UNIVERSITA’ DEGLI STUDI DI PISA
FACOLTA’ DI SCIENZE MATEMATICHE, FISICHE E NATURALI
Corso di Laurea Specialistica in Biologia Marina
Anno accademico 2008-2009
TESI DI LAUREA SPECIALISTICA
Le praterie emergenti di barriera di Posidonia oceanica e la loro regressione: il caso di studio di una prateria della Toscana meridionale (S. Liberata, Orbetello, GR)
Candidata: Letizia Barsellini
Relatore:
Prof. Alberto Castelli
Correlatore: Dott. Mauro Lenzi
1 Ai miei genitori, con immensa stima
2 Indice 1. Riassunto ... 3 2. Abstract ... 5 3. Introduzione ... 7 3.1. Posidonia oceanica ... 7
3.2. Studio delle praterie di P. oceanica ... 10
3.3. Le praterie emergenti: barrier reef ... 16
3.4. La regressione delle praterie di P. oceanica nel Mar Mediterraneo ... 16
5. Materiali e Metodi ... 24 5.1. Area di Studio ... 24 5.2 Disegno di campionamento ... 25 5.2.1 Balisages ... 25 5.2.2. La prateria di P. oceanica ... 26 5.2.3. Contaminanti ... 27
5.2.4. Caratteristiche trofiche dell’ecosistema e tessitura sedimentaria ... 31
5.3. Analisi statistica ... 35
6. Risultati ... 41
6.1. La prateria di P. oceanica ... 41
6.2. Granulometria ... 42
6.3. Carbonio organico e Carbonio, Azoto e Fosforo Totali ... 43
6.4. Materia organica ... 45
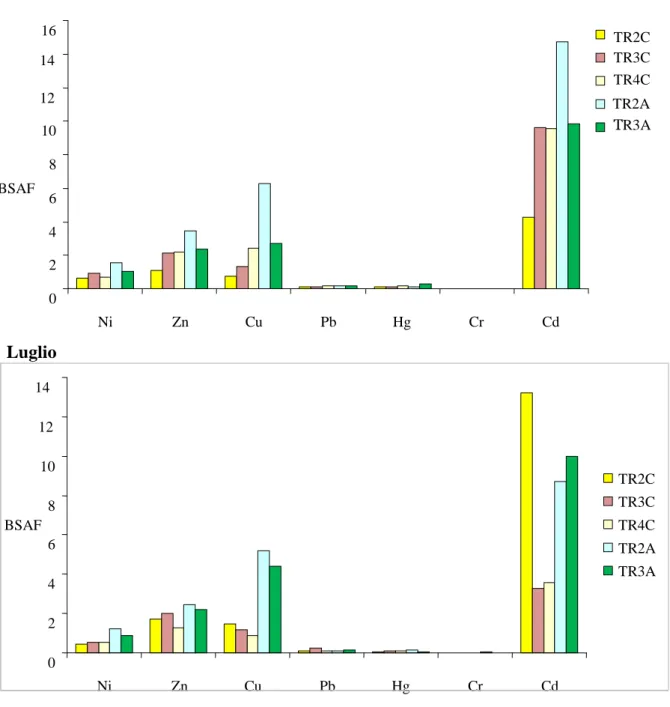
6.5. Elementi in tracce ... 46
6.6. Idrocarburi Policiclici Aromatici (IPA) ... 48
7. Discussione ... 76
8. Conclusioni ... 91
Letteratura Citata ... 93
3 1. Riassunto
Le praterie emergenti di barriera di Posidonia oceanica e la loro regressione: il caso di studio di una prateria della Toscana meridionale (S. Liberata, Orbetello, GR)
Le praterie di Posidonia oceanica costituiscono un ecosistema complesso e diffuso delle aree costiere del Mediterraneo, giocando un ruolo fondamentale nell’ecologia della zona neritica. Queste praterie risultano molto sensibili al disturbo prodotto dalle attività umane e possono essere utilizzate quali indicatori della qualità delle aree in cui sono insediate. A partire dagli anni ’70, esse hanno subito una regressione in tutto il Mediterraneo, le cui cause vanno cercate sia nella crescente pressione antropica sulla fascia costiera, che in fattori naturali.
Questa tesi si avvale di uno studio in campo di una prateria emergente della Toscana meridionale (S. Liberata, Orbetello, GR), che costituisce una barriera costiera, determinando una riduzione dell’idrodinamismo nelle aree retrostanti, prossime alla costa. Le barriere naturali emergenti di P. oceanica sono formazioni di elevata importanza naturalistica. Esse determinano peculiari caratteristiche ambientali nelle aree retrostanti, le quali si arricchiscono in materia organica e consentono lo sviluppo di insediamenti fitobentonici tipici.
L’obiettivo principale dello studio è stato quello di verificare lo stato di salute della prateria in oggetto, in rapporto agli stress a cui è stata finora sottoposta. Pertanto è stata condotta una indagine ambientale al fine di descrivere la prateria attraverso i parametri biometrici, densità e biomassa, di verificare la presenza di contaminanti nelle diverse componenti dell’ecosistema (alghe, angiosperme, sedimenti) e di definire lo stato di trofia attraverso la determinazione di C, N, P in P. oceanica, macroalghe e sedimento, e, per quest’ultimo, di materia organica e materia sedimentabile.
La prateria è risultata in espansione verso il largo con la tendenza ad innalzarsi nella parte centrale; quest’ultimo fatto ha comportato un’ulteriore riduzione dell’idrodinamismo e quindi del ricambio delle masse d’acqua, causando probabilmente una sofferenza della prateria vicina alla costa. Quest’ultima è in rarefazione con numerose chiazze a matte morte. Tuttavia, la carenza di materia organica nell’area di retro-prateria ed una dinamica sedimentaria che non ne consente l’accumulo, contrariamente al passato, ne precludono la presenza della tipica prateria mista Cymodocea nodosa - Nanozostera noltii - Caulerpa
4 prolifera. Secondo studi pregressi, questo fenomeno sarebbe riconducibile all’incremento della balneazione.
Quest’area è inoltre soggetta all’immissione di IPA che si rilevano sia nei sedimenti che nelle foglie di Posidonia, con concentrazioni superiori al livello di “background”, che fanno variare la contaminazione tra moderata e molto alta. Questi contaminanti sono attribuibili per lo più a processi pirolitici, provenienti probabilmente dal vicino insediamento portuale, che presenta un traffico di natanti che è andato via via intensificandosi negli ultimi trent’anni. Al contrario, i livelli dei metalli nei vari comparti ambientali, ad eccezione del mercurio per cui esiste una anomalia geologica naturale nell’area, sono fra i più bassi tra quelli riportati in letteratura per aree costiere.
In conclusione, questa prateria risulta ancora vitale e dinamica, per quanto in flessione della densità fogliare rispetto agli anni ’80, ma sta subendo una molteplice pressione antropica, soprattutto da parte di contaminanti IPA, e da parte del disturbo sedimentario prodotto dalla balneazione.
5 2. Abstract
The emerging meadows of Posidonia oceanica barrier and their regression: a case study of a meadow in the Southern Tuscany area (S. Liberata, Orbetello, GR)
The meadows of Posidonia oceanica are a complex and widespread ecosystem along the Mediterranean coastline, playing a key role in the neritic zone ecology. These meadows are very sensitive to human activities and can be used as indicators of its native ecosystem. Since the '70s, they have suffered from a regression in the whole Mediterranean area, caused by the increasing human pressure on the coastline, and natural processes.
This thesis is based on a field study carried out an emerging meadow located in the Southern Tuscany area (S. Liberata, Orbetello, GR). This meadow is a coastal barrier-reef that leads to a reduction of hydrodynamics in the back-reef areas, near the coast. Natural barriers of emerging P. oceanica meadows have a high naturalistic importance. They determine specific environmental characteristics in the back-reef areas, which are enriched in organic matter and allow the development of typical phytobenthic settlements.
This study mainly aims at verifying the condition of P. oceanica meadow located in the area, in relation to the stresses which it has been subjected to so far. Therefore an environmental survey was carried out in order to describe the meadow through biometric parameters, density and biomass, to verify the presence of contaminants in different parts of the ecosystem (algae, angiosperms, sediments) and to define the trophic state (C, N and P content in P. oceanica, macroalgae and sediment), and organic matter in the sediment and sedimentation degree.
It has turned out that the meadow was slowly spreading offshore, and particularly rising in the central part. This caused a further reduction of hydrodynamism and water exchange, that is probably the cause of the rarefaction in the part of the meadow nearest the coastline; in fact there were several dead matte patches. However, the poor presence of organic matter in the back-reef area, and a sedimentation dynamics that prevents the accumulation, probably did not allow the typical mixed meadow of Cymodocea nodosa - Nanozostera noltii - Caulerpa prolifera to settle.
Moreover it has been found the presence of PAHs in both, sediment and leaves of Posidonia, whose concentrations were above the background level and contamination ranged
6 from moderate to very high. Such contamination was mainly due to pyrolytic processes, caused by the nearby marina, in which the traffic of boats has been increasing over the years. On the contrary, levels of metals in biota and sediment are among the lowest of those reported in the literature for coastal areas, except mercury for which there is a natural geological anomaly in the area.
In conclusion, this meadow is still vital and dynamic, but is undergoing different human impacts: contamination by PAHs and sediment disturbance that alters the dynamics of inshore sedimentation.
7 3. Introduzione
3.1. Posidonia oceanica
Posidonia oceanica (L.) Delile è una fanerogama marina endemica del Mediterraneo appartenente al subphylum Angiospermae, classe Monocotyledon, ordine Alismatales, famiglia Posidoniaceae (APG II, 2003).
