1. INTRODUZIONE
1.1
Le piante e gli stress da freddo e da congelamento
1.1.1. Effetti delle basse temperature sulle cellule e sui tessuti vegetali
La temperatura è uno dei più importanti fattori che condizionano la di-stribuzione geografica delle piante - organismi pecilotermi e sessili - e la produttività delle specie agrarie. Le basse temperature rallentano, infatti, le reazioni enzimatiche e modificano la conformazione delle ma-cromolecole, influenzando la maggior parte dei processi biologici e pro-ducendo una complicata rete di effetti indiretti, che rende arduo de-terminare quali siano i processi interessati più seriamente da tali con-dizioni ambientali.
Molte specie vegetali di origine tropicale e subtropicale, ivi compre-se piante coltivate di grande importanza economica come mais, riso, pomodoro, patata, fagiolo, soia, cotone e molte specie da frutto, manife-stano riduzioni nette della velocità di crescita e di sviluppo a tempera-ture comprese fra 0°C e 12°C. Esse, in altri termini, vanno soggette a danni da gelo, che consistono in cambiamenti fisici e fisiologici sinto-matici di una condizione di stress.
Al contrario, le piante di origine temperata (inclusi cereali invernali come il grano tenero, l'orzo e la segale, Brassica spp., Spinacia oleracea e la specie modello Arabidopsis thaliana) hanno evoluto, in maggioran-za, meccanismi che consentono loro non soltanto di continuare a
cresce-re e svilupparsi a basse temperatucresce-re, ma altcresce-resì di sopportacresce-re condi-zioni di congelamento (fino a -25°C circa nelle specie erbacee annuali) successive ad un periodo di gelo. Una più studiate delle risposte biolo-giche alla temperatura è, appunto, la cosiddetta acclimatazione al
freddo, termine con cui si indica il complessivo incremento di
tolleran-za al congelamento acquisito da talune specie in seguito all'esposizione a temperature basse ma superiori al punto di congelamento. Il fenome-no si fa particolarmente affascinante nel caso delle specie arboree pe-renni, che alle latitudini più elevate si sono adattate a fronteggiare temperature inferiori ai -40°C, sviluppando adattamenti particolari come la dormienza (Welling e Palva, 2006).
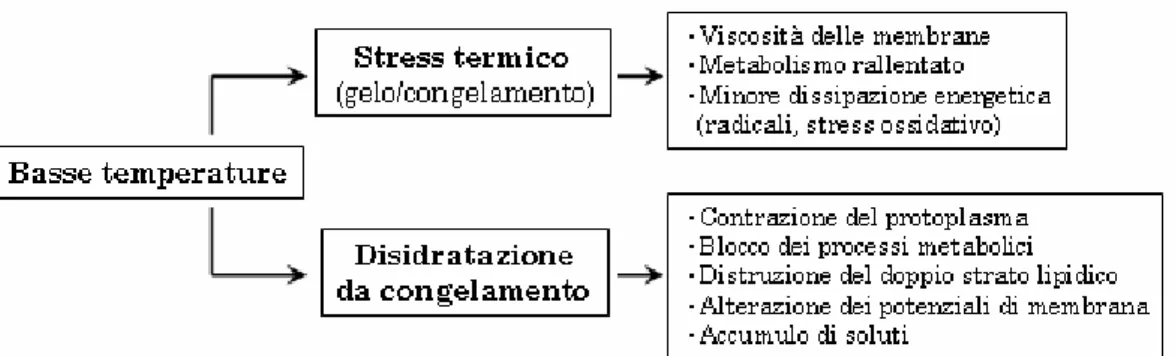
Figura 1.1. Componenti principali dello stress da basse temperature ed effetti al
livello cellulare (adattato da Beck et al., 2004).
La dizione "stress da basse temperature" si presta a indicare collet-tivamente condizioni tanto di gelo, o freddo che dir si voglia, quanto di congelamento, ovvero l'insieme degli stress derivanti sia da temperatu-re non permissive ma superiori allo zero (tipicamente comptemperatu-rese nell'in-tervallo 0-12°C) sia inferiori agli 0°C. Quantunque correlati vicende-volmente ed in parte condivisi, gli effetti sulle piante dell'uno e dell'al-tro regime termico non sono tuttavia coincidenti, infatti le risposte di tolleranza al gelo e quelle di acclimatazione e di tolleranza al
congela-mento costituiscono processi fisiologici distinti, benché in parte sovrap-posti fra loro, ciascuno dei quali richiede l'attivazione da parte della pianta di un insieme di meccanismi specializzati. Con una semplifica-zione estrema, la ragione ultima di questa differenza può essere ricono-sciuta nel fatto che agli effetti termodinamici del freddo si affiancano, nel caso del congelamento, le conseguenze della nucleazione del ghiac-cio nei tessuti (Figura 1.1).
L'abbassamento della temperatura, nelle cellule di organismi non omeotermi come le piante, rallenta i flussi metabolici influenzando le cinetiche enzimatiche, in accordo con l'equazione di Arrhenius:
k = A·e-Ea/RT (essendo k la costante di velocità della reazione, Ea la
sua energia di attivazione, A il fattore preesponenziale, R la costante dei gas e T la temperatura assoluta); in aggiunta, la viscosità dei mezzi cellulari aumenta al calare della temperatura, e con essa cala il tasso di diffusione delle molecole. Appare comprensibile, quindi, che gli adat-tamenti essenziali degli organismi psicrofili e psicrotrofi mirino a ga-rantire l'espressione delle attività enzimatiche a livelli appropriati (Zecchinon et al., 2001).
Il secondo tipo di effetto termodinamico diretto esercitato dall'ab-bassamento della temperatura sui componenti cellulari consiste nel cambiamento di conformazione a cui vanno soggette molecole dalle strutture complesse come i polipeptidi: a temperature basse può diven-tare termodinamicamente favorevole, per certe proteine globulari, e-sporre regioni non polari al solvente acquoso, a scapito della funzionali-tà e della stabilifunzionali-tà della molecola stessa. Anche le membrane subiscono modifiche strutturali, benché solo al di sotto del loro punto di congela-mento, mentre a temperature superiori sono interessate da alterazioni dello stato fisico e della composizione chimica. Da tutto ciò deriva la
necessità di proteggere queste molecole biologiche, attraverso agenti che ne permettano il mantenimento allo stato nativo.
Quando la temperatura scende ulteriormente e oltrepassa il punto di congelamento, il suo effetto sui tessuti viene a realizzarsi attraverso il fenomeno della transizione di fase dei fluidi; le molecole d'acqua, in particolare, possono aggregarsi in cristalli di ghiaccio sia spontanea-mente (nucleazione omogenea), sia grazie alla catalisi di sostanze nu-cleatrici (nucleazione eterogenea). Per quanto riguarda la prima eve-nienza, essa ha scarse probabilità di verificarsi a temperature di poco inferiori allo zero, poiché la massa di acqua pura necessaria per la for-mazione di un singolo cristallo stabile di ghiaccio è estremamente grande anche a -30°C (si può calcolare, ad esempio, che essa corrispon-de a 2.3·10312 kg a -10°C). Pertanto, previa rimozione delle sostanze
nucleatrici, le piante hanno la possibilità di mantenere le proprie cellu-le non congelate in presenza di temperature ben al di sotto dello zero: il fenomeno prende il nome di "supercooling" e resta possibile fino alla temperatura di -38.5°C, alla quale anche un volume molto piccolo di acqua pura va incontro a nucleazione omogenea; tale limite termico in-feriore viene, inoltre, leggermente abbassato dalle molecole di soluto contenute nella matrice cellulare, che contribuiscono per circa 1-2°C.
Questi elementi, nonché l'esistenza nei tessuti di barriere fisiche capaci di ostacolare la crescita dei cristalli di ghiaccio - assai rapida, una volta iniziata - permettono, in via teorica, alla pianta di evitare il congelamento fino all'estremo di -40°C (Pearce, 2001). Essi, tuttavia, sono contrastati dalla presenza, difficilmente eliminabile, di nucleatori eterogenei, che possono trovarsi tanto sulla superficie esterna dei tes-suti quanto al loro interno (nucleatori estrinseci o intrinseci): si tratta di proteine batteriche (secrete dai cosiddetti batteri INA, ice nuclea-tion-active), molecole biologiche prodotte dalla pianta stessa e particelle
organiche ed inorganiche. Vari aspetti della nucleazione eterogenea nelle piante sono ancora ignoti, ma è possibile affermare che questo sia il principale fenomeno responsabile, in ambienti umidi, del congela-mento dei tessuti vegetali a temperature di poco inferiori allo zero (Pe-arce, 2001).
La formazione di ghiaccio nei tessuti ha luogo generalmente al di fuori della membrana plasmatica, poiché i fluidi extracellulari possie-dono un punto di congelamento più alto del citoplasma e ospitano di norma nucleatori eterogenei. Il cambiamento di stato dell'acqua abbas-sa il potenziale idrico extracellulare e provoca, di conseguenza, la per-dita osmotica di acqua dalla cellula e la sua plasmolisi; la quantità to-tale di acqua rimossa dalla cellula dipende sia dal potenziale idrico in-tracellulare che dalla temperatura di congelamento, che influenza di-rettamente il potenziale extracellulare finale.
È riconosciuto, dunque, come la disidratazione cellulare sia il danno primario arrecato ai tessuti dallo stress da congelamento, il che spiega perché esso mostri numerosi tratti in comune con gli stress salini, idrici e osmotici. Benché l'analisi degli effetti dello stress da congelamento sia concentrata prevalentemente sulla cellula, è opportuno ricordare, per completezza, che esso produce altresì conseguenze più macroscopi-che sui tessuti e sugli organi della pianta, quali l'alterazione della struttura in seguito all'incorporazione di grosse masse di ghiaccio, la frattura dei fusti degli alberi dovuta a tensioni interne nel legno, l'u-stione dei tessuti congelati esposti al vento o al sole, l'insorgenza di ma-lattie favorita dalle lesioni create dal congelamento e fenomeni di em-bolismo nei vasi xilematici (Pearce, 2001).
