Alteraciones de la bomba de calcio
en la hipertensión arterial esencial
Javier Sobrino Martínez
Aquesta tesi doctoral està subjecta a la llicència Reconeixement 4.0. Espanya de Creative
Commons.
Esta tesis doctoral está sujeta a la licencia Reconocimiento 4.0. España de Creative
Commons.
DE
CALCIO
EN LA
HIPERTENSION
ARTERIAL
ESENCIAL
Tesis
presentada
por Javier Sobrino Martínez
para
aspirar
al
Grado
de Doctor
enMedicina
Marzo,
1998
Facultad de Medicina
Universidad de Barcelona
Titular del
Departamento
de Medicina de laUniversidad de
Barcelona,
CERTIFICA: Que la tesis doctoral que lleva
por título "Alteraciones de la bomba de calcio en la
hipertensión
arterialesencial",
realizada por D.Javier Sobrino Martínez y
dirigida
por el quesuscribe,
está en condiciones de ser leída ante eltribunal
correspondiente.
Lo que se hace constar a los efectos
del
Departamento
de Medicina de la Universidadde
Barcelona,
CERTIFICA: Que la tesis doctoral que lleva
por título "Alteraciones de labomba de calcio en la
hipertensión
arterialesencial",
realizada por D.Javier Sobrino Martínez y
dirigida
por el quesuscribe,
está en condiciones de ser leída ante eltribunal
correspondiente.
Lo que se hace constar a los efectos
Profesor Asociado del
Departamento
de Medicinade la Universidad de
Barcelona,
CERTIFICA: Que la tesis doctoral que lleva
por título "Alteraciones de la bomba de calcio en la
hipertensión
arterialesencial",
realizada por D.Javier Sobrino Martínez y
dirigida
por el quesuscribe,
está en condiciones de ser leída ante eltribunal
correspondiente.
Lo que se hace constar a los efectos
Al Prof. Dr. Antonio
Coca,
director de lapresente
TesisDoctoral,
que meofreció la
posibilidad
de iniciarme comoinvestigador
en el estudio de lahipertensión
arterial y pusoa mi
disposición
los medios técnicos necesarios para ello.Además,
sinsu constante estímulo y su
participación
en la elaboración final del manuscrito estatesis no se habría realizado.
Al Dr.
Alejandro
de laSierra,
codirector de esta TesisDoctoral,
artífice de lahipótesis
detrabajo
delproyecto,
por su constanteayuda,
apoyo y colaboración en laestructuración,
elaboración y redacción de laTesis,
así como por su constanteestímulo y
exigencias.
Al Prof. Dr. Alvaro
Urbano-Márquez,
codirector de esta TesisDoctoral,
por laconfianza
depositada
en mitrabajo
asistencial einvestigador.
A la Dra. M. T.
Aguilera,
por su constante apoyo y suayuda
en los distintosaspectos
de las técnicas de laboratorio.Al Dr.
Miquel
Torres,
por su constante estímulo para la finalización de estaTesis.
Al
personal
del Laboratorio deBioquímica,
Hormonología
yToxicología,
por sudesinteresada colaboración en el
procesamiento
de las muestrassanguíneas
ypermitirme
la utlización de sus instalaciones.A todos los
pacientes
queconstituyen
el material humano de esta tesis.Al
Hospital
Clínici Provincial de Barcelona por la concesión de una Beca queha hecho
posible
la realización delproyecto
deinvestigación
queconstituye
esta tesisl.
Justificación
111. Revisión
bibliográfica
51.
La
hipertensión
arterial esencial
comoenfermedad de la
membrana celular
61.1. Introducción 6
1.2. El sodio en la
etiopatogenia
de lahipertensión
arterial esencial 101.2.1. Aumento de la concentración intracelular de Na+ 11
1.2.2. Alteraciones de la difusión
pasiva
de Na+ 121.2.3. Alteracionesen la ATPasa Na+-K+ 12
1.2.4. Alteraciones en el
cotransporte
Na+ -K+-CI" 141.2.5. Alteraciones en el
contratransporte
Na+ -L¡+ 151.2.6. Alteracionesen el intercambiador Na+-H+ 15
1.2.7. Alteracionesen el intercambiador
HC03"¡CI"
161.2.8. Alteraciones en el intercambiador Na+
_Mg2+
172. Metabolismo celular del calcio
182.1. Introducción 18
2.2. Los canales de calcio 19
2.3. La ATPasa
Ca2+-dependiente.
La bomba deCa2+
202.3.1. Estructura y funcion de la bomba de calcio 20
2.3.2. La interaccion de la bomba de calcio con la calmodulina 24 2.3.3.
Regulacion
de la bomba de calcio 262.3.4. La bomba de calcio del reticulo
sarcoplasmico
272.4. El
contratransporte
Na+-Ca2+
283.
Hipertensión
arterial y calcio
303.1.
Papel
del consumo de calcio en lahipertension
arterial 303.1.1. Evidencias
epidemiológicas
313.1.2. Marcadores
biológicos
323.1.3. Estudios de intervención 33
3.2. El calcio intracelular en la
hipertensión
arterial 343.3. Anomalías de la bomba de calcio en la
hipertensión
arterial 373.3.1. Alteraciones de la
fijacion
de calcio por las membranas celulares. 37 3.3.2. Alteraciones de la bomba de calcio en lahipertension
arterialexperimental
, 393.3.3. Alteraciones de la bomba de calcio en la
hipertension
arterialesencial . . . .. 44
pacientes
1.1.
Grupo
control
511.2.
Grupo
de
pacientes
conhipertensión
arterial esencial
512.
Métodologías
522.1. Medida de la
presión
arterial
522.2.
Metodología
del análisis
bioquímico
y hormonal
532.3.
Metodología
del
ecocardiograma
532.4.
Determinación de los
parámetros
cinéticos de la ATPasa
Ca2+
-dependiente
542.4.1.
Preparación
de los eritrocitos 552.4.2. Procedimiento de carga 55
2.4.3. Determinación de
Sr2+
yCa2+
intraeritrocitarios 562.4.4. Determinación del
flujo
deSr2+
562.4.4. Cálculo de la afinidad
aparente
de la ATPasaCa2+
-dependiente
para el
Sr2+
intracelular y de la velocidad máxima de la salida deSr2+
572.4.5. Calculo de la afinidad
aparente
para elCa2+
intracelular 593.
Análisis estadístico
613.1. Pruebas
estadísticas
utilizadas
613.2.
Soporte
informático.
. . . .. 61V.
Resultados
631.
Características
generales
de la serie
641.1.
Grupo
control
641 .1 .1.
Descripción
de losparámetros
641.2.
Grupo
de
pacientes
conHTA esencial
681.2.1.
Descripción
de losparámetros
682.
