4.3.5 THE GENUS METAXYTHERIUM
Order SIRENIA Illiger, 1811 Family DUGONGIDAE Gray, 1821
Subfamily HALITHERIINAE (Carus, 1868) Abel, 1913 Genus Metaxytherium Christol, 1840
Synonymy – See Domning, 1996.
Type species – Metaxytherium medium (Desmarest, 1822) Hooijer, 1952
Included species – M. arctodites Aranda-Manteca, Domning and Barnes, 1994; M.
crataegense (Simpson, 1932) (= M. calvertense Kellogg, 1966) Aranda-Manteca,
Domning and Barnes, 1994; M. floridanum Hay, 1922; M. krahuletzi Depéret, 1859; M.
petersi Abel, 1904; M. medium (Desmarest, 1822) Hooijer, 1952; M. serresii (Gervais,
1847) Depéret, 1895; M. subapenninum (Bruno, 1839) Fondi and Pacini, 1974.
Range - The genus Metaxytherium showed a wide geographical and stratigraphical distribution during the Miocene: Metaxytherium remains have been collected in several Miocene deposits of the Mediterranean, Paratethyan, northeastern Pacific, southeastern Pacific, northeastern Atlantic, and western Atlantic - Caribbean regions. In contrast, during the Pliocene, Metaxytherium survived only in the Mediterranean area and became extinct in the Middle Pliocene.
Emended diagnosis – Halitheriine dugongids characterizad by a zygomatic-orbital bridge of maxilla elevated about 1 cm above the alveolar margin [c. 11(1)], differ from the Hydrodamalinae in having a supracondylar fossa of exoccipital present [c. 67(1 or 2)]; a dorsolateral border of exoccipital rounded and more or less smooth, not flangelike [c. 70(0)]; lacrimals present [c. 91(1)], an horizontal mandibular ramus broad [c. 128(1)], with ventral border strongly concave [c. 122(3)] and retaining incisor tusks [c. 139(0)]..
Remarks – Metaxytherium dentition: The lifetime dental formula of the genus
Metaxytherium is presumed to be I 1/0, C 0/0, DP 3/3, M 3/3 [c. 139(0); c. 143(1); c.
144(1); c. 150(0); c. 155(2); c. 157(2)].
As observed by Domning (see e.g. Domning, 1988), the cheek teeth of Metaxytherium spp. erupt sequentially throughout the animal’s growth to maturity. The most anterior teeth are worn out and shed as new ones erupt at the rear, and the empty alveoli are eventually obliterated by mesial drift of the posterior teeth. In adult Metaxytherium, the cheek dentition is typically reduced to DP5-M3/dp5-m3 or M1-M3/m1-m3; and of
these, M1/m1 and frequently M2/m2 are so heavily worn that no enamel pattern remains. In old age, even the anterior molars may be lost.
With age the molars advance: the distance between m3 and the rear of the coronoid canal (retromandibular space) increases, and the position of the upper molars relative to the posterior border of the zygomatic-orbital bridge of maxilla changes.
The molars of Metaxytherium show considerable individual variation in size, complexity, and cusp arrangement, and most of this variation is not of value for systematics.
Metaxytherium krahuletzi Depéret, 1895
(Plates 11-14; Figures 17-18)
Synonymy – See Domning & Pervesler, 2001
Type series – KME nos. GII 21, 22, 25, 26, 29, 34, six isolated molars
Type locality – Schindergraben, Eggenburg, Austria. Previously called the Erste Mediterranstufe, Horner Schichten, Liegendsande, or Loibersdorfer Schichten; now termed the Burgschleinitz Formation. Burdigalian in age.
Referred specimens – Main specimens listed in Appendix 2
Range – Early Miocene (early to late Burdigalian), Central Paratethys and possibly Early Miocene (middle Aquitanian to late Burdigalian-early Langhian), Euro-North African western and central coasts.
Emended diagnosis – Metaxytherium characterized by the following primitive features: zygomatic process of squamosal narrow throughout its length [c. 83(0)] and sternum tripartite; and in which the nasals are sometimes united in midline [c. 31(0 or 1)], the exoccipitals sometimes meet in midline [c. 66(0 or 1)], the ventral extremity of jugal is sometimes under the posterior edge of orbit [c. 85(1 or 2)].
Remarks - M. krahuletzi has been recently revised by Domning and Pervesler (2001). They recorded the species with confidence only from the Burdigalian Central Paratethys deposits of Austria and Switzerland and maybe also from Horné-Strháre (Slovakia) and Léognan (France). Moreover, they cited the skeleton, now lost, from the Burdigalian of Beaucaire (France) named Metaxytherium beaumontii by Christol in Blainville (1844) and supposed to represent M. krahuletzi by Depéret and Roman (1920: 31); some specimens from Klattau (Bohemia, Czech Republic) referred by Pia and Sickenberg (1934) to M. krahuletzi, but which Domning and Pervesler did not have the opportunity to examine; and two skull fragments from the Burdigalian of Gebel Zelten (Libya) described as Metaxytherium sp. indet. by Heal (1973), which might be referable to M.
krahuletzi, but need further study (see Appendix 2).
In this study I also assign to Metaxytherium cf. krahuletzi a new French specimen (MPNRL-MAN2000) represented by a skull with associated thoracic vertebrae and ribs, from the late Burdigalian deposits of Manosque (Provence, southern France), the humerus from the upper Burdigalian of Quinta da Farinheira (Lisbon, Portugal) reported by Zbyszewski (1944), Metaxytherium aquitaniae from the Aquitanian deposits of northwestern France, and Metaxytherium catalaunicum from the upper Burdigalian – Langhian deposits of southeastern Spain.
MPNRL-MAN2000: description (Plates 11-12; Figures 17-18) – The specimen is represented by a skull with associated thoracic vertebrae and ribs. The cranium and the mandible are in anatomic connection, so the ventral side of the cranium and the dorsal side of the mandible are not visible. They are deformed, mediolaterally flattened with the right side still partially embedded in the molasse, and the posterior portion incomplete (lacking the right exoccipital and the basioccipital).
The rib cage is also embedded in the hard sandstone matrix; the left ribs are more or less in anatomic position, the right ones are posteriorly inclined, the vertebrae are crushed, lying on the right side, but more or less in anatomic connection.
The exoccipital-supraoccipital and the interpremaxillary sutures are completely fused, and the m3 is completely erupted, which indicate that the specimen was an adult.
The state of preservation and the incomplete preparation do not permit scoring of all the morphological characters. However, the features visible include those most critical for taxonomic determination in this case; the difficulty in making a specific identification does not result from incomplete preparation.
SKULL
Premaxilla – The rostrum is incomplete and compressed, but long enough to be supposed enlarged relative to the cranium [c. 3(1)]. Only the dorsal portion is preserved, but the dorsal keel is damaged so that the dorsal surface of the premaxillary symphysis appears to be concave. The posterior end of the rostrum seems to be upraised to form a slight boss in lateral view [c. 10(1?)]. The lateral edges are thin, not flaring. The tusks and the alveoli are not preserved, but, judging from the development of the rostrum, they were probably small, extending less than half the length of the symphysis [c. 140(0)].
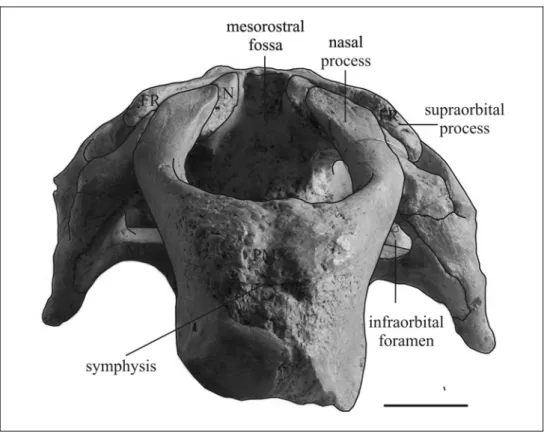
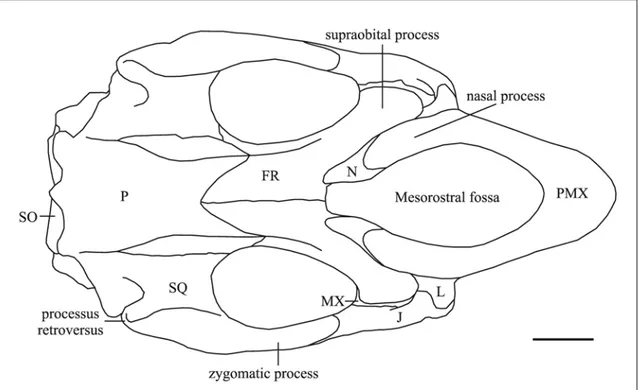
The nasal process is long [c. 7(0)] and thin at its posterior end [c. 6(0)]. It contacts the frontal [c. 9(1)], the lacrimal [c. 1(1)], and abuts against the nasal, but the sutures with these bones are unclear. The premaxillary-maxillary suture is partially visible and its anterior end is estimated to be about 5 mm posterior to the dorsal tip of the symphysis. The contact with frontal and lacrimal appears to be about 86 mm long.