Le funzionalità cellulari possono essere in vario modo compromesse dalla disidratazione indotta da congelamento, ma le ripercussioni prin-cipali si osservano sul plasmalemma, sulle endomembrane e su quelle
organellari. In particolare, il comportamento del primo è stato estensi-vamente studiato su protoplasti di cereali sottoposti al congelamento, e le evidenze raccolte consentono di classificare in tre categorie le modifi-cazioni a carico di questa struttura (Uemura et al., 1995; Xin e Browse, 2000).
Un primo tipo di lesione è detto "expansion-induced lysis" (EIL) e consiste nella perdita irreversibile di superficie di membrana durante la disidratazione, tale per cui pressione idrostatica generata dalla rei-dratazione della cellula conduce alla sua lisi prima del raggiungimento del volume originario. Si nota infatti che, nelle cellule non acclimatate, l'applicazione dello stress provoca l'invaginazione della membrana e il distacco di vescicole endocitotiche, mentre le cellule acclimatate trat-tengono la membrana nella sua intera lunghezza, mediante estrusioni esocitotiche che permettono la nuova distensione cellulare. In protopla-sti di Arabidopsis, il fenomeno ha luogo nell'intervallo termico compre-so fra -2°C e -4°C.
A temperature inferiori, la disidratazione spinta porta in stretto contatto reciproco membrane cellulari diverse, da cui può derivare la loro separazione laterale di fase: il doppio strato viene distrutto dal-l'aggregazione di alcuni lipidi di membrana in una struttura invertita a simmetria esagonale (indicata come fase HexII), che si traduce in una perdita irreversibile di impermeabilità e funzionalità (indicata come LOR, "loss of osmotic responsiveness") dopo la fine dello stress. La mag-gior predisposizione a incorrere in tale comportamento è esibita dalla membrana plasmatica e cloroplastica.
Entrambi i tipi precedenti di modificazione si verificano solo rara-mente in piante acclimatate, nelle quali il danno da congelamento è as-sociato piuttosto con le "fracture-jump lesions". Anch'esse sono dovute a
fusioni localizzate di membrane diverse, ma i dettagli biofisici del fe-nomeno non sono ben noti.
In specie suscettibili al congelamento, come Vigna radiata e Sain-tpaulia grotei, è stata osservata la formazione di ghiaccio anche all'in-terno della cellula, evento riconosciuto come letale. L'ipotesi è che, in questo caso, il congelamento produca sulla membrana danni tali da impedirle di funzionare come barriera contro la propagazione del ghiaccio extracellulare e che, inoltre, la stessa parete cellulare sia in-capace di assolvere a questo compito, permettendo la formazione di ghiaccio nello spazio periprotoplasmico (Yamada et al., 2002).
Le medesime specie suscettibili allo stress possono essere sfruttate per lo studio degli effetti che il freddo produce al livello dell'intera cel-lula, con lo scopo di identificare, per converso, quali siano i meccanismi di protezione messi in atto dalle specie capaci di acclimatarsi.
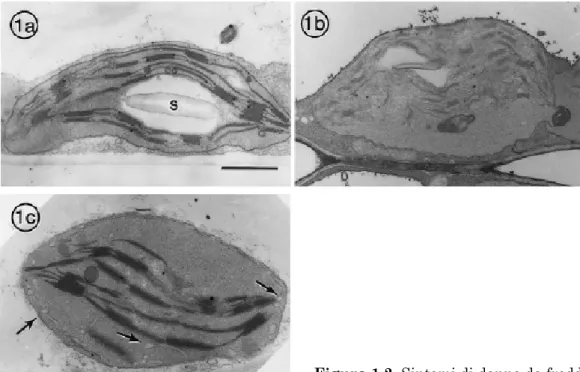
Dal punto di vista ultrastrutturale, i sintomi dello stress da freddo appaiono ampiamente condivisi fra le specie sensibili. Una manifesta-zione precoce e pressoché universale di stress è il rigonfiamento dei clo-roplasti: gli organelli e i tilacoidi si ingrossano, le dimensioni dei gra-nuli di amido si riducono e la membrana cloroplastica interna forma un reticolo periferico (Figura 1.2). È probabile che, durante lo stress, l'e-sportazione dei prodotti di degradazione dell'amido verso citoplasma sia inibita (dalla bassa forza elettromotrice, oppure dalla riduzione di capacità trasportatrice del floema) e che essi agiscano da soluti osmoti-camente attivi; il reticolo periferico stesso potrebbe venir prodotto per sopperire alle funzioni di trasporto compromesse. Il protrarsi dello stress causa l'accumulo di gocce lipidiche nel cloroplasto, l'imbruni-mento dello stroma, il disassemblaggio dei grana e, infine, anche delle membrane esterne.
Al contrario dei plastidi, i mitocondri non paiono andar soggetti ad alcuna alterazione ultrastrutturale, se non in specie ipersensibili al freddo come il cotone, la V. radiata, la saintpaulia e l'efedra, nelle quali sono stati osservati il rigonfiamento, la destrutturazione e la vacuola-rizzazione di questi organuli. Probabilmente ciò è almeno in parte do-vuto al contenimento dello stress ossidativo, che nei mitocondri vegeta-li è assicurato dalla presenza di diversi sistemi di disaccoppiamento, mediati da proteine PUMP (plant uncoupling mitochondrial proteins) quali l'ossidasi alternativa AOX, la proteina di stress CSP310 (per cold shock protein) e altri fattori disaccoppianti UCP-like della membrana mitocondriale interna, che risultano attivate, fra i vari stress, anche in risposta a quelli da basse temperature (Fiorani et al., 2005; Kolesni-chenko et al., 2005).
Figura 1.2. Sintomi di danno da freddo
in cloroplasti. 1a, cloroplasto normale di spinacio, da piante cresciute in condizioni permissive; s, amido. 1b, cloroplasti di cocomero, da piante trattate per 9h a 5°C e 1000 µmol fotoni m-2 s-1: si osservino il rigonfiamento, la distorsione dei tilacoidi e la
riduzione dei granuli di amido. 1c, cloroplasto di cotone, da piante trattate per 72h al buio, a 5°C e col 70% di umidità; le frecce indicano il reticolo periferico (da Kratsch e Wise, 2000).
La struttura cellulare che viene maggiormente alterata dal freddo, tanto in specie suscettibili quanto in specie tolleranti, resta comunque la membrana plasmatica. Numerosi studi hanno dimostrato chiara-mente che la fluidità della membrana diminuisce al calare della tempe-ratura, pur mantenendosi in fase fluida cristallina finché l'abbassa-mento termico non si faccia così pronunciato da causare le lesioni da congelamento esaminate in precedenza. È interessante, in quest'ottica, il fatto che l'imposizione di condizioni iperosmotiche (esposizione a PEG e a glucosio) produca su membrane artificiali un effetto di irrigi-dimento analogo a quello conseguente al freddo, benché i dati raccolti in tal senso richiedano ancora conferme e supplementi d'indagine (Los e Murata, 2004).
La dipendenza fra il grado di insaturazione degli acidi grassi nei fo-sfolipidi di membrana e la sua fluidità è un fenomeno assai ben carat-terizzato, anche dal punto di vista genetico (Browse e Xin, 2001): l'atti-vazione di geni codificanti per desaturasi in risposta al freddo è stata, infatti, osservata nella maggior parte degli organismi pecilotermi stu-diati, dai batteri alle piante e ai pesci. L'evento primario del danno da gelo nelle specie vegetali sensibili è ritenuto la transizione dalla fase fluida cristallina a quella di gel nella membrana, che le specie tolleran-ti sono, genericamente, in grado di prevenire attraverso il controllo de-gli isomeri insaturi e dede-gli steroli presenti all'interno della struttura. Danni secondari derivano, poi, dall'incremento delle attività di perossi-dazione lipidica associato alla componente ossidativa degli stress da basse temperature.
Da ultimo, pur senza aver la pretesa di approfondire, in questa se-de, il vasto capitolo dell'interrelazione fra il freddo e il metabolismo primario della cellula, non si può prescindere dal tratteggiare alcuni aspetti salienti del coinvolgimento delle basse temperature nella
foto-sintesi e nel metabolismo del carbonio, alla luce dei quali diventa più agevole interpretare la miriade di sfaccettature di cui si compone la ri-sposta della pianta.
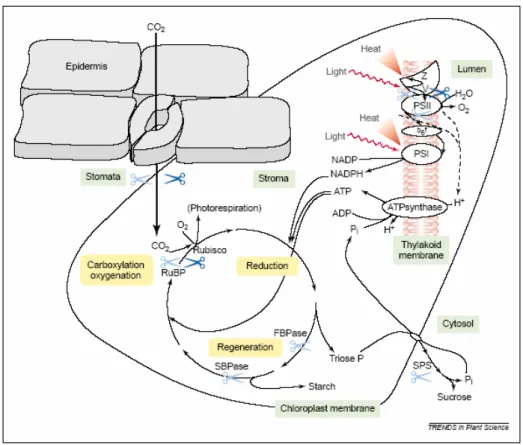
Numerosi studi dimostrano che nelle specie sensibili il freddo com-promette la funzionalità di tutti i principali componenti biochimici del-la fotosintesi, compresi del-la catena di trasporto elettronica e il ciclo di Calvin, e altera la regolazione della conduttanza stomatica (Allen e Ort, 2001).
Figura 1.3. Effetti primari del freddo sulla fotosintesi in piante termofile. V,
viola-xantina; Z, zeaxantina. Le forbici azzurre indicano i punti d'impatto delle basse tem-perature in condizioni luminose, le forbici blu quelli in condizioni di buio o bassa lu-minosità (da Allen e Ort, 2001).