Diferencias
entre
los
parámetros
cinéticos obtenidos
enel grupo control yen los
pacientes
conHTA esencial
.. 723. Relación de los
parámetros
cinéticos de la bomba de
calcio
conlos
parámetros
analizados
743.1.
Serie
global
743.1.1. Relacion de la velocidad máxima del
flujo
deSr2+
conlos
parámetros
estudiados . . . .. 743.1.2. Relacion de la constante de disociación
aparente
para elCa2+
intracelular con los
parámetros
estudiados 753.2.
Grupo
de
pacientes
conHTA esencial
763.2.1. Relacion de la velocidad máxima del
flujo
deSr2+
conlos
4.
Clasificación de los
pacientes
conHTA esencial
795.
Diferencias
clínico-biológicas
entre los
subgrupos
de
pacientes hipertensos
80VI.
Discusión
86VII.
Resumen y conclusiones
90Tabla 1. Valores medios
(± DE)
de losparámetros
clínicos, bioquímicos
yecocardiograficos
básicos en los 27 individuos queconstituyen
el grupocontrol 65
Tabla 2 Valores medios
(±
DE)
de losparámetros
cinéticos de la bomba de calcio enlos 27 individuos que
constituyen
el grupo control 66Tabla 3 Valores medios
(± DE)
de losparámetros clínicos, bioquímicos,
hormonalesy
ecocardiograficos
básicos en los 49 individuos queconstituyen
el grupo depacientes hipertensos
esenciales 69Tabla 4 Valores medios
(± DE)
de losparámetros
cinéticos de la bomba de calcio enlos 49 individuos que
constituyen
el grupo depacientes
hipertensos
esenciales 70
Tabla 5. Parámetros que muestran una relación directa con la velocidad máxima del
flujo
deSr2+
74Tabla 6. Parámetros que muestran una relación directa con la constante de
disociación
aparente
para elCa2+
intracelular 75Tabla 7.
Comparación
de los diferentesparámetros
clínicos entre lospacientes
hipertensos
esencialescon constante de disociaciónaparente
para elCa2+
intracelular normal o elevada 80
Tabla 8.
Comparación
de los diferentesparámetros bioquímicos,
hormonales yecocardiográficos
entre lospacientes
hipertensos
esenciales con constante de disociaciónaparente
para elCa2+
intracelular normal o elevada . . . .. 84Figura
1. Ciclo de reacción de la ATPasaCa2+-dependiente
, 24Figura
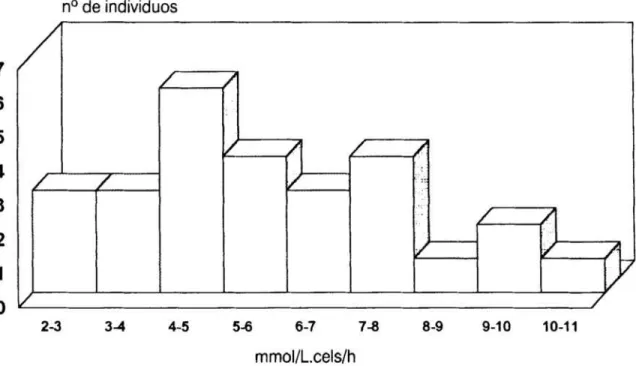
3.Histograma
de la distribución en frecuencias de los valores individuales de la velocidad máxima delflujo
desr2+
en los 27 individuos queconstituyen
elgrupo control 66
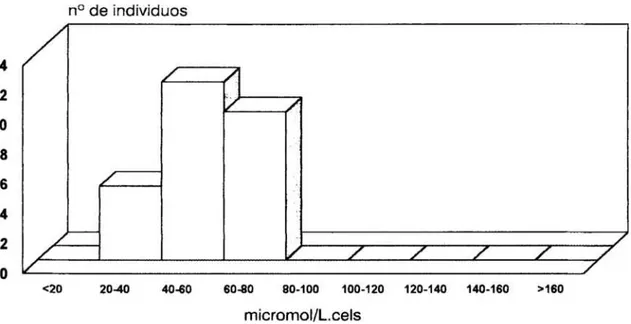
Figura
4.Histograma
de la distribución en frecuencias de los valores individuales de la constante de disociaciónaparente
para elCa2+
intracelular en los 27individuos que
constituyen
el grupo control 67Figura
5.Histograma
de la distribución en frecuencias de los valores individuales dela velocidad máxima del
flujo
deSr2+
en los 49 individuos queconstituyen
elgrupo de
pacientes hipertensos
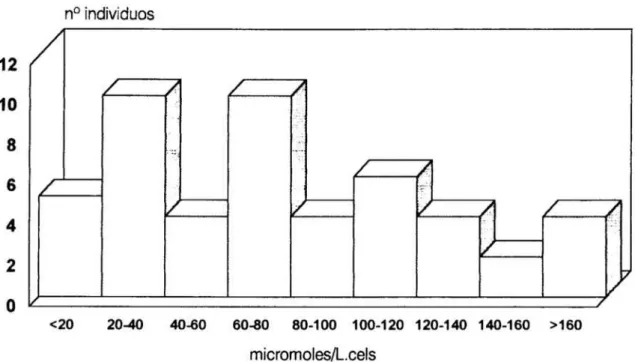
esenciales , 70Figura
6.Histograma
de la distribución en frecuencias de los valores individuales dela constante de disociación
aparente
para elCa2+
intracelular en los 49 individuos queconstituyen
el grupo depacientes hipertensos
esenciales 71Figura
7.Diagrama
de barras de los valores medios(±
desviaciónestándar)
de lavelocidad máxima del
flujo
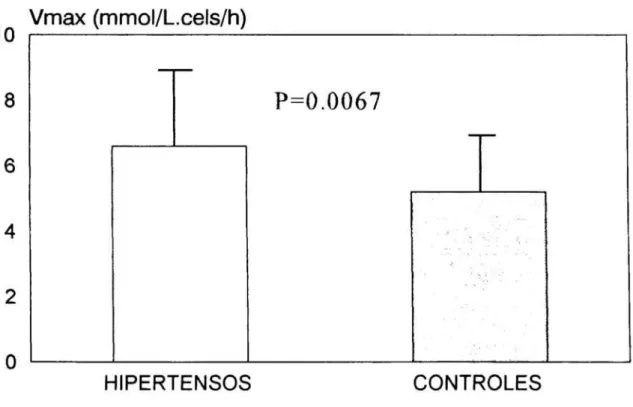
deSr2+
en los 49hipertensos
esenciales y los 27controles 72
Figura
8.Diagrama
de barras de los valores medios(±
desviaciónestándar)
de laconstante de disociación
aparente
para elCa2+
intracelular totalO<ca)
en los49
hipertensos
esenciales y los 27 controles 73Figura
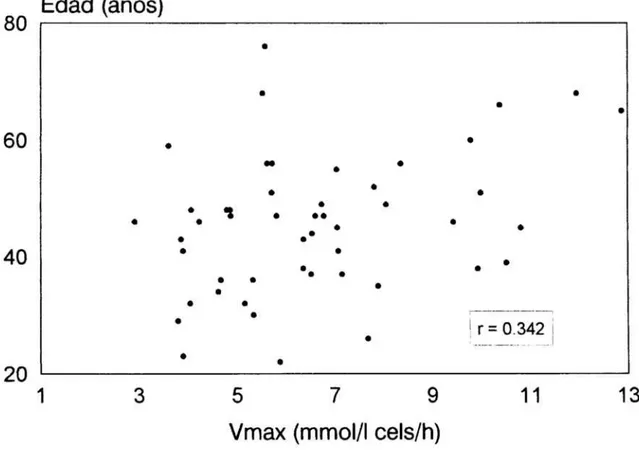
