Capitolo 1
Il sistema visivo
In questo capitolo vengono illustrate l’anatomia e la fisiologia dell’apparato visivo. Sono quindi descritti l’occhio, il suo apparato motorio e le vie neurali che portano il segnale visivo all’elaborazione corticale.
1.1 Cenni sull’apparato visivo
L’apparato visivo è costituito da organi pari e simmetrici, situati, in gran parte, nella regione anteriore della testa, al di sotto della fronte ed ai lati della radice del naso. Di questi organi il principale è senza dubbio l’occhio o bulbo oculare; gli altri (organi
accessori dell’occhio) si possono suddividere in apparato motore (conferisce all’occhio
la motilità) ed apparato protettore (difende il bulbo da eventuali agenti dannosi).
L’apparato motore permette l’orientazione dell’occhio verso determinati settori dell’ambiente, rendendo quest’ultimo un fondamentale mezzo di esplorazione dello spazio circostante.
La sua particolare organizzazione e le sue connessioni con l’encefalo fanno sì che l’occhio possa trasmettere ai centri analizzatori un complesso di segnali che riportano le immagini del mondo esterno.
1.2 Il bulbo oculare
L’occhio può essere considerato come un’appendice dell’encefalo: con questo, infatti, resta perennemente connesso. Il bulbo oculare è provvisto di un peduncolo nervoso, il
nervo ottico, che lo pone in continuità con formazioni del diencefalo, attraverso le quali
gli stimoli visivi vengono trasmessi ai centri nevrassiali per l’attività integrata delle immagini e per le attività riflesse.
posteriore (costituito dalla sclera) ha un raggio di curvatura di 12 mm e forma i restanti cinque sesti posteriori.
Il diametro anteroposteriore del bulbo oculare, asse anatomico, è la linea che unisce il centro della cornea (polo anteriore) al centro della sclera (polo posteriore) e misura circa 24 mm. È detto asse ottico quel tratto di asse anatomico compreso tra il centro della cornea e quello della retina. L’asse visivo o linea della visione (“line of sight”) si identifica nel tragitto seguito dai raggi luminosi provenienti da un oggetto fissato con lo sguardo che passando dal centro della cornea (vertice) e proseguendo per il punto
nodale (posto poco al davanti della faccia posteriore del cristallino), giungono alla fovea centralis, zona retinica della visione distinta. La linea di visione non coincide con
l’asse ottico poiché diverge lateralmente ed indietro andando a formare con questo un angolo, che, in condizioni normali, è di circa 7°. L’equatore è una circonferenza tracciabile sulla superficie esterna del bulbo, giacente sul piano perpendicolare all’asse anatomico ed equidistante dai poli anteriore e posteriore. [1]
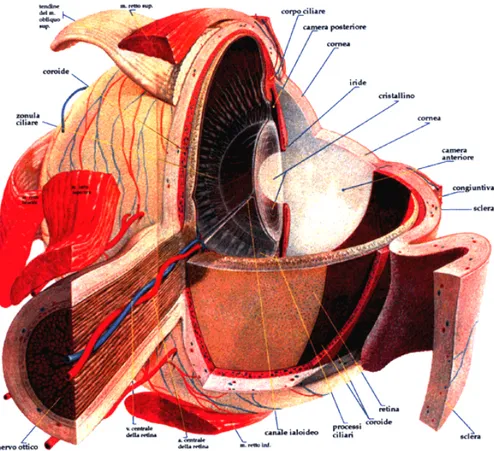
La struttura dell’occhio (fig. 1.2.1) è costituita da tre strati concentrici.
Lo strato esterno è una lamina fibrosa che comprende la cornea (trasparente) con il suo epitelio, la congiuntiva e la sclera (opaca). Lo spessore della sclera è variabile da 0.5 a 1 mm dal foro di emergenza del nervo ottico alla cornea.
Lo strato intermedio è una lamina vascolare che comprende l’iride e la coroide. L’iride è uno sfintere le cui fibre muscolari lisce sono disposte radialmente e circolarmente in modo tale da poter aprire o chiudere la pupilla, comportandosi quindi come un diaframma. La coroide è particolarmente ricca di vasi sanguigni che vanno ad irrorare gli strati esterni della retina. Gli strati interni invece ricevono sangue da rami dell’arteria centrale della retina, che decorrono parallelamente al nervo ottico.