Quando le condizioni di freddo sono imposte in assenza di luce, le conseguenze sulla successiva attività fotosintetica derivano
principal-mente dall'interferenza col metabolismo dei carboidrati, dall'inibizione dell'attività della rubisco (riconosciuta come un bersaglio primario du-rante lo stress da freddo) e della chiusura stomatica, dei quali l'inibi-zione del fotosistema I sembrerebbe essere un riflesso (Figura 1.3). Il declino della sintesi di saccarosio e l'accumulo di intermedi fosforilati della fotosintesi costituiscono, infatti, eventi pressoché istantanei in seguito all'applicazione dello stress, che risultano in un'immediata li-mitazione dell'attività dell'ATP sintasi tilacoidale; fra le concause di ta-li eventi potrebbero trovarsi la distruzione del ritmo circadiano di atti-vità della saccarosio fosfato sintasi e l'inibizione della mobilizzazione notturna dell'amido fogliare.
Allorché, invece, lo stress termico è concomitante con l'esposizione alla luce piena, i danni maggiori consistono nella fotoinibizione e nella distruzione del controllo redox delle bisfosfatasi del ciclo di Calvin. An-che le conseguenze ultrastrutturali sui cloroplasti delle specie sensibili, infatti, si aggravano decisamente al crescere dell'irradianza, poiché l'imposizione di condizioni di luce, piuttosto che di buio, modifica in maniera sostanziale la situazione di stress a cui la pianta è sottoposta, aggiungendo a quello da freddo anche lo stress foto-ossidativo.
Il calo della temperatura abbassa i tassi di reazione, limitando la dimensione dei sink dell'energia luminosa assorbita: in tale frangente, il fotosistema II può subire il fotodanneggiamento, con conseguenze non rapidamente reversibili sul trasporto elettronico tilacoidale, inoltre è possibile che i componenti danneggiati del centro di reazione vengano sostituiti con maggiore difficoltà, a causa della fluidità ridotta delle membrane. Al di là delle compensazioni dinamiche nell'efficienza quantica del fotosistema, l'eccesso di energia luminosa viene almeno in parte distribuito in sink alternativi come gli antiossidanti - che inter-vengono anche nella disattivazione delle ROS prodotte - e dissipato
sot-to forma di calore. Tuttavia, si ritiene che le maggiori conseguenze del-lo stress ricadano sugli enzimi assimilatori piuttosto che sul PSII: il cambiamento dello stato redox del cloroplasto sembrerebbe, cioè, com-promettere drammaticamente la regolazione di enzimi come la sedoep-tulosio 1,7-bisfosfatasi (SBPasi) e la fruttosio 1,6-bisfosfatasi (FBPasi), nonché la rubisco attivasi (ipotizzando l'esclusione di danni diretti alla rubisco).
Per quanto concerne la regolazione stomatica, infine, resta proble-matico stabilire se la chiusura degli stomi osservata a seguito del fred-do dipenda da una diretta inibizione della fotosintesi a causa dello stress oppure sia, viceversa, un effetto diretto della temperatura che porta con sé una riduzione di ci e il declino della fotosintesi.
1.1.2. La risposta agli stress da basse temperature
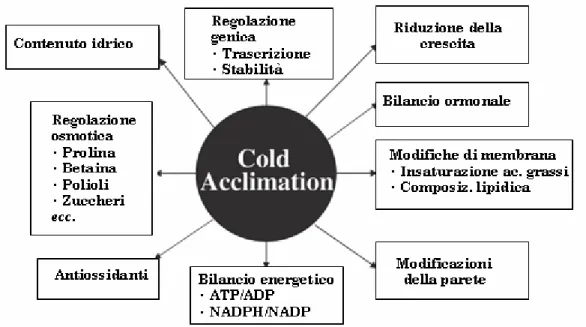
L'acclimatazione va considerata come il processo adattativo, innescato dal freddo, nel quale la pianta predispone le difese contro gli effetti del-lo stress da congelamento susseguente, cioè acquisisce nei suoi confron-ti caratterisconfron-tiche di tolleranza valide a proteggerla nel momento in cui la fuga dallo stress (che consiste nell'impedire la formazione del ghiac-cio nei tessuti) non sia più possibile. Data la sua importanza agronomi-ca, il fenomeno ha richiamato immediatamente l'attenzione degli stu-diosi, che nel tempo ne hanno individuato gli aspetti fenotipici e strut-turali, dapprima, e biochimici, in un secondo momento: decenni di in-dagini e osservazioni hanno permesso, cioè, di stilare un catalogo assai esteso dei cambiamenti fisiologici e biochimici associati alla risposta di acclimatazione.
Essa, dunque, comprende cambiamenti del metabolismo tanto pri-mario quanto secondario, la modifica del bilancio ormonale e del tasso di crescita dell'organismo, l'adattamento funzionale e strutturale delle membrane, la sintesi di soluti compatibili, di enzimi e molecole antios-sidanti, nonché di una quantità di altri prodotti, non ancora caratteriz-zati, dall'azione protettiva rispetto al congelamento, e l'alterazione del contenuto idrico dei tessuti in vista della disidratazione da congela-mento (Figura 1.4). È evidente, in altri termini, che l'acclimatazione è un carattere complesso che incide profondamente sul metabolismo del-la pianta (queldel-la che alcuni Autori hanno ribattezzato "sindrome da freddo" delle piante), qualificandosi come un vero e proprio processo di sviluppo. L'osservazione che la tolleranza al congelamento sia inducibi-le piuttosto che costitutiva è un primo segno della portata dei costi me-tabolici che l'insieme dei meccanismi protettivi comporta: altrettanto importante rispetto all'acclimatazione è, infatti, per la sopravvivenza della pianta, il fenomeno inverso di de-acclimatazione, che si innesca rapidamente al ritorno di condizioni permissive.
Non sono, per inciso, mancati studi genetici che hanno tentato di le-gare al carattere multigenico della tolleranza al congelamento QTL e geni maggiori, nell'ottica del miglioramento genetico delle specie agra-rie, in special modo del grano tenero e dell'orzo. I loci maggiori respon-sabili della resistenza sia vegetativa che riproduttiva al congelamento sono stati mappati sul cromosoma 5H dell'orzo e sugli omeologhi, sin-tenici, 5A, 5B e 5D del frumento; ad essi sono strettamente associati i loci Vrn che controllano la vernalizzazione in questi cereali (Francia et al., 2004; Reinheimer et al., 2004).
L'onnicomprensività della risposta di acclimatazione renda talora disagevole separare gli eventi legati all'acquisizione di tolleranza al congelamento da quelli di mera risposta al freddo. Un aiuto, durante
questo sforzo di distinzione, proviene dagli elementi che rendono non del tutto sovrapponibili fra loro gli effetti prodotti dal freddo e dal con-gelamento sulle cellule (Figura 1.1). In altri termini, in condizioni di gelo saranno certamente associate all'acclimatazione almeno le difese riconducibili alla componente di disidratazione, che nello stress da freddo è assente.
Figura 1.4. Principali processi interessati dall'acclimatazione (adattato da Xin e
Bro-wse, 2000).
Con i loro studi sui geni COR, Guy e colleghi presentarono, nel 1985, le prime prove sperimentali del verificarsi di cambiamenti dell'e-spressione genica durante l'acclimatazione, aprendo la strada alla di-mostrazione dell'ipotesi - avanzata da Weiser e colleghi fin dal 1970 - che l'acquisizione di tolleranza al congelamento avesse una base gene-tica. Dal momento della scoperta dei geni COR ad oggi, l'analisi mole-colare della risposta al freddo e al congelamento è stata di gran lunga più incentrata sugli aspetti genomici e trascrizionali, piuttosto che me-tabolici e proteomici del problema. In particolare, i recenti progressi
nella tecnologia dei microarray, applicati alla biologia delle basse tem-perature, hanno reso possibile l'identificazione di numerosi geni che contribuiscono alla risposta finale allo stress, la scoperta di nuove vie regolative e la dissezione di quelle già note, nonché l'elucidazione dei rapporti fra le risposte al freddo e altre vie di segnalazione.
Secondo una classificazione comune a tutti gli studi di risposta agli stress ambientali, i prodotti scoperti e identificati vengono ripartiti in due categorie, l'una comprensiva delle molecole funzionali, responsabili cioè degli adattamenti finali allo stress, l'altra delle molecole associate alla trasduzione del segnale e all'espressione genica (quali protein chi-nasi e fosfatasi, fattori di trascrizione, trasportatori di membrana ed enzimi del metabolismo dei fosfolipidi). L'analisi molecolare è giunta a dimostrare non soltanto come tali molecole siano organizzate in una re-te di vie cellulari di segnalazione e risposta, ma anche come taluni trat-ti della reazione al freddo si sovrappongano e si intreccino con partrat-ti della risposta cellulare a stress ambientali diversi, principalmente di natura osmotica. Svariati studi sono stati, infatti, dedicati alla rico-struzione e alla comparazione dei profili trascrizionali relativi agli stress da siccità, salinità e basse temperature, e all'applicazione di ABA esogeno (Seki et al., 2003 e 2002; Shinozaki et al., 2003). All'origi-ne dell'interazioAll'origi-ne fra questi stress si può riconoscere, in virtù di quan-to esposquan-to precedemente, l'esigenza, al livello cellulare, di rispondere all'elemento comune rappresentato dalla disidratazione.
Per quanto riguarda i soli prodotti funzionali implicati nella difesa dallo stress da congelamento, fra di essi sono inclusi enzimi del meta-bolismo primario (alcol deidrogenasi, saccarosio fosfato sintasi, β-amilasi, fenilalanina-ammonio liasi) e secondario (calcone sintasi), en-zimi legati all'adattamento della composizione delle membrane (desa-turasi FAD8, enzimi biosintetici degli steroli) e della parete, proteine
connesse al metabolismo specifico degli acidi nucleici alle basse tempe-rature (l'RNA elicasi DEAD box LOS4, cfr. Gong et al., 2005; proteine di legame all'RNA di tipo GR-RBP, cfr. Kim et al., 2005; il regolatore trascrizionale FRY2, cfr. Xiong et al., 2002b) e alla sintesi proteica spe-cifica (LOS1, fattore di allungamento della traduzione, cfr. Guo et al., 2002), enzimi della biosintesi di soluti compatibili (∆-pirrolina-5-carbossilato sintasi, galattinolo sintasi), enzimi detossificanti e scaven-ger dello stress ossidativo, trasportatori di membrana, proteine inte-grali coinvolte nel mantenimento del potenziale elettrochimico (come RCI2A, cfr. Medina et al., 2005), β-glucosidasi (come SFR2, cfr. Thorlby et al., 2004), componenti dell'apparato fotosintetico, chaperoni moleco-lari e deidrine.
