione
È un genere al quale appartengono specie perenni, con crescita attiva in Primavera, Estate e Autunno. Il genere comprende 9 specie tra cui la più diffusa è Cynodon dactylon (L.) Pers. Tale specie è diffusa largamente fra il 45°N e il 45°S (Anderson et al., 1993; Harlan and de Wet, 1969). È molto utilizzata per i tappeti erbosi sportivi per la sue caratteristiche: rapida ripresa dopo l'usura, crescita veloce, tessitura medio-fine e elevata densità.
Cynodon dactylon (L.) Pers è ottimale per le zone calde e
siccitose. Non ha bisogno di una particolare tessitura del terreno e si adatta a pH che variano fra 5 e 8. Viene definita una specie tollerante alla salinità anche se ci sono notevoli differenze fra le varie cultivars (K.Razmjoo, J.Sugiura,
S.Kaneko, 1997). È infatti coltivata sui suoli salini della
Central Valley in California, irrigata con acque saline e usata per il pascolo del bestiame (Kaffka, 2009).
Non è però altrettanto tollerante all'ombra. Con intensità luminose inferiori al 60% rispetto alla piena luce l'apparato radicale si indebolisce, gli internodi e le foglie si allungano. Il tappeto erboso diminuisce la sua densità con crescenti problemi di malattie e infestanti.
Produce sia stoloni che rizomi in relazione alle condizioni di disponibilità di nutrienti e luce. La ramificazione degli stoloni e dei rizomi è ridotta in corrispondenza di una riduzione della luminosità e della disponibilità di nutrienti. Gli stoloni permettono alla Cynodon di andare in cerca di luce, mentre i rizomi servono come organi di stoccaggio di risorse e contengono meristemi (Ming Dong and Hans de Kroon, 1994).
Introduzione
Tra le macroterme più utilizzate la Cynodon è la più resistente alla siccità, la più veloce ad insediarsi e ha rizomi/stoloni molto estesi.
Gli ibridi della Cynodon dactylon sono stati ottenuti dall'incrocio fra Cynodon dactylon e Cynodon transvaalensis.
Questi, in generale, hanno una minore larghezza fogliare, minor evapotraspirazione, maggior resistenza alla salinità, sopportano tagli più bassi, necessitano di più azoto e possono essere propagate solo vegetativamente. Un ruolo di fondamentale importanza per la realizzazione e lo studio di nuovi ibridi è attribuito alla stazione sperimentale di Tifton in Georgia. Qui le ricerche iniziarono nel 1946.
Le prove sperimentali che vi sono state condotte sono state incentrate sulla ricerca di nuove varietà di specie da tappeto erboso che potessero soddisfare varie esigenze, testandone anche la resistenza all'usura, l'esigenza di elementi nutritivi etc. Il primo obiettivo di questa stazione sperimentale fu quello di sviluppare una nuova Cynodon con caratteristiche più adatte per i greens dei campi da golf e con propagazione vegetativa.
Burton (1991) riassunse i punti salienti della storia della stazione sperimentale. La prima varietà lanciata sul mercato fu la Tiflawn nel 1952. Questa venne poi superata perché risultò avere una tessitura troppo spessa e una crescita eccessiva per i greens. Per realizzare un ibrido con tessitura più fine venne incrociata la Tiflawn con la Cynodon
transvaalensis. A seguito vennero realizzate due migliori e
più utilizzate varietà: la Tifgreen nel 1956 e la Tifway nel 1960. Questa risultò migliore sia per il colore, più scuro, che per una maggiore resistenza al freddo, una crescita precoce in primavera, maggior resistenza agli stress biotici e
Introduzione
abiotici e una maggior rigidità delle lamine che la rende migliore per l'uso nei greens. Altra varietà realizzata a Tifton è la Tifdwarf nel 1965. Si scoprì dopo vari test che la varietà nana si trattava di una mutazione naturale della Tifgreen. Nel 1970 vennero inoltre sottoposte all'esposizione con raggi gamma Tifgreen, Tifway e Tifdwarf per ottenere mutanti.
Introduzione
1.4 Zoysia spp.
Il genere Zoysia (fam.: Poaceae, sottofam.: Chloridoideae, tribù: Zoysieae) è formato da circa 10 specie di piante perenni, macroterme. È originaria dell'Asia orientale. Il naturale areale di distribuzione del genere si estende dalla Nuova Zelanda all'isola di Hokkaido in Giappone comprendendo la Polinesia e le regioni sulla costa del Sudest asiatico
(Engelke and Anderson, 2003).
La Zoysia viene utilizzata nelle regioni calde e nelle zone di transizione per i tappeti erbosi sportivi, compresi quelli da golf, ma anche per i tappeti erbosi ornamentali.
I vantaggi che presenta sono una buona resistenza all'ombreggiamento, alla siccità, un'eccellente resistenza al calpestamento e una buona resistenza al freddo. Da sottolineare la bassa necessità di inputs che la Zoysia richiede. Fra le macroterme nella famiglia delle Poaceae, il genere Zoysia è uno dei generi che comprende le specie più tolleranti al sale e alle basse temperature.
La resistenza al freddo è più alta rispetto alla Cynodon, infatti entra più tardi in dormienza e riprende prima l'attività vegetativa. Un motivo per cui può non essere scelta per tappeti erbosi sportivi di frequente utilizzo è che ha una scarsa velocità di recupero, nonché di insediamento e germinazione.
È caratterizzata dall'avere un'alta densità, una tessitura medio-fine. Produce sia rizomi che stoloni.
La Zoysia viene usata di frequente nelle zone di transizione per esempio per i fairway, tees e i rough grazie
Introduzione
alla sua grande resistenza alle alte temperature, alla buona densità e una bassa richiesta di trattamenti chimici.
Le tre specie più utilizzate e quindi di importanza economica più rilevante sono: Z.japonica (Steud.), Z.matrella (L.) Merril, Z.tenuifolia Will.ex Trin. Queste tre specie sono tutte originarie del Giappone. Il Giappone è considerato il centro di differenziazione del genere Zoysia.
Tra le varie cultivar possiamo citare "Cavalier" (Z.
matrella), "Diamond" (Z. matrella), "Palisades" (Z. japonica).
Queste cultivars permettono di avere un tappeto erboso di alta qualità, ma non sono abbastanza resistenti al freddo per l'utilizzo nelle zone di transizione (Morris, 1996 e 2001).
Da uno studio è emerso che in Indiana la Diamond e la Cavalier hanno avuto una mortalità invernale del 95% e 45% rispettivamente, mentre la Zoysia japonica Meyer non ha subito alcun danno (Patton and Reicher, 2007).
La Zoysia "Meyer", prende il nome da Frank N.Meyer che raccolse i semi di Zoysia in Korea nel 1905. Si tratta di una Zoysia japonica migliorata, è stata selezionata in base alla larghezza delle foglie (media tra Z. japonica e Z. matrella) e alla resistenza al freddo.
