CAPITOLO 1 : INTRODUZIONE
1.1 IL CORALLIGENO: CENNI STORICI SULLE CONOSCENZE
Nel Mar Mediterraneo, i sistemi litorali profondi (tra i 25-30 m. e i 150-200 m. di profondità) sono generalmente colonizzati da popolamenti bentonici dominati da organismi calcarei denominati “coralligeno”.
Questi popolamenti si sviluppano su fondi rocciosi o su pianori sabbiosi e sono costituiti da blocchi di bioconcrezioni (concrezioni di origine organica) originati principalmente da talli morti di alghe calcaree crostose e secondariamente da animali biocostruttori.
Tali blocchi costituiscono un substrato duro sul quale si sviluppano popolamenti altamente diversificati.
I primi studiosi ad utilizzare il termine coralligeno sono stati Marion (1883), per designare le formazioni concrezionate (graviers coralligènes) presenti nel Golfo di Marsiglia tra i 30 e i 70 m. e Pruvot (1894, 1895) nella regione di Banyuls-sur-mer.
Il termine coralligeno significa produttore di corallo e si riferisce all’abbondanza di corallo rosso (Corallium rubrum) presente in questo tipo di fondo.
Pérès & Picard (1964) definirono la biocenosi coralligena come un climax di una serie evolutiva di comunità che si sviluppano nel piano circalitorale in condizioni di stabilità dei parametri ambientali quali salinità e temperatura e di forte riduzione della luce.
La biocenosi coralligena secondo questi Autori presenta due caratteri essenziali:
a) è legata a substrati duri tanto primari tanto secondari (derivati da concrezionamento) ; b) è nettamente sciafila nonostante sia a dominanza vegetale.
Inoltre sempre secondo Pérès & Picard tale biocenosi esiste in due tipi di stazioni : 1) di orizzonte inferiore della roccia litorale, cioè ai piedi delle falesie sommerse e di
superfici rocciose emergenti dal fondale mobile, quando la profondità determini una sufficiente diminuzione della luce o all’ingresso di grotte ;
2) di piattaforma, con substrato duro realizzato da un concrezionamento biologico, principalmente dovuto ad alghe calcaree, a partire da un fondo originariamente mobile. Le condizioni ecologiche che portano alla formazione della biocenosi coralligena sono: salinità e temperatura relativamente costanti ed illuminazione molto ridotta anche se sufficiente allo sviluppo di alghe calcaree.
Laubier (1966) a seguito degli studi condotti sulle formazioni coralligene delle coste di Albères (Francia), sostenne che non si poteva parlare di una singola biocenosi del coralligeno, ma che questo fosse un complesso di biocenosi che popolano i numerosi microhabitat creati all’interno di queste formazioni.
Sarà & Pulitzer- Finali (1970) continuano il loro riesame critico sulla classificazione dei fondi coralligeni, affermando che anche per il coralligeno pugliese e ligure, gli organismi costruttori determinano la formazione di un substrato duro secondario che è alla base di un complesso di biocenosi, in cui cioè occorre distinguere gli elementi dell’epifauna che vivono alla superficie della formazione e la ricca endofauna che popola gli interstizi colmi di sedimento.
Quindi secondo questi Autori Il concetto di biocenosi coralligena è stato applicato da Pérès & Picard a due popolamenti distinti. Il primo sarebbe costituito da una epifauna e da una epiflora di affinità sciafila che si sviluppa su substrati primari (non concrezionati), quando si verifica una notevole attenuazione della luce; il secondo si riferirebbe alla formazione di blocchi concrezionati prodotti principalmente dall’attività di alghe calcaree,
ma anche di alcuni animali, in cui un ruolo importante dal punto di vista del trofismo e della dinamica del complesso biocenotico viene svolto dalla ricca endofauna.
Per concludere, secondo Sarà & Pulitzer- Finali (1970) i termini “biocenosi coralligena” e “coralligeno” nel senso di una singola biocenosi devono essere scartati.
L’uso dell’ aggettivo coralligeno è invece opportuno solo con l’accezione datagli da Marion (1883) e poi da Pruvot (1894), ad indicare cioè delle formazioni derivanti da concrezionamenti di origine biologica.
Gli elementi sciafili dell’epifauna e dell’epiflora andrebbero invece inquadrati nelle biocenosi di substrato duro del piano circalitorale.