Lo strato interno, o nervoso, dell’occhio è costituito appunto dalla retina, la cui porzione funzionale ricopre tutta la superficie posteriore dell’occhio, ad eccezione della macchia ceca, che corrisponde all’emergenza del nervo ottico. L’acuità visiva è massima nella regione centrale della retina che è definita macula lutea. La fovea centralis, un piccolo affossamento situato nel centro della macula, è la regione sulla quale vengono messi a fuoco gli stimoli visivi e costituisce il punto di fissazione, ovvero il punto sul quale vengono messi a fuoco i raggi luminosi quando gli occhi fissano un oggetto. Una curiosità: l’origine del nome “retina” deriva dalle prime osservazioni fatte su questo tessuto che sembrava circondare l’umor vitreo “come la rete di un cacciatore circonda la preda”.
Fig. 1.2.1: Struttura anatomica dell’occhio.
La messa a fuoco dell’oggetto da visualizzare è espletata dalla cornea (per 2/3, con un potere di +40 Diottrie) e dal cristallino (per il restante 1/3, con un potere massimo di +20 Diottrie). Quest’ultimo (fig. 1.2.2) è il più importante dei due avendo la capacità di modificare il proprio potere grazie al meccanismo dell’accomodazione. Proprio a causa di quest’ultima, il cristallino ha dimensioni variabili; comunque nella sua posizione di “riposo”, che si ha quando l’occhio è in assenza di accomodazione, mentre fissa un punto all’infinito (l’infinito accomodativo corrisponde a circa 6 metri di distanza), ha mediamente un diametro di 10 mm ed uno spessore di 5 mm. Il meccanismo di supporto del cristallino è costituito da legamenti sospensori (o fibre zonulari) che si inseriscono alla parete dell’occhio a livello del corpo ciliare, che è adibito alla contrazione del cristallino dovuta allo stimolo accomodativo.
Fig. 1.2.2: Il cristallino di un occhio proietta le immagini invertite sulla retina, analogamente a quanto fatto dall’obiettivo di una fotocamera.
La trasmissione della luce dall’esterno alla retina senza alcune distorsioni è garantita dall’umor vitreo e dall’umor acqueo, mezzi fluidi, rifrattivi, trasparenti che determinano la forma del globo oculare (funzione di sostegno), e da alcuni pigmenti, che hanno la funzione di ridurre la diffusione della luce. L’umor vitreo si parlerà più diffusamente in seguito (cap.2).
1.3 L’apparato muscolare
L’apparato motore dell’occhio comprende i muscoli estrinseci dell’occhio, retti e
obliqui, il muscolo elevatore della palpebra superiore; le fascie orbitali, il setto orbitale
e la fascia bulbare, nonché gli spazi orbitali, interfasciale e retrofasciale.
I muscoli estrinseci sono muscoli striati che si diversificano notevolmente da quelli delle restanti parti del corpo sia perché posseggono un’innervazione molto più ricca (sono quelli dotati delle più piccole unità motorie), sia per il fatto che le fibre che li costituiscono presentano maggiori variazioni morfologiche ed istochimiche. Sono stati distinti, infatti, tre tipi di fibre muscolari:
− spesse (diametro da 10 a 38 mm, provviste, in genere, di 2-3 nuclei), presenti soprattutto alle estremità dei muscoli e provviste di terminazioni nervose multiple;
− sottili (diametro da 13 a 30 mm, costituite da miofibrille molto sottili, organizzate in lamine), poco rappresentate in tutte le porzioni dei muscoli e provviste di un’unica terminazione nervosa;
− granulari (diametro da 13 a 38 mm, con miofibrille dalla morfologia irregolare, organizzate quasi a formare una rete), prevalenti nelle porzioni centrali dei muscoli e provviste di un'unica terminazione nervosa [1].