La Zoysia venne utilizzata per la prima volta nei campi da golf nel 1950 quando la Meyer venne distribuita sul mercato.
Per molto tempo è stata la cultivar più utilizzata nelle zone di transizione per la sua buona resistenza al freddo. La Meyer ha comunque dei punti deboli: una tessitura grossa rispetto alle cultivars di Z. matrella, si insedia e recupera lentamente (Fry and Dernoeden, 1987), è inoltre suscettibile a contrarre alcune malattie fra cui Rhizoctonia solani (Green et
Introduzione
Nuove cultivars sono entrate in uso dopo che alcuni ricercatori della Texas A & M University negli anni '80 raccolsero del germoplasma in Asia. Tra queste possiamo citare le seguenti cultivars a propagazione vegetativa: ‘Cavalier’ (Z. matrella) (Engelke et al., 2002a), ‘Crowne’ (Z. japonica)
(Engelke et al., 2002b), ‘Diamond’ (Z. matrella) (Engelke et al., 2002c), e ‘Palisades’ (Z. japonica) (Engelke et al., 2002d).
Negli esperimenti condotti in condizioni controllate di laboratorio ‘Belair’ (Z. japonica), ‘Chinese Common’ (Z. japonica) e la Meyer hanno avuto una ricrescita dai rizomi esposti a -18°C, mentre non sono stati osservati tessuti vivi nelle cultivars: Cavalier, Crowne, Emerald (Z. japonica x Z.
pacifica)e la Palisades dopo l'esposizione a temperature
inferiori a -10 C (Dunn et al., 1999). Test condotti in campo in
Indiana hanno confermato i risultati di laboratorio: Diamond fu la più danneggiata dal freddo (98-100 %), seguita dalla
Palisades (31-61%) e la Cavalier (17- 45%); mentre Meyer e la Chinese Common non subirono danni (Patton and Reicher, 2007).
1.5 Resistenza al freddo di Cynodon e Zoysia
Cynodon spp. (Bermudagrasses), per le sue qualità, viene
utilizzata frequentemente anche nelle zone di transizione. In queste aree come in Italia queste specie subiscono frequentemente danni nel periodo invernale. Questo crea molti problemi per i gestori di tappeti erbosi. I manutentori fanno molti sforzi per ritardare l'entrata in dormienza della
Cynodon e limitarne i danni da congelamento e questi sforzi si
moltiplicano se dal tappeto erboso in questione ci si aspetta un alto standard qualitativo.
Introduzione
Alcuni inverni possono essere talmente freddi da provocare un'alta mortalità di esemplari all'interno del tappeto erboso e questo è causa di notevoli perdite economiche.
Per la difficoltà di prevedere l'entità del freddo invernale e delle condizioni di campo sono stati fatti anche molti studi di laboratorio allo scopo di definire la resistenza alle basse temperature e/o al congelamento. Queste prove vengono fatte sia per vedere le potenzialità di utilizzo della Cynodon nelle zone di transizione, per testare la resistenza di nuove cultivars e la possibile interferenza che possono avere eventuali trattamenti con fertilizzanti e/o regolatori di crescita. Un altra applicazione è quella di poter selezionare genotipi resistenti per brevettare nuove varietà.
Sappiamo da alcuni studi che la resistenza al freddo varia molto all'interno della stessa specie a seconda della varietà presa in considerazione. GN-1 -5.9°C Baby -6.7°C TifSport -7.2°C Quickstand -8.0°C Midlawn -8.4°C
Tabella 1.3. Tolleranza al congelamento di varietà di Cynodon da Fairway propagate vegetativamente.
(Anderson at al., 2005) Arizona Common -5.6°C Mirage -6.1°C Jackpot -6.3°C Guymon -7.4°C
Introduzione
Yukon -7.6°C
Tabella 1.4. Tolleranza al congelamento di varietà di Cynodon da Fairway da seme. (Anderson at al., 2005) Champion -4.8°C Floradwarf -4.9°C MS-Supreme -5.2°C MiniVerde -5.8°C Tifeagle -6.0°C Tifdwarf -6.5°C Tifgreen -6.5°C
Tabella 1.5. Tolleranza al congelamento di varietà di Cynodon da Green propagate vegetativamente.
(Anderson at al., 2005)
Zoysia spp. si spinge più a Nord delle altre specie
macroterme da tappeto erboso resistendo a temperature più basse, anche se la variabile varietà può influenzare molto il confronto fra le specie.
Spesso le nuove cultivars per tappeti erbosi sportivi più performanti hanno un'inferiore resistenza alle basse temperature.
Ci sono differenze nella resistenza al freddo delle varie specie e cultivars di Zoysia e sono state valutate negli anni con diversi studi di campo e di laboratorio.
Attraverso gli esperimenti fatti la Meyer ha dimostrato una maggiore resistenza al congelamento della Zoysia matrella
(Rogers et al., 1977).
Nuove cultivars messe sul mercato dopo Meyer hanno una migliore tessitura, resistenza all'ombra, alla siccità, agli attacchi di insetti e si stabiliscono più velocemente in confronto a Meyer (White and Engelke, 1990; Reinert and Engelke,
Introduzione
2001; White et al., 2001; Patton et al., 2007; Karcher et al., 2005).
A seguito di studi di resistenza alle basse temperature è emerso che le nuove cultivars sono meno resistenti di Meyer
(Dunn et al., 1999). Patton e Reicher (2007) hanno determinato
la resistenza al congelamento di alcune cultivars di Zoysia trovando che questa resistenza varia da 8,4°C (Diamond) a -11,5°C (Meyer e Zenith). Tra le cultivars di Zoysia matrella le meno resistenti sono Diamond, Royal e Zorro; fra quelle di Zoysia japonica è la Victoria la meno resistente. Le cultivars di Zoysia japonica, Meyer, Zenith, Palisades, El Toro,
Companion e J-36, invece hanno mostrato tutte una migliore e
simile resistenza al congelamento. Sappiamo però che la resistenza al congelamento testata in laboratorio non rispecchia sempre la resistenza alle basse temperature in natura (Anderson et al., 2002). Questo è dovuto ai molteplici fattori addizionali che possono interferire in condizione di campo (copertura nevosa, umidità del suolo e oscillazioni della temperatura) (Patton e Reicher, 2007). In più le condizioni di campo possono favorire una maggiore "acclimatazione" rispetto a quella indotta nella camera di crescita (Anderson et al., 2002). In sintesi dallo studio di Patton e Reicher emerge che le cultivar di Zoysia japonica hanno una maggior resistenza al freddo e al congelamento e fra queste le più resistenti sono quelle propagate da seme.