Quello che segue è un approfondimento sulle classi di composti, fra queste appena elencate, che producono i tratti più riconoscibili della ri-sposta di acclimatazione, al di là dei fenomeni, già analizzati, che inte-ressano le membrane cellulari.
Proteine COR, osmoliti compatibili e chaperoni molecolari
Numerosi studi riportano la sintesi di soluti compatibili (metaboliti di basso peso molecolare che stabilizzano le molecole organiche durante gli stress osmotici e contribuiscono al mantenimento del turgore senza interferire con i processi metabolici) in risposta alle basse temperature, con un probabile effetto protettivo di tipo sinergico; è stata, infatti, os-servata la partecipazione di aminoacidi, composti dell'ammonio qua-ternario e polioli, considerate le classi principali di osmoliti compatibili.
La glicin betaina sembra contribuire in parte all'acquisizione di tol-leranza al congelamento in Arabidopsis (Xing e Rajashekar, 2001), no-nostante essa non sia inclusa fra le specie accumulatrici di tale compo-sto e, per contro, molte specie sensibili allo stress producano betaine.
Ancor più controverso è il ruolo individuale della prolina nella difesa dagli stress da disidratazione: essa è assente in alcuni mutanti costitu-tivamente tolleranti al congelamento, ma forti indizi della sua impor-tanza derivano dai mutanti esk1 (vedi sotto) e dalla sua sintesi in linee CBF-costitutive di Arabidopsis (Cook et al., 2004), che ben si correla con l'induzione del gene P5CS precedentemente osservata. Il coinvol-gimento di diversi altri soluti compatibili è, inoltre, testimoniato dagli esperimenti di ricostruzione del metaboloma da stress (Browse e Lan-ge, 2004; Kaplan et al., 2004).
Fra le sostanze che contribuiscono al mantenimento, alle basse temperature, della corretta conformazione delle molecole sensibili sono inclusi anche gli chaperoni molecolari, che partecipano al processo di ripiegamento delle proteine. Da questo punto di vista, sia le HSP70 e HSP90 che le sHSP sono state correlate alla tolleranza al freddo in va-rie specie, benché non sia noto se esse si comportino, in vivo, come cha-peroni, come stabilizzatrici delle membrane oppure in ambedue i modi (Lopez-Matas et al., 2004).
Come si accennava in precedenza, a partire dal 1985 ad oggi si sono individuati numerosi geni fortemente indotti dal gelo - contrassegnati cioè da livelli di espressione da 50 a 100 volte maggiori in condizioni di freddo rispetto che a 20-25°C - cui sono stati di volta in volta attribuiti i nomi di COR (cold-regulated), LTI (low temperature-induced), KIN (cold-induced), RD (responsive to dehydration) ed ERD (early responsi-ve to dehydration). Fra quelli maggiormente espressi si trovano geni come COR6.6, COR15a, COR47, COR78/RD29A, RD29B e KIN1-2, che sono stati, perciò, utilizzati come marcatori della risposta al freddo in molti studi molecolari. Geni omologhi sono stati isolati e clonati in una varietà di specie vegetali, ma solo alcuni di essi hanno trovato un'attri-buzione funzionale.
Nel tentativo di determinarne l'importanza relativa nell'acquisizio-ne di tolleranza, molti di essi sono stati sovraespressi in Arabidopsis, ma, attualmente, soltanto il gene COR15a è stato dimostrato contri-buire in una certa misura alla tolleranza al congelamento delle piante transgeniche non acclimatate (Shimamura et al., 2006; Artus et al., 1996). Tuttavia, la trascurabilità dell' effetto protettivo di COR15a al livello della pianta intera e, parimenti, l'inefficacia degli altri COR con-fermano che l'utilità di tali prodotti risiede nella loro azione collettiva piuttosto che individuale. Il polipeptide codificato da COR15a (il primo dei COR a ricevere una caratterizzazione) è una piccola proteina stro-matica (9.4 kDa di massa) che, una volta persa la sequenza segnale du-rante l'ingresso nel cloroplasto, è designata come COR15am; il suo ruo-lo, apparentemente incongruo, di protezione della membrana plasmati-ca dalla LOR viene spiegato con l'ipotesi che, delle due membrane coin-volte, quella cloroplastica possieda la maggior propensità a promuovere la transizione alla fase HexII, e che quindi COR15am protegga l'intero sistema preservandone il componente più debole. Dal punto di vista funzionale, il polipeptide altera la curvatura intrinseca della membra-na intermembra-na del cloroplasto, con cui interagisce mediante diverse α-eliche anfipatiche (Steponkus et al., 1998).
I prodotti di diversi geni COR (come COR6.6, COR47, ERD10 ed ERD 14 in Arabidopsis, WCOR140 in T. aestivum e CuCOR19 in Citrus unshiu, per fare soltanto alcuni esempi) sono stati identificati come deidrine, proteine appartenenti alla vasta famiglia LEA D11 (late em-briogenesis abundant), la cui sintesi è stata osservata in un'ampia gamma di organismi in risposta a stress accomunati da una componen-te di disidratazione: nelle piancomponen-te, esse si accumulano, infatti, negli stress da basse temperature, siccità e salinità, ed in seguito all'applizione di ABA (Campbell e Close, 1997; Close, 1997). Alla medesima
ca-tegoria appartengono, oltre ai COR, numerosi geni denominati DHN/RAB/LEA.
Le deidrine vegetali sono identificate dalla presenza di un dominio aminoacidico conservato, indicato con la lettera K perché ricco in Lys, corrispondente a un'α-elica putativa, ripetuto fino a undici volte lungo la sequenza. Esse contengono, inoltre, altri domini (S, Y e Φ) meno conservati, che si trovano ripetuti e arrangiati fra loro in numerosi mo-di mo-diversi, il che dà luogo ad una notevole varietà mo-di conformazioni (ef-ficacemente descritta dall'apposita notazione YSK). Le strutture se-condarie predominanti sembrano essere gli avvolgimenti casuali mati dalle regioni Φ. La composizione aminoacidica risultante è for-temente idrofilica.
Nonostante gli sforzi compiuti, l'esatta funzione delle deidrine in condizioni di disidratazione resta elusiva, comunque si ritiene che i numerosi membri della famiglia ricoprano ruoli almeno in parte distin-ti (come indicato dalla grande varietà dei profili di espressione genica). L'attività principale delle deidrine è considerata la protezione delle membrane cellulari e degli enzimi sensibili al congelamento come la ca-talasi, la lattato deidrogenasi e le α-amilasi, che può essere ottenuta at-traverso più meccanismi: interazioni idrofobiche delle α-eliche anfifili-che con le molecole target, formazione di strati coesivi da parte degli avvolgimenti casuali, inibizione della perossidazione lipidica e attività di scavenging (Hara et al., 2004, 2003 e 2001). Anche per queste pro-teine valgono le medesime considerazioni sugli effetti sinergici fatte a proposito dei COR (Puhakainen et al., 2004).
Zuccheri
Il ruolo centrale del metabolismo primario nell'acquisizione di tolleran-za al congelamento emerge dai recenti studi che mostrano la serie di
eventi che, durante l'acclimatazione, impediscono l'inibizione della sac-carosio fosfato sintasi (SPS) e della fotosintesi dovute alla bassa tempe-ratura (Stitt e Hurry, 2002). Nelle foglie di piante di Arabidopsis cre-sciute a temperature permissive e trasferite a 4°C si osserva, nell'arco di 30 minuti, l'attivazione post-traduzionale dell'SPS, con un conse-guente aumento dell'affinità per il substrato nell'enzima, e, nei pochi giorni successivi, l'induzione dei geni cFBP (FBPasi citosolica) e SPS; in aggiunta a questo, sembra che si verifichi un parziale trasferimento di Pi dalle riserve vacuolari al citosol, tale da permettere l'incremento
della fosforilazione dei substrati senza il consumo del Pi libero.
La piena acclimatazione, tuttavia, si compie soltanto nelle foglie svi-luppatesi al freddo, nelle quali all'aumento selettivo di attività degli enzimi coinvolti nella sintesi del saccarosio si affianca un analogo effet-to su tutti gli enzimi del ciclo dell'assimilazione foeffet-tosintetica del carbo-nio. Il risultato dipende sia dall'induzione dei geni codificanti che dal contenuto proteico superiore delle foglie cresciute al freddo (minor con-tenuto idrico, maggior volume relativo del citoplasma rispetto al vacuo-lo, maggior concentrazione di proteine in rapporto al peso fresco). Lo studio dei mutanti pho1 e pho2, alterati nella [Pi] del germoglio, ha
ri-velato che alcuni di questi elementi della risposta sono innescati dalle gravi limitazioni di Pi subite dalla pianta durante la prima fase dello
stress.
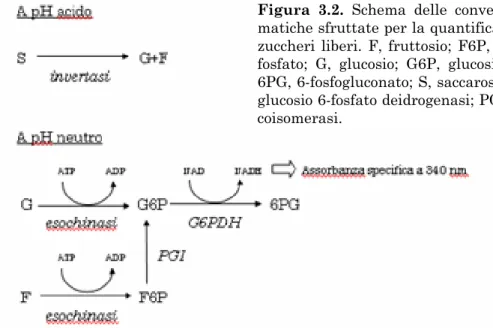
Alcuni studi molecolari hanno, recentemente, messo in luce le con-nessioni fra il metabolismo dell'amido e lo sviluppo della risposta di ac-climatazione, già emersi dalle indagini ultrastrutturali e biochimiche (Figura 1.5). Il gene codificante per un'isoforma cloroplastica della β-amilasi, BMY8, risponde positivamente alle basse temperature e la sua espressione (probabilmente preceduta, nel breve periodo, dall'attiva-zione post-trascrizionale dei trascritti preesistenti) è correlata con la
produzione da stress di maltosio. BMY7 e BMY8 sembrano le uniche β-amilasi attivate da stress termici (Kaplan e Guy, 2005 e 2004). Inoltre, Yano et al. (2005) hanno riportato l'intervento del gene GWD/SEX1 (starch excess1), codificante per una α-glucano/acqua dichinasi (una classe di enzimi regolatori globali del catabolismo dell'amido), nell'ac-cumulo di zuccheri liberi durante la prima fase dello stress, ossia entro le prime 24 h di esposizione al freddo. SEX1 è attivato dal freddo, tut-tavia non sembra promuovere di per sé la risposta di degradazione del-l'amido, bensì piuttosto rispondere omeostaticamente a tale evento, golato da componenti situati a valle, inoltre l'effetto di sex1 risulta re-versibile nell'arco di una settimana.