The mesorostral fossa is long and narrow and the external nares appear to be retracted and enlarged beyond the level of the anterior margin of the orbit [c. 8(1?)]. A nasal incisure at posterior end of mesorostral fossa seems to be absent [c. 37(0?)], but these
characters could be altered by compression and deformation. The rostral deflection is uncertain; probably circa 60°.
Nasal – The nasal region is abraded and incomplete, so the dorsal exposure of the nasal cannot be clearly delineated; but processes of the frontals seem to separate the nasals in the midline [c. 31(1)].
Lacrimal – The lacrimal is about 5 cm long and has a prominent ridge, 3 cm long, on its lower lateral surface. It abuts posterodorsally against the supraorbital process of the frontal and anterodorsally against the nasal process of premaxilla, and it is anteromedially in contact with the jugal. The contact with the maxilla is unclear. A nasolacrimal canal is absent [c. 91(1)].
Frontal – The supraorbital process is well developed, about 2 cm thick dorsoventrally with a long, acute anterior point and a small posterolateral corner [c. 36(1)], flattened in the horizontal plane, with a dorsal surface inclined relatively gently ventrolaterad [c. 43(0)]; not divided by deep dorsoventral grooves indenting its lateral margin. A large orbicular apophysis is present on each side. The lateral crests are not significantly overhanging. The medial wall of the temporal fossa is formed by a thin lamina orbitalis of the frontal [c. 38(0)] and a crista orbitotemporalis is present. The median portion of the frontal roof is convex [c. 42(0)] with a low longitudinal crest. The anterior portion is not well preserved and the contacts with nasal and premaxilla are not visible. The interfrontal suture is fused.
Parietal – The parietal is 74 mm long in the midline and 119 mm long laterally. The interparietal suture is fused. The fronto-parietal suture is concave in shape. The posterolateral corners of the roof are indented by the squamosals. The roof is nearly rectangular, not constricted [c. 46(2)], posteriorly flat and anteriorly slightly concave [c. 45(1)]. The temporal crests are lyriform, low, broad, confined to the lateral edges; minimum external distance between crests 74 mm, minimum internal distance 37 mm (Type B from Domning, 1988). A sagittal crest is absent [c. 51(1)]. A median bump is present just in front of the external occipital protuberance.
Supraoccipital – The right ventral portion of the supraoccipital is lacking. The supraoccipital is hexagonal in outline with rounded dorsolateral corners; it forms an
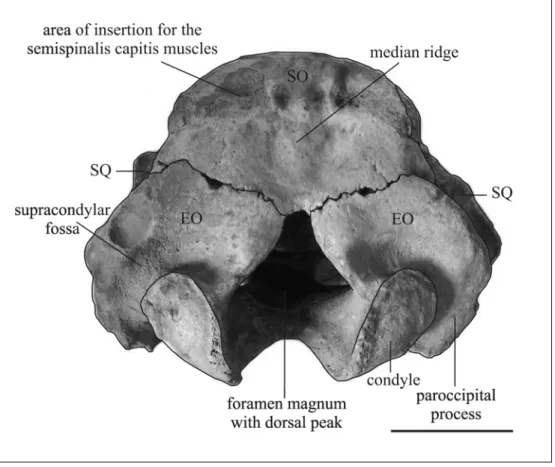
angle of 125° with the after part of the parietals. The parietal-supraoccipital suture is fused. The external occipital protuberance is very large and thick anteroposteriorly; it rises above the plane of the parietal roof, and the median ridge below it is present but short; the nuchal crest is distinct at its lateral ends near the squamosals, and the lateral border is thick, rounded and not overhanging. The areas of insertion for semispinalis capitis muscles are flattish, face posterodorsad, and extend about halfway to the ventral end of the bone. The lower part of the supraoccipital is convex laterally below the semispinalis insertions. The sutural surfaces for the exoccipitals are not distinct. The ratio of width to height is about 1.21.
Exoccipitals – Only the dorsal portion on the left side is preserved. The bone appears to be fused with the supraoccipital. The dorsolateral border is rounded, smooth and not flangelike [c. 70(0)]. The supracondylar fossa is deep and extends across the entire width of the occipital condyle [c. 67(1)].
Maxilla – Only a small dorsal portion is visible laterally. The ventral side and the zygomatic-orbital bridge are concealed by matrix. The infraorbital foramen is not visible in anterior view because of the compression.
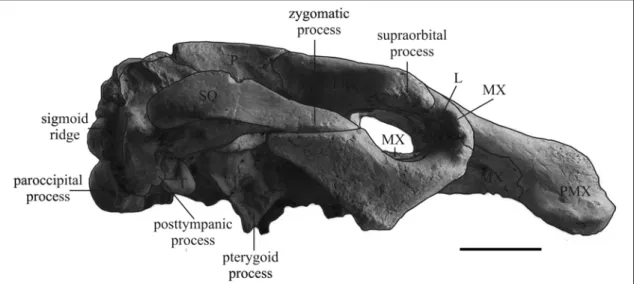
Squamosal – The squamosal is dorsally in contact with the squared posterior part of the parietal roof [c. 76(1)]. The sigmoid ridge is prominent, dorsally slightly sharp and ventrally more rounded. The surface of the cranial portion dorsal to the zygomatic root is nearly flat. The postglenoid process and the postarticular fossa are distinct and well developed. The processus retroversus is present, moderately inflected [c. 77(1)], and does not project below the line of the suture with the jugal; its posterior end is incomplete, but distinct dorsal and ventral protuberances are present. The temporal condyle is not visible. The zygomatic process is 160 mm long, not much broader posteriorly than anteriorly [c. 83(0)]. The posterodorsal edge of the process is almost straight (slightly concave) in outline and convex laterad. The external auditory meatus is partly obscured, but seems to be about as wide anteroposteriorly as high [c. 82(1?)]. Jugal – The preorbital process is relatively flat and thin [c. 88(1)], but with a protuberance on the posterior side. It presumably does not contact the premaxilla [c 87(0?)]. The ventralmost point of the jugal lies clearly under the orbit itself [c. 85(2)], as mentioned by Domning and Pervesler (2001: 43). The ventral tip is thick and rounded;
the border behind is smooth. The ventral margin of the orbit is concave, smooth, broad but not overhanging [c. 90(0)]. There is a raised postorbital process against which the squamosal abuts [c. 86(1)]. The posterior process is incomplete, but seems to be as long as the diametre of the orbit [c. 89(0?)], tapering rapidly in outline with a longitudinal shallow sulcus on the lateral side.
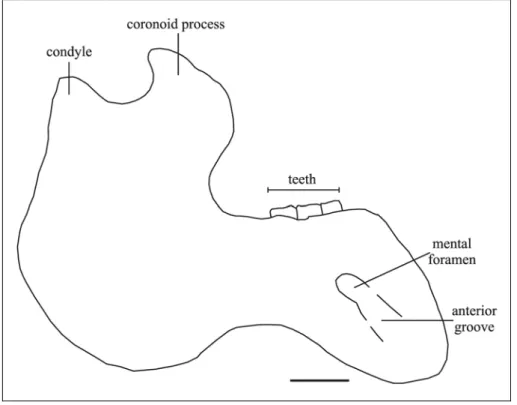
Mandible – Left and right dentaries are preserved, but they are mediolaterally flattened. The left is exposed, but the right is enclosed in the matrix. The mandible is in anatomic position and the dorsal extremity of the ascending ramus is therefore covered by the zygomatic arch. The condyle, partially exposed, is elliptic and laterally overhanging. The mandibular notch and coronoid process are not exposed. The base of the anterior border of the coronoid process is exposed and extends slightly anterior to the base of the process [c. 126(1)]. The posterior border is thick and rounded. It has no distinct processus angularis superior, but has a broadly convex outline beginning well below the condyle [c. 125(2)]. Lateral to m3 there is a shelf that slopes steeply. The horizontal ramus is deep and short [c. 128(1)], with a strongly concave ventral border [c. 122(3)] that is not tangent to the mandibular angle [c. 129(1)]. The ventral border is thickened and rounded anteriorly, tapering posteriorly. Only one large mental foramen is present [c. 123(1)], and it lies at the level of deflection of the dorsal edge of the ramus. The deflection of the masticating surface is abrupt, about 64° (60° according to Domning & Pervesler, 2001: 44). The masticating surface is incomplete and mediolaterally compressed. The lateral edge is thin and overhanging. The posteroventral side of the symphysis is concave and the anteroventral side is convex transversely. The symphyseal suture is not well preserved, but appears to be fused. The labial side of m3 is visible. The distance between m3 and the coronoid process (retromandibular space) is about 25 mm.
Fig. 17: Skull of MPNRL-MAN2000 from Manosque (Provence, France) in a) left lateral; b) posterior; and c) dorsal views. Scale bar: 10 cm (modified from Sorbi, in press).
POSTCRANIAL BONES
Vertebrae – The cervical and the most anterior thoracic vertebrae are missing. Twelve thoracic vertebrae and a very small fragment representing a thirteenth thoracic vertebra are preserved in anatomic sequence (Ta-m). They are crushed, fragmented and lying on the right side. The centra seem to be crushed and only the left sides of the neural spines and the left transverse processes are exposed. The neural spines are better preserved than the transverse processes. The first vertebrae are partially overlapping. The
overlapping diminishes from the first to seventh vertebra and the following vertebrae are in anatomic position.