9. Correlacion directa entre la velocidad máxima delflujo
deSr2+
y la edad delos
pacientes hipertensos
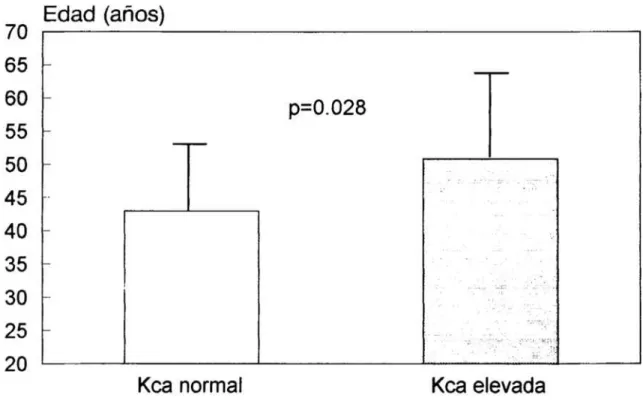
esenciales 76Figura
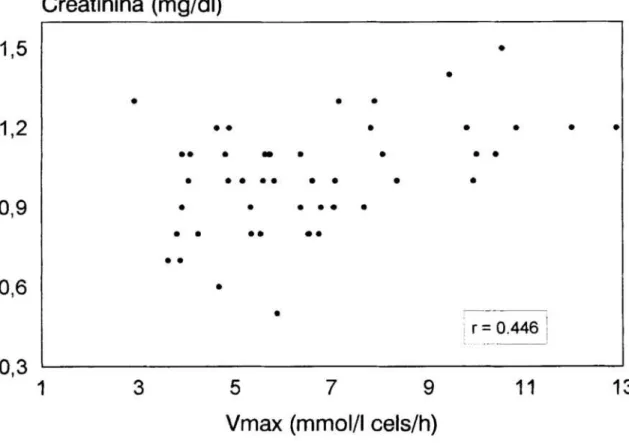
10 Correlacion directa entre la velocidad máxima delflujo
deSr2+
y las cifras decreatinina de los
pacientes hipertensos
esenciales 77Figura
11 Correlacion directaentre la constante de disociaciónaparente
para elCa2+
intracelulary las cifras de creatinina de los
pacientes hipertensos
esenciales. . . .. 78
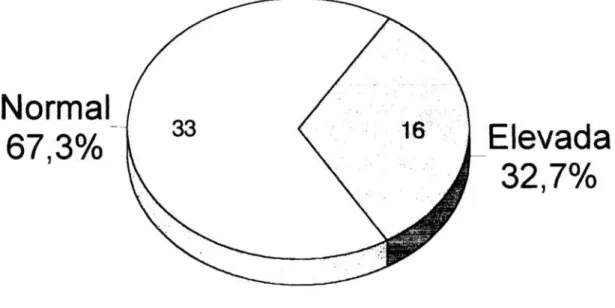
Figura
12 Gráfico de sectores que muestra laprevalencia
de la alteración de la constante de disociaciónaparente
para elCa2+
intracelular en lapoblación
constante de disociación
aparente
para elCa2+
intracelular 81Figura
14Diagrama
de barras que muestra los valores medios(±
desviaciónestándar)
de la creatinina
plasmática
en los dossubgrupos
depacientes hipertensos
esenciales clasificados enfunción de la constante de disociación
aparente
para el
Ca2+
intracelular 82Figura
15Diagrama
de barras que muestra los valores medios(±
desviaciónestándar)
del ácido úrico en los dos
subgrupos
depacientes
hipertensos
esenciales clasificados en función de la constante dedisociaciónaparente
para elCa2+
intracelular 83
Figura
16Diagrama
de barras que muestra los valores medios(±
desviaciónestándar)
de la velocidad máxima del
flujo
deSr2+
en los dossubgrupos
depacientes
hipertensos
esenciales clasificados en función de la constante deAMP AMPc ATPasa Bomba
(-)
HTA HTAe Hto Adenosinmonofosfato Adenosinmonofosfato cíclico AdenosintrifosfatasaPaciente con disminución de la afinidad
aparente
de la ATPasa
Ca2+
para elCa2+
intracelularHipertensión
arterialHipertensión
arterial esencialHematocrito
Constante de disociación
aparente
para elCa2+
Presión arterial
Presión arterial diastólica Presión arterial media
Presión arterial sistólica
Hormona
paratiroidea
Retículo
endosarcoplásmico
Ratas
espontaneamente
hipertensas
Velocidad máxima
Ratas normotensas
Wistar-Kyoto
Kca
PA PAD PAM PAS PTH RS SHR WKYLa
hipertensión
arterial(HTA)
esencial es la enfermedad másfrecuente de cuantasafectan a la humanidad y su
prevalencia
se cifra entre el 20 y el 25 por ciento de lapoblación
adulta. Gran númerode estudios demuestran que a mayorpresión
arterial(PA),
mayores son la morbilidad y la mortalidad de causacardiovascular;
lo queavala el hecho de que
constituya
uno de losprincipales
factores deriesgo
cardiovascular. La evidencia acumulada hasta el momentohace suponer que la HTA
esencial se desarrolla por la acción combinada de dos grupos de factores: factores
genéticos,
demostrados por la existencia de unaimportante
agregación
familiar deesta enfermedad y factores ambientales diversos. Entre estos últimos se han
implicado
las dietas ricas en clorurosódicoypobres
encalcio,
laingesta
alcohólica,
las dietas
hipercalóricas
y el estrés.En la última decada ha existido un creciente interés ante la
posibilidad
de que laHTA esencial
pueda
estar causada poruna alteración en la homeostasis intracelularde
algunos
iones,
especialmente
el Na+ y elCa2+.
El avanceproducido
en elconocimiento del metabolismo iónico a nivel celular y más
concretamente,
de losmecanismos que
regulan
sutransporte
a través de las membranascelulares,
hapermitido
la detección de diversas anomalías caracterizadas por un déficit en lafunción de las
proteinas transportadoras.
