R E S E A R C H
Open Access
Community analysis of the abundance
and diversity of biting midge species
(Diptera: Ceratopogonidae) in three
European countries at different latitudes
Tim W. R. Möhlmann
1,2*, Uno Wennergren
2, Malin Tälle
2, Guido Favia
3, Claudia Damiani
3, Luca Bracchetti
3,
Willem Takken
1and Constantianus J. M. Koenraadt
1Abstract
Background: The outbreaks of bluetongue and Schmallenberg disease in Europe have increased efforts to
understand the ecology of Culicoides biting midges and their role in pathogen transmission. However, most studies have focused on a specific habitat, region, or country. To facilitate wider comparisons, and to obtain a better understanding of the spread of disease through Europe, the present study focused on monitoring biting midge species diversity in three different habitat types and three countries across Europe.
Methods: Biting midges were trapped using Onderstepoort Veterinary Institute light traps at a total of 27 locations in Sweden, the Netherlands and Italy, comprising farm, peri-urban and wetland habitats. From July 2014 to June 2015 all locations were sampled monthly, except for during the winter months. Trapped midges were counted and identified morphologically. Indices on species richness, evenness and diversity were calculated. Community
compositions were analysed using non-metric multidimensional scaling (NMDS) techniques.
Results: A total of 50,085 female midges were trapped during 442 collection nights. More than 88% of these belonged to the Obsoletus group. The highest midge diversity was found in Sweden, while species richness was highest in the Netherlands, and most specimens were trapped in Italy. For habitats within countries, diversity of the trapped midges was lowest for farms in all countries. Differences in biting midge species communities were more distinct across the three countries than the three habitat types.
Conclusions: A core midge community could be identified, in which the Obsoletus group was the most abundant. Variations in vector communities across countries imply different patterns of disease spread throughout Europe. How specific species and their associated communities affect disease risk is still unclear. Our results emphasize the importance of midge diversity data at community level, how this differs across large geographic range within Europe, and its implications on assessing risks of midge-borne disease outbreaks.
Keywords: Culicoides, Midge sampling, Species diversity, OVI trap, Community ecology
* Correspondence:[email protected]
1Laboratory of Entomology, Wageningen University and Research, P.O. Box 16, 6700, AA, Wageningen, the Netherlands
2IFM Theory and Modelling, Linköping University, 581 83 Linköping, Sweden Full list of author information is available at the end of the article
© The Author(s). 2018 Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
Background
Worldwide around 1400 species of Culicoides (biting midges, Diptera: Ceratopogonidae) have been de-scribed [1]. At present, the European Interactive Culicoides Key [2, 3] includes 110 species. A minority of these species has thus far been described as im-portant vectors for arthropod-borne viruses (arbovi-ruses) [4, 5]. Biting midges from the Culicoides Obsoletus group [6–11], C. imicola Kieffer, 1913 [12], C. pulicaris(Linnaeus, 1758) [11, 13, 14], and C. punc-tatus (Meigen, 1804) [10, 14–19] are important vectors occurring in Europe. Of the over 50 viruses isolated from biting midges, several are of major international signifi-cance [4]. Animal diseases caused by viruses such as Akabane virus, bovine ephemeral fever virus, African horse sickness virus (AHSV) and bluetongue virus (BTV) are all transmitted by biting midges. Both infection with AHSV and bluetongue are of such international sig-nificance that they are listed by the World Organisa-tion for Animal Health (OIE) as focus diseases that can have serious socio-economic or public health consequences, and are of major importance for inter-national trade [20].
Outbreaks of midge-borne viruses in Europe, e.g. BTV since 2006, and more recently Schmallenberg virus (SBV) during 2011–2013 [21, 22], have had a great impact on the European livestock sector [23–25]. Milk production by infected livestock is often re-duced, and the virus affects unborn calves and lambs when the mother becomes infected during gestation, resulting in non-viable offspring. These impacts, in addition to livestock movement restrictions, result in considerable economic losses. Factors such as inten-sified transportation of livestock and the rise in global temperature may further increase the risk of arbovirus outbreaks in Europe. Rising temperatures create opportunities for vector populations to in-crease rapidly, and allow viruses to complete their extrinsic incubation period in vectors faster, which both imply an increased potential for pathogen transmission [26].
Although research following the outbreaks of BTV and SBV has improved our knowledge of the ecology of Culicoides biting midges, these studies often focus on a specific habitat, region, or country [27–31]. Therefore, it is difficult to make direct comparisons among the results of these studies. Thus, to facilitate wider comparisons on biting midge communities at European level, we aimed to simultaneously sample midge species distribution, abundance, and diversity in three habitat types within three representative countries at different latitudes in Europe, where large differences in environmental characteristics could be expected.
Methods Midge trapping
Adult midges were trapped using Onderstepoort Veterinary Institute (OVI) light traps. A 30 cm 8 W fluorescent black light tube was used to attract midges [1]. When in close proximity of the trap, midges were sucked in by the down-draught fan, which was pow-ered by the main grid or a 12 V, 24–32 Ah battery [14, 17, 32]. The top of the trap was placed at a height of 1.5–2 m and traps were at least 100 m apart to prevent interference between them. The collection bucket had a capacity of 500 ml and larger insects were excluded by polyester netting (mesh size 2–4 mm) placed around the light source of the trap. The bucket was filled with 50 ml water-soap solution.
Sampling procedures
Traps were placed in three countries at different latitudes: Sweden (surroundings of Linköping 58° 24'38.9"N, 15°37'17.5"E), the Netherlands (surround-ings of Wageningen 51°57'53.3"N, 5°39'46.4"E), and Italy (surroundings of San Benedetto del Tronto 42° 56'58.1"N, 13°52'42.6"E). Within each country, farm, peri-urban and wetland habitats were selected. Selec-tion criteria for habitat type and trap locaSelec-tion have been described in [33]. In brief, each habitat type was represented by three unique sampling locations. Traps were positioned in these locations within 50 m of open stables of dairy cattle (‘farm’), a residential property (‘peri-urban’) or standing water (‘wetlands’). Habitat types mostly matched the classification of the CORINE European Land cover database [34].
From July 2014 to June 2015, except for the months December, January and February (and March for Sweden), monthly collections were performed for six consecutive days in each of the countries. Traps were ac-tive for 24 h and were emptied and rotated among the sampling locations between 08:00 h and 17:00 h the next day. Midges were sorted and stored at -20 °C in Eppendorf tubes containing 70% ethanol solution.