Introduzione
1.6 Congelamento e stress causato dalle
basse temperature
Ogni specie e varietà ha il suo range di temperature ideali per la crescita e la riproduzione. Questo fa sì che le piante siano distribuite geograficamente in dipendenza alle temperature massime e minime che si registrano in quell'areale.
Uno dei primi effetti delle basse temperature è il rallentamento del metabolismo. Vengono rallentate le reazioni enzimatiche e questo ha delle gravi conseguenze su moltissimi processi biologici causando vari danni indiretti. Un altro effetto causato dall'abbassamento di temperatura è il cambiamento di conformazione dei polipeptidi. Le membrane subiscono modifiche strutturali, fisiche e chimiche.
Le basse temperature possono dare vari tipi di stress che si possono differenziare in stress causati da basse temperature, ma superiori allo zero e in condizioni di vero e proprio congelamento con esposizione a temperature inferiori a 0°C. Alcune piante rispondono al freddo con una risposta biologica detta "acclimatazione al freddo". Riescono cioè ad incrementare progressivamente la loro resistenza alle basse temperature dopo l'esposizione a basse temperature, ma inferiori al punto di congelamento. Le risposte di tolleranza al freddo e al congelamento non sono uguali, ma si considerano processi fisiologici distinti. Le lesioni da raffreddamento (chilling) si presentano quando siamo in presenza di temperature troppo basse per una crescita normale, ma non così basse da formare ghiaccio. Nel caso del congelamento abbiamo la formazione di cristalli di ghiaccio all'interno dei
Introduzione
tessuti. La formazione dei cristalli di ghiaccio si verifica fuori dalla membrana plasmatica poiché i fluidi extracellulari hanno un punto di congelamento più alto rispetto a quello del citoplasma.
Il danno più importante che possiamo attribuire allo stress da congelamento è la disidratazione cellulare. Questa è provocata dalla formazione di ghiaccio che abbassa il potenziale idrico extracellulare. In conseguenza a questo l'acqua si allontana dall'interno della cellula fino ad arrivare alla plasmolisi, ovvero la rottura della membrana cellulare. È per questo motivo che i vari stress da congelamento, salino e idrico possono dar luogo a sintomi simili.
La disidratazione a seguito della formazione di ghiaccio viene spiegata con il concetto di osmolalità che si misura come moli di soluto disperse in un kg di acqua. In presenza di due compartimenti contenenti liquido separati da una membrana semipermeabile non abbiamo passaggio di acqua se l'osmolalità è mantenuta uguale e costante nei due comparti. Nel caso l'osmolalità si alzi abbiamo un passaggio di acqua dal compartimento meno concentrato a quello più concentrato. La formazione di ghiaccio aumenta la concentrazione del liquido extracellulare portando a una fuoriuscita dell'acqua dalla cellula. L'osmolalità è dipendente dalla temperatura (kelvin) se consideriamo una soluzione ideale parzialmente congelata:
86 , 1 273 T
Introduzione
Fig 1.1
L'acqua si sposta dal citoplasma verso l'apoplasto
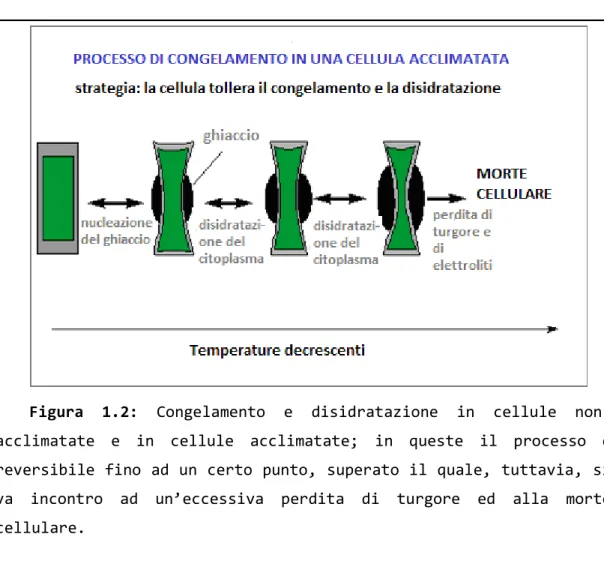
La disidratazione provocata dalla formazione di ghiaccio nei tessuti non acclimatati porta al collasso della cellula, mentre in quelli acclimatati è reversibile fino a un certo livello di danno.
Introduzione
Figura 1.2: Congelamento e disidratazione in cellule non-acclimatate e in cellule non-acclimatate; in queste il processo è reversibile fino ad un certo punto, superato il quale, tuttavia, si va incontro ad un’eccessiva perdita di turgore ed alla morte cellulare.
Durante un congelamento rapido il liquido cellulare si mantiene liquido anche se a temperature ben inferiori al punto di congelamento. Il ghiaccio da punti di potenziale osmotico meno negativo si forma mediante il fenomeno di nuclezione del
ghiaccio (Burke e Lindow, 1990).
La nucleazione avviene grazie all'azione di vari tipi di molecole (polisaccaridi, amminoacidi) che assumono la funzione di nucleatori del ghiaccio. Il raffreddamento veloce detto
supercooling è un fenomeno durante il quale, nella fase
iniziale, la temperatura scende sotto la temperatura di congelamento del citosol e del vacuolo senza che ci sia formazione di ghiaccio. Questo fenomeno è sfruttato dalle piante arboree per evitare la disidratazione delle cellule. In alcune specie l'acclimatazione consiste proprio in un
Introduzione
adattamento per il quale viene eliminata la possibilità di nucleazione del ghiaccio nel protoplasto. In questo caso è possibile un supercooling molto forte senza avere formazione di ghiaccio. Tutto questo ha un limite minimo che è -40°C, temperatura alla quale i cristalli di ghiaccio si formano anche senza punti di nucleazione.
1.7 Metabolismo e tolleranza al freddo
La tolleranza al freddo nelle piante erbacee è stata spesso correlata al metabolismo di queste, in particolare al contenuto di carboidrati di riserva, a quello degli zuccheri solubili e delle proteine.
Su Arabidospis thaliana è stato fatto nel 2004 uno studio che metteva a confronto il metabolismo di due genotipi diversi in relazione alla tolleranza alle basse temperature (Klotke et
al., 2004). Da questo studio è emerso che mentre il contenuto
totale di zuccheri solubili era molto simile nei due genotipi, c'erano notevoli differenze fra i genotipi per quanto riguardava il rapporto fra zuccheri solubili e amido.
Il genotipo meno tollerante risultava avere una maggiore concentrazione di amido nelle foglie e quindi una maggiore presenza totale di carboidrati non strutturati nelle foglie non acclimatate. Il genotipo più resistente, invece, accumulava più zuccheri in particolare saccarosio e raffinosio piuttosto che esosi nelle foglie. Il raffinosio e il saccarosio sono soluti con una funzione più crioprotettiva degli esosi (Crowe et al., 1990) e questo può spiegare la maggiore tolleranza al freddo del genotipo in questione. È emerso dallo studio un forte ruolo crioprotettivo del raffinosio.