Figura 1.5. Modello per l'idrolisi
del-l'amido durante lo stress da freddo. R1, GWD/SEX; AMY, α-amilasi; DBE, enzima deramificante; PHS, fosforilasi b; BMY, β-amilasi; DPE1-2, enzimi disproporzionanti plastidi-co e citosoliplastidi-co; MEX, esportatore del maltosio; GLT, trasportatore del glu-cosio (da Kaplan e Guy, 2005).
L'accumulo di zuccheri solubili durante l'acclimatazione, in condi-zioni di luce, e la loro correlazione positiva con la tolleranza acquisita al congelamento sono fatti ben documentati in Arabidopsis (Wanner e Junttila, 1999) e molte altre specie, e confermati, dal punto di vista ge-netico, dal comportamento di mutanti di Arabidopsis come sfr4 ed esk1: l'incapacità del primo di acclimatarsi e prevenire i danni da LOR alla membrana plasmatica è causata dal mancato accumulo di zuccheri so-lubili in risposta al freddo (Uemura et al., 2003), mentre il secondo, al
contrario costitutivamente tollerante al congelamento, produce elevati livelli di zuccheri e di prolina anche a temperature permissive (Xin e Browse, 1998). Il ruolo degli zuccheri liberi viene giustificato ipotiz-zando diverse funzioni, a partire da quelle certe di soluti compatibili e di sostanze crio- e osmoprotettrici per arrivare ad altre più indirette di regolatori delle vie di risposta al freddo (Stitt e Hurry, 2002).
Glucosio e fruttosio assolvono, dunque, a funzioni di protezione, ma appaiono molto meno efficaci di disaccaridi come il saccarosio e il mal-tosio. Quest'ultimo (Figura 1.5) sembra agire da soluto compatibile nel-la difesa delle proteine stromatiche e delle membrane tinel-lacoidali, ma è possibile che il suo ruolo sia più complesso, poiché esso contribuisce al-la produzione del glucosio e del saccarosio citosolici (Kapal-lan e Guy, 2005).
In aggiunta ai carboidrati di basso peso molecolare, altri composti contribuiscono a determinare la tolleranza allo stress. I fruttani sono i primi polisaccaridi per i quali è stata provata la capacità di interagire direttamente con membrane lipidiche e di stabilizzarle durante il con-gelamento, forse ascrivibile alla loro struttura lineare (Hincha et al., 2000). Anche per la famiglia degli oligosaccaridi derivati dal raffinosio è stato dimostrato un ruolo nella protezione delle membrane dalla dis-seccamento, in qualità di molecole in grado di sostituire l'acqua nei gu-sci di idratazione, e, in aggiunta, la via del raffinosio risulta fortemente indotta a valle dei CBF, tuttavia una funzione crioprotettiva degli RFO non è stata, finora, dimostrata (Zuther et al., 2004).
Fra gli zuccheri alcol, trealosio e mannitolo sono i principali collega-ti all'osmoprotezione e allo scavenging dello stress ossidacollega-tivo. Il primo (Garg et al., 2002) si accumula in grandi quantità soltanto in alcune specie tolleranti al disseccamento (come le "resurrection plants"),
men-tre nelle almen-tre si trova a livelli molto bassi, forse a causa dell'attività delle trealasi.
Proteine anti-congelamento
Le AFP (antifreeze proteins) costituiscono una particolarissima classe di proteine, rinvenute in organismi filogeneticamente distanti come batteri, funghi, pesci artici, insetti svernanti e piante acclimatate, che non risultano accomunate da alcuna similarità strutturale bensì solo dalla loro omologia funzionale. Nelle piante, queste proteine vengono rilasciate nell'apoplasto, sulla superficie esterna della parete cellulare, dove agiscono abbassando la temperatura di congelamento dei fluidi in maniera non colligativa. A differenza di tutti gli altri soluti, le AFP non vengono escluse dai cristalli di ghiaccio in crescita, bensì incorporate nella loro matrice grazie ai siti funzionali che permettono loro di legar-si alla superficie del ghiaccio; la loro presenza all'interno previene la ri-cristallizzazione del ghiaccio e limita, probabilmente, i danni sulla cel-lula al momento del disgelo (Griffith e Yaish, 2004). È stato riportato che, oltre a inibire la cristallizzazione del ghiaccio, le AFP aumentano la stabilità delle membrane artificiali in modo specifico (Tomczak et al., 2001).
Sorprendentemente, le AFP isolate dalle piante esibiscono similari-tà di sequenza con proteine PR, e come queste sono condizionate nello svolgimento della loro attività dal calcio. Si ritiene, infatti, che sia i ci-cli di congelamento/scongelamento che la presenza di cationi incidano sulla struttura secondaria di queste proteine e sui loro processi di oli-gomerizzazione, indispensabili dal punto di vista funzionale (Stres-sman et al., 2004).
1.2
Regolazione trascrizionale dei geni indotti da
bas-se temperature
1.2.1. Reti di regolazione trascrizionale
Lo studio dei geni coinvolti nella risposta finale agli stress che, come il congelamento, condividono una componente di disidratazione cellulare ha messo in risalto il ruolo dell'acido abscissico e, con esso, l'esistenza di almeno quattro sistemi indipendenti per la regolazione dell'espres-sione genica sotto l'insieme di tali stimoli ambientali.
Ad orientare per prime in questa direzione sono state le analisi, di fondamentale importanza, svolte sull'espressione nei mutanti aba (A-BA-deficient) e abi (ABA-insensitive) dei geni RD29A/LTI78/COR78 e COR15a, entrambi inducibili sia dal freddo che dagli stress idrici: la lo-ro attivazione da stress in quel backglo-round genetico plo-rova, infatti, l'e-sistenza di vie ABA-indipendenti nella trasduzione del segnale. Tutta-via essi risultano parimenti indotti dall'ABA esogeno in condizioni normali di crescita, pertanto sembrerebbero appartenere anche a vie ABA-dipendenti autonome rispetto alle prime. La giustificazione dei ri-svolti controversi che l'acido abscissico conserva nella segnalazione de-gli stimoli da bassa temperatura risiede, forse, nell'incapacità da parte di questi ultimi - a differenza di quelli legati alla disidratazione - di promuovere l'accumulo del fitoregolatore endogeno a livelli sufficien-temente alti da innescare la trascrizione di geni a valle come RD29B (Shinozaki e Yamaguchi-Shinozaki, 2000). Ma la scoperta recente di geni addizionali nella sottofamiglia CBF, e in particolare di CBF4, im-plicati nella regolazione ABA-dipendente sembrerebbe dischiudere pro-spettive nuove sull'esistenza di vie ulteriori per l'espressione dei geni di risposta al freddo (Yang et al., 2004).
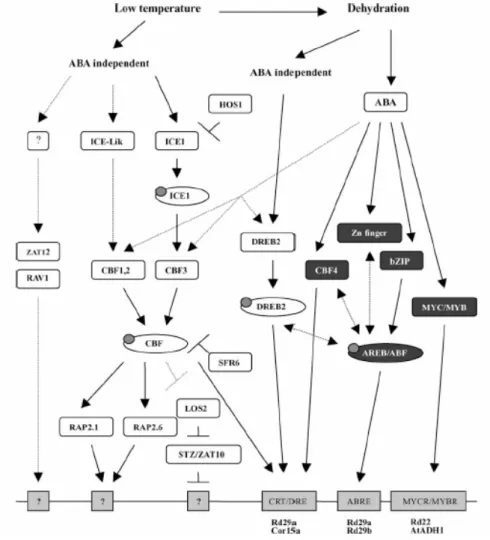
Il quadro dei sistemi di controllo dell'espressione genica, arricchitosi nel tempo con l'identificazione di elementi cis-acting e fattori trascri-zionali trans-acting multipli, associati alla trascrizione dei geni regolati dal freddo, continua, dunque, tuttora ad espandersi. Lo stato attuale delle conoscenze al riguardo, complessivamente schematizzato nella Figura 1.6, viene discusso nel dettaglio all'interno di questo Paragrafo.
Figura 1.6. Modello della rete regolativa della risposta alle basse temperature e alla
disidratazione. I fattori trascrizionali dipendenti dall'ABA sono ombreggiati, quelli indipendenti sono in chiaro. I cerchietti simboleggiano modifiche post-trascrizionali o post-traduzionali. Nei rettangoli in basso, i siti di legame dei fattori di trascrizione e, sotto, alcuni promotori rappresentativi che li contengono. Le doppie frecce indicano un ipotetico cross-talk e le linee discontinue una possibile regolazione (da Yang et al., 2005).
1.2.2. La via CBF
La scoperta della via CBF di risposta al freddo, che rappresenta a tut-t'oggi il sistema genetico meglio caratterizzato nell'ambito dell'acclima-tazione, è stata resa possibile dall'isolamento e dall'analisi dei geni COR, per la precisione dal riconoscimento, mediante mutazio-ne/delezione dei promotori, delle regioni cis-acting responsabili della loro sensibilità al freddo e alla disidratazione. Tali regioni, presenti in una o più copie all'interno di molti dei promotori esaminati, sono state denominate DRE/CRT (dehydration-responsive element/C-repeat) e ABRE (ABA-responsive element).