Sample identification
All female midges were identified to species level in collections that contained less than 100 individual midges. For collections that contained more than 100 midges (14% of the collections), a random sub-sample of at least 50 individuals was identified as an estimation of the species composition of the total collection. All iden-tifications were performed using the Interactive Identifi-cation Key for Culicoides (IIKC) [2, 3]. Morphologically similar species were recorded as belonging to a group or complex [35,36].
Statistical analyses
Species diversity indices were calculated with the Simpson’s Index of Diversity: 1−D ¼ 1−Pniðni−1Þ
NðN−1Þ , Shannon-Wiener
Diversity Index: H0¼ −PRi¼1pi lnðpiÞ, and the
Shannon-Wiener evenness: E¼ H0
lnðSÞ . Diversity indices were
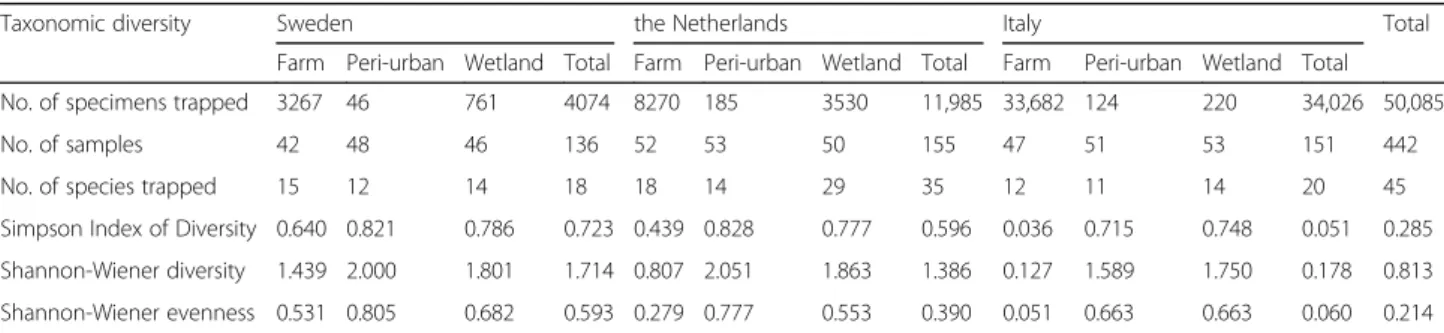
calcu-lated for each of the three countries as well as for farm, peri-urban and wetland habitats. To better understand whether sufficient trapping efforts had been made for a reasonable estimate of species diversity, a rarefaction curve of the species and the number of trapped midges was cre-ated with the rarecurve function within the vegan version 2.9.2. [37] package in R version 3.2.3. [38].
Non-metric multidimensional scaling (NMDS) analyses were used to evaluate the combined effects of country, habitat and diversity on the midge community compos-ition [39]. An NMDS analysis can deal with abundant null measures in a dataset and calculates a reliable best model fit for shortest distances between the elements. The degree of stress calculated within this analysis indicates the reli-ability of the plot that is generated with NMDS, whereby lower stress corresponds to a higher reliability of the plot. For values above 0.3 the NMDS ordination plot is consid-ered arbitrary. For NMDS analyses, the metaMDS func-tion with the Bray-Curtis dissimilarity metric was used. All data were analysed using the statistical software pack-age R version 3.2.3. [38].
Results
A total of 442 trap collections were performed in Sweden, the Netherlands and Italy (Table 1). In 305 (69%) of these collections one or more biting midges were trapped, whereas in the remainder of the collec-tions (31%), no biting midges were trapped. A total of 50,729 biting midges were trapped during this study. Of these specimens, 7818 (15.4%) female midges were identified to species level. After identification, a total of 50,085 female (98.7%) biting midges were estimated to be trapped. Other individuals either were males or dam-aged to the extent that they could not be identified
morphologically. A total of 45 midge species were found for the three countries combined. Of all female biting midges trapped during the field study, the number of specimens trapped was highest for the Obsoletus group (88.6%), followed by C. punctatus (2.3%), C. pulicaris (2.2%) and C. festivipennis (Kieffer 1914) (1.6%).
The rarefaction curves for each of the three countries are beyond their exponential growth and start to level off (Fig. 1). Although more sampling efforts would in-crease the number of species expected to be found (mostly in Sweden and the Netherlands), we believe that our sampling effort was sufficient for obtaining a repre-sentative number of species for the three countries.
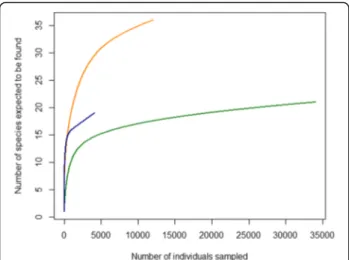
Although the lowest numbers of samples and specimens were trapped in Sweden, the highest species diversity was found there. The lowest values of species diversity were found in Italy, but most specimens were trapped in this country (Table1). The lowest midge species diversity was found at farms in all countries. The highest midge species diversity was found for midges trapped in peri-urban habitats in Sweden and the Netherlands, while the midge diversity was almost similar for peri-urban and wetlands in Italy (Table 1). Catches from peri-urban habitats had the lowest number of specimens, while the highest num-ber of specimens were trapped at farms in all countries.