The neural spines are laterally flattened with a slight backward slope. Their full height is around 100 mm. The posterior side of each spine has a rounded protuberance; posteriorly this protuberance gradually becomes confined to the lower part of each spine. The posterolateral edge of each spine is notched just above the postzygapophysis; the notch seems to become less developed from the eighth vertebra. The summit of each spine is expanded (transverse width about 20-25 mm; anteroposterior length around 60 mm), flattened, with anterior and posterior corners rounded. The transverse processes are massive and thick. Their mediolateral length is about 25 mm, anteroposterior length about 42 mm, and dorsoventral thickness about 10 mm. They are deformed, but seem to be horizontal and anterior to their neural spines. The zygapophyseal articular planes are nearly horizontal on anterior vertebrae and seem to slope inward beginning with the seventh vertebra.
Ribs – Ten ribs in anatomic sequence (Ra-j) and a small fragment (R?) are preserved on the left side; seven ribs posteriorly inclined are preserved on the right side (Re-g Ri-l). Their contacts with the vertebrae are not visible. The ribs are massive and thick, smooth with a low ridge (iliocostalis tendon insertion) on the posterior edge; this ridge becomes longer on the posterior ribs. The shafts are elliptical in cross-section and maintain their diametres for most of their length. The distal ends are flattened, swept back, and taper in the last 10 – 20 cm.
Fig. 18: Ribs and thoracic vetrebrae of MPNRL-MAN2000 from Manosque (Provence, France) in dorsal view. Scale bar: 10 cm (modified from Sorbi, in press).
MPNRL-MAN2000: comparison and discussion – This specimen differs from the Trichechidae in having a large estimated rostral deflection (about 60° for the premaxilla and 64° for the mandible); an enlarged rostrum [c. 3(1)]; a squamosal with a processus retroversus [c. 77(1)]; a supposed deeply rounded external auditory meatus [c. 82(1?)], and a mandible with a deep and short horizontal ramus [c. 128(1)] and a strongly concave ventral border [c. 122(3)]. Moreover it differs from Anomotherium
langewieschei and Miosiren kocki in having a thin lamina orbitalis of the frontal [c.
38(0)] and processes of the frontals which seem to separate the nasals in the midline [c. 31(1?)]; and it particularly differs from Miosiren kocki in having supposed small tusk alveoli [c. 140(0?)].
MPNRL-MAN2000 differs from the Dugonginae in having the supraorbital process of the frontal flattened in the horizontal plane, with a dorsal surface inclined relatively gently ventrolaterad [c. 43(0)], the preorbital process of the jugal relatively flat and thin [c. 88(1)], and the ventral margin of the orbit not overhanging [c. 90(0)].
Moreover, it differs from “Halitherium” bellunense and Rytiodus spp. in having the nasal process of the premaxilla thin at its posterior end [c. 6(0)] and a long nasal process
of the premaxilla [c. 7(0)]. In particular it differs from Rytiodus spp. in having supposed small I alveoli [c. 140(0?)], and a convex median portion of the frontal roof [c. 42(0)]; and from “Halitherium” bellunense in having a squamosal extending to the temporal crest [c. 76(1)].
All the morphological characters identified for MPNRL-MAN2000 are consistent with the Halitheriinae species.
MPNRL-MAN2000 differs from Halitherium christolii in having the supraorbital process of the frontal dorsoventrally thickened with a posterolateral corner [c. 36(1)]. This character is scored as derived in all the Old World Neogene Halitheriinae (Domning, 1994: table 1); therefore MPNRL-MAN2000 appears to be closer to the Old World Neogene Halitheriinae. Among them M. aquitaniae is represented only by a few fragmentary specimens which could be referred to M. krahuletzi (see below); M. petersi is considered a species evolved in isolation in the Carpathian Basin (Domning, pers. comm.); M. catalaunicum is considered a synonym of M. medium by Domning (1996) but it could be referred also to M. krahuletzi (see below). Therefore the only Euro-North African Early to Middle Miocene halitheriine species recognized and widespread are M.
krahuletzi and M. medium.
MPNRL-MAN2000 appears to be primitive among the Metaxytherium species in retaining the following characters: nasal incisure at posterior end of mesorostral fossa absent [c. 37(0)] and zygomatic process of squamosal narrow throughout its length [c. 83(0)].
C. 37. This character is scored as primitive in M. krahuletzi and in M. catalaunicum, while in M. medium and M. floridanum a nasal incisure at posterior end of mesorostral fossa is usually present [c. 37(1)] and it separates the nasals.
C. 83. This character is usually scored as primitive in M. krahuletzi: “zygomatic process usually not much broader posteriorly than anteriorly; process frequently sinuous and rather narrow throughout its length” (Domning & Pervesler, 2001: 22). It is primitive also in MV 1210; while it is polymorphic in M. floridanum, and derived in M. medium,
M. serresii, and M. subapenninum.
On the other hand MPNRL-MAN2000 has a dorsoventrally broad horizontal ramus of the mandible, with a ratio of minimum height of horizontal ramus (MO) to total length of mandible (AB) of 0.27, and with a mandibular deflection of about 64°.
The MO/AB ratio appears to be higher than the typical ratio in M. krahuletzi mandibles graphed by Domning and Pervesler (2001: fig. 10). The mandibular deflection also appears to be greater than those observed in M. krahuletzi (53° - 63°, from Domning &
Pervesler, 2001). In M. catalaunicum it is about 66°, in the M. medium specimen MPUN 18403 is about 65° and in the M. medium specimen MNHN 1921-10 it seems to be 75°, but the anterior end is damaged and this renders the datum questionable.
In conclusion, I refer MPNRL-MAN2000 to M. cf. krahuletzi. MPNRL-MAN2000 could represent a late form of M. krahuletzi with a slightly greater mandibular deflection and a slightly greater ratio of minimum height of horizontal ramus (MO) to total length of mandible (AB).
Metaxytherium aquitaniae: description (Plate 13) – The species was founded by Pilleri (1987) on the basis of a parieto-supraoccipital skullcap and a cast of dorsal endocranium (NHMB 09.917a), collected from the Aquitanian deposits of Courtebotte near Gans (Gironde, SW France). Pilleri (1987) referred to this species also a fragment of left scapula (NHMB 09.918) and five fairly small skull fragments (NHMB 09.917b) from the same locality.
The original label indicated “Rhytiodus capgrandi”, but Pilleri (1987) considered it incorrect, compared the specimens with Metaxytherium krahuletzi and, on the basis of differences and affinities, considered these specimens as belonging to a new species of
Metaxytherium. Domning (1996) maintained the species as possibly valid, but
considered its status and affinities uncertain.
The following description supplements Pilleri’s description (1987).
NHMB 09.917a is a skullcap constituted by parietals and supraoccipital fused to each other. The frontoparietal, the parietal-squamosal and the supraoccipital-exoccipital sutures are completely open; while the interparietal and the parietal-supraoccipital sutures are perfectly fused.
The parietals bear massive, rounded, lyriform, low temporal crests with posteriorly rounded lateral edges. The crests show their maximum development in the centre with a width of about 21 mm; they diverge posteriorly and become progressively lower. The apexes of the crests are separated by a minimum of 43 mm across a median valley about 20 mm wide (TYPE C from Domning, 1988). An anteromedially directed groove separates the posteromedial side of each crest from the nuchal crest. The nuchal crest is
distinct from the external occipital protuberance. The cranial vault is trapezoidal in anterior view and 24 mm thick in anterior midline. The parietal roof is flat [c. 45(1)]. A sagittal crest is absent [c. 51(1)]. The parietals are 70 mm long in the median line, 83 mm wide at the level of posterior corner of the temporal crests, posteriorly constricted to 68 mm at the level of indentations by squamosals, and anteriorly narrower at 57 mm.
The suture with the frontal is concave in dorsal view. The cranial portion of the squamosal is absent, but the lateral sutures of the parietals indicate that the squamosal reaches the level of the roof and slightly indents the parietal [c. 76(1)].
The internal occipital protuberance is distinct and rounded; the bony falx cerebri is sharp and long [c. 47(1)], anteriorly reaches the frontoparietal suture and posteriorly contacts the internal occipital protuberance with a rounded vertex.
The supraoccipital is hexagonal in outline, 74 mm wide, 50 mm high, 11 mm thick; with rounded dorsolateral corner, not prominent in outline. It forms with parietal an angle of about 137°.
The external occipital protuberance is very low, but rises slightly above plane of parietal roof. The surface below it is medially flat, without a median crest, and laterally concave and rugose at the level of the areas of insertion for semispinalis capitis muscles. These areas are wider than high, extending no more than halfway to ventral end of supraoccipital, and are ventrally delimited by two transverse ridges. The supraoccipital-exoccipital sutures meet at an angle of about 144° and are separated in midline by a small pit [c. 66(1)].
NHMB 09.918 is a fragment of left scapula 180 mm long with glenoid fossa. I refer to Pilleri (1987: 43-45) for its description and measurements.
Metaxytherium aquitaniae: comparison and discussion – The shape and the size of the specimens are suggestive of those of Metaxytherium krahuletzi as observed also by Pilleri (1987); and the characters pointed out by him in NHMB 09.917a (temporal crests very rounded and thick; arched parietal and deep supraoccipital) do not appear to be sufficient to assign the specimen to a new species, since parietals and supraoccipitals are extremely variable in the Metaxytherium species, especially in temporal crest morphology (e.g. Domning, 1988). This is also true in M. krahuletzi, in which three types of temporal crests (A, B and C) are described (Domning & Pervesler, 2001).