Il primo dei due elementi contiene il motivo conservato A/GCCGAC, inizialmente dimostrato essere essenziale per l'attivazione ABA-indipendente del gene RD29A a seguito di sollecitazioni da freddo e di-sidratazione. Successivamente, è stato confermato che una singola co-pia della sequenza DRE/CRT è sufficiente per l'espressione dei geni che la possiedono, in risposta a stress dalla trasduzione indipendente dal-l'ABA, qual è considerato il freddo secondo il paradigma corrente (Ya-maguchi-Shinozaki e Shinozaki, 2005). È possibile che anche la strut-tura delle regioni circostanti l'elemento cis-acting sia importante per garantire l'interazione con i fattori trascrizionali, dal momento che non tutti i geni a valle di DREB1A, identificati da Maruyama et al. (2004), risultano essere target diretti del fattore di trascrizione corrispondente, pur contenendo il DRE/CRT.
Il secondo elemento cis-acting, ABRE, contiene la sequenza conser-vata C/GACGTGGC, affine alla G-box ed è il principale regolatore della trascrizione genica ABA-dipendente. Per l'avvio della trascrizione non è sufficiente un singolo elemento ABRE, diversamente dal caso
prece-dente, bensì è richiesta la presenza di almeno due copie in tandem del-la sequenza o di un elemento di accoppiamento addizionale.
Il passo successivo è stato la scoperta dei fattori di trascrizione DREB1/CBF (DRE/CRT-binding factor) che riconoscono l'elemento DRE/CRT (Stockinger et al., 1997). Ben presto i geni CBF1/DREB1B, CBF2/DREB1C e CBF3/DREB1A, presenti in tandem sul cromosoma 4 di Arabidopsis, hanno conquistato il ruolo di snodi principali ("master switches") nella regolazione della risposta al freddo. I loro trascritti, in-fatti, si accumulano nell'arco di 15 minuti dall'applicazione delle basse temperature (sia di gelo che di congelamento) e circa due ore dopo l'e-sposizione al freddo raggiungono un picco di espressione transiente, che coincide con l'attivazione dei geni target e appare legato, in am-piezza, al gradiente termico imposto. Inoltre, la sovraespressione dei geni CBF1 e CBF3 in Arabidopsis si traduce nell'espressione costituti-va dell'intera batteria di geni COR e in un incremento della tolleranza al congelamento, nelle piante transgeniche, in assenza dello stimolo da freddo. Geni omologhi ai CBF sono stati identificati in varie specie er-bacee diverse da Arabidopsis e crescenti evidenze suggeriscono che questa via di risposta sia conservata nelle piante superiori (Yang et al., 2005).
Le proteine DREB1/CBF, codificate dai geni CBF, fanno parte della superfamiglia AP2/ERF, di cui costituiscono una sottofamiglia a parte, contraddistinta dalla presenza di due sequenze aminoacidiche fian-cheggianti il dominio di legame AP2/EREBP, come questo conservate nelle proteine ortologhe. Si tratta di attivatori della trascrizione, dei cui meccanismi funzionali ancora poco è noto: l'ipotesi più recente è che esse agiscano almeno in parte richiamando sui promotori dei geni target complessi di modificazione della cromatina; in questo senso,
pro-teine come SFR6 (vedi sotto), ADA2 e GCN5 potrebbero influenzare la funzionalità dei CBF (Van Buskirk e Thomashow, 2006).
I maggiori sforzi profusi finora sono stati, invece, concentrati sull'i-dentificazione dei geni controllati dai CBF, ossia sulla ricostruzione del regulone CBF (ove con "regulone" si intende l'unità trascrizionale con-trollata da un unico fattore di trascrizione, attraverso presenza del re-lativo elemento cis-acting nei promotori dei geni a valle), cui sono con-siderati appartenenti tutti i geni nettamente espressi in risposta sia al-la bassa temperatura che alal-la sovraespressione ectopica dei CBF. Il profilo trascrittomico che emerge da svariati esperimenti di microarray assegna al regulone CBF geni codificanti per un'ampia gamma di cate-gorie proteiche, che comprendono fattori trascrizionali, componenti del-le vie di trasduzione del segnadel-le, enzimi biosintetici, crioprotettori e al-tre proteine di sal-tress, e numerosi geni dalla funzione sconosciuta (Vo-gel et al., 2005; Fowler e Thomashow, 2002; e altri): l'insieme dei geni a valle include, pertanto, tanto elementi coinvolti nell'ulteriore espres-sione genica e nella regolazione della trasduzione del segnale quanto componenti responsabili della risposta finale allo stress. Su tale insie-me, i diversi membri della famiglia CBF non sembrano mostrare speci-ficità di target, poiché non sussistono differenze significative nei profili di espressione a valle dei tre geni, che, del resto, esibiscono una simila-rità molto alta nelle sequenze nucleotidiche (Van Buskirk e Thoma-show, 2006).
L'appartenenza al regulone di molti geni codificanti per fattori di trascrizione, acclarati o putativi, diversi dai CBF può essere indizio di un meccanismo regolatorio complesso, realizzato per mezzo di sotto-reguloni interni al sistema CBF. Due fattori di trascrizione controllati dai CBF e ritenuti membri di sotto-reguloni sono, per esempio, RAP2.1
e RAP2.6, i cui geni risultano sovraregolati dall'espressione costitutiva dei CBF1-3 (Fowler e Thomashow, 2002).
Quest'organizzazione potrebbe spiegare aspetti altrimenti sorpren-denti, fra cui l'assenza degli elementi DRE/CRT nei promotori del 20% circa dei geni del regulone (Vogel et al., 2005) e il fatto che non tutti i circa 100 geni attribuiti siano indotti, bensì ve ne sia un certo numero di repressi, malgrado le proteine CBF funzionino solamente come atti-vatori trascrizionali. In effetti la sovraespressione ectopica dei geni DREB/CBF reca con sé effetti pleiotropici che appaiono legati non sol-tanto agli elevati costi metabolici associati al mantenimento costitutivo dei meccanismi di acclimatazione, ma anche alla reale repressione di geni attivi nei processi di sviluppo e crescita: ad esempio, il gene STZ/ZAT10, bersaglio di DREB1, codifica per un fattore di trascrizio-ne di tipo C2H2-ZF (zinc-finger) che è stato associato alla soppressione
di molti geni appartenenti alle vie fotosintetiche e del metabolismo dei carboidrati (Nakashima e Yamaguchi-Shinozaki, 2006).
Un altro aspetto del funzionamento della via CBF che ha ricevuto considerevoli attenzioni è la regolazione dei geni DREB1/CBF mede-simi in risposta alle basse temperature. La tempestività dell'espressio-ne di CBF1, CBF2 e CBF3 in risposta al trattamento da freddo fa sup-porre che alcuni componenti della via di trasduzione del segnale e della via regolativa della trascrizione siano già presenti a temperature nor-mali e che vadano soggetti ad attivazione trascrizionale o post-traduzionale in conseguenza del segnale termico. Il problema è stato affrontato da diversi punti di vista, sia attraverso lo studio dei promo-tori dei geni CBF che attraverso l'impiego della genetica inversa.
ICE1 (inducer of CBF expression 1), la cui esistenza era stata ipotiz-zata da Gilmour e colleghi, è stato, dunque, identificato mediante il clonaggio posizionale della mutazione ice1, che influenza sia
l'espres-sione del transgene promotore DREB1A::LUC che la tolleranza al fred-do e al congelamento delle piante di Arabifred-dopsis (Chinnusamy et al., 2003): in questo tipo di screening genetico, linee transgeniche recanti il costrutto di fusione promotore-reporter vengono mutagenizzate e, al-l'interno della popolazione M2 risultante, vengono selezionati gli indi-vidui caratterizzati da una risposta bioluminescente anomala sotto stress da freddo, sui quali viene compiuta l' analisi genetica. Nel caso di ice1, siamo in presenza di una mutazione dominante negativa che sopprime quasi totalmente l'accumulo dei trascritti CBF, pertanto il gene in questione è considerato un regolatore a monte dei CBF.
ICE1 codifica per un fattore di trascrizione di tipo MYC-bHLH (basic helix-loop-helix), la cui funzione di attivatore trascrizionale è ri-volta solo verso DREB1A/CBF3 ma non verso gli altri due geni, il che pare sintomatico di differenze nei meccanismi di espressione dei mem-bri della famiglia genica. L'analisi molecolare dei promotori DREB1 ha permesso di identificare elementi cis-acting fondamentali per la rispo-sta ad ICE1 (ICEr1 e ICEr2); ICE sembra riconoscere sequenze senso multiple localizzate all'interno all'interno di due dei sei box con-servati che si trovano nei promotori dei geni CBF (Zarka et al., 2003). Il gene è espresso costitutivamente, accumulandosi nel nucleo anche a temperatura normale, e potrebbe, quindi, esser soggetto ad attivazione da freddo, in accordo con l'ipotesi di partenza: in particolare, è stato proposto il meccanismo di fosforilazione/defosforilazione della proteina (o, indirettamente, di un cofattore).
In definitiva, l'induzione da stress dell'intera via CBF, principale responsabile della tolleranza al congelamento, è un chiaro indizio dei costi metabolici che la pianta deve sostenere per difendersi dal freddo e costituisce una spiegazione indiretta del perché i sistemi di acclimata-zione non possano essere costitutivamente attivi. Benché ICE1 sia
l'u-nico fattore trascrizionale di cui è stata dimostrata direttamente l'inte-razione con i promotori CBF, un ruolo regolativo nella via CBF è stato inferito anche per vari altri geni e proteine, come ZAT12, SFR6, FRY2 e alcuni geni HOS, LOS e COS, individuati per mezzo di uno screening genetico con mutanti promotore RD29A::LUC avviato da Ishitani e colleghi.