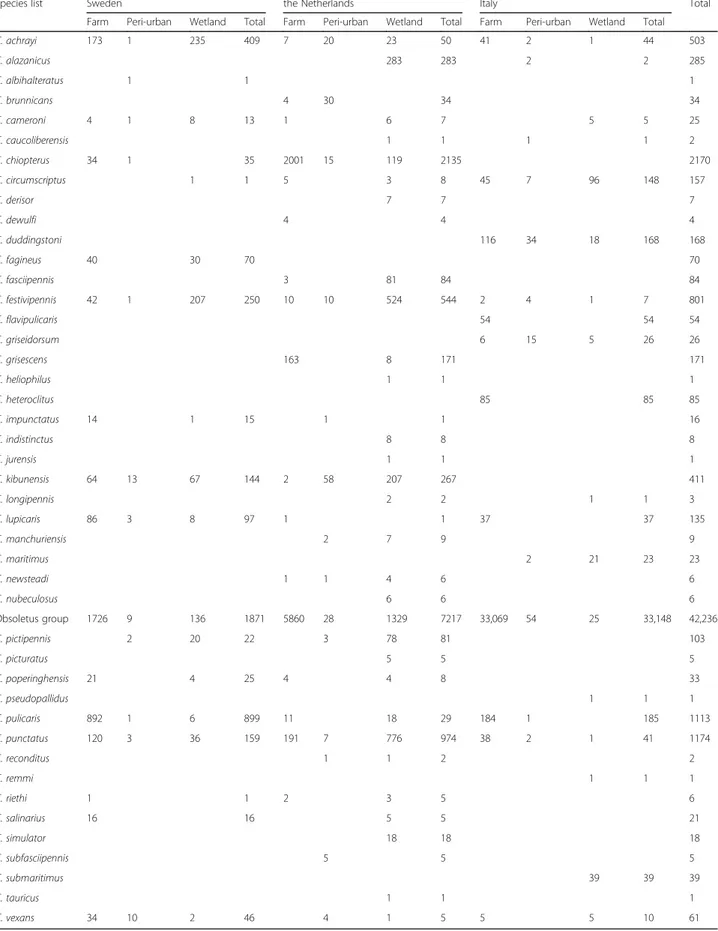
From the 4074 female midges trapped in Sweden, 18 species were identified. The most common species were from the Obsoletus group (47%), C. pulicaris (22%) and C. achrayi(Kettle & Lawson, 1955) (10%) (Table2). The dominating species among the 3,267 females trapped on farms were from the Obsoletus group (54%) and C. puli-caris(27%). From the 46 female midges trapped in peri-urban habitats, C. kibunensis (Tokunaga, 1937) (28%) and C. vexans (Staeger, 1839) (22%) were the most common, whereas the 761 specimens from wetlands in Sweden were dominated by C. achrayi (31%) and C. festivipennis(27%).
The Netherlands had the highest species richness with 35 species identified among 11,985 female midges trapped during the study period. The most common spe-cies were from the Obsoletus group (78%), C. punctatus
Table 1 Midge species diversity. Estimators of taxonomic diversity with values for the Simpson’s Index of Diversity, Shannon-Wiener diversity and Shannon-Wiener evenness for three habitats (farms, peri-urban and wetlands) in three countries (Sweden, the Netherlands and Italy)
Taxonomic diversity Sweden the Netherlands Italy Total
Farm Peri-urban Wetland Total Farm Peri-urban Wetland Total Farm Peri-urban Wetland Total
No. of specimens trapped 3267 46 761 4074 8270 185 3530 11,985 33,682 124 220 34,026 50,085
No. of samples 42 48 46 136 52 53 50 155 47 51 53 151 442
No. of species trapped 15 12 14 18 18 14 29 35 12 11 14 20 45
Simpson Index of Diversity 0.640 0.821 0.786 0.723 0.439 0.828 0.777 0.596 0.036 0.715 0.748 0.051 0.285
Shannon-Wiener diversity 1.439 2.000 1.801 1.714 0.807 2.051 1.863 1.386 0.127 1.589 1.750 0.178 0.813
(8%) and C. festivipennis (5%). For wetland and farm habitats the most abundant species were comprised of those species. The 185 midges trapped from peri-urban habitats were dominated by C. kibunensis (31%) and C. brunnicans(Edwards, 1939) (16%).
Of the 20 species trapped in Italy, the Obsoletus group was by far the most dominant with 97% of all 34,026 trapped female midges. The trapped midges from farm (33,682) and peri-urban (124) habitats were dominated by the Obsoletus group, although C. pulicaris (1%) and C. duddingstoni Kettle & Lawson, 1955 (27%) were also trapped more than other species in the two habitats, respectively. From the 220 midges trapped in Italian wetland habitats, C. circumscriptus Kieffer, 1918 (44%) and C. submaritimus Dzhafarov, 1962 (18%) were most abundant.
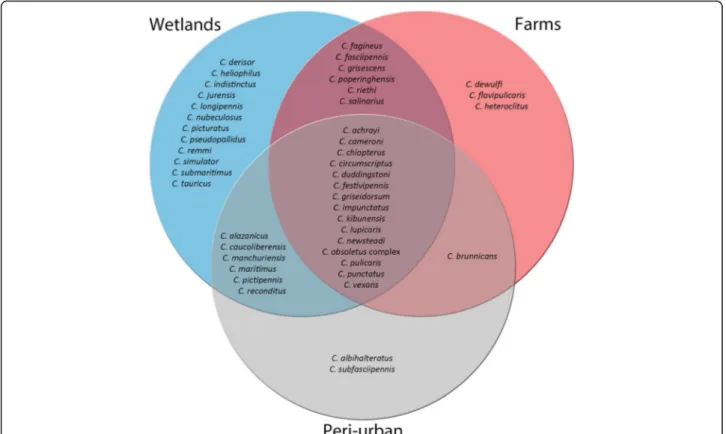
Of all midge species trapped, 38% (17/45) were unique to one of the three habitat types. Three species occurred exclusively at farms, two species in peri-urban habitats and 12 species only in wetland habitats (Fig.2). Figure3
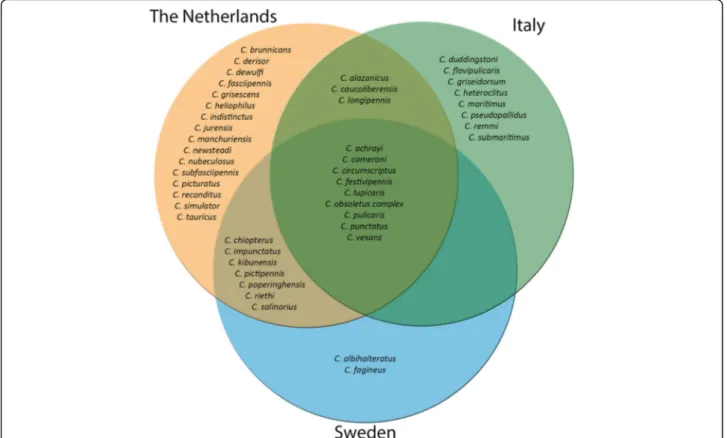
shows that more than half of the 45 species identified were trapped in only one of the countries (26/45, 58%), while 20% (9/45) of the species were trapped in all three countries. This core community included the most abundant species from the three countries: the Obsole-tus group, C. punctaObsole-tus, C. pulicaris, C. festivipennis and C. achrayi.