M. krahuletzi was recorded in the Burdigalian deposits of Central Paratethys and
probably other parts of the Euro-North African coasts, including the Aquitaine Basin (Domning & Pervesler, 2001).
In conclusion, the specimens here described could represent an older M. krahuletzi record, extending the chronostratigraphic range of the species to the Aquitanian.
Metaxytherium catalaunicum: description (Plate 14; see also Pilleri et al., 1989) - The species was founded by Pilleri (Pilleri et al., 1989) on the basis of a cranium and mandible, two molars, 3 cervical vertebrae, and a rib fragment (MV 1210) from the upper Burdigalian – Langhian deposits of Olèrdola (Mas Romeu, NE Spain). The year before, Pilleri (1988b) described the mandible of the specimen, which has an extensive pathology of the right mandibular ascending ramus. In 1996, Domning considered
Metaxytherium catalaunicum as a synonym of M. medium.
The skull is well preserved and nearly complete, but the mesorostral fossa, the temporal region and the cranial cavity are filled by a consolidated matrix. The pathological right side of the mandible is nearly complete, while just the left horizontal ramus is preserved. The space between the rami is filled by matrix.
SKULL
Premaxilla – The premaxilla lacks the anterior portion. The dorsal keel of the rostrum is thin and sharp anteriorly, and broadens posteriorly into a rugose and slightly concave surface. The posterior end of the rostrum is upraised to form a boss in lateral view [c. 10(1)]. The lateral edges are thin, not much flaring; the preserved anterior portion of the rostrum is narrow, therefore the tusk alveoli, if present, are inferred to be short, extending less than half the length of the symphysis [c. 140(0)]. The symphysis is fused. The palatal surface is rugose, not very concave. The masticating surface of the rostrum is trapezoidal in outline. The nasopalatine canal is filled by matrix.
The nasal process is long [c. 7(0)] and thin [c. 6(0)], contacts frontal [c. 9(1)], lacrimal [c. 1(1)], but not jugal [c. 87(0)]; it abuts against nasal; and ventrally it lies in a groove in the maxilla. The mesorostral fossa is long, without a posterior nasal incisure [c. 37(0)]; it reaches the level of the posterior margin of the orbit [c. 8(2)]. Its anterior end is rounded and constricted by a bulge on medial side of premaxilla. The maximum width of the mesorostral fossa lies anterior to the orbits. The rostral deflection is about 60°.
Nasal – The nasals are relatively large, thick, and massive, set in sockets in anterior margins of frontals. They are posteriorly separated by small triangular internasal processes of frontals, but anteriorly united in midline [c. 31(0)]. The dorsal exposure of nasals is about 25 mm long, and irregular in shape.
Lacrimal – The lacrimal is irregular in shape, mediolaterally flattened, without a nasolacrimal canal [c. 91(1)]. It abuts posteriorly against supraorbital process of frontal, contacts dorsally the premaxilla, medially the maxilla and anteroventrally it fits in a socket on dorsal surface of jugal. The jugal seems to not overlap its lateral side anteriorly.
Frontal – The supraorbital process is well developed, about 2 cm or more thick dorsoventrally with a small posterolateral corner [c. 36(1)], flattened in a horizontal plane, with a dorsal surface inclined more or less ventrolaterad [c. 43(0)]; not divided on its lateral margin. An orbicular apophysis is absent. The lateral crests are in continuity with the temporal crests of parietal, and not overhanging. The temporal region is obscured by matrix, so the development of the lamina orbitalis of frontal is not observable. The internasal processes are small, extending just 15 mm between the nasals. The medial portion of the frontal roof is slightly convex [c. 42(0)] and fragmented, and the interfrontal suture appears to be open. The fronto-parietal suture is not discernible.
Parietal – The posterolateral corners of the roof are indented by the squamosals [c. 76(1)]. The parietals are fused to the supraoccipital and to each other. A sagittal crest is absent [c. 51(1)]. The temporal crests are lyriform, low, not much overhanging, smoothly rounded, with dorsal surface rugose and separated by about 2 cm, with a concave roof (TYPE C from Domning, 1988). The roof is rectangular, without an intertemporal constriction [c. 46(2)], and slightly concave [c. 45(1)]. An emissary foramen is absent. A median bump just in front of the external occipital protuberance is present. The ventral side is not visible.
Supraoccipital – The supraoccipital is subrectangular in outline, 61 mm high and 83 mm wide (ratio 1.36), with rounded dorsolateral corners. It forms with the after part of the parietals an angle of 95°. The external occipital protuberance rises slightly above the plane of parietal roof; it is larger in the centre, and below it a low and short median ridge is present. The nuchal crest is distinct at its lateral ends near squamosals. The areas of insertion for the semispinalis capitis muscles are flattish, smooth, extending more than halfway to ventral end of supraoccipital. The lateral borders of supraoccipital
are rounded, not very thick. The suture with the exooccipitals is M-shaped with a wide central angle of 135°, without a median notch.
Exoccipital – The exoccipitals are fused to each other in midline, and meet in a suture 20 mm long dorsal to foramen magnum [c. 66(0)]. The foramen magnum is triangular in shape, but without an acute dorsal peak. The dorsolateral border is about 2-2.5 cm thick, and has a smoothly rounded posterior edge, posteriorly slightly overhanging but not flangelike, forming a rugose surface at the level of the top of supracondylar fossa. The fossa is deep, extending across entire width of occipital condyle [c. 67(1)]. The left condyle is complete, and reaches below level of tip of paroccipital process. The paroccipital process is short [c. 65(1)], with rugose posterior surface and a posteroventrally curved flange.
Basioccipital – It bears a pair of convex rugosities for the longus capitis muscles. The posterior slope is steeper than the anterior.
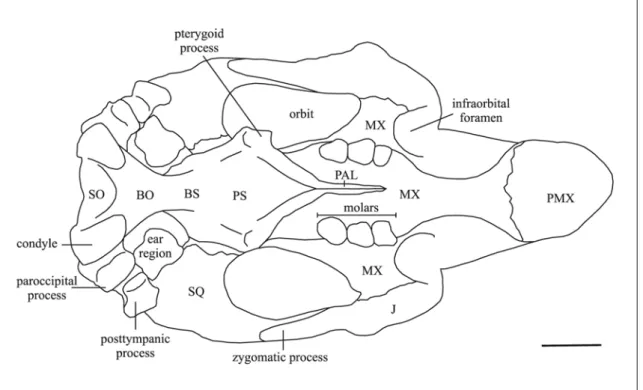
Sphenoid and Pterygoid Regions - Just the ventral surface is visible. The basisphenoid appears to be fused to the basioccipital, indicating maturity of the specimen. The pterygoid process is long and laterally inclined; its ventral tip is enlarged, convex and rugose; its posterior side is dorsoventrally concave.
Palatine – The palatine forms with the alisphenoid the anterior extremity of the pterygoid process. The anterior ends of the palatines are not clearly distinct. The median sutural contact between palatines extends forward from the level of M3 to probably just beyond the posterior edge of the zygomatic-orbital bridge [c. 99(0?)]. The posterior border is incised [c. 97(1)].
Maxilla - The alveolar portion of maxilla is heavy and massive; its dorsal side bears a vertical plate that helps form wall of temporal fossa, but its development is not visible because of the matrix. The edges of palatal surface are lyriform: the surface narrows forward of molars, becomes narrowest forward of anterior edge of zygomatic-orbital bridge, then widens at posterior end of rostral masticating surface. The palatal gutter is narrow and not very deep, with rounded edges and a low median crest. The palatal and rostral surfaces meet in a smooth but abrupt curve. The zygomatic-orbital bridge is large and anteroposteriorly long, and elevated about 9 mm above alveolar margin [c. 11(1)].
The bridge slopes slightly posteroventrad; the anterior edge is thinner than the posterior. The posterior opening of maxillo-premaxillary canal is obscured by matrix. The infraorbital foramen is large, more or less round [c. 13(1)]; it is filled by matrix, but it seems not to be obstructed [c. 20(0?)].
Squamosal – The squamosal indents the posterolateral corners of the parietal roof [c. 76(1)] and extends to the temporal crests. The posterior edge along mastoid foramen is rounded. The prominent sigmoid ridge extends from dorsal end of mastoid foramen to ventral end of posttympanic process, forming a strong laterad-projecting flange with its posteroventral edge broad and rugose. The postglenoid process and postarticular fossa are distinct and well developed. The temporal condyle is irregularly oval. The processus retroversus is moderately inflected, not projected below line of suture with jugal [c. 77(1)]; its posterior end is smooth, with dorsal and ventral protuberances. The posttympanic process is present [c. 73(1)]. The external auditory meatus is nearly rounded [c. 82(1)]. The zygomatic process is not much broader posteriorly than anteriorly, but narrow throughout its length [c. 83(0)]; its posterodorsal edge is convex laterad; its anterior tip is long, reaching the level of the posterior extremity of supraorbital process.