Le fanerogame marine si distinguono dalle alghe per la riproduzione sessuale, in quanto possiedono organi specializzati ben visibili (phaneros in greco significa “manifesto” e gamos “nozze”) e perché costituite da radici, fusto (detto “rizoma” a causa del suo habitus ipogeo) e foglie.
I rizomi di P. oceanica sono fusti modificati che presentano la caratteristica di accrescersi sia in senso orizzontale (rizoma plagiotropo o tracciante) che verticale (rizoma ortotropo). I rizomi plagiotropi sono i primi a colonizzare i sedimenti ed hanno la funzione di ancorare la pianta al substrato, grazie alla presenza di radici sul lato inferiore; i rizomi ortotropi, crescendo in altezza, contrastano il progressivo insabbiamento dovuto al continuo processo di sedimentazione e permettono in tal modo di sfruttare la luce e lo spazio disponibili (Mazzella et al., 1986). Questi due tipi di accrescimento sono all’origine della formazione delle cosiddette “mattes”, tipiche strutture a terrazzo costituite dall’intreccio di più strati di rizomi, di radici e di sedimento intrappolato tra questi (Fig. 1). Solo la sommità di questa formazione è ricoperta da piante vive (Mazzella et al., 1986), mentre gli interstizi sono riempiti dai sedimenti intrappolati nella volta fogliare (Boudouresque et al., 1984). La velocità di innalzamento delle matte è stata stimata in circa 1 m al secolo (Molinier & Picard, 1952).
Le foglie nascono dai rizomi ortotropi, sono nastriformi, con apice arrotondato, di colore verde brillante, con una larghezza in media di 1 cm e possono superare anche 1 m di lunghezza (Buia et al., 1986). Nella parte superiore del rizoma è situato l'apice vegetativo da cui si originano le foglie che sono organizzate in ciuffi; ogni ciuffo contiene in media 6-7 foglie che si distinguono in adulte, intermedie e giovanili (Fig. 2a). Le foglie adulte sono costituite dalla base, che inserisce la foglia al rizoma e dal lembo (lamina), che è la parte fotosintetizzante. Le foglie intermedie, senza base, hanno una lunghezza maggiore di 5 cm mentre le giovanili, anch'esse senza base, hanno una lunghezza inferiore a 5 cm (Giraud, 1979). L’accrescimento si origina da un meristema basale anziché apicale; tale adattamento
8 permette la crescita della lamina fogliare anche quando l’apice, che ne diviene la parte più vecchia, va incontro per primo a fenomeni di degenerazione (Mazzella et al., 1986). Le foglie più giovani e più corte sono interne, mentre le foglie più vecchie sono più esterne. Con il sopraggiungere della stagione autunnale, si assiste ad una produzione di nuove foglie giovanili e al distacco a livello della ligula (linea concava situata tra la base e il lembo) delle foglie adulte, che però lasciano la base attaccata al rizoma, conferendogli una struttura a scaglie. Le basi, o scaglie, grazie al loro maggior contenuto di lignina, rimangono fissate al rizoma e circondandolo con una particolare struttura protettiva detta “ manicotto” (Fig. 2a).
La pianta si riproduce sia sessualmente con fiori e frutti, che asessualmente per stolonizzazione. I fiori detti “spighe” (Fig. 2b), a causa del loro aspetto, sono ermafroditi, raggruppati (da 4 a 10) in un’infiorescenza avvolta da due brattee floreali e provvista di un peduncolo inserito al centro del ciuffo di foglie. La fioritura si verifica verso Settembre-Ottobre nelle praterie superficiali (fino a 15 m di profondità) e verso Novembre in quelle più profonde (Mazzella et al., 1986). Dopo l’impollinazione, l’ovario fecondato porta a maturazione un frutto chiamato “oliva di mare” (Fig. 2c) che raggiunge la maturazione nei mesi di Marzo-Aprile. Il frutto giunto a maturazione si stacca dalla pianta madre e galleggiando alcune settimane grazie alle sostanze oleose presenti nel pericarpo, viene trasportato dalle correnti anche per lunghe distanze. Alcune “olive di mare” finiscono spiaggiate lungo le coste, mentre quelle rimaste in mare, con la deiscenza del pericarpo, lasciano cadere i semi sul fondo: in presenza di adeguate condizioni chimico fisiche e del substrato si ha la germinazione.
Quando P. oceanica incontra condizioni ambientali favorevoli, colonizza ampie aree di fondo marino formando praterie, che si estendono dalla superficie fino a 30 - 35 metri di profondità, spingendosi fino a 40 metri in acque molto limpide; la sua crescita è infatti strettamente condizionata dalla presenza di luce.
La pianta necessita di una forte illuminazione e per questo la trasparenza dell’acqua e la temperatura sono fattori che ne limitano lo sviluppo; può vivere entro un ampio intervallo di temperatura (tra 10° C e 28° C), tollerando sbalzi di temperatura relativamente importanti (Boudouresque & Meinesz, 1982).
Diverso è il comportamento rispetto alla salinità: P. oceanica è stenoalina, per cui non si insedia in prossimità degli sbocchi fluviali ed è assente nelle aree salmastre.
9 la densità, che corrisponde al numero di foglie per unità di superficie pari a 1 m2 in relazione alla densità dei fasci fogliari, si possono distinguere praterie molto dense, dense, rade, molto rade e semipraterie (Giraud, 1977). La densità è generalmente più elevata nelle praterie superficiali e va progressivamente diminuendo in profondità. Per quanto riguarda la distribuzione di una prateria a P. oceanica, si distingue un “limite superiore” ed un “limite inferiore”: il primo è il punto in cui ha inizio la prateria partendo dalla linea di costa, ed è sempre molto netto, mentre il secondo è il punto di maggiore profondità a cui termina la prateria, il quale può avere conformazioni diverse.
P. oceanica riveste un ruolo fondamentale nell’economia generale delle aree costiere di fondo mobile per l’equilibrio e la ricchezza sia a livello biologico che fisico.
In quanto organismo vegetale, P. oceanica contribuisce in maniera cospicua all’ossigenazione delle acque, grazie alla sua attività fotosintetica: Bay (1978) stimò che 1 m² di prateria produce giornalmente da 4 a 20 litri di ossigeno.
Le praterie di P. oceanica rivestono una notevole importanza per l’elevatissima diversità ecologica di questa comunità, costituendo una sorta di “nursery” per gli avannotti dei pesci e allo stesso tempo rappresenta sia una zona di riproduzione, di deposizione, di riparo dai predatori che una fonte di cibo per molti pesci, cefalopodi e crostacei. Molti organismi utilizzano foglie e rizomi come substrato (es., epifite) e molti altri trovano rifugio all’interno della “matte” (meiobenthos), così che la prateria costituisce un complesso ecosistema.
La grande quantità di sostanza organica prodotta (può raggiungere i 20000 Kg/ha per anno di sostanza organica secca) costituisce una fonte di cibo diretta e indiretta per numerosi organismi ed è il punto di partenza di una complessa rete trofica, che coinvolge sia la catena del detrito che quella del pascolo.
Infine P. oceanica riveste un ruolo di primaria importanza nella stabilizzazione di rive e fondali. Nel primo caso gli accumuli sulla spiaggia di foglie morte della fanerogama dette dai francesi “les banquettes” riducono l’azione erosiva del moto ondoso sul litorale, assorbendo l’energia che si libera al frangivento dell’onda. E’ stato stimato che la regressione di un solo metro di prateria può provocare la perdita di 15-18 m di litorale sabbioso (Mazzella et al., 1986). Nel secondo caso l’apparato radicale e stolonifero della pianta garantisce un effetto di stabilizzazione del fondo marino.
Numerose ricerche hanno validato, inoltre, P. oceanica come bioindicatore grazie ad alcune caratteristiche che rendono questa fanerogama marina particolarmente adatta per studi
10 ambientali e per il monitoraggio della fascia costiera. Questa specie è infatti dotata di: sensibilità ai cambiamenti ambientali (Augier, 1985; Maserti et al., 1988; Grauby et al., 1991; Pergent, 1991), ampia distribuzione geografica (Boudouresque et al., 1990a), longevità e risulta facile al prelievo di campioni (Boudouresque et al., 1990b). È stata inoltre dimostrato che Posidonia oceanica è capace di accumulare un vasto spettro di inquinanti, come composti organoclorurati, lindano, policlorobifenili (Chabert et al., 1984), alcuni radionuclidi artificiali (Florou et al., 1985; Calmet et al., 1988, 1991) e metalli in traccia (Augier et al., 1977; Chabert et al., 1983; Maserti et al., 1988; Malea & Haritonidis, 1989; Gnassi-Barelli et al., 1991). Sia per queste proprietà, che per la possibilità di studiare il processo in termini dinamici, attraverso la datazione delle scaglie, ad esempio con la lepidocronologia (Pergent, 1990), è stato ipotizzato che tale pianta possa conservare la memoria di contaminazioni ambientali (Calmet et al., 1988; Carlotti et al., 1992; Pergent-Martini & Pergent, 1995; Romeo et al., 1995).
3.2. Studio delle praterie di P. oceanica
Le praterie di Posidonia oceanica sono le più importanti e le più abbondanti tra le praterie di Angiosperme del Mar Mediterraneo e, grazie alla loro ampia distribuzione, alla loro caratteristica ecologica di specie “sedentaria” e alla loro suscettibilità alle mutevoli condizioni ambientali, sono utilizzate come indicatori biologici della qualità e della salute delle acque (Pergent-Martini & Pergent, 2000).