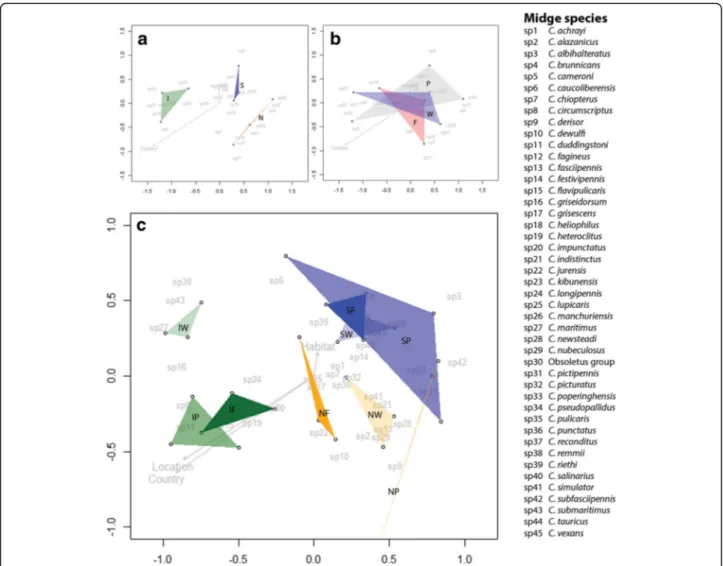
Combining the presence and abundance of different midge species trapped for the different countries and habitat types into one statistical analysis provides add-itional information about vector communities associated with specific regions. Dissimilarity matrices resulting from NMDS analyses show clear differences in biting midge community composition among the areas investigated in countries at different latitudes (P = 0.002,
stress = 0.084) (Fig. 4a). Differences in communities were not found among habitats (P = 0.976, stress = 0.084) (Fig.4b). However, looking at habitats within each coun-try, midge communities were found to be different for some of the habitats (Fig.4c). Midge communities among Dutch (P = 0.048, stress = 0.081) and Italian (P = 0.040, stress = 0.0594) habitats were significantly different from each other. For habitats in Italy this difference was mainly driven by the wetland habitat (Fig. 4c). Midge communities among habitats in Sweden were compar-able (P = 0.577, stress = 0.099).
Discussion
Biting midge community composition clearly differed among the areas we investigated at different latitudes. This is illustrated by the diversity indices (Table1), which was highest in Sweden, followed by the Netherlands, and then Italy. In addition, the Venn-diagram (Fig. 3) shows that 57% of the trapped midge species were found in only one of the countries. Finally, the dissimilarity matrix (Fig. 4a) distinguishes distinct midge communities among countries. Although communities varied among the areas investigated for the countries, a core community of midges seems to be present neverthe-less (Fig. 3). This core community includes the five most abundant species from the three countries: the Obsoletus group, C. punctatus, C. pulicaris, C. festivi-pennis and C. achrayi. While this core community oc-curs throughout Europe and across different habitats, it cannot be assumed that their contribution to disease spread is similar in all countries as temperature, inter-action with other (host) species, and genetic variation within midge species [4,40] also vary throughout Europe. However, these known midge vector species are present in a core community throughout Europe, and with rising temperature as a consequence of climate change, and continued increase in animal transport, it is expected that disease outbreaks will increase throughout Europe. In-creasing temperature will simultaneously affect the rapid increase of midge vector populations, and at the same time allow viruses to complete their extrinsic incubation period in vectors faster, which both imply an increased potential for pathogen transmission [26].
While community composition clearly differed among the sampled areas at different latitudes, communities were similar among habitat types (Fig. 4a, b). However, when differentiating habitats within countries, there were marked habitat effects on community composition (Fig. 4c). Habitat communities from farm, wetland, and peri-urban sites differed within the Netherlands and Italy, while communities in Sweden were more similar to each other. These results are comparable to diversity and com-munity composition found for mosquitoes in Europe [33]. Both mosquito and biting midge communities show clear
Fig. 1 Rarefaction plot of sampling effort. The plot shows the number of species expected to be found for the number of individuals sampled for Sweden (blue), Italy (green) and the Netherlands (orange)
Table 2 Midge species abundance. List of midge species with number of specimens for each country (Sweden, the Netherlands and Italy) and habitat type (farms, peri-urban and wetlands)
Species list Sweden the Netherlands Italy Total
Farm Peri-urban Wetland Total Farm Peri-urban Wetland Total Farm Peri-urban Wetland Total
C. achrayi 173 1 235 409 7 20 23 50 41 2 1 44 503 C. alazanicus 283 283 2 2 285 C. albihalteratus 1 1 1 C. brunnicans 4 30 34 34 C. cameroni 4 1 8 13 1 6 7 5 5 25 C. caucoliberensis 1 1 1 1 2 C. chiopterus 34 1 35 2001 15 119 2135 2170 C. circumscriptus 1 1 5 3 8 45 7 96 148 157 C. derisor 7 7 7 C. dewulfi 4 4 4 C. duddingstoni 116 34 18 168 168 C. fagineus 40 30 70 70 C. fasciipennis 3 81 84 84 C. festivipennis 42 1 207 250 10 10 524 544 2 4 1 7 801 C. flavipulicaris 54 54 54 C. griseidorsum 6 15 5 26 26 C. grisescens 163 8 171 171 C. heliophilus 1 1 1 C. heteroclitus 85 85 85 C. impunctatus 14 1 15 1 1 16 C. indistinctus 8 8 8 C. jurensis 1 1 1 C. kibunensis 64 13 67 144 2 58 207 267 411 C. longipennis 2 2 1 1 3 C. lupicaris 86 3 8 97 1 1 37 37 135 C. manchuriensis 2 7 9 9 C. maritimus 2 21 23 23 C. newsteadi 1 1 4 6 6 C. nubeculosus 6 6 6 Obsoletus group 1726 9 136 1871 5860 28 1329 7217 33,069 54 25 33,148 42,236 C. pictipennis 2 20 22 3 78 81 103 C. picturatus 5 5 5 C. poperinghensis 21 4 25 4 4 8 33 C. pseudopallidus 1 1 1 C. pulicaris 892 1 6 899 11 18 29 184 1 185 1113 C. punctatus 120 3 36 159 191 7 776 974 38 2 1 41 1174 C. reconditus 1 1 2 2 C. remmi 1 1 1 C. riethi 1 1 2 3 5 6 C. salinarius 16 16 5 5 21 C. simulator 18 18 18 C. subfasciipennis 5 5 5 C. submaritimus 39 39 39 C. tauricus 1 1 1 C. vexans 34 10 2 46 4 1 5 5 5 10 61
differences among areas in the three representative coun-tries, while these communities are different for habitats only within the countries studied. This suggests that local habitat factors can be important for vector community composition, but that ecological factors at large geograph-ical distances between sites have a more significant impact.