Introduzione
Gli zuccheri come il glucosio, il saccarosio e il raffinosio aiutano a mantenere la membrana cellulare integra durante il congelamento, preservando le caratteristiche fisiche della membrana (Caffrey et al., 1988; Crowe et al.,
1988).
In più gli RFO (raffinose family oligosaccharides) come il raffinosio e lo stachiosio possono impedire la cristallizzazione del saccarosio a basse temperature (Caffrey
et al., 1988; Koster and Leopold, 1988). Gli zuccheri, incluso il
glucosio e il saccarosio possono proteggere la struttura delle proteine durante il congelamento (Carpenter and Crowe, 1988).
Si parla qui di una naturale capacità di resistere alle basse temperature senza ricorrere all'acclimatazione. Di certo però la tolleranza al freddo è una carattere di resistenza poligenico e non può essere riferito direttamente a un solo metabolita. Ad esempio un aumento nella concentrazione di zuccheri solubili in genotipi non aventi particolari capacità di acclimatazione non aumenta la tolleranza al freddo. Questo suggerisce che non abbiano un ruolo solo gli zuccheri solubili, ma anche vari amminoacidi come le proline e le glutammine che sono state trovate in concentrazione maggiore nel genotipo più resistente. Le proline hanno la capacità di favorire gli aggiustamenti osmotici e la protezione delle strutture cellulari durante la deidratazione. È stato anche dimostrato che durante il freezing in vitro le proline sono in grado di stabilizzare le proteine (Carpenter, Crowe e Arakawa, 1990). I risultati dello studio suggeriscono un ruolo indiretto del saccarosio come substrato metabolico per la sintesi di altri composti crioprotettivi e non una diretta funzione crioprotettiva dello stesso.
Introduzione
In generale le molecole che giocano un ruolo nel favorire la resistenza al freddo sono:
• lipidi: i lipidi costituendo la membrana cellulare condizionano con le loro caratteristiche fisiche le caratteristiche di questa. Spesso le piante più resistenti al freddo hanno una maggior quantità di acidi grassi insaturi come costituenti della membrana. Questo avviene perché con una maggior quantità di acidi grassi insaturi si abbassa la temperatura alla quale i lipidi di membrana cominciano il graduale cambiamento da fase liquida a semi-cristallina. Durante il processo di acclimatazione si ha un aumento degli acidi grassi insaturi come protezione dal freddo (Williams et al.,
1988; Palta et al., 1993; Uemura e Steponkus, 1997)
• proteine: è stato riconosciuto il ruolo di alcune proteine con funzione antigelo. Queste si legano alla superficie dei cristalli di ghiaccio rallentandone o impedendone l'accrescimento all'interno della cellula.(anti-freeze proteins, AFPs) (Wisniewski et al., 1999).
• zuccheri: contribuiscono alla stabilizzazione delle membrane durante la disidratazione causata dalle basse temperature, si accumulano nelle pareti cellulari e limitano la crescita dei cristalli.
Introduzione
1.8 Tecniche agronomiche per aumentare la
resistenza al freddo
Il tasso di mortalità invernale delle piante in campo dipende principalmente dalla temperatura del terreno e dell'interfaccia suolo-aria, ma è correlato anche a un'altra serie di variabili: il grado di acclimatazione delle piante, il livello di idratazione dei tessuti, la velocità del congelamento (più è rapido, più è alto il tasso di mortalità), la velocità di scongelamento (anche in questo caso, più è rapido, più è alto il tasso di mortalità), il numero di cicli di congelamento e scongelamento, la durata del periodo in cui si mantengono le temperature che causano il freezing. Tra questi fattori quello che riguarda il livello di idratazione è il più importante (Beard, 1984).
Per evitare che il contenuto idrico all'interno dei tessuti sia eccessivo e possa essere negativo per la vitalità delle piante in caso di temperature sotto lo zero, è necessario favorire un buon drenaggio del terreno con le tecniche di coltivazione.
Altri fattori che influenzano la mortalità invernale sono: il livello di azoto utilizzato per la concimazione, la quantità di potassio, l'altezza di taglio del tappeto erboso, la presenza o meno del feltro.
La presenza del feltro, al contrario, è negativa perché questo alza il livello delle corone esponendole maggiormente. Inoltre questo è in grado di impregnarsi e trattenere l'acqua aumentando il livello di idratazione.
È stata osservata una maggiore mortalità invernale in caso di altezze di taglio basse. Questo si verifica perché una
Introduzione
maggiore quantità di biomassa favorisce l'accumulo dei carboidrati che hanno un ruolo centrale nell'acclimatazione, in più essa ha la funzione di proteggere le corone ostacolandone l'esposizione alle basse temperature.
Non esagerare con le concimazioni azotate e le irrigazioni, aumentare la dose di potassio, eliminare il feltro sono tutte tecniche che sfavoriscono la mortalità invernale (Beard, 1984).
Alcuni ricercatori hanno dimostrato che elevati livelli di potassio e fosforo giocano un ruolo fondamentale nell'aumentare la sopravvivenza agli stress da basse temperature (Roberts, 1993; Johansson, 1994).
Al contrario, un'abbondante fertilizzazione azotata effettuata in tarda stagione crea un tappeto erboso rigoglioso che è particolarmente suscettibile ai danni da freddo e alle malattie.
1.8.1 Il ruolo dei fertilizzanti
Un buon programma di fertilizzazione dovrebbe: aumentare la crescita della parte aerea senza aumentare troppo la crescita verticale dei germogli, migliorare il colore del tappeto erboso, stimolare la crescita radicale e favorire il processo fotosintetico. Tra gli effetti non desiderati che uno sbagliato piano di fertilizzazione può causare si riscontrano: un'eccessiva crescita che obbliga a rendere più frequenti i tagli e una riduzione della tolleranza agli stress fra cui lo stress da calpestamento, la resistenza alla siccità e la resistenza alle basse temperature (Duff, 1976).
M.D. Richardson, (2002), testò l'effetto dell'applicazione in tardo autunno di Trinexapac-ethyl e fertilizzazione azotata
Introduzione
sulla morfologia, la qualità e la tolleranza alle basse temperature della Cynodon dactylon 'Tifway'. Dallo studio è emerso che le applicazioni di azoto in tardo autunno hanno avuto il principale effetto di diminuire il periodo di dormienza della 'Tifway'. D'altro canto a questo effetto positivo della fertilizzazione azotata si affiancano almeno due conseguenti effetti negativi. Uno è quello di favorire l'incremento delle infestanti annuali invernali, l'altro quello di rendere le piante più suscettibili alle gelate tardive.