Este hecho hapermitido
sentar las basesmoleculares de la relación existente entre el consumo de sal y la HTA esencial.
En el Servicio de Medicina Interna General del
Hospital
Clinic i Provincial deBarcelona ha existido desde hace ya varios años un marcado interés en la
investigación
de los mecanismosimplicados
en laetiopatogenia
de la HTA esencial.El estudio del
transporte
de Na+ a través de la membranacelular fué iniciado por eldirector de esta
tesis,
el Dr. A Coca1,
que demostró la existencia de una alteracióndel
flujo
catiónico activo en los eritrocitos de lospacientes hipertensos
esencialesyde sus
hijos
normotensos.Posteriormente,
el Dr. A de laSierra2 pudo
clasificar alos
hipertensos
esencialesensubgrupos
atendiendo a lasanomalíasen los sistemasde
transporte
transmembranario de Na+ y a la existencia de características clínicasdiferenciales. La
presente
tesis doctoral continua la línea iniciada por esteGrupo
deInvestigación
ypretende
aportar
nuevosaspectos
sobre laimplicación
del Na+ ysurelación con el
Ca2+
ysus sistemas detransporte,
dado que el contenido intracelularde estos dos ionesy su
regulación
parecen estar interrelacionados. Puesto que elNa+ intracelular
ejerce
su efecto presora través del incremento en el contenido deCa2+,
resultalógico
pensar que una alteración en los mecanismos queregulan
eltransporte
de este catión y, más concretamente la ATPasaCa2+-dependiente,
pueden
tenerrelación con laetiopatogenia
de la HTA. Cabe decir alrespecto
queunode los miembros de nuestrogrupo ha
puesto
de manifiesto la existencia de unaalteración en la función cinética de dicha ATPasa en un
pequeño
número depacientes
hipertensos
esenciales".
La
presente
tesis doctoralpretende
demostrar la existencia de unsubgrupo
dehipertensos
esenciales caracterizados por anomalías de la ATPasaCa2+
dependiente
entre lospacientes
afectos de HTA esencial de nuestro medio.No
obstante,
este estudio no se detiene a un nivelpuramente
experimental.
Cabeesperar que con una muestra
amplia,
estesubgrupo
dehipertensos
esencialespresente
características clínicas que lo diferencie del resto dehipertensos,
hechoque parece vislumbrarse en nuestros
trabajos previos,
aunque las muestrasutilizadas han sido
generalmente
pocosignificativas.
Laposibilidad
de estableceruna correlación clínico-celular
supondría
un pasoimportante
hacia la redefinición dela HTA esencial como un
síndrome,
en el que se incluirían diferentes entidadesnosológicas
diferenciables tanto en elaspecto
clínico como en el de labiología
celular.
1
.La
hipertensión
arterial
esencial
como
enfermedad de la membrana celular.
1.1.
Introducción.
El conocimiento acumulado hasta la
actualidad,
hace suponer que lahipertensión
arterial esencial
(HTAe)
se desarrolla por la acción combinada de al menos dosgrupos defactores: factores
genéticos
y factores ambientales de diversaíndole,
entre los que se han
implicado
el excesivo consumo de sal y las dietas con unpobre
contenido encalcio,
magnesio
ypotasio.
La
posibilidad
de que la HTAepueda
ser causada por una alteración en lahomeostasis intracelular de estos
iones,
ha sido motivo de un creciente interésen la última década. El avance
producido
en el conocimiento del metabolismoiónicoa nivel celular y, más
concretamente,
de los mecanismos queregulan
sutransporte
a través de las membranascelulares,
ha hechoposible
la detecciónde diversas anomalías caracterizadas porun deficit en lafunción de las
proteinas
transportadoras.
Estas alteraciones descritas a nivel del metabolismo iónicocelularhan
permitido
sentar lasbases moleculares de la relación existenteentreel consumo de
sodio, calcio,
magnesio
ypotasio,
y la HTAe.El
progresivo
incremento de las cifras depresión
arterial(PA)
que se observa conel aumento de edad en las civilizaciones desarrolladas parece ser un evento
alimentos".
En los últimos 5.000años,
el desarrollo de latecnología
necesariapara la obtención de la sal común
procedente
de las minas o del agua del mar hapermitido
al serhumano la conservación de los diferentes alimentos enforma desalazón. En aras a evitar el crecimiento bacteriano y
fúngico,
la conservaciónmediante sal común
requiere
de altas concentraciones de cloruro sódico porgramo de
tejido.
Probablemente,
elpaladar
de las sucesivasgeneraciones
se haido
adaptando
de maneraprogresiva
a estas altas dosis y su resultado ha sidoel uso de sal como
apreciado
condimento. Este hechopodría
serlaexplicación
de que el consumo habitual de cloruro sódico sea del orden de 10 a 30 veces
superior
a las necesidadesfisiológicas
del serhumanos.
Ambard y
Beauiard"
sugirieron,
hace ya más de 80años,
que el excesivoconsumo de sal en la dieta
podía
serelresponsable
del aumento de las cifras dePA de
algunos
individuos. Desdeentonces,
gran numero de estudiosepidemiolóqicos'", experímentales'?"
ycnnícos":"
han dadosoporte
a lasprimeras
observaciones y corroborado lainequívoca
relación entre el consumode sal y la HTAe.
El intento de
extrapolar
conclusionesemergentes
de datosepidemiológicos
aexperiencias
individuales en la clínica hagenerado
innumerables controversias.Desde un
punto
de vistageneral,
lahipótesis "sal-hipertensión"
sostiene que lamayoría
de los humanos son sensibles a laexposición
a la sal. Sinembargo,
únicamente
algunos
individuosresponden
con descensos depresión
frentea lasimilar a lo que sucede en el animal de
experimentación".
Contrariamente a la sal
común,
cuya relación con la HTA sesospecha
desdehace casi 100
años,
lasprimeras
observaciones que relacionaban el consumo decalcio con el
riesgo
cardiovascular fueronpublicadas
hace apenas tresdecadas
16.
Estostrabajos
inicialespermitieron
demostrar que las comunidadesque consumían un agua másdura
presentaban
indices másbajos
de muerteporenfermedad
cardiovascular":".
En los últimos
años,
diversos estudiosepiderniolóqicos'P'"
han identificado elbajo
consumo de calcioen la dieta como unfactor deriesgo
para el desarrollo dela HTA. Esta relación ha
persistido
inclusodespués
de controlar variables talescomo la
edad,
raza, sexo, pesoy consumo de alcohol. Otrosfactoresimplicados
por dichos
estudios,
tales como las dietas deficitarias enpotasio,
magnesio
ofósforo
pueden
serexplicadas
por el hecho de que laprincipal
fuente de calciode la alimentaciónes tambien la
principal
fuente de estos otroscomponentes.