Questi muscoli sono allocati nell’orbita, in seno al suo corpo adiposo, e si inseriscono nella sclera e sono in numero di sei: quattro retti, rispettivamente superiore, inferiore, mediale e laterale, e due obliqui, superiore ed inferiore. Sono piccoli e lunghi, un po’ appiattiti parallelamente alla parete dell’orbita, dal cui fondo si dipartono, ad eccezione dell’obliquo inferiore. I muscoli retti si inseriscono all’emisfero anteriore, gli obliqui all’emisfero posteriore dell’occhio.
I quattro retti (superiore, inferiore, mediale o interno, laterale o esterno), fuoriescono da un ispessimento anulare della periorbita, definibile come anello tendineo comune, il quale circonda il tratto superiore e mediale del foro ottico, aderendovi, e l’estremo mediale della fessura orbitale superiore. Tale anello è in forma di imbuto allargato anteriormente e circonda il nervo ottico: i quattro muscoli sorgono dal rispettivo segmento dell’anello, e si recano in avanti divergendo in modo da costituire un imbuto sempre più ampio, alla cui apertura anteriore corrisponde il bulbo oculare e il cui asse è rappresentato all’incirca dal nervo ottico.
Divenuti tendinei, i muscoli perforano, in corrispondenza all’incirca dell’equatore del bulbo oculare, la fascia bulbare e, giunti così nello spazio interfasciale, si inseriscono alla sclera.
La lunghezza media dei muscoli reti è 40 mm. Il superiore e l’inferiore formano con la linea visuale un angolo θ di circa 25°; il mediale e il laterale decorrono secondo il meridiano orizzontale dell’occhio. Il retto mediale è il più robusto, il superiore il più gracile. I tendini dei quattro muscoli retti si inseriscono alla parte corrispondente della sclera, per la larghezza ciascuno di 1 cm, secondo linee un po’ convesse in avanti, quasi parallele al solco sclerale (quelle dei retti superiore ed inferiore ne divergono un po’ lateralmente).
Fig. 1.3.1: Muscoli dell'occhio osservati dal lato temporale. a, globo oculare; b, nervo ottico; c, c', palpebre superiore ed inferiore; d, seno mascellare; e, processo pterigoideo dello sfenoide; f, foro grande rotondo; g, volta dell'orbita; h, seno frontale; i, nervo sopraorbitale; k, setto orbitale. 1, muscolo elevatore della palpebra superiore; 2, retto superiore; 3, retto inferiore; 4, 4', retto laterale, sezionato dalla sua parte media; 5, retto mediale; 6, obliquo inferiore; 7, tendine del muscolo obliquo superiore; 8, anello dello Zinn.
Il muscolo obliquo superiore (o grande obliquo) è il più lungo e sottile ed il meno appiattito di tutti. Sorge dal fondo dell’orbita per mezzo di un breve tendine, sul contorno mediale del foro ottico dalla periferia dell’anello tendineo e dalla periorbita; decorre in avanti lungo l’angolo supero-mediale dell’orbita e, presso l’adito di questa, trapassa in un lungo e sottile tendine cilindroide, il quale si impegna in un semianello fibro-cartilagineo, la troclea, fissato alla periferia della fossetta trocleare dell’osso frontale. Scorrendo quivi per mezzo di un connettivo molto lasso, il tendine si flette lateralmente e indietro secondo un angolo di 40-50° e, mentre va sempre più appiattendosi, passa al di sotto del tendine del retto superiore e si inserisce sulla parte alta del quadrante supero-laterale della sclera, dietro all’equatore, secondo una linea lunga 11 mm, un po’ concava in avanti e medialmente.
Il muscolo obliquo inferiore (o piccolo obliquo) è il più breve di tutti, ha origine in corrispondenza dell’angolo infero-mediale dell’orbita, e presenta dei brevi tratti tendinei nella periorbita davanti della base della cresta lacrimale posteriore. Abbraccia il tendine del retto inferiore ed il bulbo oculare, nel quale si innesta nella parte alta del quadrante
infero-laterale, dietro all’equatore, tramite un tendine appiattito di lunghezza pari a 10 mm [3].