Da uno studio condotto nel Dipartimento di Biologia dell'Università di Pisa in collaborazione con il CeRTES (Centro Ricerche Tappeti Erbosi Sportivi) con lo scopo di testare la relazione fra la LT50 (median lethal temperature,
temperatura che uccide il 50% della popolazione) di Zoysia
matrella 'Zeon' con l'applicazione in tardo autunno di due
differenti dosi di azoto (15 e 30 g m-2), emerge l'azione positiva della fertilizzazione azotata sul ritardo dell'entrata in dormienza. L'entrata in dormienza è stata ritardata di più di un mese (Pompeiano et al., 2011).
La capacità delle cellule vegetali di sopravvivere alle basse temperature è stata spesso associata alle caratteristiche osmotiche del tessuto vegetale e la capacità degli osmoliti di prevenire il congelamento dell'acqua intra e extra cellulare. (Hsiao, 1973).
Gilbert e Davis (1971) dimostrarono che è maggiore il vantaggio di applicare dosi bilanciate di azoto e potassio, piuttosto che eccedere o nell'uno o nell'altro elemento.
MD Richardson (2002) ha evidenziato che l'applicazione di azoto nel tardo autunno non ha effetto sulla tolleranza alle basse temperature quando il potassio non è limitante.
Introduzione
Più recentemente, nel 2005, D.E. Webster e J.S. Ebdon condussero uno studio con lo scopo di valutare l'interazione dell'azoto e del potassio sulla resistenza alle basse temperature del Lolium perenne L..
In particolare gli obiettivi dello studio erano quelli di: valutare la resistenza alle basse temperature in fase di deacclimatazione in relazione all'azoto e il potassio, valutare i conseguenti effetti sull'idratazione delle corone, individuare la LT50, misurare il tasso di crescita dei
germogli, la concentrazione di potassio nei tessuti, la quantità di potassio scambiabile nel terreno e l'incidenza delle malattie conseguenti alle basse temperature. Vennero distribuite cinque dosi crescenti di azoto ognuna in combinazione con tre concentrazioni di potassio. Da questo studio è emersa una correlazione negativa fra tolleranza al freddo e idratazione delle corone, tasso di crescita dei germogli e contenuto di potassio nei tessuti.
La sopravvivenza degli esemplari e la LT50 sono correlate
direttamente alle applicazioni di azoto e potassio e alla loro interazione. La maggior tolleranza al freddo è stata osservata con l'applicazione in tardo autunno di concentrazioni basse di azoto accompagnate da medio - alte concentrazioni di potassio. D'altra parte simili concentrazioni di potassio distribuite con alte concentrazioni di azoto hanno aumentato lo stress da basse temperature e anche gli attacchi da parte di funghi che si presentano solitamente con le basse temperature (es.
Typhula incarnata).
Un aspetto che favorisce la mortalità a basse temperature è il contenuto di umidità delle corone. La concentrazione più alta di azoto applicata in tardo autunno, che è risultata essere la più dannosa dal punto di vista della tolleranza al
freddo, aumenta significativamente il contenuto idrico delle corone. In più l'applicazione di alte concentrazioni di azoto favoriscono, rispetto a dosi più basse, un aumento del tasso di crescita delle foglie e una conseguente accelerazione della riduzione delle riserve di carboidrati.(Hull e Smith, 1974).
Scopo della tesi
Capitolo 2
Scopo della tesi
Con questa sperimentazione è stato possibile studiare la resistenza al congelamento di due specie macroterme da tappeto erboso molto usate: Zoysia matrella (L.) Merr. cv. ‘Zeon’ e
Cynodon dactylon x C. transvalensis Burtt. Davy cv. ‘Tifway’.
Questo con lo scopo di valutare la possibile espansione del loro areale di utilizzo, ovvero la possibilità di utilizzarle in zone di transizione minimizzando i probabili danni invernali e sfruttando al tempo stesso i vantaggi già elencati che l'uso di macroterme può portare.
L'obiettivo è anche quello di mettere in relazione la resistenza al congelamento con il metabolismo in funzione dell'applicazione di due diverse forme azotate (NH4+, NO3-).
In particolare le caratteristiche del metabolismo vengono studiate prima del congelamento. Questo permette di analizzare le possibili influenze delle diverse concentrazioni di amido (negli stoloni), zuccheri solubili (nei vari organi) sulla mortalità delle piante e il tasso di crescita delle porzioni rimaste vitali dopo il congelamento delle piante.
È stato anche valutata la concentrazione di pigmenti fogliari per le varie tesi nei vari tempi per correlare l'attività vegetativa con l'uso delle due forme azotate e la mortalità.
Scopo della tesi
Allo stesso momento, è stato possibile valutare l'eventuale effetto positivo o meno di una applicazione di due tipi di fertilizzanti azotati antecedente al congelamento. I fertilizzanti utilizzati sono stati solfato d'ammonio e nitrato di calcio.
Spesso nelle sperimentazioni sulla resistenza al congelamento questa è stata correlata all'uso di fertilizzanti azotati per valutare se l'applicazione di azoto in tardo autunno possa avere un effetto positivo sul metabolismo in modo da aumentare la resistenza al congelamento.
Materiali e metodi
Capitolo 3
Materiali e metodi
3.1 Insediamento delle cultivars
Le cultivars sono state insediate il 27 Aprile 2011 presso il Dipartimento di Biologia dell'Università di Pisa.
Le cultivars utilizzate sono state: Cynodon dactylon × C.
transvaalensis Burtt. Davy cv. ‘Tifway’ e Zoysia matrella (L.)
Merr. cv. ‘Zeon’. Queste sono state propagate vegetativamente raccogliendo fitomeri (segmenti di 1 o 2 centimetri di stolone comprendenti anche radici, corona e foglie).
Il substrato dove sono stati posti i fitomeri è stato messo in vassoi per la semina da 160 fori, ogni cella aveva 5 cm3 di volume.
Il mezzo di crescita era costituito da torba di sfagno e sabbia vulcanica.
Le piante sono state per 24 settimane in serra con una temperatura di 23°C (±7°C) giorno e notte.
L'acqua è stata fornita per subirrigazione a seconda del bisogno per prevenire l'appassimento delle piante.
L'altezza di taglio è stata mantenuta a 2,5 cm prima e dopo l'inizio dei trattamenti.
Le piante mature sono poi state acclimatate in una camera di crescita per 8 settimane prima dell'inizio dei trattamenti.
Materiali e metodi
Le condizioni della camera di crescita sono state: 22°C (±1°C) di temperatura, un fotoperiodo di 12 ore e un'intensità luminosa di 90 µmol m-2s-1.
L'azoto è stato distribuito per concimazione fogliare in due forme diverse al tempo zero. Sono stati usati due diversi fertilizzanti azotati: il solfato d'ammonio [(NH4)2SO4] e il
nitrato di calcio [Ca(NO3)2].