Negli ultimi anni, è stato osservato un'allarmante declino di molte popolazioni di P. oceanica nel Mediterraneo soprattutto nella parte Nord-Ovest del bacino (Ardizzone et al., 2006; Boudouresque et al., 2006), attribuito a disturbi naturali ma anche di natura umana (Duarte, 2002). Infatti il fenomeno è andato aumentando negli ultimi anni di pari passo con l'aumento della pressione antropica lungo la fascia costiera. P. oceanica esercita un ruolo importante nelle zone costiere, ed essendo appunto sensibile al degrado ambientale è oggi oggetto di notevole interesse a causa della tendenza alla progressiva e diffusa regressione.
Data l'importanza delle praterie di P. oceanica, al fine di tutelarne la conservazione è necessario valutarne lo stato di salute. Il loro studio prevede l'uso di descrittori che vengono distinti in: fisici, fisiogeografici, strutturali e funzionali.
11 rientrano nel gruppo dei descrittori fisici e si possono ottenere analizzando, ad es., le immagini raccolte attraverso l’uso di una telecamera subacquea. La morfologia, il profilo della costa, l'estensione batimetrica, la morfologia del fondale, le condizioni idrodinamiche, le caratteristiche dei sedimenti all'interno della prateria, le caratteristiche nelle zone di erosione e quelle sui limiti, sono requisiti che è necessario conoscere al fine di scrivere correttamente la situazione delle praterie (Buia et al., 2003).
A seconda delle caratteristiche delle aree colonizzate e delle specie strutturanti queste aree, le fanerogame marine danno origine a differenti tipologie di praterie.
A seconda della natura del substrato, le praterie si distinguono in praterie su sabbia, praterie su roccia e praterie su matte. Tutte le fanerogame si sviluppano su substrato sabbioso e/o su matte, ma solo P. oceanica può formare praterie su roccia.
Una seconda tipologia di praterie è connessa al numero di specie che la compongono; infatti quando una prateria è formata da una sola specie è detta pura o monospecifica, mentre quando più specie contribuiscono alla sua formazione è detta mista o plurispecifica. Le praterie miste sono più frequenti sotto costa, mentre quelle pure sono caratteristiche a profondità maggiori.
In rapporto alla distribuzione della pianta sul fondo si può determinare una terza tipologia. Le praterie sono così denominate "omogenee" quando le fanerogame sono ripartite uniformemente su tutta la sua estensione e denominate "disomogenee" in caso contrario.
Integrando la seconda e la terza tipologia si può distinguere quattro categorie: pura omogenea, pura disomogenea, mista omogenea e mista disomogenea.
Infine si possono distinguere praterie continue, discontinue e discrete a seconda che il fondo sia interamente colonizzato, interrotto dalla presenza di canali o interrotto dalla presenza di radure. La tipologia delle praterie può essere ricavata attraverso osservazioni dirette in immersione tramite operatori subacquei (nel caso di praterie di estensione limitata) o utilizzando il Side Scan Sonar (soprattutto nel caso di praterie di vaste dimensioni).
Tra i descrittori fisiografici rientra anche la tipologia del limite superiore e inferiore. Il limite superiore delle praterie corrisponde alla batimetrica più bassa alla quale la prateria ha inizio, mentre il limite inferiore corrisponde alla batimetrica più profonda a livello della quale la prateria termina. La profondità del limite superiore dipende dalle condizioni idrodinamiche e dalla pendenza del fondale, mentre la profondità del limite inferiore dipende dalla trasparenza delle acque e dalle caratteristiche biologiche delle specie. P. oceanica sviluppa
12 praterie molto strutturate e complesse e il suo limite inferiore presenta un aspetto diverso in base alle caratteristiche ambientali (Buia et al., 2003). Per questa specie quindi, il limite inferiore è classificato in quattro tipologie (Meinesz & Laurent, 1978; Pergent et al., 1995) (Fig. 3):
il limite inferiore è di tipo “progressivo” quando si presenta con un'alta percentuale di rizomi plagiotropi, ovvero disposti parallelamente alla direzione della pendenza, e la matte risulta poco strutturata o assente. Questo tipo di limite sta ad indicare sia un avanzamento della prateria sia che la luce non risulta un fattore di limitazione;
il limite inferiore è di tipo “netto” quando la prateria termina in modo repentino e sono presenti rizomi plagiotropi e ortotropi insieme e la matte è assente. Questo tipo di limite indica che la luce è un fattore limitante;
il limite è di tipo “erosivo” quando prevalgono i rizomi ortotropi e si osserva un gradino di matte esposta; il fattore che limita la progressione in questa tipologia è l'idrodinamismo;
il limite è di tipo “regressivo” quando si presenta con matte morta sulla quale si possono trovare isolati alcuni ciuffi di piante vive. Questo tipo di limite indica il declino della prateria per uno o alcuni dei numerosi fattori per cui ciò può avvenire.
In base alla profondità (p) del limite inferiore, Pergent et al. (1995) hanno proposto una scala di valutazione della trasparenza delle acque (Tab. 1).
Le osservazioni del limite inferiore, forniscono informazioni circa i cambiamenti che possono essere avvenuti per cause naturali e cambiamenti per impatti di natura antropica (progressione o regressione della prateria). Attraverso il limite inferiore può essere valutata anche la dinamica della prateria nel tempo e per eseguire questo monitoraggio possono essere utilizzate tre tecniche diverse. La tecnica operativamente più semplice è quella del "Balisage", che prevede il posizionamento di corpi fissi (per esempio blocchi di cemento) in siti prescelti usati come punti di riferimento. Confrontando le fotografie che vengono scattate annualmente da tre diverse angolazioni viene valutata la dinamica della prateria. Le altre tecniche prevedono l'uso di fotografie e/o Side Scan Sonar; quest'ultimo strumento è molto più complesso e ha costi di applicazione maggiori rispetto al balisage ma rispetto a questo dà la possibilità di valutare una superficie molto più ampia in un arco di tempo ridotto.
Per valutare lo stato di una prateria, soprattutto se misurato su scala temporale pluriannuale, il descrittore più importante è la densità. Essa consta nella determinazione del
13 numero dei fasci fogliari su l’unità di superficie (fsc m-2). La densità può essere determinata in situ contando direttamente i fasci fogliari presenti all’interno di una superficie campionaria di forma quadrata, oppure in laboratorio, dopo la raccolta dei ciuffi fogliari contenuti all’interno di una superficie campionaria. La superficie di campionamento attualmente ritenuta sufficiente a individuare un’area minima ottimale per valutare la densità è di 3600 cm2 (Cormaci et al., 2003; AAVV, 2008).
Per quanto riguarda il numero di campioni per area di campionamento e il numero di repliche per ciascuno di questi da effettuare, questo può variare in accordo la omogeneità e l’articolazione del fondo della prateria. Per le praterie di P. oceanica, Giraud (1977b), propone una scala di valutazione in relazione alla densità che permette di distinguere sei tipi di praterie; solo in seguito questa è stata ridotta a cinque tipi (Tab. 2), considerando il fatto che quando il numero dei fasci per m2 è inferiore a 50, questi non sono ritenuti sufficienti a definire una prateria (Buia et al., 2003).
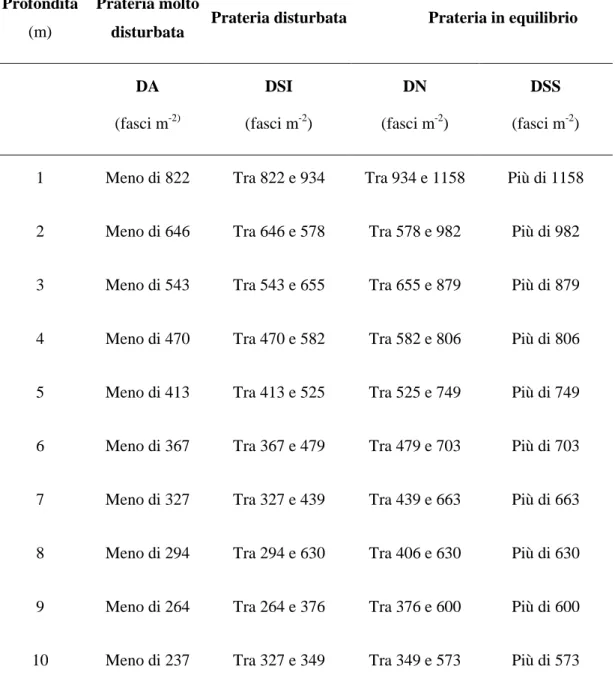
Più recentemente è stata proposta da Pergent et al. (1995) e da Pergent-Martini & Pergent (1996), una classificazione delle praterie per la quale la densità è rapportata alla profondità e ad altri fattori ambientali, come torbidità e disturbo antropico (Tab. 3). Si possono distinguere:
praterie in equilibrio, dove la densità è molto alta (DSS) o dove rientra nella norma (DN);
praterie disturbate, dove la densità è bassa (DSI);
praterie molto disturbate, dove la densità è molto bassa (DA).