Le formazioni coralligene sarebbero poi riconducibili a due tipologie: quelle infralitorali, che si trovano tra i 20 e 30 m. di profondità e quelle circalitorali più profonde.
Cinelli & Colantoni (1993) adottano questo tipo di distinzione ma suggeriscono l’uso del termine più generale di “popolamento” al posto di biocenosi per indicare il complesso di organismi associati alle formazioni coralligene.
Nel 2006 Ballesteros et al. hanno definito l’habitat coralligeno un substrato duro di origine biogena prodotto principalmente dall’accumulo di alghe coralline incrostanti che crescono in condizioni di scarsa illuminazione.
1.2 STRUTTURA E CARATTERISTICHE DEI POPOLAMENTI
1.2.1
Biocostruzione
Per biocostruzione s’intende la capacità di alcuni organismi di formare strutture calcaree persistenti in grado di aumentare l’eterogeneità del substrato.
I popolamenti si sviluppano su fondali rocciosi o sabbiosi, in acque con valori re-lativamente costanti di temperatura, idrodinamismo, salinità e con illuminazione ridotta.
Dal momento che il popolamento è costituito principalmente da alghe, la diminuzione dell’irraggiamento deve essere compreso tra 2-3% e il 0-0.5%, in quanto una diminuzione maggiore non consentirebbe lo sviluppo di questi organismi vegetali (Garrabou & Ballesteros, 2000). Questi requisiti ecologici li possiamo ritrovare nel Mediterraneo a differenti profondità in relazione alle condizioni ambientali locali.
E’ possibile, infatti, trovare il coralligeno già ad una profondità di 18 m lungo le coste Toscane, dove le acque presentano un elevato carico di sedimenti, mentre ad esempio lungo le coste della Corsica il coralligeno non compare prima dei 60 m, a causa dell’elevata trasparenza delle acque.
Gli organismi biocostruttori, sia animali che vegetali, producono una struttura calcarea che dopo la morte rimane fissa al substrato. L’accumulo di queste strutture, dopo la morte degli organismi, può condurre alla formazione di un substrato duro permanente, in cui solo lo strato più esterno è vivo (Laborel, 1987). Queste formazioni possono crescere per centinaia d’anni o com’è stato recentemente calcolato per alcuni banchi coralligeni, anche per migliaia d’anni (Sartoretto et al.1996).
Nel Mediterraneo è stato stimato un tasso di produzione dei carbonati ad opera degli organismi biocostruttori dell’ordine di 103 g CaCO
3 m -2
produzione paragonabile a quella stimata per i reef tropicali (Chave et al., 1972, Heiss, 1995).
Immaginando di fare una sezione verticale lungo la formazione coralligena vedremo, se presente, un primo strato di roccia con la presenza di organismi endoliti, un secondo strato formato da concrezioni coralligene con fauna cavitaria all’interno, un terzo strato incrostante vivente formato da alghe, poriferi, briozoi, antozoi ecc., un quarto strato caratterizzato da alghe, antozoi, poriferi, briozoi, ascidie, ed infine un quinto strato colonizzato da gorgonie con associato una fauna epibionte.
1.2.2 Componente algale
Le specie più rilevanti dal punto di vista quantitativo per lo sviluppo del coralligeno appartengono al phylum Rhodophyta e alle famiglie Corallinaceae e Peyssonelliaceae.
Le Corallinaceae dette comunemente alghe coralline incrostanti sono i più importanti organismi costruttori del coralligeno (Laborel 1961, Faubier 1966, Sartoretto 1996).
La tassonomia di questo gruppo di alghe è molto complessa e la nomenclatura di queste specie cambia continuamente.
Feldmann (1937) identificò le quattro più importanti alghe calcaree responsabili della creazione del coralligeno nella regione di Banyuls-sur-mer: Lithophyllum frondosum,
Neogoniolithon mamillosum, Mesophyllum alternans tra le Corallinaceae e Peyssonnelia rosa-marina tra le Peyssonnelliaceae.
Le stesse specie sono riportate per quanto riguarda il coralligeno studiato nel Golfo del Leone ( Boudouresque 1973, Ballesteros 1992).
Ballesteros (2000) ritiene che tra le Peyssonelliaceae sia da considerarsi fondamentale per la biocostruzione del coralligeno oltre a P. rosa-marina anche P. rubra. E’ interessante notare che nonostante il genere Peyssonnelia sia molto abbondante come alga incrostante vivente, è completamente assente nei ritrovamenti fossili (Bosence 1985, Ballesteros 1992); questo è dovuto al fatto che il contenuto di carbonato di calcio di queste specie è più basso rispetto a quello delle altre corallinacee (Laubier 1966, Ballesteros 1992).