La prova prevede anche il controllo al quale non viene applicato alcun fertilizzante azotato.
Il liquido è stato nebulizzato per facilitare l'assorbimento fogliare. La dose utilizzata è stata di 10 g/m2 di azoto che corrisponde alla dose di azoto comunemente applicata per via fogliare
3.2 Trattamenti
La prova consiste nel sottoporre le piante all'esposizione a temperature sotto zero. Per cinque volte; zero, una, due, quattro e otto settimane dall'inizio dei trattamenti (t0, t1, t2, t4 e t8); un vassoio per ogni tesi è stato messo in un apposito congelatore.
Il giorno prima di ogni trattamento è stato eseguito il taglio delle piante riportandole a un'altezza di 2,5 cm.
A partire da una temperatura iniziale di -5°C ogni ora la temperatura è stata abbassata di 2°C fino ad arrivare a -19°C (-5°C, -7°C, -9°C, -11°C, -13°C, -15°C, -17°C, -19°C). Prima di ogni cambiamento della temperatura cinque piante per ogni tesi sono state prelevate. Le piante che hanno subito il trattamento saranno poi destinate al recovery.
Materiali e metodi
Testare la resistenza al congelamento delle piante nella loro interezza, ovvero con le radici delle piante nel substrato, è importante perché rende le condizioni dell'esperimento più simili a quelle di campo (Repo e Ryyppö, 2008).
Per ogni tesi, controllo, NH4+ e NO3-, un plateau
rettangolare con 40 alveoli (5x8) è stato sottoposto a congelamento e uno al campionamento non distruttivo.
3.3 Campionamento
Il campionamento è stato eseguito in corrispondenza di ogni trattamento. Lo scopo del campionamento è quello di raccogliere materiale per le prove di laboratorio con l'obiettivo di determinare il contenuto di zuccheri solubili, di amido e pigmenti fogliari prima del congelamento. È stato sempre eseguito in tarda mattinata per far sì che le condizioni del metabolismo delle piante fossero simili per ogni trattamento.
La metodologia di campionamento seguita prevedeva di raccogliere dei campioni di circa 100 mg ognuno. Sono stati raccolti campioni per ogni organo e per ogni tesi, in particolare: sei campioni di foglie, sei campioni di rizomi, tre di steli e tre di radici per il controllo, lo stesso numero di campioni per la tesi trattata con NH4+ e per quella
trattata con NO3-.
Le analisi di laboratorio sono state condotte in duplicato con un campione extra di scorta per quanto è riguardato l'analisi degli zuccheri solubili e dell'amido, mentre in triplicato per l'analisi dei pigmenti fogliari.
Materiali e metodi
Tre campioni di foglie, tre di rizomi e tre di radici per ogni tesi sono stati dedicati all'estrazione degli zuccheri.
Per l'estrazione dell'amido sono stati destinati tre campioni per ogni tesi di rizomi e per quella dei pigmenti i tre campioni di foglie restanti.
3.4 Recovery
Il plateau sottoposto a congelamento è stato poi lasciato in camera di crescita per quattro settimane per il recovery.
Durante il recovery le piante non vengono tagliate, ma vengono irrigate regolarmente. Sono state lasciate crescere liberamente per andare poi, dopo le quattro settimane, a misurare il peso fresco della parte epigea vitale di ogni alveolo.
In questo modo, sono stati raccolti due diversi dati importanti, la vitalità di ogni alveolo, quindi la sopravvivenza degli alveoli alle varie temperature e il tasso di ricrescita dato dal peso fresco di tessuti verdi.
I dati raccolti vengono utilizzati per costruire una curva sigmoide usando la seguente equazione
Sopravvivenza = a + (b - a)/ {1 + exp [c (Tm - T)]
dove "a" rappresenta il livello minimo, "b" rappresenta il livello massimo di sopravvivenza, "c" è la pendenza della linea nel punto di flesso, Tm è la temperatura corrispondente al punto di flesso (LT50) che causa un danno mortale al 50%
Materiali e metodi
3.5 Analisi degli zuccheri solubili
3.5.1 Estrazione dei metaboliti
Sono stati sottoposti all'estrazione degli zuccheri solubili tutti e quattro gli organi campionati (foglie, rizomi, radici e steli). L'estrazione è stata fatta in duplicato per ogni organo di ogni tesi di ogni tempo diverso.
Ogni campione è stato omogeneizzato in un mortaio. Successivamente è stata aggiunta una quantità pari a 12 volumi di HCLO4 5,5%. Il campione è stato mantenuto nel ghiaccio per
almeno un'ora. A seguito è stato centrifugato per 10 minuti a 10000 rpm.
Il surnatante è stato recuperato e neutralizzato con K2CO3
3,5 M. La neutralizzazione si ottiene con un volume di base pari a 1/10 del volume totale di acido. Il processo di neutralizzazione libera anidride carbonica causando la formazione di bolle quindi va eseguito lentamente con piccole quantità alla volta di base. Quando la formazione di bolle si arresta la neutralizzazione è completata. A questo punto è stato controllato il pH con una cartina tornasole. È stato accettato un range di pH fra 5 e 9 perché non ha influenza sull'attività degli enzimi utilizzati in seguito.
Successivamente il campione è mantenuto per almeno un'altra ora nel ghiaccio e poi nuovamente centrifugato per 10 minuti per permettere la completa formazione e precipitazione del sale. Il surnatante viene prelevato e ne viene misurato il volume che servirà per calcolare la concentrazione di zuccheri.
A questo punto i campioni vengono messi in congelatore e conservati fino alla determinazione dei metaboliti.
Materiali e metodi
3.5.2 Procedura per la determinazione
enzimatica degli zuccheri solubili
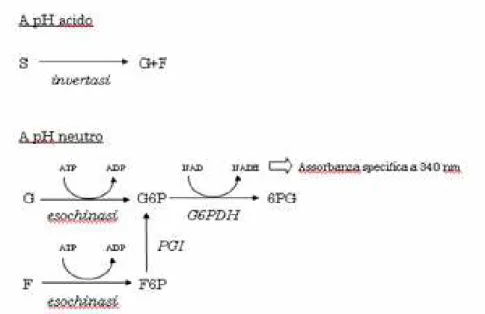
La quantificazione degli zuccheri liberi presenti nel campione si basa sul principio illustrato in figura
Figura 3.1 Schema delle conversioni enzimatiche sfruttate per la quantificazione degli zuccheri liberi. F, fruttosio; F6P, fruttosio fosfato; G, glucosio; G6P, glucosio fosfato; &PG, 6-fosfogluconato, S, saccarosio. G6PDH, glucosio 6-fosfato deidrogenasi, PGI, fosfoglucoisomerasi.