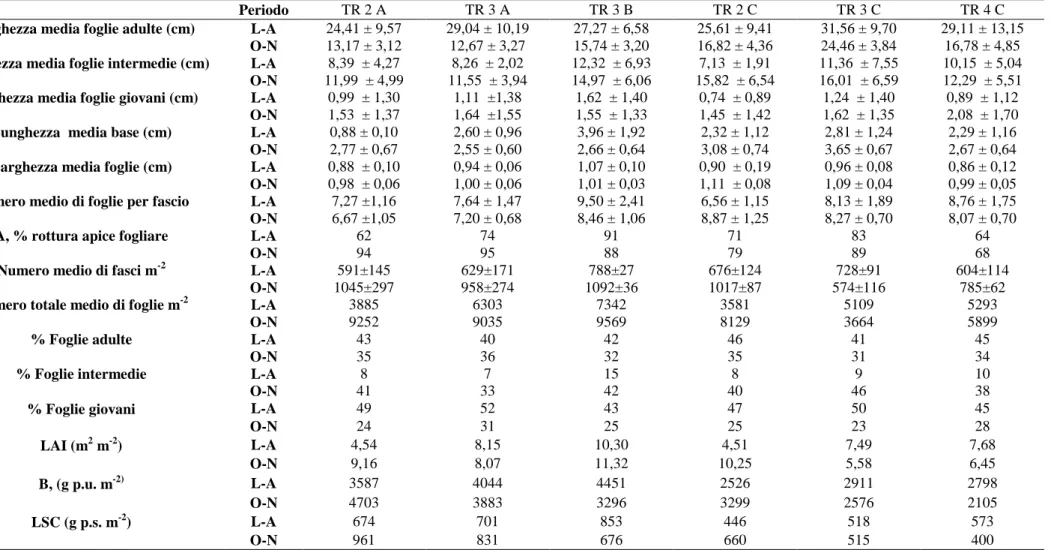
Il fascio fogliare (fsc) costituisce l'unità funzionale per la descrizione di una prateria (Buia et al., 2003). Ad ogni fascio fogliare corrisponde una porzione ipogea costituita dai rizomi e dalle radici. La lunghezza, la larghezza e la biomassa di foglie, rizomi e radici, quali descrittori funzionali definiti “sintetici”, permettono di ricavare, attraverso calcolo, i descrittori funzionali definiti “derivati”, i quali consentono di delineare lo stato di vitalità delle piante costituenti la prateria e gli eventuali danni procurati da fattori ambientali, biologici e antropici.
Le foglie sono classificate, in accordo con Giraud (1977a), in tre categorie: giovanili (fg), con lunghezza inferiore a 50 mm e privi di base;
intermedie (fi), con lunghezza superiore a 50 mm e prive di base; adulte (fa), con lunghezza superiore a 50 mm e provviste di base.
14 Per determinare le caratteristiche biometriche ogni unità funzionale è scomposta nelle sue componenti (foglie, rizomi e radici) che sono poi analizzate separatamente. Le foglie vengono separate nelle tre categorie per poter così ottenere anche il numero per classe oltre al numero totale. Viene stimata la larghezza, misurata a circa metà del lembo, della foglia adulta (La), e la lunghezza del lembo fogliare, misurata dalla ligula (se presente) o dal punto di inserzione sul rizoma fino all'apice. Per ogni foglia adulta viene stimata anche la lunghezza della base (b), misurata dal suo punto di inserzione sul rizoma fino alla concavità della ligula. I valori ottenuti dall'analisi biometrica, vengono mediati al fine di ottenere la lunghezza e la larghezza media di ogni classe di foglie e di basi. Questi valori sono detti descrittori funzionali sintetici e sono espressi in centimetri o in millimetri. Si può ricavare inoltre il numero medio di foglie per classe e per ciuffo.
La biomassa è distinta in epigea e ipogea e indica rispettivamente il peso dell'apparato fogliare e il peso di rizomi e radici. La biomassa può essere espressa in grammi per m2, di peso fresco o peso secco. Il rapporto tra biomassa ipogea e epigea delle diverse specie di angiosperme marine spiega il loro differente ruolo svolto negli ecosistemi.
Appartengono alla classe dei descrittori funzionali anche altri descrittori detti “derivati”, ottenuti per calcolo dai descrittori sintetici. Questi sono:
Indice di superficie fogliare (Leaf Area Index, LAI) (m2 m-2). Esso viene calcolato moltiplicando la superficie fogliare media di un fascio per la densità della prateria per metro quadrato.
Indice di biomassa fogliare (Leaf Standing Crop, LSC) (gps m-2). Viene calcolato moltiplicando la biomassa fogliare media per fascio per la densità della prateria riferita al metro quadro; per Posidonia oceanica nel calcolo di questo descrittore non viene considerata anche la biomassa delle basi.
Coefficiente A. Questo parametro rappresenta la percentuale di foglie adulte e intermedie che hanno subito la rottura degli apici; esso viene calcolato dividendo il numero totale di foglie adulte e intermedie con gli apici rotti, per il numero totale di foglie osservate. A è spesso in relazione con lo stress idrodinamico e di pascolo da parte di erbivori, e può essere usato quindi come indice dell'impatto naturale sulle foglie più lunghe. È inoltre possibile distinguere il danno meccanico da quello biologico; infatti gli erbivori lasciano delle tracce sulle foglie che sono caratteristiche per le varie specie.
15 La crescita è un altro descrittore funzionale e rappresenta l'aumento in quantità (espressa in termini di numero, peso e lunghezza) che un organismo registra in un preciso intervallo temporale (Buia et al., 2003). La crescita delle fanerogame può essere stimata facendo uso di metodi diretti ed indiretti. La tecnica diretta prevede la marcatura delle parti della pianta come foglie o rizomi, mentre quella indiretta prevede l’analisi dei caratteri permanenti che sono lasciati dalle foglie sul rizoma, come le cicatrici a livello dei nodi o la valutazione dello spessore delle scaglie presenti sui rizomi (lepidocronologia). La lepidocronologia si basa sullo stesso principio della dendrocronologia e della sclerocronologia, applicate rispettivamente allo studio della crescita delle piante terrestri e dello scheletro dei bivalvi (Peirano et al., 2005). Essa misura lo spessore delle scaglie che rimangono attaccate al rizoma una volta che il lembo fogliare è caduto. Lo spessore delle scaglie varia ciclicamente da un massimo nel periodo estivo-autunnale ad un minimo in periodo invernale- primaverile. Le scaglie più sottili corrispondono alle basi delle foglie che crescono durante l’estate il cui lembo cade in inverno. L’individuazione di due minimi successivi identifica il corrispondente anno lepidocronologico e la parte di rizoma prodotta durante questo periodo rappresenta l’unità di crescita annuale. L’unità di crescita annuale comincia al primo internodo prodotto dopo la scaglia più sottile e termina all’ultimo internodo prodotto prima della successiva scaglia sottile. Partendo dalla data del campionamento, con questa tecnica è possibile retrodatare il rizoma e valutare il numero di foglie prodotte per gli anni lepidocronologici individuati. Tra le fanerogame marine, quest’analisi è applicabile solo a P. oceanica, in quanto solo in questa specie le scaglie rimangono attaccate al rizoma per lunghissimo tempo, anche centinaia di anni, formando una specie di manicotto (Roméo et al., 1995; Buia et al., 2003; Mayot et al., 2005). La capacità di “memorizzare” i dati ambientali da parte degli organismi ipogei appare estremamente affidabile e riproducibile, rendendo questa specie, e in particolare l'analisi lepidocronologica, accoppiata con la determinazione dei contaminanti, uno degli approcci più promettenti per il monitoraggio delle aree costiere. Per esempio, Pergent-Martini (1995) ha dimostrato che raggruppando le scaglie di Posidonia per anni lepidocronologici, si possono osservare annualmente le variazioni nel contenuto dei metalli in tracce. Queste variazioni, non sono casuali ma riflettono variazioni nella concentrazione ambientale dei contaminanti.
16 3.3. Le praterie emergenti: barrier reef
Le praterie emergenti di Posidonia oceanica, oltre ad essere delle peculiarità di elevata importanza naturalistica, costituiscono delle vere e proprie barriere frangiflutti (Fig. 4). Il fenomeno produce condizioni particolari nella zona di retro prateria, che in seguito alla riduzione del moto ondoso, risulta essere una zona calma, con un sedimento molto fine e ricco di materia organica, tale che permette lo sviluppo di insediamenti fitobentonici tipici (Molinier & Picard, 1952). Il sedimento grossolano viene, infatti, trattenuto dalle fronde della pianta. Man mano che, attraverso la crescita verticale, la prateria si innalza, solo la parte più esposta ad intenso idrodinamismo rimane vitale, mentre la zona meno esposta inesorabilmente muore. Il ridotto idrodinamismo ed il conseguente ridotto rinnovo delle acque, rende il retro prateria una zona a caratteristiche “lagunari”. Gli autori francesi, che più di altri si sono maggiormente e per primi occupati di questi aspetti, utilizzano un termine specifico, “lagon”, per descrivere tale peculiarità. A queste condizioni corrisponde un insediamento vegetale con specie pioniere, come Caulerpa prolifera, tra le alghe fotofile e Cymodocea nodosa, tra le angiosperme; successivamente si può insediare, fino a prevalere, Nanozostera noltii, che predilige substrati ricchi di materia organica.
Le prime formazioni di P. oceanica a soffrire dell’antropizzazione della fascia costiera sono state proprio le barrier reef, le quali sono da tempo in regressione in tutto il Mediterraneo, soprattutto in conseguenza dello sviluppo caotico dei porti turistici (Boudouresque et al., 1975, 1985). In Italia queste formazioni sono molto rare, e vanno ricordate soprattutto quelle di Portofino (Issel, 1918) e dello Stagnone di Marsala (Calvo & Fradà Orestano, 1984).
3.4. La regressione delle praterie di P. oceanica nel Mar Mediterraneo
Già da metà dello scorso secolo si sta manifestando un diffuso fenomeno di regressione delle praterie di P. oceanica (Fig. 5). Tale fenomeno può essere imputato a cause di diversa origine, sia antropiche che naturali.