Jugal – The ventralmost point lies under the posterior edge of the orbit, forward of the anterior tip of the zygomatic process of the squamosal [c. 85(1)]. The ventral tip is thickened, roughened, slightly turned medially. The preorbital process is relatively flat and thin [c. 88(1)]; it does not contact the premaxilla [c. 87(0)]. The ventral margin of orbit is broad, concave but not significantly overhanging [c. 90(0)]. The zygomatic process is nearly horizontal, tapered in outline, longer than diametre of orbit [c. 89(0)]. A raised postorbital process in front of tip of squamosal seems to be absent [c. 86(0)]. Auditory Apparatus – Poorly preserved.
Mandible – The right side of the mandible is nearly complete, marked by an extensive abscess on the ascending ramus. This forms a sort of a crater 93 x 115 mm in diametres and about 40 mm deep, open caudally, with thick edges (see Pilleri, 1988b). On the left side only the horizontal ramus is preserved.
The right condyle is incomplete; it appears to be dorsally subtriangular and overhanging. The mandibular notch is incomplete and the coronoid process is broken,
but the ventral portion of its anterior border is preserved and extends far anterior to the base [c. 126(1)].
The posterior border of the right mandible is altered by the extensive pathological crater, but the processus angularis superior seems to be absent and the posterior border seems to have a broadly convex outline beginning well below the condyle [c. 125(2?)]. The horizontal ramus is dorsoventrally broad [c. 128(1)], with ventral border strongly concave [c. 122(3)], not tangent to angle [c. 129(1)].
The mental foramen is better seen on the left side. It lies at the level of deflection of dorsal edge of ramus; it is large with an anterior groove; small accessory foramina seem to be absent [c. 123(1)]. The deflection of masticating surface is about 66°. The mandibular foramen is covered by matrix. The mandibular dental capsule is exposed posteroventrally. The masticating surface is heart-shaped, notched posteriorly and narrowed anteriorly, and rugose with vestigial alveoli identifiable on both sides; the lateral edges are convex and overhanging. The symphysis is broad. The symphyseal suture appears to be incompletely fused.
DENTITION: The tusks are not preserved, but the narrow anterior portion of the premaxilla suggests short and small alveoli.
The lifetime dental formula is presumed to be I 1/0, C 0/0, DP 3/3, M 3/3, as in other
Metaxytherium [c. 139(0); c. 143(1); c. 144(1); c. 150(0); c. 155(2); c. 157(2)].
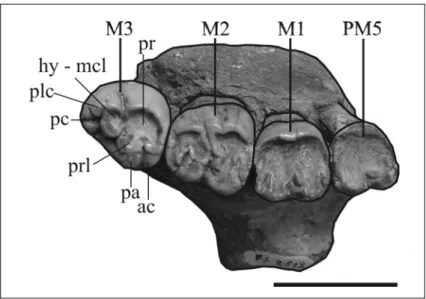
The molars are large and well developed. The molar enamel is smooth, about 1 mm thick. The molars, besides their natural wear, are abraded and damaged
UPPER DENTITION
DP3 – Not preserved; alveolus filled with spongy bone. DP4 – Not preserved; alveolus filled with spongy bone.
DP5 – Only fragments of the crown are preserved on the left side, and the two labial roots are preserved on the right side.
M1 – Heavily worn on the left side, with only the presence of two transverse rows of cusps discernible; crown broken on the right side.
M2 – Moderately worn; only the presence of two transverse lophs is discernible. It differs from M1 primarily in its larger size.
M3 – Moderately worn; larger than M2. LOWER DENTITION
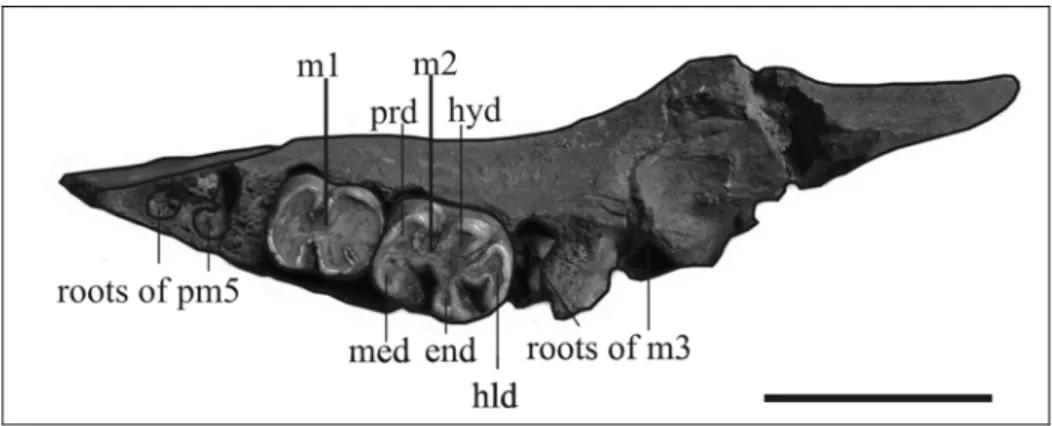
dp3 - Not preserved. dp4 – Not preserved.
dp5 – Not preserved, but its alveolus, filled with spongy bone, is present on the right side.
m1 – Not preserved, but its alveolus, filled with spongy bone, is present on the right side.
m2 – Preserved on the right side, heavily worn. It is a rectangular tooth, with two transverse lophids and a hypoconulid.
m3 – Broken on the right side; preserved but incomplete on the left side. It is a rectangular tooth, longer than the m2; the hypoconulid is longer, making the crown more elongate. It is slightly worn, anterolabially broken, so that just the metaconid and the entoconid with the median ridge are preserved on the lingual side. The hypoconulid consists of three cuspules plus an anterior median spur. The small posterior basin is blocked by the contact of the anterior median spur and the sharp median ridge extending back from the centre of the hypolophid. This median ridge does not bend labiad, so it does not form a Y-shaped hypoconulid.
POSTCRANIAL BONES: The atlas and axis are present in the consolidated sediment; an incomplete third cervical vertebra and a rib fragment are also preserved. They seem not differ from those of other Metaxytherium:
Metaxytherium catalaunicum: comparison and discussion - MV 1210 belongs to the genus Metaxytherium in having a zygomatic-orbital bridge of maxilla elevated above palate [c. 11(1)], a supraorbital process of frontal dorsoventrally thickened [c. 36(1)], a supracondylar fossa of exoccipital present and variably developed [c 67(1 or 2)], a dorsolateral border of exoccipital not flangelike [c. 70(0)], a lacrimal without nasolacrimal canal, but still large [c. 91(1)], a mandible with ventral border strongly concave [c. 122(3)] and horizontal ramus broad dorsoventrally [c. 128(1)].
MV 1210 appears to be primitive among the Metaxytherium species in retaining the following characters: nasals united in midline [c. 31(0)], nasal incisure at posterior end of mesorostral fossa absent [c. 37(0)], exoccipitals united in midline [c. 66(0)], zygomatic process of squamosal narrow throughout its length [c. 83(0)], ventralmost point of jugal under the posterior edge of the orbit [c. 85(1)].
C. 31. This character is polymorphic in M. krahuletzi and in M. serresii, while M.
medium and M. floridanum usually show the derived state. Also in MPNRL-MAN2000
C. 37. This character is scored as primitive in M. krahuletzi, in MPNRL-MAN2000 and in M. subapenninum, while it is polymorphic in M. serresii. On the contrary in M.
medium, M. crataegense, M. arctodites and M. floridanum a nasal incisure at posterior
end of mesorostral fossa is usually present [c. 37(1)]. It separates the nasals, and is particularly developed in M. arctodites [c. 37(2)].
C. 66. This character is polymorphic in M. krahuletzi. M. medium, M. floridanum, M.
crataegense, and M. arctodites show the derived state, while 1 of 5 M. serresii and 2 of
3 M. subapenninum specimens show the primitive state, possibly as a condition of advanced life-age.
C. 83. This character is usually scored as primitive in M. krahuletzi: “zygomatic process usually not much broader posteriorly than anteriorly; process frequently sinuous and rather narrow throughout its length” (Domning & Pervesler, 2001: 22). It is primitive also in MPNRL-MAN2000; while it is polymorphic in M. floridanum, and derived in
M. medium, M. serresii, and M. subapenninum.
C. 85. This character is polymorphic in M. krahuletzi and in M. floridanum, while M.
medium and the Pliocene M. serresii and M. subapenninum always show the derived
state 2. MPNRL-MAN2000 shows state 2.
On the whole the character pattern of MV 1210 appears to be more consistent with M.
krahuletzi than with M. medium or any other Metaxytherium species, especially
concerning the absence of a nasal incisure and the shape of the zygomatic process of the squamosal.
In conclusion, I refer MV 1210 to Metaxytherium krahuletzi. Therefore MV 1210 could be considered the best preserved skull of M. krahuletzi.
MV 1210 is considered late Burdigalian – early Langhian in age (see Chapter 3); therefore it could represent the youngest record of M. krahuletzi, extending the range of the species to the early Langhian. However, a more detailed chronostratigraphic study is necessary in order to establish the exact age of the specimen.