1.3.1 Le formazioni fasciali dell'orbita
II bulbo oculare è mantenuto nell’orbita da un sistema di fasce che rappresenta la soluzione ideale del problema della sospensione di uno sferoide in una cavità conica. La porzione prevalente del sistema è rappresentata dalla capsula di Tenone (fig. 1.3.2) o fascia del bulbo; si tratta di una formazione fibroelastica che avvolge il bulbo oculare dal nervo ottico, dove si unisce con le sue guaine, al limbus, dove si fonde con la congiuntiva anteriormente. Tra la capsula e la congiuntiva è compreso lo spazio sottocongiuntivale e, posteriormente, tra la capsula e la sclera, è delimitato lo spazio episclerale.
Fig. 1.3.2: Le varie guaine muscolari e la capsula di Tenone (delineate in blu).
Come già detto, i muscoli oculari perforano la capsula per inserirsi alla sclera e presentano perciò una porzione extra e una intracapsulare. Nella porzione extracapsulare ogni muscolo è avvolto da una guaina (figura 1.3.2), la cui superficie
raggiungono le rispettive pareti orbitarie andando a costituire i cosiddetti legamenti
d’arresto. Quest’ultimi hanno lo scopo di limitare la contrazione e il rilasciamento
muscolare e di conseguenza gli spostamenti eccessivi in avanti e in dietro del bulbo oculare. I retti verticali ed obliqui presentano legamenti d’arresto poco sviluppati, costituiti dal legamento di Whitnall e dal legamento di Lockwood. Il primo è un’espansione fibrosa situata nella parte superiore dell’orbita e formata dalla fusione delle guaine del retto superiore, dell’obliquo superiore e del muscolo elevatore della palpebra superiore. Il secondo è un’espansione connettivale posta nella parte inferiore dell'orbita e formata dalla fusione delle guaine del retto inferiore e dell’obliquo inferiore. Queste espansioni aponeurotiche, fissate ai bordi mediale e laterale dell’orbita, intervengono funzionalmente nel corso dei movimenti verticali dell'occhio, limitandone gli spostamenti.
L’intero sistema fasciale, inoltre, esplica la sua funzione nel limitare le contrazioni muscolari, riducendo gli spostamenti antero-posteriori dell’occhio, e conservandone il centro di rotazione relativamente costante in rapporto alla piramide orbitaria; in virtù dell’intervento dei legamenti d’arresto i movimenti oculari risultano smorzati, graduati ed armoniosi [4].
1.4 L’apparato nervoso
Il sistema visivo possiede i circuiti nervosi più complessi di tutti i sistemi sensoriali. Il nervo acustico contiene circa 30.000 fibre, mentre il nervo ottico ne contiene più di un milione. È anche per questo che, tra gli organi di senso, la vista è senza dubbio quello più adatto ad esplorare lo spazio sulla lunga distanza.
L’efficienza visiva, inoltre, è elevata sia durante le ore diurne che nella relativa oscurità. Questo è dovuto alla straordinaria versatilità del nostro apparato fotorecettore costituito dalla retina. Infatti i recettori retinici più evoluti, i bastoncelli, sono in grado di percepire lo stimolo apportato anche solo da un singolo fotone. Sono quindi i più sensibili recettori sensoriali, anche se non i più veloci, dato che le cellule acustiche sono circa 100 volte più rapide di quelle visive nella registrazione dell'impulso[5].
La presenza di fotorecettori (i coni ed i già citati bastoncelli) e di organi che li assemblino costituendo “occhi” deve aver assicurato vantaggi selettivi decisivi, dato che i 6 phyla animali che li posseggono (su un totale di 30 phyla) includono ben il 95% delle specie conosciute. Vengono di seguito trattate le vie visive centrali.
1.5 Le vie visive centrali
Il flusso delle informazioni visive può essere analizzato dividendolo in due stadi successivi: dalla retina al mesencefalo e al talamo e, dal talamo alla corteccia visiva primaria.
1.5.1 Dalla retina al mesencefalo e al talamo
È opportuno, in prima battuta, mettere in evidenza come le immagini retiniche siano diverse da quelle del campo visivo, poiché le prime sono invertite rispetto alle seconde. Il campo visivo è la parte del mondo esterno che viene vista dai due occhi in assenza di movimenti del capo. È possibile definire la metà destra e sinistra del campo visivo quando le fovee di entrambi gli occhi fissano un sol punto nello spazio.
monoculare: la luce che proviene dalla regione temporale di ciascun emicampo visivo
colpirà soltanto l’emiretina nasale dell’occhio omonimo (o emiretina nasale
ipsolaterale).
