4. DISCUSSIONE
4.1. Diversità genetica all’interno dei campioni
L’analisi delle 75 sequenze del gene mitocondriale Cyt b di Paracentrotus
lividus ha consentito di stimare la diversità genetica all’interno dei campioni delle
singole località. La diversità aplotipica si è rivelata relativamente bassa per il campione di Galway (h = 0.400), con quattro animali che presentano il medesimo aplotipo. Questo risultato era atteso, poiché il campione è stato raccolto in una popolazione artificiale di un impianto di acquacoltura, in cui gli individui derivano da fecondazione in vitro a partire da pochi riproduttori. L’effetto è quindi quello di un pronunciato inbreeding con forte riduzione della variabilità genetica. Per tutti gli altri campioni sono stati invece riscontrati valori di diversità aplotipica molto alti, paragonabili a quelli ricavati da Duran et al. (2004), che hanno utilizzato il gene mitocondriale codificante per la subunità I della citocromo ossidasi (COI) nella stessa specie. Se osserviamo l’intervallo di variazione dei valori di diversità aplotipica del presente lavoro, omettendo il campione atlantico di acquacoltura, sono stati ottenuti valori compresi tra 0.700 e 1.000, mentre Duran et al. (2004) hanno rilevato valori compresi tra 0.844 e 1.000. Analogamente la diversità nucleotidica (π), che nel nostro caso varia da 0.002 a 0.010, è dello stesso ordine di grandezza rispetto a quella ottenuta da Duran et al. (2004), che hanno rilevato valori compresi tra 0.0035 e 0.0098. È comunque interessante notare come i campioni provenienti dal Mar Adriatico, insieme a quelli atlantici di Baiona, presentino valori di π nettamente più bassi degli altri.
4.2. Struttura genetica
Le modalità di riproduzione degli invertebrati marini bentonici sono cruciali nel determinare il potenziale per la dispersione della maggior parte delle specie e, di conseguenza, per i livelli di strutturazione genetica e di flusso genico tra popolazioni o sottopopolazioni. Pertanto, non sorprende che il grado di divergenza genetica tra popolazioni di specie con larve planctotrofiche a lunga vita pelagica sia tipicamente
basso, in quanto il flusso genico può avvenire anche a distanze considerevoli. È da notare comunque come studi più o meno recenti abbiano trovato eccezioni a quanto detto finora, evidenziando casi in cui larve a lunga vita pelagica non determinano ampia dispersione. Burton (1983), in una revisione su questo argomento, ha riportato e commentato diversi casi reperibili in letteratura in cui si riscontra eterogeneità genetica su scala locale a dispetto di stadi larvali planctonici prolungati. Tra questi organismi sono da menzionare il crostaceo decapode Homarus americanus (Tracey
et al., 1975), il mollusco bivalve Mytilus edulis (Koehn, 1975) e l’echinoide Arbacia punctulata (Marcus, 1977). Tra i lavori più recenti sono da riportare alcuni casi,
riguardanti specie di pesci i) della barriera corallina caraibica, come il labride
Thalassoma bifasciatum (Swearer et al., 1999) e il gobide Elacatinus evelynae
(Taylor & Hellberg, 2003); ii) della barriera corallina australiana, come il pomacentride Pomacentrus amboinensis (Jones et al., 1999); iii) del mollusco bivalve del Mare del Nord Machoma baltica (Luttikhuizen et al., 2003); iv) degli echinoidi di acque temperate Heliocidaris spp. e Strongylocentrotus spp. (Palumbi & Wilson, 1990; Palumbi & Kessing, 1991; McMillan et al., 1992; Edmands et al., 1996). Nell’echinoide Echinotrix diadema, per il quale sono stati utilizzati mtDNA e alloenzimi come marcatori molecolari, i risultati suggeriscono un flusso genico avvenuto in tempi recenti attraverso l’intero Oceano Pacifico, probabilmente legato all’effetto periodico del Niño (Lessios et al., 1998). Molti di questi casi riflettono flusso genico più o meno recente che si verifica per dispersione naturale; non è comunque da trascurare un altro meccanismo, che spesso rappresenta un vero e proprio problema ecologico, da ascrivere soprattutto all’ultimo secolo: si tratta del trasporto antropico di organismi marini, spesso su scala globale, attraverso le acque di sentina delle navi (Carlton & Geller, 1993; Lodge, 1993). Da rilevare che fattori abiotici e biotici possono ridurre in maniera drastica la dispersione larvale. Ad esempio, particolari fenomeni oceanografici come “eddies” e “gyres” possono avere molteplici effetti: possono impedire la dispersione su larga scala delle larve, costringendole a rimanere confinate in un moto circolare per l’intera durata della loro vita planctonica, contribuendo quindi ad un relativo isolamento e conseguente differenziamento genetico tra siti distanti. Per contro, gli stessi fenomeni possono favorire il rimescolamento delle larve originate da gameti prodotti da differenti pool di
