2 Materiali e metodi
2.1 Animali
Gli animali utilizzati in questo lavoro, sono topi transgenici RasGRF1 Knock Out (RKO) ed
ERK1 Knock Out (EKO) testati, in ogni esperimento, in associazione ai loro fratelli Wild Type (WT), serviti come gruppo di controllo. La distinzione dei genotipi è stata effettuata
nei laboratori di biologia molecolare dell’Istituto di Neurofisiologia del Consiglio Nazionale delle Ricerche di Pisa, attraverso la Reazione polimerascia a catena (polymerase chain reaction, PCR), una tecnica che permette di ricavare un alto numero di copie del DNA di interesse - senza i tagli e le ricuciture tipici del clonaggio, purché se ne conosca almeno in parte la sequenza. I RKO sono stati ottenuti secondo le modalità indicate da Brambilla e collaboratori (Brambilla et al. 1997). In questi topi la proteina che manca è RasGRF1. È importante sottolineare che in questa linea genetica si manifesta il cosiddetto fenomeno dell’imprinting genico parentale. Si tratta di un meccanismo di regolazione epigenetica a seguito del quale alcuni loci o intere regioni cromosomiche di mammifero, vengono inattivate dopo la fecondazione dell’uovo o durante lo sviluppo embrionale, in modo tale che nell’individuo neoformato, solamente un allele (di derivazione paterna o materna) risulti attivo. A seconda, quindi, dello specifico gene, si ha inattivazione paterna o materna ed espressione monoallelica. Nel caso specifico di RasGRF1 solamente gli alleli provenienti dal cromosoma 9 del padre saranno espressi nella progenie. Quindi solo il genotipo del padre (+/+, +/- o -/-) è il fattore determinante nel prevedere il genotipo della progenie, mentre quello della madre non è influente in quanto darà sempre luogo ad alleli repressi. In seguito a questo fenomeno, non sono state riscontrate differenze di espressione proteica tra i topi transgenici eterozigoti (RasGRF1 +/-) ed omozigoti (RasGRF1 -/-). Da un punto di vista fenotipico, i topi RKO non presentano né differenze istologiche, né morfologiche, rispetto ad i fratelli WT (+/+). Inoltre, sono vitali ed in grado di riprodursi.
Gli EKO sono stati ottenuti secondo le modalità indicate da Pagès e collaboratori (Pagès et
al. 1999). In questi topi la proteina mancante è ERK1. Differentemente da quanto fatto con
gli animali della linea dei RasGRF1, non sono stati effettuati esperimenti con gli animali eterozigoti della linea di ERK1: si sono quindi utilizzati i topi ERK1 -/- mentre i fratelli ERK1 +/+ sono stati inseriti nel gruppo dicontrollo.
Tutti gli animali sono stati generati nel laboratorio di Neurobiologia della Scuola Normale Superiore presso l’Istituto di Neurofisiologia del Consiglio Nazionale delle Ricerche di Pisa. Gli accoppiamenti sono stati eseguiti con topi C57BL/6 (“linea genetica pura”).Nel complesso i topi utilizzati negli esperimenti sono tutti di età comparabile. La loro assegnazione nei vari gruppi esaminati, è stata resa il più omogenea possibile, tenendo conto del genere e del genotipo. Gli animali sono stabulati in condizioni di temperatura e umidità costanti, con un ciclo di luce/buio di 12:12 ore, con inizio della fase di luce alle 6:00 A.M. Acqua e cibo, sono a disposizione ad libitum. I test comportamentali, e la stabulazione degli animali, sono stati condotti in linea con i protocolli approvati dal Ministero Italiano per la Ricerca Scientifica.
2.2 Test comportamentali
Con l’intento di rendere il disegno sperimentale il meno stressante possibile per gli animali e per aumentare il loro grado di abituazione alla manipolazione, abbiamo messo in atto degli accorgimenti mirati. Innanzitutto, a circa due mesi dopo la nascita e prima di essere sottoposti ai vari test, per ogni topo è stata eseguita la manovra cosiddetta di handling (dall’inglese manipolazione). Si tratta di una semplice strategia finalizzata a far abituare gli animali ad essere manipolati dallo sperimentatore e consiste nel sollevarli per la coda (prassi comune per prelevare un topo) e tenerli - liberi di muoversi - nel palmo della mano senza nessun altro tipo di stimolazione. Questa era eseguita per 4-5 giorni prima dei compiti. Inoltre, abbiamo eseguito i test in orari simili e in particolare seguendo un ordine temporale preciso. Abbiamo notato, infatti che tra gli esperimenti eseguiti ve ne sono alcuni particolarmente debilitanti per i topi, a differenza di altri.