Sul substrato secondario concrezionato si verifica l’ insediamento e lo sviluppo di altre specie vegetali ed animali che creano il complesso biocenotico del coralligeno (Bianchi, 2001).
Secondo Ballesteros (2006) nelle acque più superficiali si sviluppa l’associazione
Lithophyllo-Halimedetum tunae, caratterizzata dall’alga costruttrice Mesophyllum alternas ,
dalle le alghe erette, che si sviluppano sul concrezionamento, Halimeda tuna e Flabellia
petiolata e da numerose specie del genere Peyssonnelia. Più in profondità le alghe erette
tendono a diminuire e dominano altre specie di alghe coralline costruttrici: Lithophyllum
frondosum, L. cabiochae e Neogoniolithum mamillosum, Peyssonnelia rosa-marina;
l’associazione che caratterizza questo popolamento è il Rodriguezelletum strafforellii. Altre alghe molto comuni in questa associazione appartengono ad alcuni taxa di Rhodophyta foliose come i generi Kallymenia, Fauchea, Sebdenia, Rhodophyllis, Predea o ad alghe Chlorophyta incrostanti come Palmophyllum crassum.
Studi sulla componente specifica del coralligeno hanno portato all’identificazione di specie presenti da 7700 anni ed hanno evidenziato che non ci sono stati cambiamenti di specie (Sartoretto et al. 1996).
La componente algale delle comunità del coralligeno comprende tra il 33 ed il 48 % di tutta la flora del Mediterraneo (Boudouresque 1973); tra le circa 315 specie algali sono compresi inoltre numerosi endemismi (Boudouresque 1985).
1.2.3 Componente faunistica
La fisionomia del coralligeno è spesso caratterizzata da molte specie animali, alcune delle quali importanti per l’accrescimento della bioconcrezione stessa, come ad esempio i madreporari (Corallium rubrum, Leptopsammia pruvoti, Madracis pharensis,
Cladocora cespitosa, ecc.) ed i briozoi (Myriapora truncata, Pentapora fascialis, Sertella beaniana, ecc.).
Tra le altre specie animali troviamo spugne (Petrosia ficiformis, Axinella cannabina,
Chondrosia reniformis, ecc.), cnidari (Parazoanthus axinellae, Cerianthus membranaceus,
ecc) ed ascidie (Halocynthia papillosa, ecc.).
Laubier (1966) fu il primo autore che enfatizzò la grande biodiversità delle comunità coralligene identificando 544 specie di invertebrati nella regione di Banyuls.
Più tardi studiando le comunità coralligene di Marsiglia, Hong (1980) trovò 682 specie mentre altri autori (Ros et al. 1984) riportarono 497 specie di invertebrati nelle Isole Medes.
In accordo con Hong (1982) quattro differenti categorie di invertebrati possono essere distinte in base alla loro posizione e al significato ecologico nella struttura del coralligeno:
1.Fauna che contribuisce al consolidamento della struttura creata dalle alghe calcaree (briozoi, policheti, serpulidi, coralli e spugne ); include il 24% delle specie totali.
2.Criptofauna che colonizza le cavità più piccole della struttura; rappresenta il 7% delle specie totali e include molluschi, crostacei e policheti.
3.Epifauna e endofauna che ricopre il 67%. 4. Specie che erodono il substrato (1%).
1.2.4
Bioerosione
Feldmann (1937) descrive l’abbondanza di organismi che erodono le concrezioni calcaree, in particolare la spugna Cliona viridis , il bivalve Lithophaga litophaga e e altri anellidi.
Possono essere distinti tre tipi di organismi che erodono:
- “Brucatori” rappresentati dai ricci di mare (Laubier 1966) le cui variazioni in abbondanza e taglia influenzano grandemente la quantità annuale di carbonato di calcio eroso; il ruolo bioerosivo dei ricci è però limitato dalla loro bassa densità (1-3 individui in 25 m2) (Sartoretto 1996).
- “Microperforatori” che includono alghe blu-verdi (cianobatteri), alghe verdi e funghi (Hong 1980).