Le attività umane rappresentano un fattore di possibile degradazione e distruzione delle praterie. L’emissione nelle acque costiere di scarichi di origine urbana crea una condizione di elevata torbidità, dovuta sia all’aumento del tripton, che al massiccio sviluppo di organismi
17 fitoplantonici; il notevole carico organico provoca anche un eccessivo sviluppo di epifiti vegetali sulle foglie di Posidonia. Il risultato è una diminuzione dell’intensità luminosa che raggiunge la pianta e una conseguente riduzione della sua attività fotosintetica.
A tal proposito, Peres (1984) ha osservato lungo i litorali più urbanizzati del Mediterraneo (Francia, Italia e Spagna), una diminuzione della densità fogliare della prateria e un calo della copertura del fondale occupato. Il restringimento della fascia di fondo colonizzata è dovuto a una netta risalita del limite inferiore, contemporanea ad una discesa in profondità del limite superiore. Gli scarichi urbani oltre all’aumento di torbidità, provocano la regressione delle praterie, a causa della grande quantità e varietà di sostanze chimiche ad essi associate, come detergenti e idrocarburi, che alterano la biosintesi dei pigmenti fotosintetici (Augier & Maudinas, 1979; Panayotidis et al., 1990).
Un’ altra importante minaccia che provoca regressione delle praterie, è collegabile allo stress meccanico legato all’attività di pesca ed agli ancoraggi ripetuti (Augier & Boudouresque, 1970; Peres, 1984). Secondo Peres (1977), le reti a strascico, arando il substrato, estirpano le foglie e i rizomi della pianta, provocando una diminuzione della densità della pianta e delle sostanze umiche, che si riflette sui meccanismi di crescita.
Tra le alterazioni legate indirettamente alle attività umane, possiamo anche aggiungere l’introduzione di specie alloctone. Queste potrebbero competere con P. oceanica per il substrato. L’esempio più importante di occupazione del substrato da parte di specie alloctone è rappresentato dall’espansione di Caulerpa taxifolia (Vahl) C. Agardh e Caulerpa racemosa (Forsskal), alghe tropicali appartenenti al phylum Chlorophyta (De Villele & Verlaque, 1995; Ceccherelli et al., 2000; Piazzi et al., 2001; Ceccherelli et al., 2002).
E’ probabile che nessuna di queste fonti di disturbo sia sufficiente da sola a produrre la regressione delle praterie a cui stiamo assistendo, ma che si tratti piuttosto di un effetto sinergico tra i vari fattori considerati.
Bisogna infine tenere conto, tra le possibili cause di regressione, i fattori di origine naturale. Moliner & Picard (1952) e successivamente Augier & Boudouresque (1970), in studi sulla regressione della “récife - barriere” della baia di Port-Cros, hanno ipotizzato che il rimaneggiamento della prateria possa essere dovuto ad una risposta di P. oceanica alle attuali condizioni climatiche del Mar Mediterraneo. Secondo questi autori, una conferma di ciò sarebbe rilevabile nella sporadicità dei fenomeni di fioritura e fruttificazione, che si verificano di norma solo a seguito di estati molto calde. La graduale rarefazione delle praterie rende
18 evidente e pressante il problema della loro salvaguardia.
Al fine di poter intraprendere concrete azioni di controllo, è necessario delineare in modo preciso l’estensione e le caratteristiche della prateria in un dato momento, e seguirne l’evoluzione nel tempo mediante successivi monitoraggi.
19 Figura 1: disegno schematico della ”matte“ e dello strato dei rizomi e delle foglie di P.
oceanica
a b c
Figura 2: il fascio fogliare (a), l’ infiorescenza (b), il frutto (c) detto “oliva di mare” di P. oceanica
Figura 3: Tipologie di limiti inferiori per le praterie di P. oceanica. 1- Limite progressivo. 2- Limite netto. 3- Limite erosivo. 4- limite regressivo.
20 Figura 4: Schema di “barrier reef”
21 Profondità del limite inferiore Valutazione
< 15 m acque estremamente poco
trasparenti (EPT) > 15 m < 25 m acque poco trasparenti (PT)
> 25 m < 35 m acque trasparenti (T)
> 35 m acque altamente trasparenti (AT)
Tabella 1. Scala di valutazione della trasparenza delle acque in rapporto alla profondità del limite inferiore della prateria di P. oceanica.
Tipo Densità (fasci m-2) Valutazione
Tipo I > 700 Prateria molto densa (MD)
Tipo II 700-400 Prateria densa (D)
Tipo III 400-300 Prateria rada (R)
Tipo IV 300-150 Prateria molto rada (MR)
Tipo V 150-50 Semiprateria (SP)
22 Profondità
(m)
Prateria molto
disturbata Prateria disturbata Prateria in equilibrio
DA (fasci m-2) DSI (fasci m-2) DN (fasci m-2) DSS (fasci m-2)
1 Meno di 822 Tra 822 e 934 Tra 934 e 1158 Più di 1158
2 Meno di 646 Tra 646 e 578 Tra 578 e 982 Più di 982
3 Meno di 543 Tra 543 e 655 Tra 655 e 879 Più di 879
4 Meno di 470 Tra 470 e 582 Tra 582 e 806 Più di 806
5 Meno di 413 Tra 413 e 525 Tra 525 e 749 Più di 749
6 Meno di 367 Tra 367 e 479 Tra 479 e 703 Più di 703
7 Meno di 327 Tra 327 e 439 Tra 439 e 663 Più di 663
8 Meno di 294 Tra 294 e 630 Tra 406 e 630 Più di 630
9 Meno di 264 Tra 264 e 376 Tra 376 e 600 Più di 600
10 Meno di 237 Tra 327 e 349 Tra 349 e 573 Più di 573
Tabella 3. Classificazione delle praterie in funzione della densità dei fasci e della profondità. Sono considerate solo le profondità fino a 10 metri (Pergent et al., 1995)
23 4. Scopo della tesi
Questa tesi si inserisce nel quadro delle attività di ricerca del Laboratorio di Ecologia Lagunare e Acquacoltura della Orbetello Pesca Lagunare srl, volte a definire lo stato delle praterie di Posidonia oceanica insediate lungo la costa della provincia grossetana a Sud della foce dell’Ombrone.
La ricerca su cui si basa la parte sperimentale di questa tesi ha avuto come obiettivo principale stabilire se e in che misura le attività umane del tratto di costa di S. Liberata, dove si insedia una piccola prateria in parte emergente, a costituire un barrier-reef, abbiano indotto evidenti alterazioni a questo insediamento o, comunque, abbiano esercitato un impatto rilevabile.
In subordine, la determinazione di parametri nutrizionali nei tessuti e nei sedimenti e della tessitura sedimentaria e del contenuto in materia organica, ai limiti superiore (area di retro-barriera) e inferiore, furono diretti a stabilire lo stato trofico dell’ecosistema e le dinamiche collegate, con lo scopo di contribuire alla comprensione delle cause della recente dinamica dei popolamenti fitobentonici, soprattutto nell’area di retro-barriera.
Il primo obiettivo trova ragione nella intensa attività portuale diportistica, che si è sviluppata nel canale detto “Navigabile”, adiacente al canale di comunicazione mare-laguna della Laguna di Orbetello, a partire dagli anni ’70 e che attualmente si è andata configurando come una vera e propria area portuale con un movimento di circa 1000 imbarcazioni da diporto. Il secondo obiettivo trova ragione:
nella vicinanza della prateria con un canale mare-laguna di una laguna che negli ultimi 20 anni ha raggiunto valori di elevata eutrofia, che veicola, pertanto, grandi quantità di detrito organico e, talvolta, di fitoplancton;
nel recente sviluppo della balneazione nel breve tratto della spiaggia di S. Liberata, a seguito della creazione di un’area di sosta attrezzata per camper di fronte alla spiaggia stessa, che ha determinato l’incremento dei fruitori di almeno un ordine di grandezza.
24 5. Materiali e Metodi
5.1. Area di Studio
L’area oggetto del presente studio è costituita da un tratto di costa esteso 400 m circa, nella parte più meridionale della Toscana, a sud di Grosseto, denominato S. Liberata. Questa località è l’ultimo tratto del tombolo della Giannella, che, traendo origine dagli apporti terrigeni del fiume Albegna, unisce la costa continentale con Monte Argentario e separa il bacino di Ponente della laguna di Orbetello dal mare (Fig. 6).
Nell’area sommersa antistante la spiaggia di S. Liberata è presente una formazione naturale emergente di P. oceanica (Fig. 6), che ha determinato la formazione di un’area retrostante la barriera a ridotta idrologia, la quale, fino al 2003, era caratterizzata da un popolamento misto costituito dalle Angiospermae Cymodocea nodosa Ucria e Nanozostera noltii (Hornemann) Tomlinson et Posluzny e dalla Chlorophyta Caulerpa prolifera Forsskal (Lenzi, 1987).
L’area di studio è delimitata a sinistra, cioè verso Monte Argentario, dai manufatti dei canali artificiali di comunicazione tra il mare e la laguna di Orbetello: il Canale di Nassa, che è il canale vero e proprio di vivificazione del bacino lagunare, dove al suo termine lagunare sono poste le strutture per la pesca, detti lavorieri, ed il Canale Navigabile, che in passato consentiva la navigazione di imbarcazioni per il trasporto di greggi di lavorazioni industriali dal mare ad una fabbrica di prodotti chimici sita in Orbetello Scalo. Non essendo più navigato da molto tempo, e allo scopo chiuso alla sua foce lagunare, questo canale ha assunto sempre più i connotati di piccolo porto turistico.
Inoltre, per ovviare ai fenomeni erosivi che insistono lungo buona parte del tombolo di Giannella, fin dagli anni ’70, lungo il tratto più meridionale del tombolo sono state realizzate, a più riprese, con sassi e materiale terroso, quattro barriere frangiflutti; l’ultima delle quali, costruita nel 1996, è quella più vicina all’area di studio e alla prateria in oggetto.