Some of the biting midge species that were morpho-logically identified, were thus far not known to be present in the studied countries. Of 18 midge species identified for Sweden, two species (C. cameroni and C. fagineus) could not be confirmed by the IIKC [2] or literature [17,41] (see Additional file 1: Table S1 for an overview). For the Netherlands, 12 species (C. brunnicans, C. cameroni, C. caucoliberensis, C. derisor, C. indistinctus, C. jurensis, C. longipennis, C. manchuriensis, C. picturatus, C. reconditus, C. simulator and C. tauricus) were not earlier described [10,42,43], and for Italy three species (C. achrayi, C. cameroni and C. vexans) were not found in literature [44, 45] or distribution maps of the IIKC. Because most of these species are known to be present in countries surrounding the countries studied here, we expect that the distribution is correct but was simply not confirmed before. We will continue to work with these samples and confirm the findings with barcoding techniques before adding them to current distribution lists.
From the European core midge community identified in this study, at least three species are (potential) vectors of pathogens. The Obsoletus group was the most abun-dant trapped in all countries, especially in farm habitats. Species in this group are known to transmit both BTV and SBV [6–11], and considered the most important midge vector species in Europe. However, the Obsoletus group consists of several species [C. chiopterus (Meigen, 1830); C. dewulfi Goetghebuer, 1936; C. obsoletus (Meigen, 1818) (s.s.); C. scoticus Downes & Kettle, 1952; and C. montanus Shakirzjanova, 1962] for which mor-phological identification is difficult and very laborious [41]. Culicoides identification remains a challenge, espe-cially for specific groups or complexes of species [3,36]. As new techniques such as DNA (barcode) sequencing and MALDI TOF (matrix assisted laser desorption/ion-isation time of flight) mass spectrometry [31,36,46,47] become available for identification, it will be easier to process large numbers of specimens. However, correct reference databases, morphological identifications, and the link with ecology remain essential components in Culicoidesresearch. Therefore, the European Interactive Identification Key for Culicoides (IKCC) developed by Mathieu et al. [2] is a useful tool to obtain accurate morphological identifications. Those species that are difficult or impossible to separate by morphological
Fig. 2 Venn diagram of habitats. Diagram shows the absolute presence of midge species found in farm (red), peri-urban (grey) and wetland (blue) habitats
identification, such as the Obsoletus group, can be further identified with molecular tools. Although species in the Obsoletus group are recognized as important potential vectors [48], it remains unknown what the species-specific (within the Obsoletus group) contribu-tion to pathogen transmission is. Priority should there-fore be given to investigate the Obsoletus group composition in more detail, to better understand disease dynamics.
The other two species found in this study that are (po-tential) vectors of pathogens were C. punctatus and C. pulicaris.These species were trapped in similar numbers and are known to transmit BTV and SBV [10,11,13–19]. Both species were found in all habitats and countries, al-though C. punctatus was mainly found in Dutch wetlands, while C. pulicaris was mostly present at farms in Sweden and Italy. One of the most important European BTV vectors for southern Europe, C. imicola, was not trapped during this study. Our trapping sites in Italy were further north compared to the known distribution of C. imicola [49], and results can therefore not be extrapolated for the most southern parts of Europe where C. imicola is present.
Although our sampling effort was comprehensive, as can be deduced from the rarefaction plot (Fig. 1), the study was carried out in a relatively limited area.
Culicoidesdiversity found in our trappings is, therefore, not representative for the countries as a whole. In addition, trapping with a single trap type may not accur-ately represent midge fauna diversity [42,50,51]. Never-theless, because of the consistent study design and use of the same trap type, results can be compared among the three areas in each country and habitats in this study. Midge species and their abundance can be under-or over-estimated compared to the biting rate on livestock animals [52]. Although mostly female biting midges were trapped, these were most likely not host-seeking midges, as they were attracted by a UV-light source [53]. The exact attraction mechanism for female midges towards the trap is thus far unknown. Nonetheless, the OVI trap seems to be the only effective midge trap currently available [54], and this stresses the need for appropriate monitoring methods against biting midges.
A study in Germany showed that adult midges can be trapped during winter months [16]. Although midge numbers captured in our traps were reduced in the months just before and after the winter period, it is not clear whether midges ceased their activity in our study. In addition to their activity during winter, the larval habitats of known midge vectors have not been exten-sively investigated [55–58]. With more knowledge on the habitats for both Culicoides larvae and adults, it will
Fig. 3 Venn diagram of countries. Diagram shows the absolute presence of midge species found in Sweden (blue), the Netherlands (orange) and Italy (green)
be possible to better understand the factors influen-cing differences in communities among countries and habitats.
Although the current study revealed differences in bit-ing midge communities among habitats within countries, the underlying factors for this were not identified. Biting midge species diversity was surprisingly high in peri-urban habitats (Table 1), possibly as only few specimens of different species were captured in comparable num-bers in this habitat type. In contrast, diversity was lowest at farms in all three countries, which could be explained by the high abundance of the Obsoletus group. Several of the farm collections had high abundance of midges and were therefore sub-sampled for identification. A few individuals of new species may be found if all individuals of farm samples would be identified. However, testing
this scenario by simulating additional species in our dataset did not change our conclusions on diversity indi-ces and the dominance of the Obsoletus group in farm habitats. The overwhelming abundance of this species group in farm habitats suppresses the influence on diversity measures of other species occurring on farms. Possibly, specific larval or adult habitats are present on farms that cause species from the Obsoletus group to flourish, while other species may not take advantage of these habitats.