Vengono preparate quattro diverse soluzioni con la seguente composizione (le quantità espresse sono valide per l'analisi di dieci campioni):
Materiali e metodi 1 ml di tampone Na-acetato 50 mM, pH 4.6; 1 mg di invertasi liofilizzata. • Soluzione B 1 ml di tampone Na-acetato 50 mM, pH 4.6. • Soluzione C 4 ml tampone TrisHCl 300 mM, pH 7.6; 3 ml MgCl2 10 mM; 15 mg ATP; 6 mg NADP; 10 U esochinasi;
10 U glucosio 6-fosfato deidrogenasi.
• Soluzione D
100 μl tampone TrisHCl 300 mM, pH 7.6; 20 U fosfoglucoisomerasi.
In questo caso è stata utilizzata un'aliquota di campione pari a 10 μl ed è stata aggiunta acqua distillata fino a raggiungere il volume di 200 μl, in ogni caso l'aliquota di campione utilizzata non può superare i 200 μl.
Ogni singolo campione viene preparato in duplicato in modo che uno dei due duplicati venga posto ad incubare con 100 μl di soluzione A e l'altro con 100 μl di soluzione B. Anche la curva di taratura viene preparata e posta a incubare con 100 μl di soluzione B. L'incubazione ha luogo a 37°C per almeno 30 minuti.
La curva di taratura del glucosio viene preparata a partire da una soluzione standard 1:1 (1 mg ml-1, in acqua distillata). Da questa vengono prelevati 0, 5, 10, 20, 40 e 80
Materiali e metodi
μl corrispondenti a 0, 28, 56, 112, 224 e 448 nmol di glucosio e portati al volume finale con acqua distillata.
Al termine di questa prima incubazione a tutti i campioni e gli standard vengono aggiunti 700 μl di soluzione C e successivamente vengono messi a incubare una seconda volta a 37°C per almeno 15 minuti.
Alla fine del tempo di incubazione l'assorbanza a 340 nm (picco di assorbimento dell'NADPH) di standard e campioni viene misurata e quella dei campioni viene confrontata con la curva di taratura.
Quando la lettura di tutti i campioni è stata fatta ai soli campioni trattati con le soluzioni B e C vengono aggiunti 10 μl di soluzione D e incubati a 37°C per 15 minuti. Viene nuovamente letta l'assorbanza a 340 nm e comparata con la curva di taratura.
In questo modo, dalla lettura dei campioni trattati con soluzione B e C (1) otteniamo la concentrazione di glucosio libero, da quella dei campioni trattati con soluzioni A e C (2) la somma del glucosio libero e quello derivante dall'idrolisi del saccarosio. La differenza (2)-(1) rende la concentrazione di saccarosio presente nel campione. Dall'assorbanza dei campioni trattati con soluzioni B,C e D (3) otteniamo la somma del glucosio e del fruttosio liberi, quindi dalla differenza (3)-(1) si evince la concentrazione di fruttosio libero nel campione.
Le concentrazioni ottenute vengono riportate al volume totale di estratto e al peso fresco (PF) del campione di partenza così da poter esprimere la concentrazione di zuccheri in termini di μmol g-1 PF.
Materiali e metodi
3.6 Analisi dell'amido
La procedura si basa sull'estrazione dell'amido e trattamento con iodio seguito dalla lettura dell'assorbanza.
3.6.1 Estrazione dell'amido
Per estrarre l'amido si procede alla solubilizzazione dello stesso che avviene dopo bollitura in ambiente basico.
Si preparano le seguenti soluzioni stock:
• HCl 1N (8,33 ml a 100 ml con acqua distillata) • KOH 1M (5,6 g a 100 ml con acqua distillata) Di queste l'HCl 1 N viene utilizzato non diluito mentre la soluzione madre di KOH viene diluita come segue:
• KOH 10 mM (soluzione madre diluita 1:100 con acqua distillata)
Il campione viene omogeneizzato in un mortaio con 10 ml si KOH diluito. Viene poi portato a ebollizione e fatto bollire per circa 10 secondi.
Il liquido viene travasato in una provetta tipo Falcon e riportato al volume di 10 ml con acqua distillata. Per neutralizzare il pH viene aggiunto un volume di HCl 1 N pari a un 1/100 del volume totale (100 μl).
Materiali e metodi
Successivamente vengono prelevati 50 μl del campione preparato come sopra e addizionati con 100 μl di acqua.
3.6.2 Determinazione dell'amido con iodio
Per la determinazione dell'amido con iodio vengono preparate le seguenti soluzioni:
• K2 1,3%-KI 3% (1,3 g I2 + 3 g KI a 100 ml con
acqua distillata)
• Soluzione neutralizzata (100 ml KOH 10 mM + 1 ml HCl 1 N)
La soluzione madre di iodio viene diluita come segue:
• Soluzione di iodio diluita 1:10 con acqua distillata
Per la curva di taratura viene preparata una soluzione 1:1 con amido solubile a acqua distillata. In questo caso è consigliabile utilizzare una quantità di amido solubile compresa fra 20 e 40 mg da disciogliere quindi in 20-40 ml di acqua distillata.
La soluzione viene bollita come sopra per circa 10 secondi. Da questa vengono prelevati 0, 20, 40, 60, 80 e 100 μl e portati al volume di 100 μl con acqua distillata. In seguito ad ogni standard vengono aggiunti 50 μl di soluzione neutralizzata.
Materiali e metodi
A standard e campioni viene aggiunto 1 ml di soluzione di iodio diluita.
Dopo 3-4 minuti, quando la reazione è completamente avvenuta, si può procedere alla lettura con spettrofotometro dell'assorbanza a 595 nm.
La concentrazione di amido su peso fresco di campione viene alla fine espressa in mg amido/g PF.
3.7 Analisi dei pigmenti fogliari
La concentrazione nelle foglie di clorofilla a, clorofilla b e carotenoidi è stata determinata per via spettrofotometrica, adattando il protocollo di Zhang e Kirkham
(1996).
3.7.1 Estrazione dei pigmenti
Per l'estrazione dei pigmenti i campioni di foglie sono stati travasati in delle provette tipo Falcon e sono stati aggiunti 10 ml di acetone all'80%.
I campioni vengono mantenuti al buio, alla temperatura di 4°C fino allo sbiancamento dei tessuti che avviene dopo qualche settimana.
3.7.2 Quantificazione dei pigmenti
Al termine del periodo d'incubazione in camera fredda si procede alla lettura allo spettrofotometro dei campioni.
Materiali e metodi
Il surnatante viene letto alle lunghezze d'onda di 663.2, 648.8 e 470 nm, corrispondenti ai picchi di assorbimento rispettivi della clorofilla a (chl a), della clorofilla b (chl b) e dei carotenoidi (car). Il solvente puro rappresenta il bianco.