Sulla base di queste osservazioni, quando ci siamo trovati ad eseguire test diversi sugli stessi soggetti, abbiamo eseguito primariamente gli esperimenti con un carico stressogeno minore, seguiti da quelli più impegnativi. Prima dell’ Object Recognition Test (ORT) è stato eseguito l’open field (OF)
che consente di far abituare ulteriormente l’animale all’ambiente sperimentale.
Dato che l’arena utilizzata nel test per la memoria di riconoscimento, è la stessa di quella dell’open field, abbiamo scelto di eseguire l’ORT dopo soli 2 giorni di riposo dall’OF. Questo per aumentare ulteriormente l’abituazione dell’animale al setting sperimentale. Per assicurare una condizione il più obiettiva possibile, nell’ORT è stata messa in atto la procedura dell’esperimento in doppio cieco, in cui lo sperimentatore, al momento dell’esecuzione del test, non è al corrente del genotipo murino che sta valutando. In questo modo, sono eliminate anche eventuali tendenze involontarie da parte dello sperimentatore, a deviare i risultati verso l’ipotesi preliminare che si vuole confermare.
2.2.a Open field
Il test è stato eseguito in un’arena di forma quadrata con bordi rialzati 60x60x30 cm e con le pareti interne rivestite con carta nera lucida. Il pavimento, in materiale vinilico trasparente (lexan), permette la visione di un cartoncino, posto sotto l’apparato, su cui sono tracciate una griglia con celle di 10x10 cm e il centro dell’arena di 20x20 cm. Questo è utilizzato come riferimento della motilità degli animali. L’arena è collocata in una cabina grande 100x180 cm, omogeneamente illuminata e isolata dall’ambiente circostante. Durante lo svolgimento del test, al suo interno si trovano, oltre allo strumento, gli animali e lo sperimentatore.
Il test dell’open-field (OF) viene utilizzato come test preliminare al fine di indagare parametri importanti per lo svolgimento di tests successivi, come l’attività locomotoria, l’attività esploratoria e la motivazione (Schildein et al, 2002). L’importanza di eseguire questa prova prima delle altre, sta nel fatto che riesce a far emergere eventuali deficit locomotori o fenotipi ansiosi, che potrebbero andare ad inficiare i risultati successivi e di conseguenza l’interpretazione del fenotipo del soggetto studiato. La teoria su cui si basa il test è che i topi abbiano una naturale tendenza alla attività esploratoria. Più in particolare, i roditori preferiscono ambienti poco illuminati, poco rumorosi e di dimensioni non molto ampie. Queste nozioni possono dare delle specifiche precise nell’interpretare il loro comportamento nell’apparato sperimentale. Sulla base di tali considerazioni vengono presi in esame indici attendibili dello stato ansiogeno dell’animale. Ad esempio, una riduzione dell’attività locomotoria, la diminuzione del tempo trascorso al centro dell’arena (zona ad alta vulnerabilità per l’animale) e comportamenti d’ansia come la defecazione, sono tutti indici delle caratteristiche ansiogene insite nella strumentazione. Inoltre, si nota una riduzione dell’attività esploratoria totale, data dall’abituazione dell’animale allo strumento che precedentemente risultava non familiare e quindi di maggior interesse. Il protocollo seguito deriva, con alcune modifiche, da quello presentato da Crawley e collaboratori (Crawley et al. 1997b). L’arena viene posizionata a rombo rispetto allo sperimentatore. Al fine di eliminare ogni sorta di riferimento olfattivo, dopo ogni topo inserito nell’apparato, questo viene pulito con una soluzione di alcool diluito. Il soggetto viene prelevato dalla