Infine, la spiaggia di S. Liberata, negli ultimi anni, ha subito un considerevole aumento del flusso turistico, dovuto essenzialmente alla realizzazione, a poche decine di metri, di un’area attrezzata per la ricezione dei camper. Poiché l’abitato di S. Liberata, disposto ai due margini della Strada Provinciale della Giannella, impedisce il passaggio al mare per 500 m circa, e l’erosione della spiaggia e le opere in sua difesa hanno trasformato la spiaggia, per circa 200 m, in un cumulo di sassi di riporto, che dividono così la spiaggia di S. Liberata dalla
25 restante spiaggia di Giannella, per i camperisti della vicina area attrezzata, la spiaggia di S. Liberata è pertanto l’unico tratto di spiaggia fruibile a piedi.
Dalla descrizione dell’area, si evince che essa è stata soggetta, ed è ancora soggetta, a molteplici impatti di natura antropica, anche diversificati, nel tempo.
5.2 Disegno di campionamento
Il principale obiettivo di questa tesi è stato quello di verificare lo stato di salute della prateria di P. oceanica in oggetto, in rapporto agli stress a cui è stata finora sottoposta. Pertanto il lavoro assume un carattere di indagine ambientale, per il quale si è agito sui seguenti punti:
1- stabilire una base per una futura verifica della dinamica di questa prateria;
2- stabilire un piano di campionamento per descrivere la prateria attraverso i parametri biometrici, della densità e della biomassa;
3- stabilire un piano di campionamento per verificare la presenza di contaminanti, in parti diverse dell’ecosistema (alghe, angiosperme, sedimenti);
4- stabilire un piano di campionamento per definire lo stato di trofia dell’ecosistema in esame. Questo è stato effettuato attraverso le determinazioni del contenuto C, N, P ai livelli di P. oceanica, macroalghe e sedimento. Relativamente ai sedimenti, è stata determinata la percentuale di materia organica e la quantità di materia sedimentabile.
I rilevamenti in campo e i campionamenti sono stati condotti nel 2008, in due campagne, una in Luglio e una in Novembre.
5.2.1 Balisages
Per ottemperare al primo punto, nel Luglio, ai limiti del posidonieto, sono stati posizionati 6 corpi fissi di riferimento (balisages), tre al margine superiore e tre a quello inferiore (Fig. 7, 8), al fine di poter valutare le dinamiche dei margini della prateria, nel tempo. La posizione dei balisages è stata georiferita mediante Global Positioning System (GPS) e la disposizione rispetto al margine di prateria è stata documentata attraverso fotografie da prospettive diverse.
26 5.2.2. La prateria di P. oceanica
Descrittori della prateria
In entrambe le campagne di Luglio e Novembre, lungo tre transetti costa-largo (TR2, TR3, TR4), in 6 stazioni (TR2A, TR3A, TR3B, TR2C, TR3C e TR4C) (Fig. 4), sono state condotte misure e campionamenti, per la determinazione dei parametri biometrici, della densità e della biomassa di P. oceanica. La determinazione della densità è stata effettuata su due repliche condotte direttamente in situ, mediante il conteggio dei fasci fogliari (campionamento non distruttivo) all’interno di una superficie campionaria 60x60 cm (3600 cm2) (Cormaci et al., 2003), e su una terza replica, in laboratorio, dopo la raccolta dei ciuffi fogliari (campionamento distruttivo) contenuti all’interno della stessa superficie campionaria (Fig. 9).
Tutte le fasi di campionamento sono avvenute mediante operatore subacqueo con autorespiratore ad aria (ARA), coadiuvato da personale in acqua per le basse profondità e con barca per le stazioni più profonde.
Il materiale vegetale asportato è stato insacchettato al momento del prelievo, opportunamente siglato, quindi portato in laboratorio dove è stato utilizzato per la determinazione della biomassa e per le misure biometriche.
Una volta determinato il numero delle foglie formanti i fasci fogliari, il seguente valore è stato riportato all’unità di superficie (numero dei fasci per m2, fsc m-2).
Le misure effettuate sulle foglie hanno permesso di calcolarne la superficie complessiva in m2 di tutte le foglie presenti in un m2 (LAI). Sebbene le foglie di Posidonia possiedano un parenchima clorofilliano su entrambe i lembi fogliari, per convenzione tale misura è stata espressa rispetto ad una sola pagina fogliare.
I campioni quantitativi sono stati puliti e pesati umidi, quindi un’aliquota del materiale umido è stato essiccato in forno a 90°C per 24h, per la determinazione della biomassa fotosintetica, il Life Standing Crop (LSC).
Da ciascun campione di materiale umido, successivamente al conteggio dei fasci e alla pesata, sono stati prelevati casualmente 20 fasci per le misure biometriche. Da questi subcampioni sono stati stimati, secondo Giraud (1977a), i seguenti parametri biometrici, già descritti in Introduzione: NF fsc-1, Fa fsc-1, Fi fsc-1, Fg fsc-1, Fa, Fi, Fg, La, b e A.
27 la percentuale delle foglie adulte, %Fa; la percentuale delle foglie intermedie, %Fi; la percentuale delle foglie giovani, %Fg; il numero totale delle foglie per metro quadrato, NF m-2; la lunghezza medie delle foglie adulte, Fa; la lunghezza medie delle foglie intermedie Fi; la lunghezza medie delle foglie giovani Fg; la larghezza media delle foglie adulte, La; la lunghezza medie della base, b; la percentuale delle foglie adulte e intermedie rotte, A.
Al fine di produrre un profilo della morfologia di prateria e di retro prateria, lungo i 4 transetti ortogonali alla costa (Fig. 4), è stata rilevata, mediante operatore subacqueo, la profondità (p) ogni 3m. Con lo stesso metodo, al fine di descrivere i margini della prateria e stimare la distribuzione delle matte morte, delle intermatte e dei popolamenti algali, è stato effettuato un rilevamento fotografico.
5.2.3. Contaminanti
Vegetazione
Dei sei campioni (TR2A, TR3A, TR3B, TR2C, TR3C e TR4C) (Fig. 4) prelevati per la determinazione dei parametri biometrici, della densità e della biomassa di P. oceanica, un’aliquota è stata utilizzata per rilevare le concentrazioni di contaminanti. Allo stesso scopo, inoltre, ipotizzando un possibile gradiente dei contaminanti dai manufatti dell’ingresso portuale verso la spiaggia della Giannella, sono stati raccolti, lungo i transetti TR1 e TR4, in prossimità degli estremi superiore e inferiore della prateria, campioni di: Caulerpa racemosa var cylindracea (Sonder) Verlaque, Huisman, Boudouresque; Caulerpa prolifera (Forsskål) J.V. Lamouroux; Padina pavonia (L) Lamouroux; Cladophora sp.; Cymodocea nodosa (Ucria) Ascherson; Nanozostera noltii (Hornemann) Tomlinson et Posluzny.
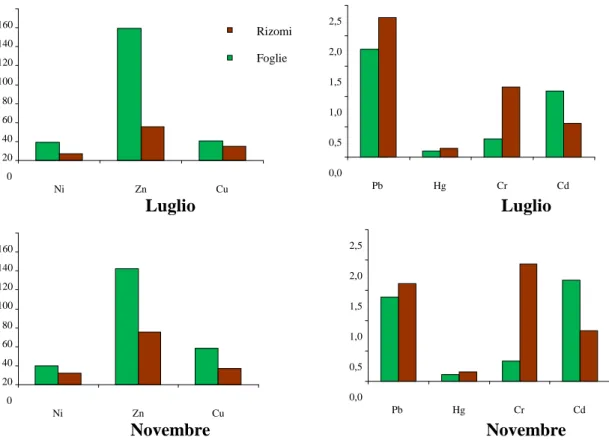
Tutti i campioni di materiale vegetale sono stati sottoposti all’analisi per la determinazione di: Mercurio (Hg), Cadmio (Cd), Piombo (Pb), Rame (Cu), Nichel (Ni), Cromo (Cr), Zinco (Zn) e delle principali 16 molecole di idrocarburi policiclici aromatici (IPA).
Tutti i campioni sono stati conservati in congelatore a -18°C, fino al momento dell’analisi. Per P. oceanica il materiale è stato suddiviso in rizomi e foglie. In entrambe le frazioni sono stati determinati i metalli, solo nelle foglie gli IPA.
28 Determinazione dei metalli
Le foglie di P. oceanica sono state raschiate con un vetrino e sciacquate in acqua di mare prelevata dal sito di campionamento, al fine di rimuovere l’epibiota ed il sedimento adeso (Dauby & Poulicek, 1995). Quest’ultima operazione si rende necessaria in quanto, negli studi eco-tossicologici, le foglie e l’epibiota non devono essere trattati come una singola unità, dal momento che i metalli sono accumulati in grado diverso nelle due componenti.
0,5 g di ciascun campione sono stati sottoposti a mineralizzazione mediante una miscela di acido nitrico (65%) e acqua ossigenata (30% v/v) in un sistema chiuso a microonde (ETHOS 900 Microwave Labstation, Milestone), in accordo con il metodo consigliato dal Manuale Milestone DG-Environment 25, Green Algae. Dopo il raffreddamento, le soluzioni ottenute sono state filtrate mediante carta da filtro (Whatman, Schleicher and Schuell) e portate ad un volume noto, aggiungendo acqua ultrapura. Per ogni carica del mineralizzatore è stata effettuata una prova di un bianco dei reattivi con le stesse modalità operative adottate per le matrici dei campioni.