Chaves et al. [59] suggested that lower diversity of vector communities is expected to increase the risk of amplification and spread of a vector-borne disease, because lower vector species diversity is thought to be correlated with higher relative abundance of some species within the community. This is in line with our
Fig. 4 Results of NMDS analyses. a Figure shows midge community compositions for Sweden (S), the Netherlands (N) and Italy (I). b NMDS analysis for farms (F), peri-urban (P) and, wetland (W) habitats based on number of midges trapped per species in each habitat and country. c NMDS analysis based on number of midges trapped per species at each location in each country and habitat (Sweden in blue: SF, SP and SW; the Netherlands in orange: NF, NP and NW; Italy in green: IF, IP and IW). The Bray-Curtis dissimilarity index was used to determine dissimilarities among midge community compositions. Stress value = 0.084 for panels a and b, which indicates a very good fit of the model. Stress value = 0.216 for panel c, which indicates a suspect fit of the model
findings, since the country (Italy) and habitat (farms) with the lowest diversity indices had the highest abun-dance of midges from the Obsoletus group. On the other hand, a theoretical study by Roche et al. [60] suggested that greater vector species richness leads to higher abun-dance, and can therefore amplify pathogen transmission. These authors showed that specific vector species in these complex community dynamics could be essential in epidemic take-off, even if these vectors are weakly susceptible to pathogen infection. Given that vector-borne diseases would require a set of multiple species that together influence the rate of transmission, under-standing the species composition of vector communities and their interactions with pathogens is becoming in-creasingly important. Ultimately, entomological field data should be used for the development of mathemat-ical and statistmathemat-ical models, to more accurately assess the effect of environmental factors on midge population dynamics and how this influences disease risks.
Conclusions
A core European midge community could be identified, with important disease vector species from the Obsoletus group, C. punctatus and C. pulicaris, as the most abun-dant in this core community. The presence of a core com-munity throughout Europe suggests that disease dynamics can be homogenous, since the core community is present in all countries and habitats. However, in-depth analysis of the complete midge community composition showed that differences were more distinct among countries than habi-tat types. In other words, although some midge species are found throughout Europe, distinctive communities can be found within each country. This suggests that specific species within countries may impose a more heterogeneous disease dynamics than expected when only looking at the core community. Although we have eluci-dated how species communities differ among countries and habitats, it is still unclear how these specific species and their associated communities affect disease risk.
Additional files
Additional file 1:Table S1. A list of the species per country found in this study. References to previous research of Culicoides fauna in the same countries are made, as well as remarks on the distribution of the species as described in the IIKC. (XLSX 14 kb)
Abbreviations
AHSV:African horse sickness virus; BTV: Bluetongue virus;
CORINE: Coordination of information on the environment; IIKC: Interactive Identification Key for Culicoides; NMDS: Non-metric multidimensional scaling; OVI: Onderstepoort Veterinary Institute; SBV: Schmallenberg virus
Acknowledgements
We would like to thank the students that helped collecting the field data; Rickard Pettersson from Sweden and Mahmod Hamdan from Italy. Farmers, nature area managers and home-owners in Sweden, the Netherlands and Italy that provided us with the essential locations needed for this research are acknowledged for their support. We also thank Marcel Dicke for provid-ing comments on a previous version of this manuscript.
Funding
This study was part of the ANIHWA-EraNet (Animal Health and Welfare) pro-gram (LIVEepi) funded by the Ministry of Economic Affairs (project number 1300018161), the Netherlands, and the Swedish Research Council (FORMAS), Sweden. In Italy, the work was supported by grants from the Italian Ministry of Education, University and Research (MIUR) (Prin 2012 protocol
2012T85B3R). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Availability of data and materials
The dataset supporting the conclusions of this article is included within the article and its additional file.
Authors’ contributions
TWRM, UW, GF, WT and CJMK designed the study. TWRM, MT, CD and LB collected data. UW, GF, WT and CJMK provided funding and support for fieldwork. TWRM did morphological identifications, analysed data and drafted the manuscript. UW, MT, GF, CD, LB, WT and CJMK contributed to writing of the manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to participate Not applicable.
Consent for publications Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Author details 1
Laboratory of Entomology, Wageningen University and Research, P.O. Box 16, 6700, AA, Wageningen, the Netherlands.2IFM Theory and Modelling, Linköping University, 581 83 Linköping, Sweden.3Scuola di Bioscienze e Medicina Veterinaria, Università degli Studi di Camerino, 62032 Camerino, Italy.
Received: 27 September 2017 Accepted: 11 March 2018
References
1. Mehlhorn H, Walldorf V, Klimpel S, Jahn B, Jaeger F, Eschweiler J, et al. First occurrence of Culicoides obsoletus - transmitted bluetongue virus epidemic in central Europe. Parasitol Res. 2007;101(1):219–28.
2. Mathieu B, Cêtre-Sossah C, Garros C, Chavernac D, Balenghien T, Vignes Lebbe R, et al. IIKC: An interactive identification key for female Culicoides (Diptera: Ceratopogonidae) from the West Palearctic region. Proceedings of the international congress Tools for Identifying Biodiversity: Progress and Problems: 20–22 September 2010; Paris. Vignes-Lebbe R: Nimis PL; 2010. p. 201–5.
3. Mathieu B, Cetre-Sossah C, Garros C, Chavernac D, Balenghien T, Carpenter S, et al. Development and validation of IIKC: an interactive identification key for Culicoides (Diptera: Ceratopogonidae) females from the Western Palaearctic region. Parasit Vectors. 2012;5:137.
4. Mellor P, Boorman J, Baylis M. Culicoides biting midges: their role as arbovirus vectors. Annu Rev Entomol. 2000;45(1):307–40.
5. Carpenter S, Lunt HL, Arav D, Venter GJ, Mellor PS. Oral susceptibility to bluetongue virus of Culicoides (Diptera: Ceratopogonidae) from the United Kingdom. J Med Entomol. 2006;43(1):73–8.
6. De Liberato C, Scavia G, Lorenzetti R, Scaramozzino P, Amaddeo D, Cardeti G, et al. Identification of Culicoides obsoletus (Diptera: Ceratopogonidae) as a vector of bluetongue virus in central Italy. Vet Rec. 2005;156(10):301–4.
7. Savini G, Goffredo M, Monaco F, Di Gennaro A, Cafiero MA, Baldi L, et al. Bluetongue virus isolations from midges belonging to the Obsoletus complex (Culicoides, Diptera: Ceratopogonidae) in Italy. Vet Rec. 2005;157(5):133–9.
8. Carpenter S, Szmaragd C, Barber J, Labuschagne K, Gubbins S, Mellor P. An assessment of Culicoides surveillance techniques in northern Europe: have we underestimated a potential bluetongue virus vector? J Appl Ecol. 2008;45(4):1237–45.