home cage e inserito nell’arena, evitando di provocargli uno stato di elevata attivazione che potrebbe ripercuotersi sul normale svolgimento dell’esperimento. Ogni singolo soggetto è rilasciato dalla medesima posizione di partenza. Una volta che il topo viene lasciato dallo sperimentatore, viene avviato un cronometro per la misurazione dei 5 minuti durante i quali è osservata la sua attività esplorativa. In particolare, gli indici che abbiamo misurato in questo test sono diversi: il numero totale degli attraversamenti che il roditore effettua sulla griglia (collocata sotto il pavimento trasparente), il numero totale dei rearings (sollevamenti sulle zampe posteriori), dei groomings (pulizia del muso), dei freezings (immobilità posturale) e degli escrementi prodotti. In aggiunta viene rilevato il tempo totale di permanenza dell’animale al centro dell’apparato. Al fine di fugare la possibilità di eventuali problematiche legate alla motilità degli animali testati, abbiamo prestato maggiore attenzione all’attività locomotoria (basal locomotion). In particolare, abbiamo valutato la distanza percorsa nell’intervallo di tempo, espressa in cm e la velocità di movimento degli animali. Scaduti i 5 minuti sperimentali, il topo viene ricollocato nella propria gabbia. In seguito, se testato ad una distanza temporale, ad esempio, di 24 ore, l’animale presenterà un effetto di abituazione all’ambiente che si tradurrà in una diminuzione del tempo totale di esplorazione. Durante lo svolgimento dell’OF per determinare l’attività locomotoria degli animali è stato usato il software EthoVision. L’EthoVision (© Noldus Information Technology b.v. 2002), è un software finalizzato alla registrazione (e analisi) automatica del movimento e del comportamento degli animali. Quando si pongono degli animali in un’arena, questi vengono videoregistrati ed i relativi dati sono automaticamente elaborati dal programma. Quando l’esperimento è terminato, è possibile riesaminare i filmati dei soggetti, ed analizzare obiettivamente i valori in modo da giudicare una vasta gamma di variabili dipendenti (parametri), come la velocità, la direzione, il tempo trascorso in varie locazioni (zone), ecc.
2.2.b Object Recognition Test
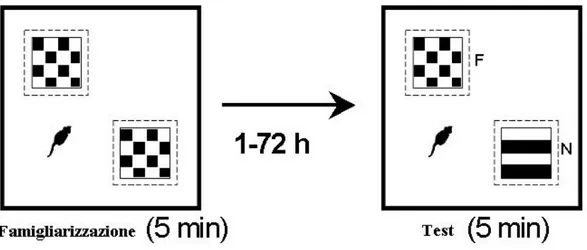
Per quanto riguarda l’apparato, viene utilizzato lo stesso descritto per l’open field test. La particolarità in questo caso è che sotto il pavimento trasparente, è posto un cartoncino sul quale sono disegnate delle sagome 15x15 (cm) indicanti il punto in cui dovranno essere collocati i due cubi che serviranno da stimolo. Sullo stesso cartoncino, è disegnato anche un contorno di esplorazione 18x18 (cm) immediatamente circostante il profilo, che indica l’area che lo sperimentatore utilizza come metro di riferimento per la valutazione dell’esplorazione. Al fine di evitare una preferenza dell’animale per uno stimolo, in base alla sua collocazione nell’arena, le sagome dei quadrati disegnati sul supporto di carta, sono equidistanti dal centro dell’arena.Gli stimoli sono due dei cubi trasparenti di Plexiglas® 15x15x15 (cm), all’interno dei quali è possibile inserire dei patterns visivi cartacei in bianco e nero, raffiguranti, ad esempio reticoli, scacchi oppure un indistinto fondo nero.
L’intero apparato si trova in una cabina grande 100x180 (cm), omogeneamente illuminata, isolata dall’ambiente circostante. Durante il test sono presenti all’interno della cabina solo i soggetti, lo strumento e lo sperimentatore.
Il protocollo originario di Ennaceur era caratterizzato dall’uso di oggetti di forma diversa, ma dimensioni paragonabili (Ennaceur A., et al, 1988). Noi abbiamo deciso di apportare delle modifiche, in riferimento a quello utilizzato da Capsoni e colleghi (Capsoni et al., 2000). In particolare, abbiamo utilizzato come stimoli, due cubi di dimensioni identiche, distinguibili solamente in base ai patterns collocati all’interno. Questa scelta si è dimostrata valida al fine di eliminare l’eventuale effetto che le conformazioni degli oggetti, possono esercitare sulla preferenza degli animali. Si tratta quindi, di un protocollo in cui la familiarizzazione, e la memorizzazione conseguente, sono basate esclusivamente su input di tipo visivo.