La determinazione di Cd, Pb, Cr, Ni e Cu è stata eseguita per spettrofotometria ad assorbimento atomico con atomizzazione elettrotermica (GF-AAS) mediante AAnalyst 700, Perkin Elmer. Al fine di minimizzare le interferenze dovute alla matrice e stabilizzare l’analita sono stati aggiunti alcuni sali modificatori. In particolare, per la quantificazione del Cd sono stati aggiunti 15 µg di palladio e 10 µg di nitrato di magnesio, per l’analisi del Pb 200 µg di ammonio fosfato e per la determinazione del Cr 50 µg di nitrato di magnesio. Hg è stato determinato in assorbimento atomico mediante generazione di vapori freddi (CV-AAS) con la tecnica di Flow Injection Mercury System (FIMS 400, Perkin-Elmer). Il dosaggio di Zn è stato effettuato attraverso spettrofotometria ad assorbimento atomico con atomizzazione in fiamma aria-acetilene (AAnalyst 700, Perkin Elmer).
Per il calcolo delle concentrazioni è stata utilizzata la retta di taratura ottenuta con il metodo delle aggiunte, che prevede l'addizione di quantità note e crescenti degli elementi da determinare a volumi uguali di campione. I risultati finali, ottenuti come media di tre letture, sono stati espressi in mg kg-1 peso secco.
La precisione analitica, espressa come coefficiente di variazione percentuale (CV%) di 5 repliche omogenee, è risultata inferiore al 10%. L'accuratezza è stata controllata con materiali di riferimento certificati dal National Institute of Standards and Tecnology (NIST): Peach Leaveas, Apple Leaves e Lagarosiphon major.
29 Determinazione degli IPA
La determinazione degli IPA si è basata sulle metodiche EPA 3545 A, 3610, 8310 che prevedono tre fasi distinte: estrazione degli analiti delle matrici, purificazione degli estratti e quantificazione degli analiti. Per l’estrazione è stata impiegata la tecnica di estrazione pressurizzata con solventi (ASE, Accelerated Solvent Extraction). Il metodo consente di estrarre da matrici solide e semisolide, molecole insolubili in acqua o scarsamente solubili in essa. Nella cella dell’ASE vengono caricati circa 2 g di allumina attivata e circa 1 g di campione liofilizzato e opportunamente miscelato con terra di diatomee. Nel caso delle foglie di Posidonia sono state scelte foglie adulte e intermedie intere (epifitate). Allumina e campione sono tenuti divisi dalla presenza di un setto di cellulosa. Per l’estrazione sono stati utilizzati i solventi esano e diclorometano, in rapporto volumetrico 3:7, alla temperatura di 60°C, e a 1500 psi di pressione, e due cicli di statica. Al fine di rimuovere lo zolfo molecolare, che potrebbe interferire nella quantificazione degli analiti, agli estratti è stato aggiunto Cu attivato. Dopo il raffreddamento, l’estratto viene concentrato a 1 ml tramite evaporatore rotante (BÜCHI). Successivamente, l’estratto viene portato a secco sotto flusso di azoto, e risospeso in 1 ml di acetonitrile per la successiva quantificazione con HPLC.
Gli IPA analizzati sono stati: naftalene; acetaftilene; acenaftene; fluorene; fenantrene; antracene; fluorantene; pirene; benzo (a)-pirene; risene; benzo (b)- fluorantene; benzo (k)-fluorantene; benzo(a)-antracene; dibenzo (a, h)- antracene; benzo (g, h, i)-terilene; indeno (1, 2, 3 c-d) pirene.
La strumentazione utilizzata per la quantificazione è costituita da Cromatografo Liquido ad Alta Prestazione (HPLC) della Dionex costituito da sistema di pompaggio multisolvente e rilevatori a UV-Vis e fluorescenza. La fase della colonna è una C18 (25 cm X 4.6 mm i.d.), l’eluizione è effettuata mediante gradiente binario di acqua/aceto nitrile (60-100% in 30 minuti) ad un flusso di 1,5 ml/minuto. La determinazione qualitativa è stata effettuata sulla base del tempo di ritenzione delle molecole utilizzando la standard PAH-Mix 9 (Dr. Ehrenstorfer). La determinazione quantitativa è stata effettuata mediante l’uso di curve di calibrazione con la tecnica dello standard esterno. Il controllo qualità è stato effettuato realizzando prove in bianco per ogni serie di analisi, prove di recupero mediante estrazione ripetute della stessa matrice da analizzare utilizzando standard di riferimento certificati (SRM 1944, NIST per il sedimento e Fucus sp. IAEA-140/OC per il materiale vegetale). Il recupero ottenuto per tutti gli analiti analizzati oscilla tra il 60 e il 93%. Il campione da analizzare (20
30 µl in acetonitrile) viene prelevato da un iniettore a siringa (loop iniector) e iniettato in colonna cromatografa. La fase mobile rappresentata dall’eluente spinge il campione attraverso la fase stazionaria della colonna. La separazione delle molecole in HPLC avviene in fase inversa, vale a dire la fase mobile è relativamente polare, mentre la fase stazionaria è apolare e la silice, di cui è composta quest’ultima fase, è modificata chimicamente tramite l’aggiunta di catene idrofobiche. In un miscuglio in cui gli analiti presentano diversa polarità, la loro separazione viene aggiunta variando la composizione della fase mobile. Utilizzando il gradiente di eluizione acqua/acetonitrile, gli idrocarburi a più basso peso molecolare, che sono anche relativamente più polari, vengono eluiti per primi e presentano quindi tempi di ritenzione più bassi; gli idrocarburi con maggior peso molecolare verranno eluiti successivamente e presenteranno tempi di ritenzioni maggiori. Lo strumento utilizzato per questa analisi presenta un sistema di rivelazione a fluorescenza e uno ad assorbimento nell’UV. Gli IPA sono stati quantificati utilizzando entrambi i sistemi; per l’assorbimento nell’UV, la lunghezza d’onda utilizzata è stata quelladi 254 nm, mentre per la fluorescenza si è utilizzato una programmata di lunghezza d’onda di eccitazione e di emissione in funzione dell’optimum di assorbimento delle varie molecole. Le concentrazioni sono espresse in ng g-1 di peso secco di campione.
Sedimento
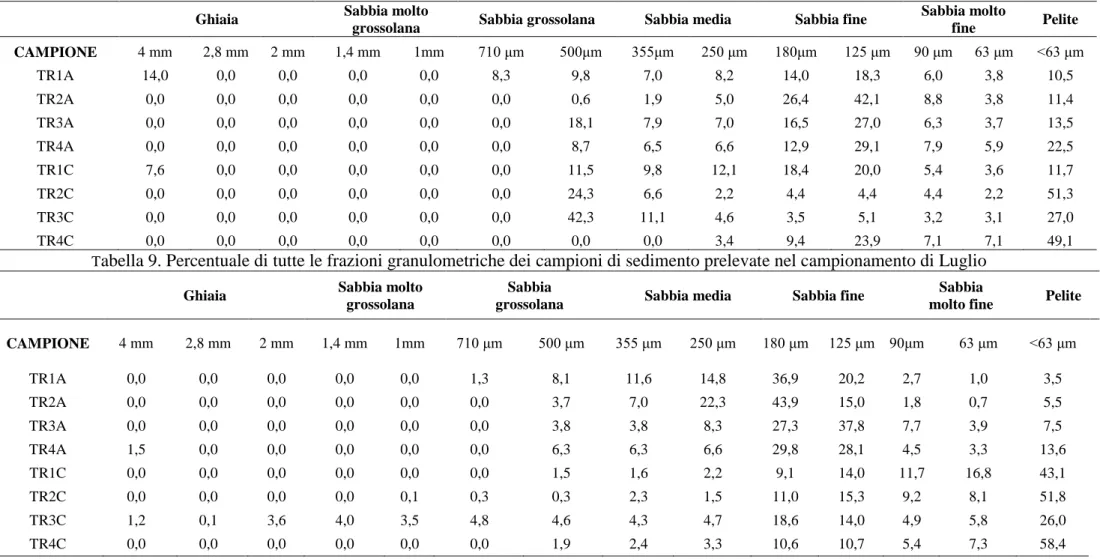
I campioni del sedimento superficiale (circa 2 etti ciascuno) sono stati raccolti, mediante operatore subacqueo, in corrispondenza del margine superiore della prateria (TR1A, TR2A, TR3A, TR4A) e del margine inferiore (TR1C, TR2C, TR3C, TR4C) (Fig 7) in Luglio (due repliche) e in Novembre (tre repliche). I campioni, raccolti nelle aree di intermatta o nel detrito di matte morta, sono stati liofilizzati, setacciati per raccogliere la frazione granulometrica < 250 µm, e, successivamente, sottoposti alle analisi per la determinazione di Hg, Cd, Pb, Cu, Ni, Cr, Zn e IPA.
Per i metalli i campioni sono stati inizialmente sottoposti a mineralizzazione mediante attacco acido in un sistema chiuso a microonde (ETHOS 900 Microwave Labstation, Milestone), in accordo con il metodo EPA 3051. In dettaglio, un’aliquota di campione di circa 0,5 g è stata digerita in acido nitrico concentrato (al 65%), utilizzando un programma di digestione che prevede il raggiungimento di una temperatura di 170°C entro 5,30 minuti ed il mantenimento della temperatura tra 170 e 175°C per 10 minuti.
31 Per la fase di quantificazione è stata seguita la stessa procedura descritta per i campioni della vegetazione.
Per la determinazione degli IPA si è fatto riferimento alle metodiche EPA 3545 A, 3610, 8310, già descritte.
5.2.4. Caratteristiche trofiche dell’ecosistema e tessitura sedimentaria
Determinazione della materia organica.