9. Elbers A, Meiswinkel R, van Weezep E. Sloet van Oldruitenborgh-Oosterbaan M, Kooi E. Schmallenberg virus detected by RT-PCR in Culicoides biting midges captured during the 2011 epidemic in the Netherlands. Emerg Infect Dis. 2013;19:106–9.
10. Meiswinkel R, Scolamacchia F, Dik M, Mudde J, Dijkstra E, Van Der Ven IJ, et al. The Mondrian matrix: Culicoides biting midge abundance and seasonal incidence during the 2006–2008 epidemic of bluetongue in the Netherlands. Med Vet Entomol. 2014;28(1):10–20.
11. Koenraadt CJ, Balenghien T, Carpenter S, Ducheyne E, Elbers AR, et al. Bluetongue, Schmallenberg - what is next? Culicoides-borne viral diseases in the 21st Century. BMC Vet Res. 2014;10:77.
12. Goffredo M, Meiswinkel R. Entomological surveillance of bluetongue in Italy: methods of capture, catch analysis and identification of Culicoides biting midges. Vet Ital. 2003;40(3):260–5.
13. Purse B, Tatem A, Caracappa S, Rogers D, Mellor P, Baylis M, et al. Modelling the distributions of Culicoides bluetongue virus vectors in Sicily in relation to satellite-derived climate variables. Med Vet Entomol. 2004;18(2):90–101. 14. Takken W, Knols BG. Emerging pests and vector-borne diseases in Europe,
vol. 1. Wageningen: Wageningen Academic Publishers; 2007.
15. Carpenter S, Wilson A, Mellor P. Bluetongue virus and Culicoides in the UK: the impact of research on policy. Outlooks Pest Manag. 2009;20(4):161–4. 16. Hoffmann B, Bauer B, Bauer C, Batza HJ, Beer M, Clausen PH, et al.
Monitoring of putative vectors of bluetongue virus serotype 8, Germany. Emerg Infect Dis. 2009;15(9):1481–4.
17. Ander M, Meiswinkel R, Chirico J. Seasonal dynamics of biting midges (Diptera: Ceratopogonidae: Culicoides), the potential vectors of bluetongue virus, in Sweden. Vet Parasitol. 2012;184(1):59–67.
18. Wilson AJ, Mellor PS. Bluetongue in Europe: past, present and future. Philos Trans R Soc Lond, Ser B: Biol Sci. 2009;364(1530):2669–81.
19. Balenghien T, Pagès N, Goffredo M, Carpenter S, Augot D, Jacquier E. The emergence of Schmallenberg virus across Culicoides communities and ecosystems in Europe. Prev Vet Med. 2014;116(4):360–9.
20. World Organisation for Animal Health. OIE-Listed diseases, infections and infestations in force in 2017. 2017. http://www.oie.int/en/animal-health-in-the- world/oie-listed-diseases-2017/. Accessed 08 Aug 2017.
21. Wernike K, Eschbaumer M, Breithaupt A, Hoffmann B, Beer M.
Schmallenberg virus challenge models in cattle: infectious serum or culture-grown virus? Vet Res. 2012;43(1):1.
22. Gubbins S, Richardson J, Baylis M, Wilson AJ, Abrahantes JC. Modelling the continental-scale spread of Schmallenberg virus in Europe: approaches and challenges. Prev Vet Med. 2014;116(4):404–11.
23. Veldhuis A, Santman-Berends I, Gethmann J, Mars M, Van Wuyckhuise L, Vellema P, et al. Schmallenberg virus epidemic: impact on milk production, reproductive performance and mortality in dairy cattle in the Netherlands and Kleve district, Germany. Prev Vet Med. 2014;116(4):412–22. 24. Van Schaik G, Berends I, Hv L, Elbers A, Vellema P. Seroprevalence of
bluetongue serotype 8 in cattle in the Netherlands in spring 2007, and its consequences. Vet Rec. 2008;163(15):441–4.
25. Saegerman C, Martinelle L, Dal Pozzo F, Kirschvink N. Preliminary survey on the impact of Schmallenberg virus on sheep flocks in south of Belgium. Transbound Emerg Dis. 2014;61(5):469–72.
26. Wilson A, Mellor P. Bluetongue in Europe: vectors, epidemiology and climate change. Parasitol Res. 2008;103(1):69–77.
27. Kiel E, Liebisch G, Focke R, Liebisch A. Monitoring of Culicoides at 20 locations in northwest Germany. Parasitol Res. 2009;105(2):351–7. 28. Venail R, Balenghien T, Guis H, Tran A, Setier-Rio ML, Delécolle JC, et al.
Assessing diversity and abundance of vector populations at a national scale: example of Culicoides surveillance in France after bluetongue virus emergence. Arthropods Vector Emerg Dis. 2012;3:77–102.
29. Fassotte C, Delecolle JC, Cors R, Defrance T, De Deken R, Haubruge E, et al. Culicoides trapping with Rothamsted suction traps before and during the bluetongue epidemic of 2006 in Belgium. Prev Vet Med. 2008;87(1–2):74–83. 30. González M, Baldet T, Delécolle JC, López S, Romón P, Goldarazena A.
Monitoring of Culicoides Latreille (Diptera: Ceratopogonidae) after BTV outbreaks, in sheep farms and natural habitats from the Basque Country (Northern Spain). Proc Entomol Soc Wash. 2013;115(1):48–69. 31. Kaufmann C, Steinmann IC, Hegglin D, Schaffner F, Mathis A.
Spatio-temporal occurrence of Culicoides biting midges in the climatic regions of Switzerland, along with large scale species identification by MALDI-TOF mass spectrometry. Parasit Vectors. 2012;5:246.
32. Venter GJ, Meiswinkel R. The virtual absence of Culicoides imicola (Diptera: Ceratopogonidae) in a light-trap survey of the colder, high-lying area of the eastern Orange Free State, South Africa, and implications for the transmission of arboviruses. Onderstepoort J Vet Res. 1994;61(4):327–40. 33. Möhlmann TWR, Wennergren U, Tälle M, Favia G, Damiani C, Bracchetti L, et
al. Community analysis on the abundance and diversity of mosquito species (Diptera: Culicidae) in three European countries at different latitudes. Parasit Vectors. 2017;10:510.
34. EEA. Corine Land Cover. Copenhagen: Off Pub Euro Comm; 2000. 35. Nolan DV, Carpenter S, Barber J, Mellor PS, Dallas JF, Mordue AJ, et al. Rapid
diagnostic PCR assays for members of the Culicoides obsoletus and Culicoides pulicaris species complexes, implicated vectors of bluetongue virus in Europe. Vet Microbiol. 2007;124(1):82–94.