Enlíneas
generales puede
decirse que un persona queconsume por término mediouna dieta con 300 mg diarios de calcio tiene un
riesgo
para el desarrollo de HTAentre un 11% Y un 14%
superior
a otro de su mismaedad,
sexo y raza queconsuma 1200 mg diarios
19.
A esta evidencia de
tipo
epidemiológico
hay
que añadir otra derivada de laexistencia de
algunos
transtornos del metabolismo del calcio detectados enpacientes
y animales afectos de HTAe. Entreellos,
el másimportante
es elMcCarron et
al26
demostró una disminución del calcio sérico total en lospacientes
hipertensos
mientras que,paradójicamente,
enalgunos
otros se puso enevidencia una correlación
positiva
entre los valores de PA y el calciosérico":
Estas observaciones contradictorias
podrían
ser debidas a la falta decorrección,
en estos
últimos,
de la relativa hemoconcentraciónpresente
en muchospacientes
hipertensos,
de modo que el aumento de los niveles de calcio sérico total seríanreflejo
del aumento de la concentración dealbúmina".
Tal comológicamante
podrían
predecir
unosniveles de calcio iónico séricobajo,
los niveles de hormonaparatiroidea
se han encontrado elevados enpacientes
hípertensos":",
en estosestudios,
el aumento de la hormonaparatiroidea
se asociaba a un aumento enla excreción urinaria de AMP cíclico. Otro indicador de la alteración del
metabolismo del
calcio,
el descenso en los niveles de fosfatosérico,
ha sidoobservada en la
HTAe22,26,29,30
altiempo
que la existencia de una correlacióninversa entre niveles de fósforo y valores de
presión
arteriaI30,31.
Otra evidencia adicional que relaciona el calciocon la HTA deriva de los estudios
terapéuticos
del grupo deMcCarron32
en los que se demuestra un descenso enlas cifras de PA en casi la mitad de los
hipertensos
esenciales tratados consuplementos
de calcio.Asimismo,
Erne etal33
demostraron un incremento de laconcentración
intraplaquetaria
de calcio enpacientes
afectos de HT Ae.En resumen, los datos de los que
disponemos
hasta la actualidad no sonconcluyentes
por lo querespecta
a la existencia de una clara relación entre unEn este
sentido,
aligual
que ocurrió hace años con elsodio,
la mayor evidenciaque
implica
dicha alteración como causa de la HTA hasurgido
de los estudios anivel del metabolismo celular de este catión.
1.2.
El
sodio
enla
etiopatogenia
de la HTA
La
hipótesis
de que la HTA esencial se relacionaba con alteraciones delcontenido intracelular de
Na',
fue descrita porprimera
vez por Tobian yBiniorr"
en
1952,
al detectar en exámenes"postmortem",
un aumento del contenidointracelular de Na+ enla fibras musculares lisas de arterias renales
procedentes
de individuos
hipertensos
esenciales. Desdeentonces,
y hasta nuestrosdías,
sehan confirmado
algunos
de los resultadosiniciales,
y se han caracterizadodiferentes anomalías
genéticas
de los sistemas enzimáticos que catalizan eltransporte
de Na+ através de las membranascelularesy,asimismo,
se hapodido
detectar la
presencia
de sustanciasplasmáticas
circulantes capaces de modularla actividad de estos sistemas en los
hipertensos
esenciales".
A pesar de quela gran
profusión
de datos referidos en la literatura médica muestran enocasiones resultados
contradictorios,
las anomalías descritas tienden a provocar,en la
mayoría
de los casos, aumentos en la concentración intracelular desodio".
La HTAees unaentidad clínica mediada eminentemente por un aumento de las
resistencias
periféricas,
lo que en gran medida es debido al aumento del tono deen los
pacientes
hipertensos,
así como losproblemas
éticos queplantea,
hancondicionado la utilización de células hemáticas circulantes en la
investigación
clínica,
especialmente
hematíes,
por su fácil obtención ymanejo.
Aunque
estascélulas no se hallan directamente
implicadas
en los mecanismos queregulan
lapresion arterial,
suestudio debe considerarse como un modelo de lo que ocurreenotros
lugares
delorganismo.
Esde destacar alrespecto,
que lamayoría
de lossistemas de
transporte
transmembranario de Na+ son comunes a todas lascélulas del
organismo
y que las alteraciones halladas en hematíes y leucocitosse han
reproducido
encélulas de fibras musculares lisasvasculares,
células deltúbulo renal y neuronas
noradrenérgicas procedentes
de animales deexperimentación.
1.2.1. Aumento de la
concentración intracelular
de
Na+
En
1960,
Losse etal37
observaron un aumento del contenido de Na+ en hematíesde individuoscon
hipertensión
esencial. Desdeentonces,
numerosos autores hancorroborado dichos
hallazgos,
tanto enpacientes
híoertensos'":"
como en ratasespontaneamente
hipertensas",
e incluso en individuos normotensos conantecedentes en familiares de
primer grado
deHTAe42.
No
obstante,
dado que el metabolismo celular del Na+depende
básicamante desus sistemas de
transporte
transmembranario,
el aumento de contenido1.2.2.
Alteraciones
de la
difusión
pasiva
de
Na+
Garay
yNazaret?
demostraron la existencia deun aumento delflujo
de salida deNa+ por difusión
pasiva
en eritrocitos dealgunos
hipertensos
esenciales,
yconsiguieron
así caracterizar a unsubgrupo
depacientes
cuyos valores dedifusión
pasiva
eran claramentesuperiores
a los límites más altos de lanormalidad. El defecto de
permeabilidad pasiva
quepresentaban
los hematíesde estos
pacientes
ha sido definido como anomalía "Leak(+)".
Este aumento ha sido confirmadotambién por Wessels y
Zurnkley",
así como porDe la Sierra et
a145,
ypuede
ser detectado entre el 1% y el 12% de loshipertensos
esenciales de nuestrapoblación".
El que en estospacientes
laconcentración intracelular de Na+ sea normal o
alta,
puede
depender
de laexistencia y
magnitud
deun aumento simultáneo de las velocidades de extrusióndeNa+ por
parte
de la ATPasa Na+-K+ y delcotransporte Na+-K+-CI-,
que actúancomo sistemas
compensadores
de esta anomalía.1.2.3.