The humerus signaled by Zbyszewski (1944) - Fragmentary sirenian remains are recorded in the Lower Miocene Portuguese deposits, ranging from the middle Aquitanian to the upper Burdigalian (Estevens, 2000; 2003a, b), and in several localities of northeastern Spain in the same Upper Burdigalian – Langhian deposits of the Valencia trough from which M. catalaunicum comes. In particular, Zbyszewski (1944, 1949) reported the occurrence of Metaxytherium remains in Portuguese Burdigalian deposits (see Appendix 2), including a right humerus from the upper Burdigalian of Quinta da Farinheira (Chelas, Lisbon) (Zbyszewski, 1944). Zbyszewski (1944) referred this humerus to M. petersi on the basis of the angle between the tubercles: this amounts to 80° in the Portuguese specimen as in M. petersi, while it was considered to be only 55° in M. krahuletzi. But now M. krahuletzi is also known to exhibit angles ranging from 60° to 85° (Domning & Pervesler, 2001: 36). I did not have the chance to examine this specimen, but the age and the characters are consistent with those of M. krahuletzi. This datum could corroborate the presence of M. krahuletzi along the European Atlantic coasts during the Early Miocene.
Metaxytherium petersi Abel, 1904
(Domning, pers. comm.)
Halitherium Cordieri Christol, sensu Peters, 1867, Jb. Geol. Reichsanst. Wien 17(2):
309.
Metaxytherium sp., Lepsius, 1882, Abh. Mittelrhein. Geol. Ver. 1(2): 165.
Metaxytherium Petersi Abel, 1904, Abh. K.-K. Geol. Reichsanst. Wien 19(2): 15, 107. Thalattosiren Petersi (Abel) Sickenberg, 1928, Denkschr. Akad. Wiss. Wien,
Math.-Nat. Kl. 101: 293.
Holotype – GBAW 2006/18/1-6, 2350-2355 Skeleton lacking skull; described by Peters (1867) and Abel (1904), termed “Typusexemplar” by Pia and Sickenberg (1934: 403); formerly in GBAW, Vienna, but now lost and probably destroyed in World War II except for distal epiphysis of radius, two carpals, one metacarpal, one phalanx, the left innominate, and a cervical vertebra.
Type locality – Hainburg an der Donau, Lower Austria. Zweite Mediterranstufe Formation. Middle Miocene (Badenian) in age.
Referred specimens – See Appendix 2
Range – Known only from the Middle Miocene deposits of Vienna Basin (Austria, Slovakia) and maybe Poland.
Remarks – This species is presently under study by Domning and Pervesler, so I do not include it in my systematic and cladistic studies.
Metaxytherium medium (Desmarest, 1822) Hooijer, 1952
(Plates 15-20; Figures 19-26)
“Lamantin fossile” Cuvier, 1809 Ann. Mus. Hist. Nat. 13: 305 pl 19 figs 12 19-23. “Moyen hippopotame fossile” Cuvier, 1821 Rech. Oss. Fossiles (2)1: 332 pl 7 fig 9.
Hippopotamus medius Desmarest, 1822 Mammologie: 388. Hippopotamus intermedius Holl 1829 Handb. Petrefactenk.: 57. Manatus fossilis Holl, 1829 Handb. Petrefactenk.: 69.
Halicore Cuvierii Christol, 1832 [partim], Ann. Sci. Indust. Midi France 2(8): 244 pl. 6:
figs. 1-3.
Halianassa Studeri Meyer, 1838 [partim] Neues Jb. Min. Geogn. Geol. Pet. 1838: 667. Halitherium cuvieri (Christol) Kaup, 1840, Neues Jb. Min. Geogn. Geol. Pet.: 675.
“Metaxytherium cuvieri” Christol, 1841, Ann. Sci. Nat. Zool. (2)15: 332 pl. 7: figs. 1,5,9-11.
M[etaxytherium]. Cordieri Christol in Blainville, 1844, Ostéogr., Genre Manatus: 130. M[anatus]. Cuvieri (Christol) Blainville, 1844 [partim], Ostéogr., Genre Manatus: 122. M[etaxytherium]. Cuvierii (Christol) Christol in Blainville, 1844, Ostéogr., Genre
Manatus: 130.
Halitherium fossilis [sic] (Holl) Gervais, 1847, Ann. Sci. Nat. (Zool.) (3)8: 221. Halianassa Cordieri (Christol) Bronn, 1848, Index Pal.: 562.
Halianassa Cuvieri (Christol) Bronn, 1848, Index Pal.: 562. Halitherium fossile Gervais, 1852, Zool. et Pal. Fra. I ed: 281.
Halitherium cordieri (Christol) Peters, 1867, Jahrb. Geol. Reichsanst. Wien 17(2): 309. Metaxytherium lovisati Capellini, 1886 , Mem. R. Accad. Sci. Ist. Bologna (4)7: 50. M[etaxytherium]. fossile (Holl) Depéret, 1895, Sitzb. Math.-Nat. Kl. K. Akad. Wiss.
Wien 104(1): 409.
Haplosiren leganyii Kretzoi, 1951, Földt Közl. 81(10-12): 438. Metaxytherium medium (Desmarest) Hooijer, 1952, Osiris 10: 114.
Holotype - MNHN Fs 2706, partial left mandible with m2-3 and roots of m1.
Type locality - Saint-Michel-de-Chasine, Maine-et-Loire, SW France. “Redonian Faluns” Formation. Serravallian-Tortonian in age.
Referred specimens - Main specimens listed in Appendix 2.
Range – Metaxytherium medium is recorded in Langhian to Tortonian deposits of Euro- North African coasts.
Emended diagnosis – Metaxytherium in which the nasal process of premaxilla contacts lacrimal [c. 1(0)] and jugal [c. 87(0)], the infraorbital foramen is moderately large [c. 13(1)], nasals are always divided in midline [c. 31(1)], the nasal incisure at posterior end of mesorostral fossa is present [c. 37(1)], the supracondylar fossa of exoccipital is usually deep and not reduced [c. 67(1)], the zygomatic process of squamosal is broader posteriorly than anteriorly [c. 83(1)], the ventral extremity of jugal usually under the orbit [c. 85(2)], the postorbital process of jugal is usually absent [c. 86(0)], the palatines extend anteriorly beyond the posterior edge of zygomatic-orbital bridge [c. 99(0)], tusks are small [c. 140(0)], and the sternum is bipartite. But none of these characters represent an autoapomorphy of the species.
The diagnosis of M. medium appears to be hard, because M. medium seems to be not distinguishable morphologically from M. crataegense and very similar to M.
floridanum, except for a postorbital process of jugal usually present [c. 86(1)] and a
shortened zygomatic-orbital bridge of maxilla [c. 14(1)] scored in this last species. I did not have the opportunity to examine these New World species and to make a direct comparison with M. medium, but a specific distinction could be questionable and these three species could represent three populations of a species with a wide distribution comparable to that of the modern dugongine Dugong dugon.
Remarks – The first specimens now referred to Metaxytherium medium were found in the Miocene deposits of the Loire Basin, belonging to the ancient “Mer des Faluns“, and referred by Cuvier (1809) to his “lamantin fossile”. In 1821, Cuvier described also an incomplete left mandible with m1-m3 from Saint-Michel-de-Chasine (Maine-et-Loire) as a “moyen hippopotame fossile”, subsequently called Hippopotamus medius by Desmarest (1822). In 1824, Cuvier described and figured this left mandible and a skullcap and some postcranial bones from Doué-la-Fontaine (Maine-et-Loire). In 1829, Holl cited some sirenian specimens as belonging to Hippopotamus intermedius and
Manatus fossilis.
In 1832, Christol considered the left mandible of Hippopotamus medius as belonging to the new species Halicore Cuvierii. In 1841, Christol reviewed the specimens referred by Cuvier (1809, 1821-1824) to the “lamantin fossile” and to the “moyen hippopotame fossile” and the two partial humeri described by Cuvier (1809) as “phoques”, and created for them the new genus Metaxytherium.
Blainville (1844) reviewed all the known sirenian fossil specimens, including those cited above, others from the Loire Basin (Angers, Rennes; Le Touraine); and specimens from the Garonne Basin (Capians; Blaye; Dax; Saint-Emilion) described also by Cuvier
(1809, 1821-1824) and Christol (1841). In 1872, Delfortrie described sirenian remains from the Garonne Basin (SW France) and referred them to the genus Halitherium. Flot (1886) described two partial skulls from Chazé-Henry as Halitherium fossile, and Cottreau (1928) described a nearly complete skeleton from Doué-la-Fontaine as
Metaxytherium cuvieri.
In 1952, Hooijer considered all the “Mer des Faluns” specimens as belonging to the same species and coined the new combination Metaxytherium medium.
In subsequent years many other sirenian specimens were found in the Miocene deposits of the ancient “Mer des Faluns”. Ginsburg and Janvier (1971) reviewed the M. medium specimens from the“Mer des Faluns”, in particular the nearly complete skeleton described by Cottreau (1928) and several bones from Chazé-Henry included in a composite skeleton. In 1942, Kaltenmark described a partial cranium from Chazé-Henry. In 2000, Plusquellec and Racheboeuf reported a new, nearly complete cranium from Tréfumel, carriére du Rouget (Côtes du Nord).
Besides the “Mer des Faluns” specimens, M. medium has been found in several Middle-Upper Miocene deposits along the Euro-North African coasts. The northernmost record is a partial skeleton from Eibergen (Netherlands) (Hooijer, 1977).