La scelta di effettuare questo test, si basa sul fatto che, dai dati in letteratura, emerge da anni la sua capacità di indagare un aspetto interessante della memoria di riconoscimento: la discriminazione di familiarità. Alla base di questo, troviamo la naturale tendenza dei roditori, ad esplorare più accuratamente, e più a lungo, stimoli nuovi (novel), in raffronto a quelli già esperiti precedentemente (familiar). Data questa particolarità, è necessario far sì che il topo familiarizzi con una coppia di patterns (fase di familiarizzazione). Dopo un determinato intervallo temporale segue la fase di test, in cui uno dei due stimoli è sostituito con un altro nuovo.
Possiamo assumere, come indice del mantenimento in memoria della traccia relativa allo stimolo familiar, la condizione in cui quest’ultimo viene esplorato meno tempo, rispetto al novel.
Al fine di ottenere una curva di memoria, possiamo intervenire variando l’intervallo temporale tra la fase di familiarizzazione e quella di test. Il test si divide in tre fasi sequenziali: la fase di abituazione, la fase di familiarizzazione e la fase di test. La fase di abituazione è inizialmente indispensabile, sia per permettere all’animale di prendere confidenza con l’apparato, che per fare in modo che il roditore risponda meno al contesto e più vigorosamente alla presentazione degli stimoli. L’animale viene inserito nell’arena (priva di stimoli) per due giorni consecutivi, e se ne osserva l’attività esploratoria, per un
totale di 5 minuti. Nel nostro caso, dato che è stato realizzato precedentemente l’open field, siamo passati direttamente alla fase successiva dell’ORT, dopo due giorni di pausa dal termine dell’OF, attribuendo a quest’ultimo la valenza di abituazione.
Nella fase di familiarizzazione, vengono inseriti nell’arena i due cubi contenenti due pattern visivi uguali. L’arena viene posizionata a rombo rispetto allo sperimentatore, in modo che quest’ultimo sia equidistante sia dal cubo di sinistra che da quello di destra, cosicché non si pregiudichino le strategie comportamentali dell’animale. Lo strumento viene pulito con una soluzione di alcool diluito, in modo da eliminare ogni riferimento olfattivo. Dopodiché, si preleva delicatamente l’animale dalla home cage e si introduce dentro l’arena. Tutti gli animali vengono rilasciati dalla stessa posizione di partenza.
Lo sperimentatore cronometrerà il tempo (in secondi) di esplorazione dei singoli cubi, per un totale di 5 minuti. Si considera un pattern esplorato, quando il topo supera il contorno di esplorazione, a patto che stia con il muso rivolto verso lo stimolo. Il solo attraversamento del contorno, non deve essere considerato come esplorazione. L’animale deve, infatti, mostrare attenzione verso il pattern.
Al termine dei 5 minuti, si ripone il topo nella gabbia, si pulisce nuovamente lo strumento, ed è quindi possibile iniziare la prossima sessione con un altro animale.
Nella fase test viene inserito nell’arena un pattern novel, e mantenuto l’altro il familiar. Questa fase permette di ottenere una misura della memoria di riconoscimento. Il test viene effettuato ad intervalli temporali precisi: 1 ora, 24 ore, 48 ore e 72 ore a partire dalla fase di familiarizzazione.
Fatto salve queste modifiche, si procede esattamente come nella fase precedente.
Ogni fase di test ha la propria familiarizzazione. Per esempio, terminato il test a 1 ora, gli animali verranno lasciati riposare per due giorni, dopodiché affronteranno una nuova fase di familiarizzazione con due nuovi stimoli (mai incontrati) familiar e dopo 24 ore sosterranno un altro test.
Questa modalità operativa rende le varie prove indipendenti una dall’altra.
Al termine dell’esperimento si valuta la performance dei soggetti, confrontando i tempi di esplorazione, e calcolando un indice di memoria così composto: (TN–TF)/(TN+TF), dove TN è il tempo di esplorazione dello stimolo novel e TF quello dello stimolo familiar.
2.2.c Analisi statistiche
L’analisi statistica dei dati ottenuti dai tests comportamentali, è stata eseguita con il supporto del programma statistico Jandel SigmaStat versione 2.0.