Alteraciones
enla ATPasa
Na+-K+
La bomba de sodio se halla virtualmente
presente
en todas las membranas delorganismo
y elflujo
de Na+dependiente
de éllapuede
variar por la acción desustancias moduladoras de su
actividad,
por modificaciones en el número deunidades
proteicas
de bomba de la memebranacelular,
o por alteraciones de laafinidad de la bomba por el Na+ intracelular.
en la
génesis
de laHTAe,
ha sido una constante en la literatura médica. En losúltimos años ha existido un renovadointerés por el aislamiento y caracterización
de inhibidores circulantesde laATPasa
Na+-�,
que ha culminado con la recientepurificación
e identificación estructural de una sustanciaendógena,
presente
enel
plasma
humano anivelessubnanomolares,
que sefija
a losreceptores
de losglicósidos digitálicos
con altogrado
de afinidad y es indiferenciable de laoubaína. Esta "oubaína-like" humana inhibe la ATPasa Na+-K+ y la entrada de
86Rb+
a lacélula,
ydisminuye
la velocidad máxima de la ATPasaNa+-K+,
por loque sería
responsable
de una anomalíaadquirida
de la bomba de Na". Lasmayores concentraciones de esta sustancia han sido detectadas en la
suprarrenales,
lo cualsugiere
que estasglándulas
y no elhipotálamo,
son lafuentedel
compuesto circulante".
En un estudio de las
propiedades
cinéticas de la ATPasa Na+ -K+ en unapoblación
dehipertensos
esenciales,
Díez etar"
consiguieron
identificar a unsubgrupo
depacientes
que, de maneraestable,
presentaban
una disminución dela afinidad
aparente
de la ATPasa Na+-K+ para el Na+ intracelular. En lamayoría
de los
hipertensos
con estedefecto,
definido como anomalía "Na+ Bomba(-)",
lavelocidad máxima de la ATPasa Na+ -K+
puede
hallarse aumentada comomecanismo
compensador
(anomalía
"bomba(+)").
Estos resultados son similaresalos observados por nuestro
grupo48
ysugieren
que, entre el 8% y el 25% de loshipertensos
esencialespresentan
una anomalía intrínseca de la bomba de Na+la necesidad de mayorescifras de Na+ intracelular para
conseguir
el mismo nivelde estimulación que un individuo normal.
1.2.4. Alteraciones
enel
cotransporte
Na+-K+-Cr
Hace ya casi dos
decadas,
Garay
yMeyer"
demostraron un descenso de laactividad del
cotransporte
Na+ -K+ -CI- enhipertensos
esenciales,
hallazgos
quefueron confirmados por los mismos
autores,
tanto al estudiar la actividad encondiciones basales como en eritrocitos sometidos a una
sobrecarga
salina49-so.
Existen marcadas diferencias
raciales,
y parece evidente que en la raza negrase
distribuye
el mayorporcentaje
de individuos con uncotransporte
anormalmente
baj051-52
Posteriormente,
Garay
etal53
constataron la existencia de unsubgrupo
depacientes
con una anomalía delcotransporte,
consistente en una disminución dela finidad
aparente
del sistema para el Na+intracelular,
este defecto establefuedefinido como anomalía "Co
(-)"
y se detecta entre el 18% y el 39% de loshípertensos".
Aligual
que la anomalía "Bomba(-)",
determina una dificultad enla extrusión de
Na",
pues serequieren
mayores concentraciones intracelularesdel catión para
conseguir
un nivel deestimulación similar al del individuo normal.En los
hipertensos
con la anomalía "Co(-)",
la velocidad máxima del sistemapuede
estaraumentada comomecanismocompensador
(anomalía
"Co(+)"),
ser1.2.5.
Alteraciones
enel
contratransporte
Na+-L¡+
En
1980,
Canessa etal55
describieron un aumento de la actividad delcontratransporte
Na+-Li' enpacientes
afectos de HTAe. Lamayoría
deinvestigadores
que han estudiado este sistema detransporte
han corroborado enaumento de la velocidad máximade este sistema en la
población
dehipertensos
esenciales55-60,
así como en individuos normotensos con antecedentes familiaresde
HTA61_
Laprevalencia
de estaanomalía,
definida como "Contra(+)",
ensujetos hipertensos
es muy variable y viene influenciada por la raza,edad,
yexistencia de historiafamiliar
previa
deHTA62-63.
En nuestro medio es una de lasanomalías más frecuentes y afecta entre el 26% y el 49% de los
hipertensos
esenciales64-65.
1.2.6. Alteraciones
enel
intercambiador
Na+-H+
Livne et
al66
describieron en 1987 la existencia de un aumento del actividad delintercambiador Na+-H+ tras
sobrecarga
ácida enplaquetas
dehipertensos
esenciales,
hallazgos
confirmados por otros autores en el mismo modelocelular".
La actividad de este intercambiador se correlaciona con las cifras depresión
arterial diastólica y suhiperactividad
se ha detectado enhipertensos
dediversas
razas",
al contrario de lo que sucede con lahiperactivación
delcontratansporte
Na+ -Li'eritrocitario,
prácticamente
inexistente entre loshipertensos
de razanegra62-63.
modelo celular eritrocitario de
hipertensos
esenciales,
defecto definido comoanomalía "Protón
(+)"
y cuyaprevalencia
oscila entre el 20% y el 80% de loshipertensos estudiados,
según
diferentesinvestiqadores'":".
siendo en nuestromedio del
60%69.
Estospacientes representan
un grupo con característicasclínicas y
biológicas
biendefinidas,
tales como un mayor índice de masaventricular
izquierda,
tendencia apresentar
valores mas elevados deglucemia,
y un
particular
perfillipídico
caracterizado por cifrassuperiores
de colesteroltotal,
LDL-colesterol y
triqlicéridos'".
No se sabe con certeza si lahiperactividad
deeste intercambiador está
genéticamente
determinada,
o es unepifenómeno
enel curso de la enfermedad
hipertensiva,
dado quealgunas
observacionesrecientes
sugieren
esta últimaposibüidad".
1.2.7. Alteraciones
enel intercambiador
HC03-¡CI-Ya en esta
decada,
Diez etal72
observaron que la actividad eritrocitaria delintercambiador
HC03-¡CI-
dependiente
del Na+ estaba anormalmente elevada enun 23% de
hipertensos.
Esta alteración deltransporte
transmembranario desodio,
definida como"Anion(+)",
permanece establea lolargo
de la evolución dela enfermedad. Al analizar las
posibles
características diferenciales de lospacientes
portadores
de esta anomalíarespecto
a loshipertensos
con unaactividad normal de este sistema de
transporte,
estos mismosautores"
observanuna frecuencia
superior
de antecedentes familiares deHTA,
así como nivelessugiere
que estos individuospodrían
representar
unsubgrupo particular
entre loshipertensos
esenciales.1.2.8. Alteraciones
enel
intercambiador
Na+-Mg2+
Muy
recientemente,
Picadoetal74
han observado una aceleración en la velocidadmáxima del intercambiador Na+
_Mg2+
en hematíes de unsubgrupo
depacientes
hipertensos,
querepresentaban
el 45% de todos loshipertensos
esencialesestudiados,
y con unapresión
arterial diastólica más elevada que la de los2.
Metabolismo celular del calcio
2.1. Introducción.
El
Ca2+
penetra
en el citosol celular desde el medio extracelular y desde elretículo
endo(sarco)plásmico
(RS)
a través de canalespropios,
cuyo controldepende
dereceptores
específicos,
demensajeros
intracelulares o de cambiosen el
potencial
demembrana,
dandolugar
a un aumento transitorio de suconcentración
citosólica".