Il disco ottico, la zona della retina da cui fuoriescono gli assoni delle cellule gangliari, è del tutto insensibile alla luce e costituisce una macchia cieca nella retina. Poiché questo è posto medialmente ad entrambe le fovee, la luce proveniente da un punto qualunque della regione binoculare non può mai andare a cadere simultaneamente su entrambe le macchie cieche e perciò, normalmente, non ci si rende conto della loro esistenza.
Nel seguire il flusso delle informazioni visive verso le stazioni cerebrali è importante tenere presente la corrispondenza fra le diverse regioni del campo visivo e le rispettive immagini retiniche. Tale corrispondenza può rivelarsi piuttosto difficile da seguire per due ragioni:
− il cristallino inverte le immagini visive (es.: la metà superiore del campo visivo si proietta sulla metà inferiore (o ventrale) della retina);
− la zona binoculare di ciascun emicampo visivo proietta su parti diverse delle due retine (es.: un punto luminoso presente nella parte binoculare dell’emicampo visivo di destra andrà a cadere sulla emiretina temporale dell’occhio sinistro e sulla emiretina nasale dell’occhio destro) (fig. 1.5.1).
Gli assoni di tutte le cellule gangliari retiniche vanno a confluire nel disco ottico dove divengono mielinici e vanno a formare, bilateralmente, i nervi ottici. Questi proiettano quindi al chiasma ottico; qui le fibre delle metà nasali di ogni retina si incrociano e proiettano all’emisfero cerebrale contralaterale (fig. 1.5.1).
I tratti ottici, infine, proiettano a tre principali stazioni sottocorticali: il pretetto, il
collicolo superiore, il corpo genicolato laterale (fig. 1.5.2). Le funzioni di questi tre
centri sono estremamente complesse. È possibile comunque schematizzare il ruolo di in rapporto all’elaborazione dell’informazione visiva:
- l’area pretettale del mesencefalo controlla i riflessi pupillari;
- il collicolo superiore è il principale controllore dei movimenti saccadici oculari;
- il corpo genicolato laterale (CGL) è la principale stazione di ritrasmissione delle informazioni afferenti destinate alla corteccia visiva.
Fig. 1.5.2: stazioni sottocorticali.
1.5.2 Dal talamo alla corteccia visiva primaria
II novanta percento degli assoni retinici va a terminare nel CGL, questo è un nucleo del talamo che costituisce la più importante stazione sottocorticale di trasmissione delle informazioni visive alla corteccia cerebrale. Se vengono a mancare queste afferenze, la percezione visiva è praticamente perduta. È quindi evidente come il CGL sia fondamentale per la visione.
Questo nucleo è organizzato in maniera tale che vi sia mantenuto un alto grado di retinotopia ed inoltre vi sia ancora una separazione anatomica di alcune caratteristiche dell'informazione visiva, qual ad esempio l’occhio da cui proviene l’impulso, o il tipo di cellula gangliare che gli ha dato origine.
I campi recettivi dei neuroni del genicolato laterale sono molto simili a quelli delle cellule gangliari della retina ed è proprio per questo che è stato a lungo considerato un semplice nucleo di ritrasmissione della via retino-striata. In realtà solo il 10-20% delle
probabile funzione sembrerebbe legata all’analisi del contrasto, a una qualche selezione attentiva degli stimoli visivi, ovvero provvederebbe a regolare l’entità, e forse la temporizzazione, del flusso di informazioni verso la corteccia striata.
Continuando il percorso dell’informazione visiva, la quasi totalità delle fibre provenienti dai neuroni del corpo genicolato termina nella corteccia striata, o area 17
di Brodmann, nota anche come corteccia visiva primaria, che nell’uomo si trova lungo i
bordi della scissura calcarina.