Nell’Open Field, abbiamo valutato con un t test e un Mann-Whitney Rank Sum Test, rispettivamente, l’attività locomotoria, prendendo in considerazione il totale della distanza percorsa, espressa in cm e la velocità media espressa in cm/sec. Nell’Object Recognition
Test, abbiamo effettuato degli studi preliminari inerenti la fase di familiarizzazione, al fine
di verificare l’efficacia e l’attendibilità del protocollo. Inizialmente abbiamo utilizzato un’ANOVA a due vie per confrontare le prestazioni valutando gli indici di memoria del gruppo dei WT contro il gruppo dei c57 ponendo come fattori il genotipo e l’indice.
Poi abbiamo valutato se una delle due locazioni dei cubi (Destra o Sinistra), fosse preferita dagli
animali (WT, RKO e EKO), determinando una differenza dei tempi di esplorazione. Abbiamo valutato i tempi della media della familiarizzazione, in tutti gli intervalli temporali, separatamente tra i tre genotipi. Per fare ciò, ci siamo avvalsi di tre Mann-Whitney Rank Sum Test, uno per genotipo.
Una volta stabilito se fosse presente, o meno, un’esplorazione preferenziale di un oggetto, abbiamo stimato le performances dei 3 genotipi al cambiamento dei patterns nell’insieme degli intervalli temporali (confrontando la media dell’esplorazione dell’oggetto familiar). Abbiamo eseguito un’ANOVA ad una via per il gruppo dei RKO ed un’ANOVA ad una via su ranghi per il gruppo dei WT e degli EKO; il fattore era il pattern (con 6 livelli).
Eliminata la variabile cubo, e la variabile pattern, abbiamo considerato se, complessivamente, ci fosse nei tre genotipi una differenza nei tempi di esplorazione, prendendo come riferimento la fase di
familiarizzazione (confrontando le medie di esplorazione al familiar), per tutti gli intervalli contemporaneamente. Ci siamo serviti del Mann-Whitney Rank Sum Test.
Alla fine di questi tests preliminari, siamo andati ad eseguire il test di memoria utilizzando gli indici scritti sopra. Per analizzare questi, al fine di individuare dei deficit di memoria nelle nostre linee murine mutanti, ci siamo serviti di un’ANOVA a due vie.
Concludendo, ci siamo proposti di analizzare i cambiamenti dei tempi di esplorazione nel passaggio dalla fase di familiarizzazione a quella del test vero e proprio. Nel caso di una differenza degli indici di memoria, eseguire questa analisi, ci permette di individuarne la causa: un decremento del tempo di esplorazione relativo all’oggetto familiare oppure un aumento del tempo di esplorazione dell’oggetto nuovo. Per questo, abbiamo suddiviso i dati tra i tre genotipi e li abbiamo analizzati distintamente, per ogni intervallo temporale, con l’ausilio di ANOVA ad una via per misure ripetute, su ranghi e non.
2.3 Registrazioni elettrofisiologiche
2.3.a Preparazione dei tessuti
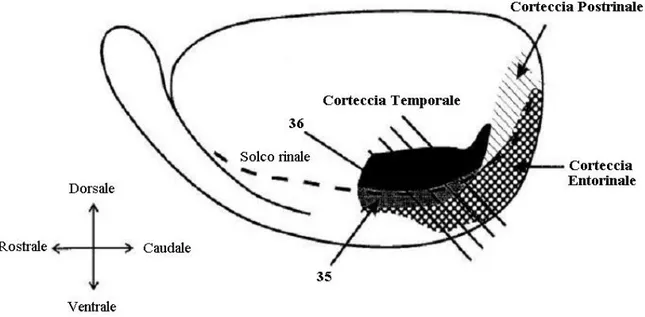
Sono state preparate fettine di encefalo che comprendessero la corteccia peririnale da topi adulti (50–90 giorni). Sono state prese tutte le precauzioni necessarie per minimizzare la sofferenza degli animali e per ridurne al minimo il numero utilizzato. Il prelievo dell’encefalo è stato effettuato in accordo con i protocolli del Ministero della Sanità. Il cervello è stato tolto il più velocemente possibile e i due emisferi sono stati divisi facendo un taglio di sezione medio-sagittale. Mentre un emisfero veniva squadrato per procedere all’affettamento l’altro è stato messo in una soluzione di simile