Un’aliquota di ciascun campione di sedimento raccolto è stata destinata alla determinazione del contenuto in materia organica (OM). Il metodo si basa sulla combustione in muffola (“loss-on-weight-by-ignition”) (Loh, 2005). L’aliquota è stata setacciata ad umido con maglia da 1 mm per rimuovere l’eventuale materiale grossolano presente (conchiglie e frammenti vegetali), quindi posta in stufa a 70°C fino a completo essiccamento. Successivamente il materiale è stato pesato e trasferito in un crogiuolo per l’incenerimento in muffola a 250 °C per 4 ore. Dopo il raffreddamento in essiccatore a temperatura ambiente, il materiale incenerito è stato pesato e riposto in muffola a 400 °C per 4 ore; una volta raffreddato se ne è registrato nuovamente il peso.
Il contenuto percentuale di materia organica labile (LOM), refrattaria (ROM) e totale (OM) è stato calcolato come segue:
(peso netto secco iniziale – peso netto a 250 °C)
OM LABILE % = __________________________________________· 100 peso netto secco iniziale
(peso netto a 250 °C – peso netto a 400 °C) OM REFRATTARIA % = _____________________________________ ·100
peso netto secco iniziale
32 Trappole sedimentarie
Per definire la dinamica sedimentaria all’interno dell’area di retroprateria e valutarne la frazione di OM presente, sono state collocate, da un operatore in ARA, evitando di sollevare il sedimento, sei trappole per la raccolta del materiale sedimentabile, in tre stazioni (TP1, TP2, TP4; Fig. 4), due per stazione, lungo il margine superiore della prateria di P.oceanica. La stazione TP1 controllava il flusso delle acque in entrata-uscita dalla retroprateria al margine sinistro della prateria emergente, la TP4 quello dell’entrata al margine destro e la TP2 costituiva una stazione interna alla retroprateria (Fig. 2, 4).
Le trappole sedimentarie sono state lasciate sul fondo per 3 giorni in Luglio e per una settimana in Novembre (quest’ultimo periodo in seguito al maltempo che ha impedito la raccolta delle trappole).
Tutto il materiale contenuto in ciascuna trappola è stato trasferito in un beker di plastica di opportuno volume, sciacquando ripetutamente e raccogliendo tutte le acque. Successivamente, il materiale è stato lasciato sedimentare e l’acqua sovrastante è stata tolta evitando di produrre disturbo al sedimentato. È stata poi aggiunta acqua bidistillata per diluire il sale presente, lasciando risedimentare e nuovamente eliminando l’acqua sovrastante. Il materiale rimasto sul fondo, sedimento e acqua residua, è stato trasferito in crogiolo e posto in stufa a 70 ºC fino a completo essiccamento. Una volta essiccato è stato rimosso e setacciato con una maglia da 1 mm, per eliminare eventuale materiale vegetale grossolano presente, quindi è stato registrato il peso. Il materiale essiccato è stato posto in muffola a 400°C, per 4h, e ne è stato successivamente registrato il peso delle ceneri residue. In Novembre, sono state determinate anche le frazioni LOM e ROM, secondo la metodica precedentemente descritta.
Determinazione delle frazioni granulometriche.
Per questa determinazione è stato fatto riferimento alla metodica ICRAM (Romano & Gabellini, 2001). I campioni di sedimento sono stati essiccati in stufa a 70 °C per 24 ore, pesati e mantenuti in condizioni di staticità per 48 ore, in una soluzione di acqua distillata e perossido di idrogeno (v.v., 2:8) per disgregare la sostanza organica e facilitare la separazione dei granuli. Successivamente i campioni sono stati lavati con acqua distillata e filtrati con un setaccio con maglia da 63 µm per separare la frazione psammitica (> 63 m) da quella pelitica (< 63 m). Dopo il lavaggio la frazione psammitica è stata posta in stufa a 70°C fino a completo essiccamento; quindi è stata analizzata con il metodo del setacciamento per separare
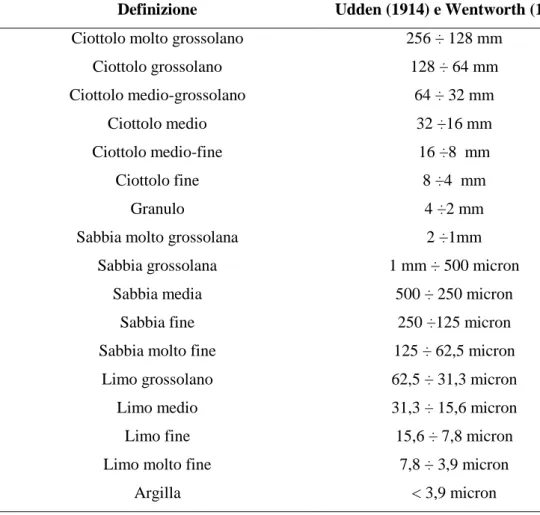
33 le varie frazioni granulometriche. Il setacciamento è stato effettuato tramite una batteria di setacci in acciaio inox (serie ASTM), scelti in modo da costruire una serie a maglia con diametri da -2φ a +4φ, decrescenti verso il basso e con intervallo di 0,5 phi (phi = -log2 del valore in mm) (Udden, 1914; Wentworth, 1922; Krumbein, 1934) (Tab 6.). La pila di setacci è stata posta su un agitatore meccanico per circa 20 minuti, le frazioni trattenute in ogni setaccio sono state pesate e i risultati espressi come percentuale del peso totale iniziale. La frazione pelitica, setacciata ad umido non è stata pesata, ma quantificata per differenza. I dati delle analisi sono stati utilizzati per determinare la distribuzione percentuale delle classi granulometriche e per l’individuazione delle principali frazioni dimensionali riportate in Tabella 7.
Carbonio organico.
La quantificazione del carbonio organico (total organic carbon, TOC) si basa sulla combustione della frazione <250μm di un’aliquota di ciascun campione di sedimento, mediante K2Cr2O7 in ambiente acido per H2SO4 concentrato e successiva titolazione indiretta (metodo Walkley Black modificato da Gaudette et al., 1974). Con tale metodo viene ossidato soltanto il carbonio più attivo, pari al 77% del totale. La quantità di K2Cr2O7 che è consumata durante la reazione è proporzionale al tenore di carbonio organico presente nel campione. L’eccesso di bicromato che non ha reagito durante l’ossidazione viene titolato con una soluzione di sale di Mohr (solfato ferroso) in presenza di difenil-ammina. Quando tutto l’eccesso è stato neutralizzato dal sale aggiunto, il colore della soluzione vira dal blu scuro al verde brillante. Per ogni serie di misure viene aggiunta anche una prova in bianco. La quantità di sale di Mohr consumata durante la titolazione è proporzionale al contenuto di carbonio organico del campione come evidenziato dalla seguente relazione:
% C org. = 10 · (1-T/S) · [1,0 N · (0,003)] · (100/w).
Dove: T sono i ml di soluzione ferrosa necessari per titolare il campione; S quelli per titolare la prova in bianco; 10 i ml di bicromato aggiunti al campione; 0,003 (= 12/4000) i meq di carbonio; 1,0 la normalità (N) del bicromato e w il peso del campione di sedimento espresso in grammi.
34 Azoto e Carbonio Totali
La determinazione del carbonio ed azoto totali (TC e TN) è stata effettuata secondo metodo ICRAM (Giani, 2001). Per la quantificazione del TC e del TN percentuali è stato utilizzato un analizzatore elementare (Perkin Elmer CHNS/O 200). Come standard si è utilizzato acetanilide; la calibrazione iniziale è stata effettuata con 4 standard di acetanilide e 2 capsule vuote a cadenza giornaliera. Quindi sono state effettuate ricalibrazioni ogni 12 campioni analizzati con 2 standard di acetanilide per ricalcolare il fattore di calibrazione ed un bianco il cui valore viene sottratto al valore di ogni singolo campione. Le concentrazioni di TC e TN sono state espresse come percentuali in peso dell’elemento sul peso secco del campione. Il limite di determinazione del metodo è di 0,01%. Relativamente ai campioni vegetali, l’azoto totale è stato denominato TON, per distinguere la variabile da quella determinata nei sedimenti.
Fosforo Totale
L'analisi del fosforo totale (TP) nel sedimento è stata effettuata in accordo al D.M. 185/99. Il metodo prevede che 0,5 g di sedimento secco siano sottoposti a mineralizzazione con acido solforico e perossido di idrogeno. La soluzione è stata alcalinizzata con NaOH fino al viraggio dell'indicatore p-nitrofenolo.
I campioni sono stati analizzati per via spettrofotometrica alla lunghezza d'onda di 882 nm, sfruttando la reazione colorimetrica dello ione fosfato con molibdato di ammonio. Con le stesse modalità operative adottate per i campioni è stata eseguita anche una prova in bianco dei reattivi e la mineralizzazione di matrici di sedimento certificate, con concentrazioni indicativamente dell’ordine di grandezza di quelle presunte nel campione. I recuperi registrati nel corso delle indagini su matrici di sedimento certificate sono compresi tra il 95 e il 100%. I risultati finali sono stati espressi come percentuale in peso del fosforo sul peso secco. Il limite di rilevabilità del metodo è pari a 0,001%.
Per la determinazione del fosforo totale nei campioni vegetali (che, per differenziarlo da quello dei sedimenti viene denominato TOP), viene effettuata una mineralizzazione ad ortofosfato mediante digestione a caldo con acido solforico, quindi portato a reagire con molibdato ammonico per la formazione di un eteropoliacido, che successivamente è stato ridotto con acido ascorbico a blu di molibdeno, quantificato per via spettrofotometrica (Murphy & Riley, 1962).