Sull’area 17 di ciascun emisfero si trova rappresentato l’emicampo visivo contralaterale in maniera tale che se la scissura calcarina venisse distesa, la corteccia striata apparirebbe come un settore circolare sul quale possono essere riportate le coordinate dell’emicampo visivo. Se si suddivide idealmente il campo visivo in piccole parti uguali, ciascuna di esse viene rappresentata su una superficie corticale tanto più ampia quanto più è vicina alla fovea; in particolare, circa la metà della massa del tessuto nervoso della corteccia è devoluta alla rappresentazione della fovea e delle regioni immediatamente circostanti (fig. 1.5.3). È quindi evidente che anche a questo livello abbiamo un’organizzazione retinotopica, tuttavia, le proprietà dei campi recettivi cellulari cambiano in maniera significativa.
Fig. 1.5.3: La corteccia visiva primaria contiene una rappresentazione ordinata di tutto il campo visivo. Ogni emicampo è rappresentato nell’emisfero contralaterale.
L’informazione visiva, una volta trasmessa alla corteccia striata dalle fibre afferenti che provengono dal CGL, circola da uno strato corticale all’altro secondo un piano
sistematico che ha appunto inizio dalle cellule stellate spinose che sono il tipo cellulare predominante nello strato IV. Le cellule stellate spinose distribuiscono le informazioni ricevute dal CGL alla corteccia, mentre le cellule piramidali attraverso collaterali dei propri assoni integrano l’attività delle cellule degli strati soprastanti e sottostanti di VI. Come si riflette una tale complessità dei circuiti corticali sulle proprietà delle cellule della corteccia stessa?
Hubel, Wiesel et al. (dagli anni ’60 in poi) hanno osservato che i requisiti necessari per ottenere una risposta da parte delle cellule degli strati soprastanti e sottostanti al IV strato erano molto più complessi rispetto a quelli che eccitano le cellule della retina e del CGL. Le cellule, infatti, rispondevano in maniera ottimale a stimoli di forma rettilinea, come una linea o una sbarretta. Queste cellule si possono suddividere in due categorie principali: cellule semplici e cellule complesse.
Le cellule semplici forniscono la loro miglior risposta ad una sbarretta luminosa dotata di un orientamento specifico. Il loro campo recettivo, infatti, presenta sia zone eccitatorie che zone inibitorie leggermente più grandi di quelle delle cellule del CGL. Ad esempio, per una cellula che abbia un campo recettivo con una zona eccitatoria rettilinea, lo stimolo ottimale deve eccitare quel tratto particolare di retina che è innervato dai recettori che stanno nella zona eccitatoria, deve avere proprietà lineari adatte e deve avere un asse di orientamento specifico.
I campi recettivi delle cellule complesse hanno, in generale, dimensioni maggiori di quelli delle cellule semplici. Anche i loro campi recettivi sono dotate di un asse di orientamento specifico; ma la posizione esatta dello stimolo, all’interno del campo recettivo, non appare essenziale, in quanto non esistono zone eccitatorie e inibitorie ben definite.
Da questo tipo di considerazioni, Hubel e Wiesel hanno ipotizzato che l’elaborazione effettuata dalle vie recettive del segnale visivo, costituisca un passaggio importante per l’analisi delle sagome degli oggetti. In effetti anche la semplice visione dei contorni può essere sufficiente al riconoscimento: la parte interna ed uniforme di un oggetto così come la superficie di uno sfondo non forniscono alcuna informazione rilevante [6].
Bibliografia
[1] http://www.units.it/~brain/visione/index.html - Appunti di oculistica. Centro interdipartimentale per le Neuroscienze B.R.A.I.N. in collaborazione con il Comitato per la Promozione delle Neuroscienze.
[2] Robert M. Berne, Matthew N. Levy (1998). Principi di Fisiologia. Casa Editrice Ambrosiana. Edizione italiana a cura di Tullio Manzoni e Fiorenzo Conti, Istituto di fisiologia umana dell'Università di Ancona.
[3] Antonio Pensa, Giuseppe Favaro (1970). Anatomia umana. Casa Editrice UTET. [4] M. Miglior, B. Bagolini, B. Boles Carenini, N. Orzalesi, L. Scullica, M. Zingirian.
Oftalmologia clinica. Casa Editrice Monduzzi Editore.
[5] Ottoboni, E. Mora, A. Salami; atti delle XIII giornate italiane di otoneurologia del 1996. Funzione vestibolare e sensibilità spaziale nell’uomo.
[6] E. R. Kandel (2003). Principi di Neuroscienze. Terza edizione italiana. Casa