Analisi dell'isolamento riproduttivo nelle orchidee mediterranee
129
0
0
Testo completo
(4)
(5)
(6)
(7)
(8)
(9)
(10)
(11)
(12)
(13)
(14)
(15)
(16)
(17)
(19)
(20)
(21)
(22)
(23)
(24)
(25)
(26)
(27)
(28)
(29)
(30)
(31)
(32)
(33)
(35)
(36)
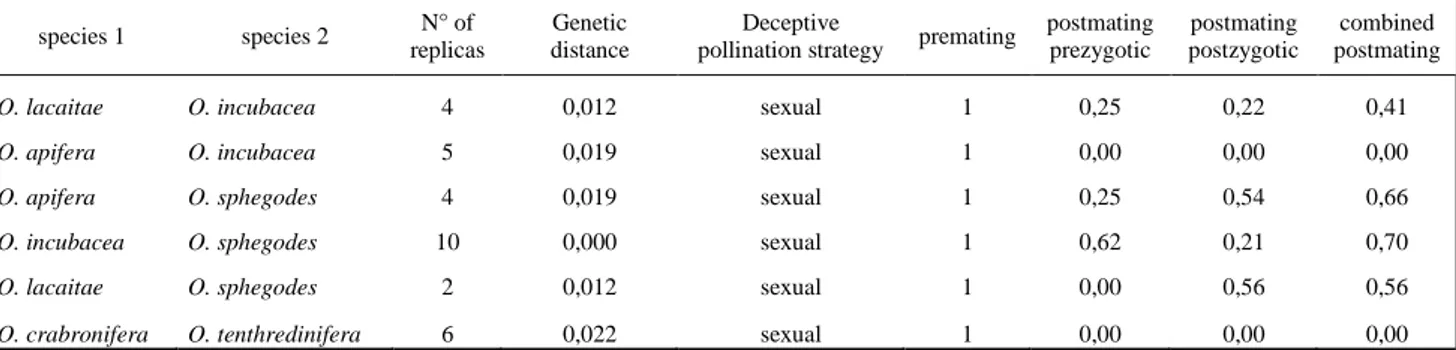
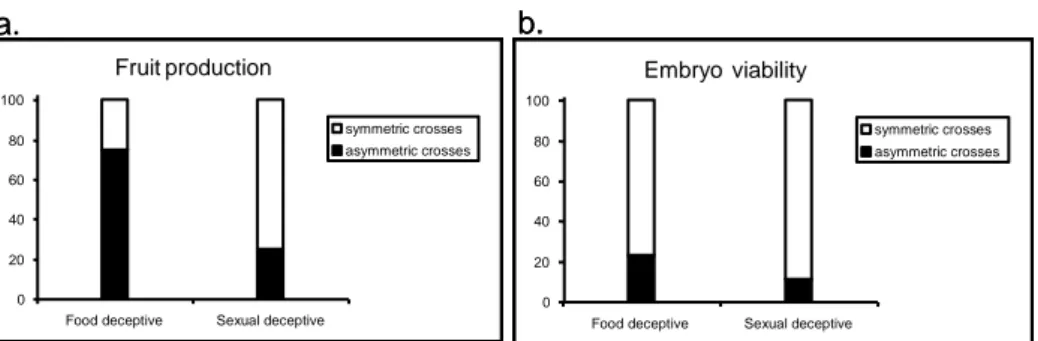
Figura
Documenti correlati
City Focus is a webbased interactive 2D and 3D GIS application to find the best place in a city to live, or to pass shorter staying. The user can select among different
In order to describe abundance and prevalence of FD-infected vectors in the cultivated and wild compartments of the vineyard agroecosystem, adults of Scaphoideus titanus were
The E-learning applied to a PE Teaching and Learning Methods and Styles lab-based ex- periment has surely entailed greater planning flexibility which “actually came in handy” when
Genotype E is found in Africa, in the sub-Saharan region, genotype F in Central and South America, genotype G in Eastern and Western Europe and genotype H, a variant
Issues such as the harmonisation between national and universal interests, the stipulation of international principles of justice, the codification of the
The American Association of Clinical Endocrinologists, the Obesity Society, and the American Society for Metabolic and Bariatric Surgery recommend the use of at least 3000 IU daily
It is hard to consider all those values in order to distinguish which SNPs are actually polymorphisms or sequencing errors, but VCF parameters can be a lot
In un secondo capoverso si richiama la giurisprudenza che in passato ha riconosciuto elementi con valenza superiore rispetto alle altre norme o leggi di rango