composizione del liquido cerebrospinale e raffreddata con ghiaccio (Soluzione di taglio, ventilata con 95% O2/5% CO2, che comprende (mM): NaCl 132.8 ; KCl 3.1 ; NaHCO3 4 ; HEPES/NaOH 10 ; K2HPO4 1 ; CaCl2 1 ; MgCl2 2 ; d-glucosio 5 ; Acido Ascorbico 1; Myo inositolo 0.5 ; Piruvato 2 ; Acido chinurenico 1). Questa soluzione presentava una concentrazione di Ca2+ inferiore rispetto al liquido cerebro spinale per ridurre l’attività necrotica durante il taglio. Allo stesso fine è stato aggiunto l’acido chinurenico, un bloccante generale dei recettori glutammatergici. Per riequilibrare lo squilibrio ionico dovuto alla riduzione di calcio è stata aumentata la concentrazione del Mg2+. La squadratura consisteva nel togliere la parte rostrale e caudale dell’emisfero in due singoli tagli eseguiti con angolazione approssimativa di 45° rispetto all’asse dorsoventrale. Una volta squadrato l’emisfero, la sua estremità rostrale è stata incollata su una piastra. Sulla stessa piastra è stato incollato un cubo 1x1x1 cm di gel di Agar 1.5%, in modo che una delle sue facce poggiasse sul lato piatto dell’emisfero formatosi nel momento della divisione. La piastra poi, è stata fissata meccanicamente sul fondo della vasca del vibratomo in modo che l’emisfero fosse compreso tra la lama e il cubetto di Agar. Quest’ultimo ha la funzione di impedire che la fette si sgretolino durante il taglio, a causa della debole consistenza dell’emisfero. La vasca era stata precedentemente riempita di soluzione di taglio e raffreddata con ghiaccio.
Figura 2.3 Il disegno mostra la disposizione dell’encefalo per il taglio.
Sono state tagliate fette dello spessore di 310/350 micron, contenenti la corteccia temporale, peririnale ed entorinale. Ogni fetta tagliata è stata rapidamente messa in una soluzione di taglio ventilata e preparata senza acido chinurenico, alla temperatura di 30°. Alla fine dell’affettamento del primo emisfero le stesse procedure sono state ripetute per il secondo. Dopo circa venti minuti, i becker contenenti le fettine sono stati lasciati a temperatura ambiente per un’ora circa.
2.3.b Metodi di registrazione
Una singola fetta veniva messa in una camerina di registrazione in cui fluiva la soluzione di registrazione (32° e 1.5 ml/min) (Soluzione di registrazione, ventilata con 95% O2/5% CO2, che comprende (mM): NaCl 132.8 ; KCl 3.1 ; NaHCO3 4 ; HEPES/NaOH 10 ; K2HPO4 1 ; CaCl2 2 ; MgCl2 1 ; d-glucosio 5 ; Acido Ascorbico 1; Myo inositolo 0.5 ; Piruvato 2 ; Glicina 0.01).
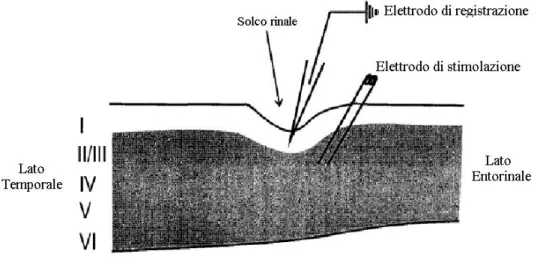
Usando elettrodi di vetro riempiti con soluzione di registrazione, sono state effettuate registrazioni extracellulari di field potentials dallo strato I della corteccia peririnale, in prossimità del solco rinale.
Figura 2.4 La figura mostra come si presentano le strutture encefaliche sulle fette. Le frecce indicano la corteccia peririnale.
L’elettrodo di stimolazione è stato applicato lateralmente rispetto a quello di registrazione ed in corrispondenza degli strati II e III. La distanza tra i due elettrodi era compresa tra 0.16 e 0.36 mm. Una volta ottenuto un segnale, sono stati osservati i valori del potenziale di campo al variare dell’intensità di stimolazione. Con l’aumento dell’intensità di stimolazione si osserva un aumento del potenziale fino ad una valore massimo di saturazione. Se questi valori vengono riportati su un grafico si osserva un andamento sigmoidale.