Metaxytherium medium is also recorded in the Middle Miocene deposits of the central
Paratethys/Vienna Basin (Domning, pers. comm.).
In the Mediterranean Basin, specimens presently referred, or referable, to
Metaxytherium medium have been recorded in several Middle to Upper Miocene
deposits (Domning, 1996; Bianucci et al., in press): several fragmentary remains come from different levels of the Pietra Leccese formation (Apulia, southern Italy) (Borgia et al., 1981; Bianucci et al., 2003, pers. obs.); Capellini (1886a, b), Lovisato (1902), Comaschi Caria (1957), and Carboni and Kotsakis (1983) reported several ribs, vertebrae and an innominate fragment from Middle to Upper Miocene deposits of Sardinia (Italy); Neviani (1886a, b, 1887), Capellini (1886a), Vazzana (1988), and Mario et al. (2000) reported several fragmentary, from Middle to Upper Miocene deposits of Calabria (southern Italy); and in 1970 a nearly complete skeleton was found in the Tortonian deposits of S. Domenica di Ricadi (Catanzaro, Calabria) and described by Moncharmont Zei and Moncharmont (1987). This skeleton represents the most complete Mediterranean M. medium specimen and the youngest M. medium record. Recently M. medium has also been recorded in the Tortonian deposits of Tuscany (Italy) (Bianucci & Landini, 2003).
In 1973, Metaxytherium medium remains were also recorded in the Tortonian deposits of Crete (Greece) (Symeonidis & Schultz, 1973). Recently, Marcopoulou-Diacantoni and Logos (2004) described other two partial crania with teeth from the island; Svana is now studying these specimens and also a new Cretan sirenian record.
Sirenian remains, which could be referred to Metaxytherium medium, were also found in the Miocene deposits of southern Spain (Cañigueral, 1952; Marin, 1992; Sendra, 1998). On the southern Mediterranean coasts, Metaxytherium medium is represented, or could be represented, by fragmentary remains from Miocene sediments of Malta (see Gatt, 2006), Tunisia (Geraads, 1989) and Libya (Boaz, pers. comm.).
Description –
BODY SIZE: The skeletons MNHN 1921-10 and MPUN 18403 as mounted (with extensive and partly conjectural restoration of the tail) measure about 2.8 m and 2.9 m in overall length respectively.
SKULL
Premaxilla – The rostrum is enlarged relative to the cranium [c. 3(1)]; it has a dorsal keel thin and sharp anteriorly, broadening posteriorly into a rugose surface that is flat or slightly convex. The posterior end of the rostrum is upraised to form a boss in lateral view [c. 10(1)]. The lateral edges are thin, not flaring much anteriorly; the sides of the anterior half of the symphysis are slightly convex where they are swollen by the small tusk alveoli, which extend less than half the length of the symphysis [c. 140(0)] (see tusk description below). The symphysis is long (e.g. >155 mm in MPUN 18403). The palatal surface is rugose, deeply concave; the right and the left halves form an angle of about 100-125°. The incisive foramen widens and shallows forward; its anterior end is undefined. The nasopalatine canal is subcircular to somewhat flattened dorsoventrally. The masticating surface of rostrum is slightly trapezoidal in outline. The opening of the premaxillary canal lies posteroventral to the incisor alveolus. The nasal process is long [c. 7(0)] and thin [c. 6(0)], contacts frontal [c. 9(1)], lacrimal [c. 1(1)], but not the jugal; it abuts against nasal; and ventrally it lies in a groove in the maxilla. The anterior margin of premaxilla-maxilla suture lies behind the posterior end of the symphysis. The mesorostral fossa is long, reaching beyond the level of the posterior margin of the orbit [c. 8(2)]. Its anterior end is rounded and constricted by a bulge on medial side of premaxilla; its posterior end is not well defined in any specimens because the nasal processes of frontals are usually anteriorly broken, but a small nasal incisure is usually
present [c. 37(1)], except in MPUN 18403 (app. 5 table 17). The maximum width of mesorostral fossa is anterior to the orbits. The rostral deflection varies from 49° to 66°.
Fig. 19: Cranium of Metaxytherium medium (ULCG 212901, cast of LPB 16001) in anterior view. Anterior portion of premaxilla not preserved. Scale bar: 5 cm.
Nasal – The nasals are relatively large, thick, and massive, set in sockets in anterior margins of frontals. They are anteriorly separated in midline by a nasal incisure and posteriorly by processes of frontals [c. 31(1)], except in MPUN 18403 where the nasal incisure is absent and the nasals are separated by the frontals. The dorsal exposure of the nasals is irregularly elliptical in shape, the anterior border is transversely concave for reception of the nasal process of premaxilla. The distance between the nasals is about 22-25 mm.
Ethmoidal region – Poorly preserved. Vomer – Not preserved.
Lacrimal – The lacrimal is irregular in shape, mediolaterally flattened, without a nasolacrimal canal [c. 91(1)], usually with a longitudinal protuberance on the
anterolateral surface. It abuts dorsally against premaxilla, it is sutured medially to maxilla and anteroventrally fits in socket on dorsal surface of jugal. The lacrimal does not contact the supraorbital process of frontal in MNHN 1921-10, where it is separated from the frontal by a distance of 25 mm, and in MPUN 18403, where the supraorbital processes are incomplete but the lacrimal preserved on the right side appears to be separated from the supraorbital process (in contrast to Moncharmont Zei & Moncharmont, 1987: 293). On the contrary, the lacrimal contacts the supraorbital process of frontal in LPB 16001. The jugal sometimes overlaps its lateral side anteriorly (e.g. LPB 16001).
Frontal – The supraorbital process is well developed, about 2 cm or more thick dorsoventrally with a small posterolateral corner [c. 36(1)], flattened in a horizontal plane, with a dorsal surface inclined more or less ventrolaterad [c. 43(0)]; not divided on its lateral margin. An orbicular apophysis is sometimes present. The lateral crests are in continuity with the temporal crests of parietal, and not significantly laterally overhanging, but sometimes dorsally overhanging (e. g. MNHN Fs5001). The thin lamina orbitalis of frontal [c. 38(0)] forms the medial wall of the temporal fossa and the lateral wall of a large hollow open anteriorly, whose medial wall was formed by ethmoid. The falciform anterior edge of the lamina orbitalis does not quite reach as far forward as the posterolateral corner of the supraorbital process. The internasal processes are not complete in any specimens, but they appear to be dorsally flat in midline or slightly convex.
The medial portion of the frontal roof is flat or slightly convex [c. 42(0)]. The anterior end of the frontoparietal suture lies about 3.5-5 cm behind the nasals. The interfrontal suture is visible in MNHN Fs 5001 and in LPB 16001.
Parietal – The cranial vault is more or less trapezoidal in coronal section anteriorly; about 2-3 cm thick in the anterior midline. The posterolateral corners of the roof are indented by the squamosals [c. 76(1)]. The parietals are fused to the supraoccipital and to each other in adults. A sagittal crest is absent [c. 51(1)]. The temporal crests are lyriform, highly variable in shape (TYPES A-E from Domning, 1988; see Appendix 5), low, confined to lateral edges of a nearly rectangular and flat roof (TYPE A); or at lateral edges of, but upraised above, a markedly concave roof (TYPE B); or smoothly rounded and separated by about 2 cm, with a distinctly concave roof (TYPE C); or with distinct, sharp or overhanging medial edges separated by more (TYPE D) or less than 2
cm (TYPE E); in these cases the intervening roof is deeply concave. When the temporal crests are highly lyriform, the parietal roof is dorsally less wide in the centre portion than in the anterior one at the frontoparietal suture. When the temporal crests are highly developed they present protuberances on the posterior lateral sides of the parietals, therefore the maximum width of the parietals lies at this level and not at the posterior end of the bones [c. 45(1); c. 46(1-2) scored as 2].
The internal occipital protuberance is usually distinct, sometimes pointed; the tentorium forms a straight transverse ridge; the transverse sulcus is narrow, usually distinct throughout its length and sometimes with shallow lateral pits; the bony falx cerebri may be sharp and long as far as frontoparietal suture, or more or less low, rounded, and anteriorly indistinct [c. 47(1)].
An emissary foramen is present in MNHN Fs2712, in MPUN 18403, and is nearly blocked in MNHN Fs5001. A small median bump just in front of the external occipital protuberance is sometimes present.
Fig. 20: Line drawing of the cranium of Metaxytherium medium in dorsal view, based mainly on ULCG 212901, cast of LPB 16001. Anterior portion of premaxilla not preserved. Scale bar: 5 cm.
Supraoccipital – The supraoccipital is nearly rectangular or hexagonal in outline with rounded dorsolateral corners. It forms with the after part of the parietals an angle ranging from 95° to 137°. The external occipital protuberance rises above plane of parietal roof. The median ridge below it is distinct, moderately to well developed especially in its central part, and long, usually reaching the ventral border of the supraoccipital. The nuchal crest is rounded, more or less convex anterad, but indistinct at its lateral end near the squamosal. The areas of insertion for the semispinalis capitis muscles are gently concave, oval, rugose, and face posterodorsad, extending no more than halfway to ventral end of supraoccipital; sometimes each areas of insertion is divided into two portion. The lower part of supraoccipital is slightly concave in midline and convex laterally. The lateral borders are thick and rounded, sloping outward at bottom, usually without overhanging upper corners. The suture with the exoccipitals is M-shaped with a wide central angle of about 130°-140°. Sutural surfaces for exoccipitals distinctly separated in midline, often by a notch.