Trasejercer
su acción es extraído de nuevo al medioextracelular y
recaptado
al interior del retículoendo(sarco)plásmico
por sistemasde
transporte
activo.La concentración de
Ca2+
extracelular es deaproximadamente
1mmolllitro,
mientras que en el interior de la célula el
Ca2+
libre citosólico alcanzaconcentraciones 10.000 veces
inferiores,
de rango nanomolar(100
a 200nmol/litro).
Existe por tanto ungradiente
transmembranario extraordinariamenteelevado".
No sólo la membranacitoplasmática
regula
elCa2+
intracelular,
pueslas membranas mitocondriales y las del RS
juegan
unimportante
papel
en lahomeostasis cálcica. En el interior de la matriz mitocondrial se acumula
calcio,
enforma de
compuestos
no iónicoscon fosfato yATp76.
Porsuparte,
el RSjuega
un
importante
papel
en laregulación
delCa2+
citosólico y en la contracción de lasfibras
musculares,
enparticular
en la contracción cíclica de la célulamiocárdica":".
Noobstante,
este retículo sirve como fuente delCa2+
necesarioencargado
deregular
constantemente su concentración intracelular. Esfundamentalmente la membrana
citoplasmática
laresponsable
de la homeostasisdel
Ca2+,
lo queconsigue
merced a unos sistemas detransporte
capaces deextraerlo al medio extracelular y mantener el
gradiente
transmembranoso.2.2. Los canales de calcio
La entrada de calcio extracelulara favor desu
gradiente
electroquimico
se realizaa través de canales
especificas
de lamembrana,
los canales decalcio,
de losque existen dos
tipos:
1/ Los
voltaje dependientes,
que se activan ypermiten
la entrada de calcioen
respuesta
a cambios delpotencial
de membrana.2/ Los
receptor-dependientes,
que se activan cuandounagonista,
comola noradrenalina o la
angiotensina
11,
actúan sobre susreceptores
situados en lamembrana celular.
En las células de la musculatura lisa vascular parecen existir ambos
tipos,
mientras que en las células cardíacas parecen ser exclusivamente
voltaje
dependiente.
De estosúltimos,
voltaje-dependientes,
existen trestipos
denominados
L,
T y N78,
Quepresentan
distintas caracteristicas cinéticas yfarmacológicas.
Tanto en las células cardíacas como en las musculares lisasvasculares no se ha
podido
demostrarla existencia de los canales N que son losque
predominan
en las celulas nerviosas. Los canales L se activan cuando ellentamente
permitiendo
aperturas
delarga
duración,
y la entrada de calcio através de estos canales sería la
responsable
a nivel cardiovascular de: elacoplamiento
excitación-contracción del músculo cardíaco y lisovascular,
de lafase 2 del
potencial
de acción cardíaco y de la velocidad dedespolarización
delos nodos senoauricularyauriculoventricular. Los canales Tse activan a niveles
de
potencial
de membrana de -80 mV y se inactivanrápidamente, permitiendo
aperturas transitorias,
y parecen estar involucrados en lagénesis
de la fase4,
de lenta
despolarización
diastólica de las celulas marcapaso cardíacas yvasculares
79.
2.3. La ATPasa
Ca2+-dependiente.
La bomba de
Ca2+.
2.3.1.
Estructura y funcion de la bomba de calcio
La ATPasa
Ca2+ dependiente
es uno de los siete u ocho mecanismos detransporte
existentes en las membranas celulares que movilizan el calcio entreel interior y el exterior de la célula. Junto con el
contratransporte
Na+-Ca2+
es elprincipal
mecanismo para la extrusión de calcio delcitoplasma
celular y estápresente
en lapráctica
totalidad de las membranas celulares. Engeneral,
laATPasa
predomina
en las celulas no excitables y elcontratransporte
en lasexcítaoles'".
Ello es debido a que estas últimasrequieren
un sistema de altacondiciones que se hallan
presentes
en el intercambiadorNa+-Ca2+.
Por elcontrario,
laATPasaCa2+-dependiente
presenta
una alta afinidad para elCa2+
yuna menor
capacidad
detransporte
y es, portanto,
laresponsable
delmantenimiento constante del
bajo
contenido de calcio existente en el interiorcelular.
La demostración en 1966 de que los eritrocitos eran capaces de extrudir
Ca2+
gracias
a una ATPasaespecifica"
originó
un vasto numero detrabajos,
realizados
principalmente
en este modelocelular,
encaminados aconseguir
lacaracterización de esta enzima. Se trata de una clásica
E1-E2
ATPasa con unaalta afinidad para el
Ca2+
«
1¡.JM),
que se inhibeespecíficamente
porbajas
concentraciones de vanadato
(K1I2
< 1¡.JM).
Gracias a esta alta afinidad estácualificada para extraer
Ca2+
continuamente de lacélula,
por lo quejuega
elpapel
másimportante
en el mantenimiento delgradiente
deCa2+
entre las célulasy el medio extracelular.
La ATPasa
Ca2+-dependiente
es unpolipéptido
con un peso molecular de138.000 daltons en su forma
monomérica,
aunque funciona en la membranacelular como un
dímer082-84.
Surepresentación
en el total deproteinas
membranarias es
baja,
con sólo un0,1 %-0,2%
del total de lasmismas,
que a suvez
constituyen
únicamente alrededor del0,5%
del peso celular.En
presencia
de la enzimareguladora
calmodulina,
laspropiedades
de laATPasa
eritrocitaria,
aquélla
que hapodido
sermejor
estudiada,
sonequivalente
alinflujo
pasivo
deCa2+
que se cifra en unos 50umol/l.cel/h
a unaconcentración internade
Ca2+
inferiora los 30nrnol/l.cels'".
Si tenemos en cuentaque la concentración extracelular de
Ca2+
se halla alrededorde1,3
mmolllpuede
establecerse un
gradiente
en contra de 40.000. De este modopuede
afirmarseque únicamente una
pequeña
parte
de lacapacidad
de la bomba se utilizahabitualmente. Si la concentraciónde calcio intracelularse satura
(1
Oumol/l.cels)
la velocidad de
transporte
puede
alcanzar los 20mmol/l.cels/h,
lo quecorresponde
a varios miles de unidades por minuto. La afinidad para el calcio esalta en el interior
(inferior
a 1�M)
Ybaja
en el exterior de la membrana(10
mM)86.
El sistema hidroliza ATP a la misma velocidad a la que se
transporta
el calcio(1
átomo de calcio
transportado
por molécula de ATPhidroíízado)".
Existen dossitioso locusde
fijación
delATP. Ellocusenzimáticopropiamente
dichopresenta
una afinidad de 2-3
�M,
operando
aisladamente a una décimaparte
de sucapacidad.