Il valore massimo e minimo dei field potentials generalmente erano ottenibili con un range di intensità di stimolazione che variava tra 0.1 e 0.8 MilliAmpère. Per ogni segnale il valore del voltaggio da scegliere come valore di base per l’esperimento, era compreso tra metà e ¾ di quello di saturazione. Tutte le registrazioni, sono state effettuate prendendo in considerazione segnali di field potentials la cui latenza di picco superava i 3 ms ed il cui valore di base line superava gli 0.3 MilliVolts. Prima di procedere col protocollo occorreva che il segnale avesse raggiunto la stabilizzazione. Per fare ciò, effettuando uno stimolo ogni 30 secondi, si constatava che l’ampiezza della risposta rimanesse approssimativamente stabile per almeno venti minuti.
Dopo venti minuti di stabilità del valore di base line si è proceduto con uno dei due protocolli:
1) LTD indotta da carbacolo: perfusione per dieci minuti di soluzione di registrazione a cui è stata aggiunta 0.05 mM di carbacolo (Cch). Successivo ritorno alla perfusione normale per 55 minuti, per lavare il Cch ed osservare il nuovo livello della base line. Per tutto l’esperimento si registra sempre effettuando una stimolazione ogni 30 secondi.
2) LTP indotta da stimolazione con ritmo theta: stimolazione con quattro treni, uno ogni 15 secondi dove ogni treno comprende dieci scariche di quattro pulsazioni a 100 Hz, con un intervallo di 200 ms tra ogni scarica. Successivo ritorno ad effettuare uno stimolo ogni 30 secondi, per 60 minuti al fine di osservare il nuovo livello della base line.
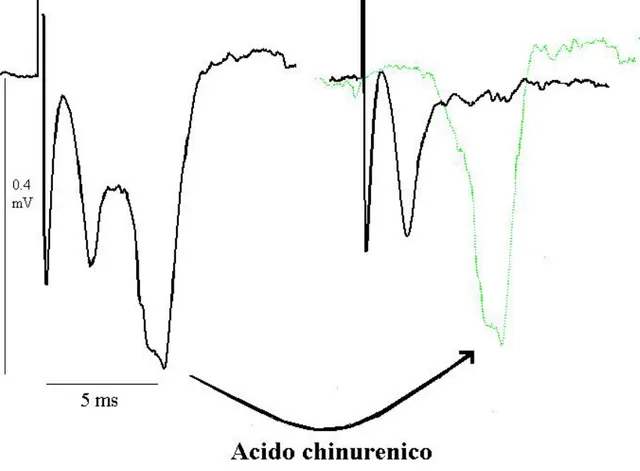
In entrambi i due protocolli, dopo la fase di osservazione della nuova base line, la fettina è stata perfusa per 10 minuti con soluzione di registrazione contenente acido chinurenico 1 mM, per poter quantificare la componente presinaptica del segnale, in modo da poter risalire con approssimazione, alla percentuale di componente post sinaptica dell’onda della base line iniziale (Figura 2.6).
Sono state escluse dai dati:
- le registrazioni in cui l’artefatto elettrico o il grosso della componente presinaptica (latenza < 2.5 ms) subivano un cambiamento di ampiezza maggiore del 20%.
- le registrazioni in cui la stima della componente postsinaptica era minore del 70% dell’onda registrata.
Figura 2.6 La perfusione di soluzione contenente 1 mM di acido chinurenico elimina completamente la componente postsinaptica del segnale (onda verde).
- le registrazioni in cui la nuova base line non raggiungeva un livello vicino al plateau ma manteneva una pendenza maggiore del 10% in valore assoluto.
2.3.c Analisi statistiche
L’analisi statistica dei dati ottenuti dalle registrazioni elettrofisiologiche, è stata eseguita usando il programma statistico Jandel SigmaStat versione 2.0.
In entrambi i due tipi di sperimentazione, sono stati analizzati i valori in percentuale delle nuove base line, rispetto a quelli delle base line iniziali. Sono stati considerati i valori relativi agli ultimi 30 stimoli prima dell’applicazione dell’acido chinurenico. E’ stato utilizzato un test ANOVA a due vie in cui i valori delle percentuali sono legate a due variabili, quella del tempo ( gli ultimi 30 stimoli registrati) e quella del genotipo (RKO, WT, EKO).