Da un'indagine su mutanti di Arabidopsis generati mediante muta-genesi chimica, Warren et al. hanno identificato sette geni di Arabidop-sis che influenzano il processo di acclimatazione, denominati SFR1-7 (sensitive to freezing): individui mutanti per cinque di essi (precisamen-te i mutanti sfr1, 2, 4, 5-1, 5-2 e 6) non mostrano alcun'al(precisamen-terazione nel-la risposta al gelo, ma acquisiscono una tolleranza al congenel-lamento molto più debole del wild type a seguito dell'acclimatazione (Thoma-show, 2001). Fra questi geni SFR6 è stato, in seguito, caratterizzato come un regolatore del flusso attraverso la via CBF: poiché l'espressio-ne dei geni marcatori COR si riduce marcatamente l'espressio-nei mutanti sfr6 trattati con basse temperature, mentre tanto i trascritti di altri geni inducibili da freddo ma privi degli elementi DRE/CRT quanto i geni CBF vengono espressi a livelli normali, la proteina SFR6 sembra esse-re attiva in un punto della via di segnalazione situato fra la trascrizio-ne dei CBF e l'induziotrascrizio-ne dei geni a valle. Studi estensivi hanno con-fermato l'azione post-trascrizionale di SFR6 rispetto ai geni DREB1/CBF e individuato un analogo effetto su DREB2, giacché i mu-tanti sfr6 si rivelano sensibili anche a stress osmotici (Boyce et al., 2003). L'esatto funzionamento della proteina nel wild type non è stato, però, ancora chiarito.
Come accennato poco sopra, un'indagine genetica di diverso tipo è stata condotta con l'ausilio della bioluminescenza, mediante mutagene-si inserzionale su linee di Arabidopmutagene-sis trasformate col costrutto
promo-tore RD29A::LUC (con un sistema identico a quello descritto a proposi-to dell'identificazione di ICE1). Diverse linee mutate sono state isolate sulla base dell'espressione anomala del gene reporter in risposta a stress osmotici, e in ragione di ciò battezzate come hos, los e cos (rispet-tivamente per high, low e constitutive expression of osmotically respon-sive genes); in seguito, vari loci interessati da queste mutazioni sono stati associati a geni, clonati e collegati con successo alle vie di segna-lazione e ai sistemi di trascrizione del freddo.
HOS1, ad esempio, è una proteina RING-finger che interviene nella degradazione dei fattori CBF e ICE1 nel corso dello stress da basse temperature, come suggeriscono la sua localizzazione subcellulare (la proteina è citoplasmatica, ma viene trasportata nel nucleo dopo l'espo-sizione al freddo), la sua omologia con le proteine animali IAP che fun-zionano da E3 ubiquitina ligasi e l'accresciuta trascrizione dei geni di risposta al freddo nei mutanti hos1, durante gli stress da basse tempe-rature (Dong et al., 2006; Lee et al., 2001).
I mutanti fiery derivano il loro nome dall'insolita intensità della luminescenza emessa dalle linee transgeniche mutate in condizioni di stress. Il gene corrispondente al locus FRY2 è stato clonato e risulta i-dentico in sequenza a CPL1 (CTD-phosphatase-like1). FRY2/CPL1 si comporta da repressore della via CBF, operando da inibitore zionale a monte dei DREB1 oppure interagendo col complesso trascri-zionale CBF. Benché i dettagli funzionali non siano noti, sulla proteina è stata rilevata la presenza di un dominio caratteristico delle fosfatasi dell'RNA polimerasi II CTD (RNPII CTD); la capacità di FRY2 di lega-re dsRNA suggerisce, inoltlega-re, la possibilità che nelle piante gli RNA siano in grado di modulare la risposta agli stress come nelle cellule a-nimali (Xiong et al., 2002b).
Nel quadro della regolazione termica del sistema CBF, infine, DREB1C/CBF2 costituisce una particolarità, dato che l'assenza degli elementi DRE/CRT nei promotori CBF porterebbe a escludere che i tre geni possano essere sottoposti ad autoregolazione. Malgrado ciò, il comportamento dei mutanti cbf2, isolati da Novillo et al. (2003) nella prima indagine loss-of-function applicata ai geni CBF, induce a ipotiz-zare per il gene in questione un ruolo di regolazione negativa nei con-fronti degli altri due. Nel modello esplicativo proposto dagli Autori, il livello stazionario di proteina CBF2 in condizioni controllo basterebbe a reprimere l'espressione di CBF1 e CBF3, mentre la medesima proteina non sarebbe più sufficiente allo scopo in seguito alla rapida attivazione termica dei due geni; CBF1 e CBF3 potrebbero contribuire, in un se-condo momento, ad attivare CBF2, innalzando i livelli della proteina e causando la nuova inibizione dei propri geni.
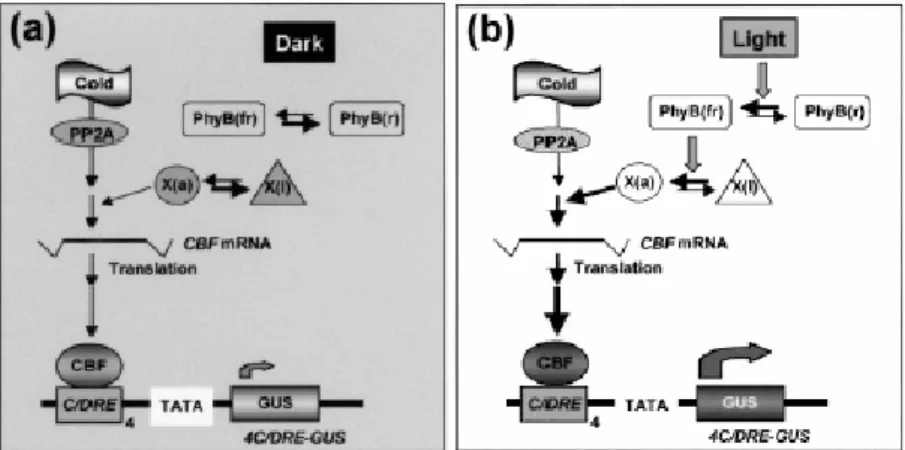
La complessità della regolazione della via CBF emerge con chiarezza dal fatto che la temperatura non rappresenta l'unico segnale in grado di attivarla, poiché altri segnali, fra i quali spicca la luce, possono af-fiancarvisi. La luce non produce soltanto una stimolo sull'espressione di CBF1-3 e di geni a valle come COR15a, ma è altresì necessaria per l'espressione dei geni di risposta mediata dagli elementi DRE/CRT: in-fatti, piante di Arabidopsis trasformate col costrutto 4C/DRE::GUS (reporter preceduto da quattro copie dell'elemento cis-acting isolato dal gene) esprimono il gene reporter soltanto a un livello basale se trattate con freddo o siccità al buio, a fronte di una forte induzione in condizioni di luce (Kim et al., 2002). La fotoreversibilità del comportamento del costrutto fa supporre l'intervento dei fitocromi (in particolare del fito-cromo B giacché l'espressione del reporter è minima nei mutanti con background genetico phyB e phyAphyB), mentre sembra da escludere il coinvolgimento della fotosintesi. Il cross-talk fra la via CBF e quella dei
fitocromi dovrebbe aver luogo in un punto a monte dei CBF e a valle della fosfatasi 2A (PP2A), come da Figura 1.7.
In aggiunta, l'espressione da freddo dei geni CBF1-3 è connessa con l'orologio circadiano, come anche quella di RAV1 e ZAT12 (Fowler et al., 2005). Piante cresciute con un fotoperiodo di 12 h esibiscono una dinamica giornaliera di accumulazione del trascritto di CBF3 a 24°C, con un massimo a ZT4 e un minimo a ZT16 (Zeitgeber time, tempo dal-l'alba soggettiva), mentre i dati relativi all'espressione ciclica di geni target marcatori (COR) restano controversi. Benché sia intuitivo asso-ciare le mutazioni nella durata del giorno con l'acclimatazione, per lo meno in riferimento a piante poliennali, l'esatto significato biologico e il contesto molecolare di queste evidenze sperimentali restano ancora da chiarire.
Figura 1.7. Modello per l'attivazione luminosa dei geni CBF. Le frecce spesse e sottili
indicano, rispettivamente, un livello attivato o basale nella segnalazione (da Kim et
al., 2002).
1.2.3. Nuove vie regolative nella risposta al freddo
Un certo numero di evidenze sperimentali indica che altre vie di segna-lazione, accanto a quella prominente del sistema DREB/CBF, mediano
la risposta della pianta alle basse temperature. Per esempio, non tutti i geni indotti dal freddo contengono gli elementi DRE/CRT, né il regulo-ne CBF non comprende la totalità dei geni attivati dallo stress; inoltre, taluni geni espressi costitutivamente hanno effetti importanti nei pro-cessi di risposta ma non richiedono l'intervento dei CBF. Attualmente, tuttavia, la conoscenza sulle nuove vie regolative si trova ad uno stadio iniziale e appare assai frammentaria se confrontata col quadro costrui-to atcostrui-torno al regulone CBF.
Xin e Browse (1998), mediante uno screening analogo a quello di Warren a colleghi, hanno isolato il già menzionato mutante eskimo1 (esk1) di Arabidopsis, che, esibendo un forte incremento nella tolleran-za costitutiva al congelamento pur sentolleran-za mostrare un'espressione co-stitutiva dei geni marcatori COR, sembra trarre la propria tolleranza allo stress dall'attivazione di vie diverse da quella CBF.
Le prime prove dirette della presenza di vie indipendenti da questa derivano dall'esame del profilo di espressione temporale in piante so-vraesprimenti CBF1-3 trasferite al gelo (Fowler e Thomashow, 2002). A favore dell'ipotesi depongono la presenza di geni dal profilo trascri-zionale simile a quello dei CBF (induzione precoce in risposta allo stress) ma non influenzati nelle piante trasformate, l'organizzazione in blocchi, durante il time-course, dei geni indotti e la constatazione che il regulone CBF rappresenta un sottoinsieme del totale dei geni attivati dal freddo. Fra quelli precocemente indotti, sono stati riconosciuti i ge-ni ZAT12 (zinc Arabidopsis transporter) e RAV1 (related to ABI3/VP1), codificanti per due fattori di trascrizione noti: il primo è una proteina ZF, caratterizzata per la prima volta da Meissner e Michael nel 1997, il secondo un fattore AP2-B3L caratterizzato da Kagaya e colleghi nel 1999.