- “Macroperforatori” che comprendono i molluschi (Lithophaga litophaga), sipunculidi, policheti e spugne (Sartoretto 1996, Martin & Britayer 1998).
Il risultato finale tra costruzione/erosione delle concrezioni è una complessa struttura dove possono essere distinti molti microhabitats. Fattori ambientali come luce, movimento dell’acqua, tasso di sedimentazione possono variare notevolmente anche tra concrezioni molto vicine tra loro.
La grande eterogeneità ambientale genera popolamenti diversi che coesistono in uno spazio ridotto.
1.2.5 Biodiversità
Secondo Laubier (1966) il coralligeno è un ecosistema maturo ad elevata biodiversità, caratterizzato da una complessa struttura ecologica.
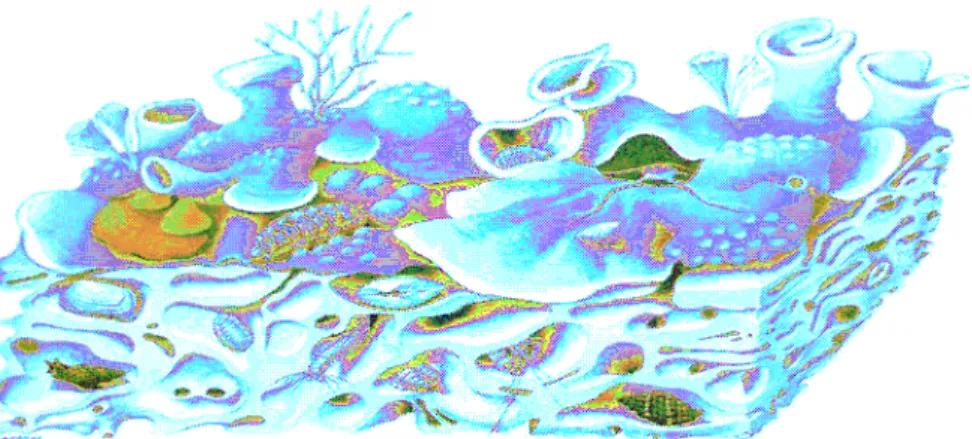
L’elevata biodiversità è dovuta all’eterogeneità del substrato, creata dall’attività degli organismi costruttori, che produce un aumento di microhabitat ed alla stratificazione del popolamento (Cocito et al. 2001, Sarà 1999; Ros et al 1984) (Figura 1.2.5).
Le stime di ricchezza specifica della comunità coralligena giungono ad un numero di 1241 specie di invertebrati e di 315 specie di macroalghe (Boudouresque 1973).
Per quanto riguarda la produzione di biomassa sono stati calcolati per il coralligeno di Banyuls-su-mer (Francia) valori fino a 400 gr/m2, paragonabili con quelli dei reef tropicali.
Le comunità del coralligeno costituiscono quindi il secondo “hot-spot” di diversità specifica nel Mediterraneo, dopo le praterie di Posidonia oceanica ( Boudouresque 2004).
Considerando la ricchezza specifica e la complessità strutturale emersa dai pochi studi che sono stati fino ad oggi compiuti sulla biodiversità del coralligeno (Laubier 1966; Pérès & Picard 1964, Ros et al.1985), è probabile che risiedono in questa comunità più specie rispetto ad ogni altra presente in Mediterraneo.
E’ da segnalare inoltre che molte specie che hanno una valenza economica nel settore della pesca, trovano rifugio in questo ambiente come ad esempio molti crostacei quali le aragoste o le magnose , numerose specie di pesci quali cernie o i gronchi.
La conoscenza del coralligeno è quindi molto importante, sia per la sua elevata biodiversità, sia per il ruolo che riveste nell’ambito della pesca e del turismo subacqueo. Recentemente infatti con lo svilupparsi l’attività subacquea, le formazioni coralligene sono diventate un’attrazione turistica a causa della ricchezza di specie e dalla varietà di forme e colori che si possono ammirare in quest’ambiente.
Questo aspetto rende questi popolamenti un’importante risorsa anche all’interno del settore del turismo ecocompatibile di grande importanza socio-economica per i parchi marini ed aree protette.
Il coralligeno è considerato inoltre un ottimo indicatore della qualità delle acque essendo costituito principalmente da organismi sessili .
Figura 1.2.5 Sezione di un banco coralligeno che mostra l’eterogeneità del substrato e i differenti microhabitats ( disegno di J.Corb).