36. Harrup L, Bellis G, Balenghien T, Garros C. Culicoides Latreille (Diptera: Ceratopogonidae) taxonomy: Current challenges and future directions. Infect Genet Evol. 2015;30:249–66.
37. Oksanen J, Kindt R, Legendre P, O’Hara B, Simpson G, Stevens M, et al. Vegan: Community Ecology Package. R package version 2.9. 2009:2. http://CRAN.R-project.org/package=vegan
38. Team RC. R: A language and environment for statistical computing. Vienna: R Foundation for Statistical Computing; 2015.
39. Young M, Scannell J, O'Neill M, Hilgetag C, Burns G, Blakemore C. Non-metric multidimensional scaling in the analysis of neuroanatomical connection data and the organization of the primate cortical visual system. Philos Trans R Soc Lond, Ser B: Biol Sci. 1995;348(1325):281–308.
40. Jones R, Foster N. Oral infection of Culicoides variipennis with bluetongue virus: development of susceptible and resistant lines from a colony population. J Med Entomol. 1974;11(3):1.
41. Nielsen SA, Kristensen M. Morphological and molecular identification of species of the Obsoletus group (Diptera: Ceratopogonideae) in Scandinavia. Parasitol Res. 2011;109(4):1133–41.
42. Elbers AR, Meiswinkel R. Culicoides (Diptera: Ceratopogonidae) host preferences and biting rates in the Netherlands: comparing cattle, sheep and the black-light suction trap. Vet Parasitol. 2014;205(1–2):330–7. 43. Naturalis Biodiversity Center.www.nederlandsesoorten.nl. Accessed 2 Jan
2018.
44. Foxi C, Pinna M, Monteys VSI, Delrio G. An updated checklist of the Culicoides Latreille (Diptera: Ceratopogonidae) of Sardinia (Italy), and seasonality in proven and potential vectors for bluetongue virus (BTV). Proc Entomol Soc Wash. 2011;113(4):403–16.
45. Gomulski L, Meiswinkel R, Delécolle JC, Goffredo M, Gasperi G. Phylogeny of the subgenus Culicoides and related species in Italy, inferred from internal transcribed spacer 2 ribosomal DNA sequences. Med Vet Entomol. 2006;20(2):229–38.
46. Kaufmann C, Schaffner F, Ziegler D, Pflueger V, Mathis A. Identification of field-caught Culicoides biting midges using matrix-assisted laser desorption/ ionization time of flight mass spectrometry. Parasitology. 2012;139(2):248–58. 47. Kaufmann C, Ziegler D, Schaffner F, Carpenter S, Pflüger V, Mathis A.
Evaluation of matrix-assisted laser desorption/ionization time of flight mass spectrometry for characterization of Culicoides nubeculosus biting midges. Med Vet Entomol. 2011;25(1):32–8.
48. Gomulski LM, Meiswinkel R, Delécolle J-C, Goffredo M, Gasperi G. Phylogenetic relationships of the subgenus Avaritia Fox, 1955 including Culicoides obsoletus (Diptera, Ceratopogonidae) in Italy based on internal transcribed spacer 2 ribosomal DNA sequences. Syst Entomol. 2005;30(4):619–31.
49. Calistri P, Goffredo M, Caporale V, Meiswinkel R. The distribution of Culicoides imicola in Italy: application and evaluation of current Mediterranean models based on climate. Zoonoses Public Hlth. 2003;50(3):132–8.
50. Elbers A, Heuvel SJ, Meiswinkel R. Diel activity and preferred landing sites in Culicoides biting midges attacking Fjord horses. Entomol Exp Appl. 2016;160(3):272–80.
51. Meiswinkel R, Elbers A. The dying of the light: crepuscular activity in Culicoides and impact on light trap efficacy at temperate latitudes. Med Vet Entomol. 2016;30(1):53–63.
52. Viennet E, Garros C, Lancelot R, Allene X, Gardes L, Rakotoarivony I, et al. Assessment of vector/host contact: comparison of animal-baited traps and UV-light/suction trap for collecting Culicoides biting midges (Diptera: Ceratopogonidae), vectors of Orbiviruses. Parasit Vectors. 2011;4:119. 53. Elbers A, Meiswinkel R. Limited attractant range of the black-light suction
trap for the capture of Culicoides biting midges (Diptera: Ceratopogonidae). J Appl Entomol. 2016;140(5):386–94.
54. Probst C, Gethmann JM, Kampen H, Werner D, Conraths FJ. A comparison of four light traps for collecting Culicoides biting midges. Parasitol Res. 2015;114(12):4717–24.
55. Foxi C, Delrio G. Larval habitats and seasonal abundance of Culicoides biting midges found in association with sheep in northern Sardinia, Italy. Med Vet Entomol. 2010;24(2):199–209.
56. Zimmer J-Y, Saegerman C, Losson B, Beckers Y, Haubruge E, Francis F. Chemical composition of silage residues sustaining the larval development of the Culicoides obsoletus/Culicoides scoticus species (Diptera:
Ceratopogonidae). Vet Parasitol. 2013;191(1):197–201.
57. Zimmer JY, Brostaux Y, Haubruge E, Francis F. Larval development sites of the main Culicoides species (Diptera: Ceratopogonidae) in northern Europe and distribution of coprophilic species larvae in Belgian pastures. Vet Parasitol. 2014;205(3–4):676–86.
58. Harrup L, Purse B, Golding N, Mellor P, Carpenter S. Larval development and emergence sites of farm-associated Culicoides in the United Kingdom. Med Vet Entomol. 2013;27(4):441–9.
59. Chaves LF, Hamer GL, Walker ED, Brown WM, Ruiz MO, Kitron UD. Climatic variability and landscape heterogeneity impact urban mosquito diversity and vector abundance and infection. Ecosphere. 2011;2(6):70. 60. Roche B, Rohani P, Dobson AP, Guégan J-F. The impact of community
organization on vector-borne pathogens. Am Nat. 2013;181(1):1–11.
• We accept pre-submission inquiries
• Our selector tool helps you to find the most relevant journal • We provide round the clock customer support
• Convenient online submission • Thorough peer review
• Inclusion in PubMed and all major indexing services • Maximum visibility for your research
Submit your manuscript at www.biomedcentral.com/submit