Si è osservato che la sovraespressione di ZAT12 causa un leggero ma riproducibile incremento nella tolleranza al congelamento, indican-do un ruolo attivo per il regulone ZAT12. Esso sembra interagire con il CBF in almeno due modi (Vogel et al., 2005), in primo luogo condivi-dendo con esso alcuni geni regolati dal freddo e in secondo esercitando su di esso una regolazione negativa, diretta o meno (dal momento che una conseguenza dell'espressione costitutiva di ZAT12 è la soppressio-ne dei geni CBF1-3 in risposta al freddo). Tuttavia, è assai probabile che il ruolo svolto da ZAT12 sia assai più complesso di quanto finora accertato, giacché il gene, di cui è stato dimostrato il coinvolgimento in una varietà di stress biotici e abiotici oltre alle basse temperature (in-fezioni batteriche, fungine e da nematodi; stress da ferita; fumigazioni di ozono; stress da metalli pesanti; applicazione di metanolo, fumonisi-ne, elicitori e UV; shock termico; siccità; stress ossidativo da H2O2;
stress luminoso), sembra avere a che fare con l'integrazione di segnali ambientali multipli. Un'ipotesi plausibile è che esso occupi una posi-zione centrale nella segnalaposi-zione cellulare degli stress mediata da spe-cie attive dell'ossigeno e che la sua espressione, durante i vari stress, sia controllata dalla combinazione di ROS e fitoregolatori (Rizhski et al., 2004; Davletova et al., 2005).
Di recente, due dei fenotipi hos sono stati ricondotti alla modifica-zione di due geni codificanti per fattori di trascrimodifica-zione, situati proba-bilmente a monte di reguloni indipendenti da CBF. HOS9 (Zhu et al., 2004) e HOS10 (Zhu et al., 2005) sono costitutivamente espressi e codi-ficano, rispettivamente, per una proteina HD (homeodomain) putativa con similarità di sequenza rispetto alle proteine WUSCHEL (WUS) e PRESSED FLOWER (PRS) di Arabidopsis, e per un fattore trascrizio-nale putativo di tipo R2R3-MYB. L'insieme dei loro geni target presen-ta una sovrapposizione solo parziale col regulone CBF.
Entrambe le mutazioni hos9-1 e hos10-1 producono conseguenze nette sulla risposta al freddo e al congelamento. I mutanti hos9 esibi-scono una marcata riduzione nella tolleranza al congelamento, dell'or-dine di 3-6°C, e un comportamento analogo, ancorché meno accentuato, nella capacità di acclimatazione, pertanto HOS9 sembra essere asso-ciato alla tolleranza costitutiva al congelamento oppure avere una fun-zione ridondante nell'acclimatafun-zione; l'accumulo dei trascritti di CBF1-3 non è alterato fra hos9 e wt, mentre quello di RD29A aumenta nei mutanti in risposta al freddo ma non a NaCl e ad ABA. Nel complesso, si ipotizza che HOS9, oltre che regolare negativamente alcuni geni a valle dei CBF, possa controllare l'espressione costitutiva di geni essen-ziali per lo sviluppo della risposta, e che quindi la perdita della sua at-tività nei mutanti risulti in un incremento compensatorio di altri geni di risposta al freddo.
Quanto a HOS10, i mutanti corrispondenti dimostrano un compor-tamento analogo a hos9 rispetto all'espressione da freddo di CBF1-3 e RD29A, ma, al contrario di quelli, perdono del tutto la capacità di ac-climatarsi e acquisiscono sensibilità anche allo stress salino. Il fattore di trascrizione HOS10 viene, infatti, correlato positivamente con la sin-tesi di ABA durante la disidratazione.
Né per HOS10 né per gli altri fattori trascrizionali di nuova identi-ficazione passati in rassegna sono stati finora trovati gli elementi cis di legame al DNA.
Anche uno dei mutanti los, precisamente los2, sensibile al freddo, è stato collegato a una via di trascrizione indipendente (Lee et al., 2002). La mutazione reprime l'espressione dei geni marcatori a valle del CBF durante lo stress da congelamento, ma non in corrispondenza di trat-tamenti da ABA, NaCl o PEG, né risulta alterare la trascrizione dei geni CBF; essa, inoltre, conferisce ai mutanti un fenotipo sensibile al
freddo, ma solo se concomitante con condizioni di piena luce. I dati rac-colti dagli Autori suggeriscono che la proteina LOS2, un'enolasi bi-funzionale, possa agire, nel compartimento nucleare, come repressore del gene STZ/ZAT10, nello stesso modo in cui alcune enolasi umane reprimono i geni contenenti motivi c-myc nei promotori (l'equivalente, nel caso delle piante, è la ripetizione a tandem di due sequenze ATG adeguatamente spaziate). Poiché STZ/ZAT10 reprime, a sua volta, i geni COR marcatori, la funzione di LOS2 nei loro confronti risulta di regolatore positivo; il sito di riconoscimento di LOS2 è stato, infatti, rinvenuto anche nei loro promotori.
Il fatto che l'enolasi LOS2 intervenga anche nella via glicolitica ci-tosolica suggerisce un nuovo modo di regolazione e apre interessanti prospettive sulla correlazione fra lo stress da freddo e il metabolismo primario. Si ipotizza che il fenotipo letale condizionale della mutazione los2 sia il risultato di un processo apoptotico innescato congiuntamente dalla regolazione genica difettiva e dai segnali di freddo e luminosità.
Il gene GIGANTEA (GI), da ultimo, codifica per una proteina nucle-are priva di omologia con le sequenze note, di cui non è tutt'oggi cono-sciuta l'esatta funzione. Sono stati, però, ottenuti mutanti gi che esibi-scono fenotipi pleiotropici con alterazioni nella risposta fotoperiodica di fioritura, nella via di segnalazione da fitocromo B, nel funzionamento dell'orologio circadiano e nel metabolismo dei carboidrati, in più la mu-tazione riduce la tolleranza costitutiva al congelamento di Arabidopsis e ne compromette la capacità di acclimatazione. Il gene GI risponde po-sitivamente alle basse temperature in maniera indipendente dai CBF (espressione di CBF1-3 e dei geni COR marcatori inalterata in gi ri-spetto a wt) e successivamente ad essi, mentre non sembra rispondere agli stress idrici e osmotici (Cao et al., 2005). Questi dati, insieme con la localizzazione subcellulare della proteina GI, fanno supporre che
es-sa sia un fattore trascrizionale che regola indipendentemente la rispo-sta al freddo, forse agendo sull'integrazione dei segnali termici e lumi-nosi nella sincronizzazione dell'orologio circadiano.
1.2.4. Cenni sulla regolazione della risposta agli stress osmoti-ci, salini e da disidratazione
Mentre gli stimoli da basse temperature si traducono in una regolazio-ne genica largamente ABA-indipendente, la trascrizioregolazio-ne dei geni di ri-sposta agli stress osmotici e idrici può dipendere o meno dalla sintesi di acido abscissico.
Dei quattro sistemi regolativi ipotizzati inizialmente a partire dagli studi genetici, almeno due dipendono dall'acido abscissico (Figura 1.6). Il primo di questi si indirizza sugli elementi cis ABRE/G-box ed è stato studiato principalmente mediante il gene RD29B, indotto in risposta agli stress salini e idrici ma non termici, che contiene due sequenze ABRE, entrambe essenziali per la sua trascrizione, nel proprio promo-tore. Esse vengono riconosciute da fattori di trascrizione di tipo bZIP (basic-domain leucine zipper) denominati AREB/ABF (ABRE-binding proteins/factors), la cui attivazione dipende da un segnale mediato dal-l'ABA - probabilmente una fosforilazione ABA-dipendente - poiché la loro attività risulta ridotta nei mutanti aba2 e abi1 e accresciuta nei mutanti era1. È interessante che un membro di questa famiglia protei-ca, AREB1/ABF2, sia un componente essenziale della segnalazione del glucosio e che la sua sovraespressione conferisca una maggior tolleran-za non solo alla siccità, ma anche a stress ossidativi e da calore (Yamaguchi-Shinozaki e Shinozaki, 2005).
Dato che la sequenza degli elementi di accoppiamento di ABRE, se presenti, risulta talora simile a quella dei CRT/DRE, non è improbabile che AREB e DREB possano interagire nell'espressione genica ABA-dipendente (Zhang et al., 2004). A monte delle proteine AREB è stato proposto l'intervento di vari peptidi e fattori trascrizionali, fra i quali si trova anche DREB1D/CBF4, un membro della famiglia CBF identifica-to molidentifica-to recentemente e non implicaidentifica-to nella risposta al freddo.
Alcuni geni inducibili da stress con la mediazione dell'ABA conten-gono, invece, sequenze diverse dalle ABRE nei loro promotori e fanno parte degli altri sistemi di regolazione della risposta ABA-dipendente. RD22, per esempio, presenta siti MYBR e MYCR, riconosciuti dai fat-tori trascrizionali MYB2 e MYC2 (una proteina di tipo MYC-like bHLH) che attivano cooperativamente il gene; il secondo di questi fun-ziona, inoltre, nel cross-talk fra ABA a acido jasmonico. Fattori di tra-scrizione addizionali, per i quali è stato proposto un ruolo nella risposta all'ABA, appartengono, poi, alle classi ZF-HD e NAC (nascent polypep-tide-associated complex), i cui siti di riconoscimento si trovano sui pro-motori dei geni come ERD1.
La regolazione ABA-indipendente della risposta al segnale esterno, infine, è ancora una volta imperniata sul sistema DREB, per la preci-sione sui fattori della sottofamiglia DREB2. Si tratta delle proteine DREB2A e DREB2B, che determinano l'induzione di quei geni appar-tenenti al regulone CBF - poiché provvisti di elementi DRE/CRT - che sono stimolati soltanto dalla disidratazione e dalla salinità, oppure an-che dalle basse temperature (come RD29A). I DREB2 sono presenti nella cellula in condizioni normali e vengono attivati da modificazioni post-traduzionali all'insorgere degli stress osmotici, come sembra con-fermare il fatto che, contrariamente al caso dei CBF, la