Además,
existe otro locusregulador
para el ATP con una afinidadentre 200 y 300
�M
que cuando estáocupado permite
el funcionamiento avelocidad
máxima".
Es
importante
destacar que el sistemarequiere
lapresencia
del ionMg2+
para suoperatividad.
La afinidad para elMg2+
en el interior de la membrana es de 15�M89,
lo que noimplica
que el sistematransporte
este catión. Dehecho,
laespecificidad
de la bomba para el calcio es altamenteselectiva'",
permitiendo
únicamente el
Sr2+
y,posiblemente
elPb2+
como alternativa altransporte
deEstá claramenteestablecido que la bomba de calcio no
contratransporta
ningún
otro catión al interior
celular,
aunque inicialmente se habíapostulado
suintercambio por
Na",
K+ oMg2+,
ni es intercambiado por unningún
anión".
Noobstante,
existen evidenciasexperimentales"
quesugieren
laposibilidad
de unintercambio por
protones,
esdecir,
que el sitio móvil defijación
delCa2+
volveríaal interior celularunido a un H+. Ello viene
apoyado
por el hecho de que cuandose
bloquea
eltransportador
de aniones conDIOS,
aquellos
aniones que pasanla membranaenforma no
disociada,
como elacetato,
estimulan eltransporte
decalcio,
mientras que los que la atraviesan en formaionizada,
como eltiocianato,
no
presentan
estapropiedad.
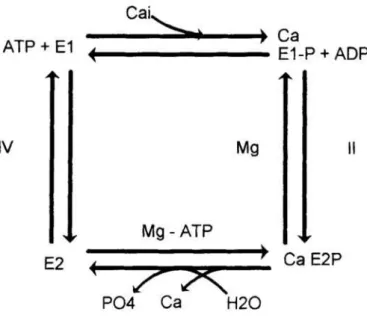
El ciclo de reacción de la bomba de calcio
(figura 1)
es similar al de otrasATPasas comola bomba de sodio.
Básicamente,
la hidrólisis del ATP fosforila laproteína
enpresencia
de calcio.La
proteína
fosforilada cambia su conformación deE1-P
aE2-P,
cambio que seve favorecido porla
presencia
deMg2+.
Durante este cambiode conformación elCa2+
es extruído al exterior paraposteriormente
desfosforilarse laproteína
yATP + E1
Cai�
---�--�----�.
Ca ....--- E1-P+ADP IVMg
IIMg
- ATP E2(
7A:
P04 Ca H20 Ca E2PFigura 1. Ciclo de reacción de la ATPasa
ca=-dependrente.
El ciclo comienza en el extremo superior izquierdoy sigue el sentido horario. E1 yE2 representan las dosconfromaciones de laproteina y E1-P YE2-Plasformas fosforiladas: El Ca2+ atraviesa lamembrana celular durante la
reacción 11. Dicha reacción requiere la presencia de
Mg2+,
mientras que la 111 necesita altas concentracionesde ATP.2.3.2.
La
interaccion
de
la
bomba
de
calcio
conla
calmodulina
Una
importante
característica de la ATPasaCa2+
-dependiente
es sucapacidad
deestimulación por la acción de la
calmodulina94,95,
proteína
soluble intracelularcon cuatro locus de
fijación
para elCa2+
que se hallapresente
en el interior detodas las células y es la
encargada
de mediar en varias de las acciones en lasque el
Ca2+
actúa comosegundo mensajero.
La calmodulina estimula la bombade calcio cuando varios de sus locus de
fijación
para este catión sonocupados.
Esta reacción provoca una interacción calmodulina-bomba de
calcio'"
que resultaPara
algunos
autores"
la ATPasaCa2+
-dependiente
tendría dosconfiguraciones:
unaforma asociada a la calmodulina y otra disociadade esta. La forma disociada
representaría
un estado debaja
afinidad donde la actividad máxima seconsigue
por una concentración de calcio libre
comprendida
entre 100 Y 300¡.JM,
presentando
un solo locus defijación
para el calcio. Por elcontrario,
en su formaasociada a la calmodulina la bomba de calcio se convierte en una enzima de alta
afinidad que alcanzasu máxima actividad para una concentración de calcio libre
citosólico de 10 a50
¡.JM
y donde el número de locus defijación
para el calcio esde tres. El
papel fisiológico
de laregulación
de la bomba de calcio por lacalmodulina no está suficientementeaclarado. Para
algunos
autores, la ATPasaCa2+
-dependiente
en su estado basal estaría disociada de la calmodulina y seríael aumento del calcio intracelular el que
provocaría,
a través de laocupación
delos locus
especifícos
de lacalmodulina,
la asociacion a la ATPasaCa2+dependiente
y elconsiguiente
aumento de su afinidad para elCa2+
y de suvelocidad máxima. Ello obtendría el resultado
deseado,
esdecir,
elrestablecimiento de la concentración basal de calcio y, alcanzado dicho
objetivo,
la calmodulina se disociaría de nuevo de la bomba. Este fenómeno tiene
implicaciones
reguladoras,
pues la calmodulina actuaría como unactivador,
capaz de modificar la conformación de la enzima desde un estado de
baja
afinidad para el
Ca2+
ybaja
velocidad debombeo,
a un estado de característicasdiametralmente
opuestas.
Noobstante,
otros autores ponen en duda estacalmodulina,
con lo que elpapel
regulador
de estaproteína
intracelular seríaprácticamente
nulo.2.3.3.
Regulacion
de la bomba de calcio
Dejando
aparte
laregulación
de la bomba decalcio por lacalmodulina,
que actúacomo un mecanismo de feed-back
negativo
ante los aumentos del calcio librecitosólico,
las características funcionales de la ATPasa de la membranaplasmática dependen
en gran medida del ambientelipídico
en el que se hallainmersa".
Losfosfolípidos,
ya sean neutros ocon carganegativa,
son esencialespara su funcionamiento enzimático en condiciones basales
1()().
Si sereconstruye
la ATPasa con un
fosfolípido
neutro como lafosfatidilcolina,
la adición de otrosfosfolípidos
con carganegativa
como lafosfatidilserina,
elfosfatidilinositolmonofosfato o el fosfatidilinositolbifosfato son capaces de
estimular de forma adicional la actividadde la ATPasa
101.
Si bien no estáprobado
que esta
regulación
tenga
algún papel fisiológico,
sí parece factible que elfosfatidilinositolmonofosfatosea un
regulador
deltransporte
deCa2+
por la bombade la membrana celular.
Se ha descrito que el GMP cíclico
ejerce
un efecto estimulador sobre eltransporte
transmembranario de calcio y que dicho efecto seproduce
medianteuna fosforilación directa de la
enzima'?"
Noobstante,
otrosautores'?'
handemostrado que dicha enzima fosforiladanotiene lascaracterísticas de la bomba