Exoccipitals – The exoccipitals are not fused to the supraoccipital or to each other. They are separated in midline with a dorsally peaked foramen magnum [c. 66(1)]. The dorsolateral border is about 2-2.5 cm thick, and has a smoothly rounded posterior edge, posteriorly slightly overhanging, but not flangelike [c. 70(0)], forming a rugose surface at the level of the top of supracondylar fossa. The fossa is more or less deep, extending across entire width of occipital condyle [c. 67(1)]. The arc of the condylar articular surface subtends an angle ranging from 94° to 120°. The condyloid foramen is usually single, double on the right side of MNHN Fs5001. The condyle is oval, more or less at the level of the tip of the paroccipital process [c. 65(1)], with the ventral end turned anteroventrad. The paroccipital process is short, with rugose posterior surface and a posteroventrally-curved flange.
Fig. 21: Cranium of Metaxytherium medium (MNHN Fs5001) in posterior view. Scale bar: 5 cm.
Basioccipital – As in other dugongids it is fused with exoccipitals and basisphenoid in adults. It bears a pair of convex rugosities for the longus capitis muscles. The posterior slope is steeper than the anterior.
Basisphenoid – The sella turcica is shallow and the tuberculum sellae is nearly flat. Otherwise does not differ from that of other dugongids.
Presphenoid – It is fused with the surrounding bones. The orbitosphenoidal crest is more or less developed and may or may not overhang the optic chiasm.
Orbitosphenoid – The optic foramen lies at the level of dorsal side of sphenorbital foramen. The bony wall lateral to the optic foramen does not bear a distinct pointed process.
Alisphenoid – The posterolateral side of the pterygoid process is long, sculptured, slightly concave dorsally and convex and rugose ventrally, laterally inclined. A slight convexity continues the forward edge of the zygomatic root anteriorly. An alisphenoid canal is absent [c. 101(1)].
Pterygoid – Fused with the surrounding bones; forms the posteromedial part of the pterygoid process. The pterygoid fossa is broad and well developed, extending above level of roof of internal nares [c. 102(1)]. The ventral tips of alisphenoid and pterygoid enclose end of palatine in slot between them anteromedially. The ventral tip of the pterygoid process is enlarged, convex and rugose.
Palatine – The palatine forms the anteromedial side and (together with alisphenoid) smooth and rounded anterolateral extremity of the pterygoid process. The palate is thin, less than 1 cm thick at the level of M2[c. 16(0)]. The anterior ends of the palatines are not clearly distinct. The median sutural contact between palatines extends anteriorly beyond the posterior edge of the zygomatic-orbital bridge [c. 99(0)]. The posterior border is incised [c. 97(1)].
Maxilla – The alveolar portion of maxilla is heavy and massive; its dorsal side bears a vertical plate that helps form wall of temporal fossa and posteriorly contacts the lamina orbitalis of frontal. The edges of palatal surface are lyriform, the surface narrows forward of molars, becomes narrowest forward of anterior edge of zygomatic-orbital bridge, then widens at posterior end of rostral masticating surface. The palatal gutter is narrow and deep with rounded edges. The palatal and rostral surfaces meet in a smooth but abrupt curve. The part of palate between toothrow and strongly deflected portion of rostrum is downturned about 27°-32° from the occlusal plane. The zygomatic-orbital bridge is large and anteroposteriorly long, slightly concave on ventral side, and elevated usually more than 1 cm above alveolar margin [c. 11(1)]. The bridge is slightly posteroventrally sloping; the anterior edge is thinner than the posterior. The posterior opening of maxillo-premaxillary canal is very large. The infraorbital foramen is large, more or less rounded [c. 13(1)], not obstructed [c. 20(0)]. Sometimes a small foramen is present behind the last molar (e.g. MNHN Fs5001).
Fig. 22: Line drawing of the cranium of Metaxytherium medium in ventral view, based mainly on ULCG 212901, cast of LPB 16001. Anterior portion of premaxilla not preserved. Scale bar: 5 cm.
Squamosal – The squamosal indents the posterolateral corners of the parietal roof [c. 76(1)] and extends to the temporal crests, except when they do not reach the posterior end of the parietal. The posterior edge along mastoid foramen is rounded. The prominent sigmoid ridge extends from dorsal end of mastoid foramen to ventral end of posttympanic process, forming a strong laterad-projecting flange with its posteroventral edge broad and rugose. The surface of cranial portion dorsal to zygomatic root is not inflated or bulging. The postglenoid process and postarticular fossa are distinct and well developed. The temporal condyle is irregularly oval. The processus retroversus is moderately inflected, not projected below line of suture with jugal [c. 77(1)]; its posterior end is smooth, sometimes with dorsal and ventral protuberances. The posttympanic process is present [c. 73(1)]. The external auditory meatus is nearly rounded [c. 82(1)] and less than 1 cm long. The zygomatic process is roughly lozenge-shaped in lateral view, broader posteriorly than anteriorly [c. 83(1)]; its medial side is relatively flat or concave, slightly inclined inward dorsally; its posterodorsal edge is convex laterad, medially turned in; its anterior tip is long, but it does not reach the level of the posterior extremity of supraorbital process.
Jugal – The ventralmost point lies ventral to orbit [c. 85(2)]. The ventral tip is thickened and the border behind it is thickened and roughened. The preorbital process is relatively flat and thin [c. 88(1)], but with a longitudinal crest on the anterolateral border, and dorsally it does not contact the premaxilla [c. 87(0)]. The ventral margin of orbit is broad, concave but not significantly overhanging [c. 90(0)]. The zygomatic process is nearly horizontal, tapered in outline, longer than diametre of orbit [c. 89(0)]; its lateral side is flat, its medial side is concave. A raised postorbital process in front of tip of squamosal is usually absent [c. 86(0)].
Fig. 23: Cranium of Metaxytherium medium (ULCG 212901, cast of LPB 16001) in lateral view. Anterior portion of premaxilla not preserved. Scale bar: 5 cm.
Ear Region – (see also Flot, 1886):
Periotic – The periotic sits in a closely fitting socket in the squamosal. The lateral surfaces are nearly smooth; the pars mastoidea and the tegmen tympani (pars temporalis) are separated by a groove about 6 mm wide, not covered by the overhanging borders of the pars mastoidea and the tegmen tympani. The pars mastoidea bears a slightly raised, rugose triangular area (processus fonticulus) on the lateral surface which fits into the mastoid foramen. The endolymphatic foramen is narrow and slitlike. The cavity above aquaeductus vestibuli is elongated laterad over medial shelf of tegmen tympani. The medial extremity of pars petrosa is rounded in outline; the dorsal side is more or less flat or with variable low convexities. The ventral end of the promontory is blunt. The fossa for origin of the stapedius muscle is present and distinct.
Tympanic – The distal border is V-shaped in outline with a blunt tip. The posterior edge is sinuous. The anterior edge is more or less straight. The external side is smooth and slightly sinuous. The internal side is convex. The proximal border is more or less concave. The posterior branch forms an angle of about 105° and is proximally expanded into a broad, nearly flat surface, which contacts the pars mastoidea; along its proximolateral edge a ridge and the sulcus tympanicus for the tympanic membrane are present. The anterior branch is short with a rounded tip incised by a longitudinal groove. Malleus – The area of orbicular apophysis on posterior end is convex. The processus muscularis is massive, swollen and elliptical in outline. A sharp horizontal ridge runs forward on lateral side from dorsal end of manubrium. The external edge of manubrium is convex. The articulation with the incus is constituted by two facets: the anterior one is larger and flat, the posterior is smaller and saddle-shaped. The angle between the incudal facets is about 70°.
Incus - As in other Metaxytherium, the anterolateral edge of crus breve is very slightly convex in outline; the crus longum (processus lenticularis) is more developed, cylindrical, and curves sharply inward. The ventral side is irregular with small tubercles and the articular surface for the malleus.
Stapes – The stapedial foramen is well developed, about 4 mm above base; the head end flattened anteroposteriorly, the basal end dorsoventrally. The head facet has no distinct overhanging lip, just a lateral protuberance. The basal facet is quadrilateral and offset backward. A deep groove for facial nerve is present.
Mandible – The condyle is elliptical, overhanging; its dorsal surface is slightly concave in the centre. The mandibular notch is deep. The posterior medial edge of condylar process is thin and sharp. The edge of angle is thickened. A processus angularis superior is absent and the posterior border of the mandible has a broadly convex outline beginning well below the condyle [c. 125(2)]. The internal and external pterygoid fossae are large. The coronoid process is well developed, with a sharp, backward-pointing tip. The anterior border extends far anterior to the base [c. 126(1)].
The horizontal ramus is dorsoventrally broad [c. 128(1)], with ventral border strongly concave [c. 122(3)], not tangent to angle [c. 129(1)]. A small protuberance is usually present below m1.
The mental foramen lies at the level of deflection of dorsal edge of ramus; it is very large with an anterior groove [c. 123(1)]. The anterodorsal margin of the mental foramen is formed by a sharp ridge. The deflection of masticating surface varies from