Le concentrazioni dei tre pigmenti fogliari vengono misurate in μg nl-1surn. e vengono calcolate a partire
dall'assorbanza (Aλ) attraverso le seguenti relazioni:
[chl a] μg ml-1 = (12.25∙A663.2) - (2.79∙A648.8)
[chl b] μg ml-1 = (21.50∙A648.8) - (5.10∙A663.2)
[car] μg ml-1 = [(1000∙A470.0) - (1.82[chl a] + 85.02[chl
b])]/198
Le concentrazioni sono riportate al peso fresco del materiale vegetale (PF) e al volume totale del surnatante (Vsurn.), per esprimere le concentrazioni in termini di mg g-1 PF:
Risultati e discussione
Capitolo 4
Risultati e discussione
4.1 Tolleranza al freddo (LT
50)
4.1.1 Cynodon
Fig. 4.1 Il grafico rappresenta l'andamento nel tempo della LT50 per le varie tesi di Cynodon.
La LT50 rappresenta la temperatura che uccide la metà dei
campioni sottoposti a trattamento di congelamento. Più è bassa, più le piante sono resistenti alle basse temperature e
Risultati e discussione
la mortalità sarà inferiore. Al contrario più si avvicina a valori alti, più la mortalità fra i campioni sarà elevata.
La LT50 per la Cynodon va da un minimo di -13,21°C al t1
del controllo a un massimo di -9,50°C al t2 della tesi trattata con la forma NH4+.
Si mantiene più bassa nel controllo indicando un livello di mortalità inferiore rispetto alle tesi trattate con le due forme azotate. Questo a sostegno del possibile effetto negativo della concimazione azotata sulla resistenza delle piante al congelamento.
Da notare il picco con il quale la LT50 raggiunge il valore
massimo (-9,5°C) alla seconda settimana della tesi NH4+, questo
indica un chiaro effetto negativo sulla resistenza al congelamento della fertilizzazione con azoto in questa forma.
La mortalità della tesi trattata con la forma NO3- si
mantiene, se pur superiore, più simile a quella del controllo eccezion fatta per il t4. Alla quarta settimana la LT50
aumenta molto superando quella della tesi NH4+. Questo indica
un'influenza negativa causata dall'applicazione di fertilizzante azotato sotto forma di NO3- sulla resistenza al
congelamento, che si manifesta alla quarta settimana.
Notiamo che, l'effetto negativo più macroscopico delle due forme azotate sulla resistenza al congelamento, si manifesta in corrispondenza di due tempi diversi a causa, probabilmente, delle diverse velocità di assorbimento delle due forme.
Da segnalare al t8 un aumento della mortalità per tutte e tre le tesi da imputare alle condizioni colturali in camera di crescita che, dopo otto settimane, hanno probabilmente influito negativamente sulla vitalità delle piante.
Risultati e discussione
4.1.2 Zoysia
Fig. 4.2 Il grafico rappresenta l'andamento nel tempo della LT50 per le varie tesi di Zoysia.
La LT50 della Zoysia varia da un minimo di -12,96°C al t2
della tesi NO3-, a un massimo di -8,80°C al t8 del controllo.
La curva presenta gli stessi picchi crescenti di Cynodon alla seconda settimana per la tesi trattata con la forma azotata NH4+ e alla quarta settimana per quella trattata con NO3-,
indicando un forte effetto negativo sulla resistenza al congelamento di queste forme azotate.
Da notare che la Zoysia nella prima settimana risulta più suscettibile di Cynodon alle basse temperature. Alla seconda settimana migliora la resistenza al congelamento e si nota un effetto positivo dell'NO3- sulla riduzione della mortalità. È
Risultati e discussione
controllo, mentre nelle altre settimane risulta sempre più resistente dei due trattamenti. Questo effetto è solo temporaneo, in quanto alla quarta settimana la mortalità nella tesi trattata con NO3- sarà molto superiore al controllo.
Da segnalare anche in questo caso un aumento di mortalità in tutte le tesi in corrispondenza e in prossimità dell'ottava settimana dovuto a un peggioramento generale delle condizioni degli esemplari stressati dalle condizioni colturali non ideali.
Dall'analisi di entrambe le curve di LT50, si può dire che,
la conseguenza dell'applicazione delle due forme azotate sulla resistenza al congelamento sia negativa in entrambi i casi e l'effetto non sia, quindi, specie specifico. Vi è però una precocità di effetto della forma ammoniacale rispetto all’altra, visto che la forma NH4+ causa un aumento della
mortalità in entrambe le specie alla seconda settimana, mentre l'NO3- alla quarta.
Risultati e discussione
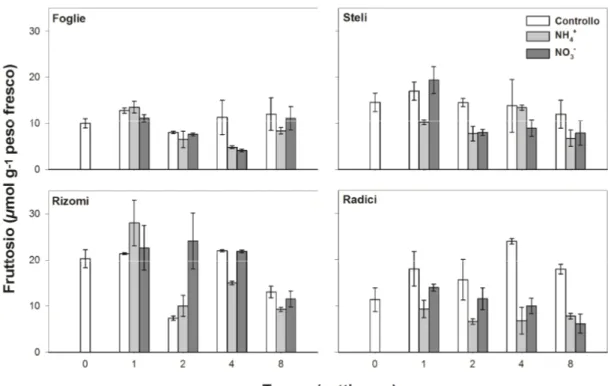
4.2 Zuccheri liberi
4.2.1 Cynodon
•
Saccarosio
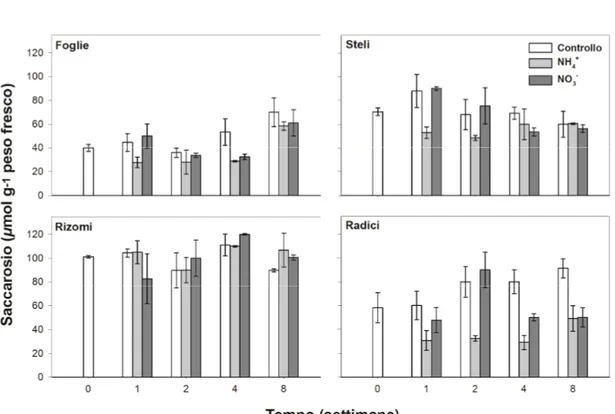
Fig. 4.3 Andamento nel tempo della concentrazione di saccarosio nelle foglie, negli steli, nei rizomi e nelle radici delle tre tesi di Cynodon.
La concentrazione di saccarosio in Cynodon varia da un minimo di 27,80 μmol g-1 al t1 delle foglie della tesi NH4+, a
un massimo di 120,00 μmol g-1 raggiunte dai rizomi della tesi NO3- alla quarta settimana.
Nei rizomi la concentrazione di saccarosio è visibilmente superiore rispetto agli altri organi, mentre le foglie hanno la più bassa concentrazione di questo disaccaride.