riproduttori, anche geograficamente distanti, risultando in una maggiore omogeneità nella composizione genetica delle future reclute. In una fase successiva, la selezione differenziale di specifici genotipi in seguito all’insediamento larvale, può indurre una caratterizzazione genetica di popolazioni le cui larve derivino da fonti multiple e siano presumibilmente eterogenee dal punto di vista genetico (Hartl & Clark, 1993). Fattori come la migrazione verticale, il nuoto e una relativa tendenza a mantenere il contatto con il fondo possono anche far sì che le larve si allontanino dal loro luogo d’origine meno del previsto (Raimondi & Keough, 1990).
L'analisi della divergenza genetica tra i campioni di Paracentrotus lividus (vedi paragrafo 3.2) ha messo in evidenza una generale omogeneità tra i 15 campioni considerati (P = 0.282 con il test esatto). Questo scarso grado di diversità genetica tra individui che si trovano anche in località geograficamente distanti concorda con quanto rilevato da Arculeo et al. (1998) e Duran et al. (2004) in P. lividus e da Palumbi (1996), Palumbi et al. (1997), Lessios et al. (2003) e Addison & Hart (2004) per altri echinoidi con un potenziale di dispersione paragonabile o superiore a quello di P. lividus. L’analisi della varianza molecolare (AMOVA) ha attribuito circa l’88% della varianza alla componente inter-individuale all’interno dei campioni (Tab. 3.3). Tale proporzione è risultata più bassa di quanto rilevato da Duran et al. (2004), che hanno ottenuto il 99% per questa componente. Situazioni di scarsa diversità genetica tra campioni geograficamente distanti non sono infrequenti negli invertebrati marini ad ampia dispersione e sono una conseguenza del fatto che individui campionati nella medesima località presentano tra loro una maggiore differenziazione genetica che non rispetto ad altri prelevati a centinaia o, talvolta, migliaia di chilometri. A questo proposito, ricordiamo per esempio i molluschi gasteropodi del genere
Siphonaria (Johnson & Black, 1982; 1984) e l’echinoide Echinometra mathaei (Watts et al., 1990). I risultati dell’AMOVA ottenuti nel presente lavoro concordano con
l’ipotesi generale di lieve strutturazione della specie, confermando un notevole effetto della dispersione larvale nel determinare l’assenza di pattern geografici riconoscibili. I valori leggermente negativi ottenuti nell’AMOVA che considera i raggruppamenti biogeografici (Tab. 3.4) sono determinati dall’assenza di una marcata strutturazione genetica e possono anche essere il risultato di un elevato tasso di “outcrossing”. Ciò significa che geni di individui campionati in località differenti possono essere più affini
tra loro di geni di individui della medesima località. Nel caso di P. lividus non è possibile parlare di “outcrossing” in termini classici, in quanto gli spermatozoi perdono la capacità di fecondare le uova nel giro di poche ore (L. Kozinkova, comunicazione personale) e la percentuale di successo fecondativo è proporzionale alla concentrazione degli spermatozoi; la fecondazione avverrà quindi necessariamente entro breve termine, tra gameti prodotti da individui relativamente vicini. Successivamente la larva può percorrere distanze notevoli, trasportando altrove l’aplotipo materno. Alla generazione successiva l’aplotipo può disperdersi nuovamente a partire da più siti e così via. I valori della Φ-statistica ottenuti sono risultati tutti significativi al test di permutazione (P < 0.05), indicando l’assenza di una situazione di panmissia e la presenza di strutturazione genetica che può essere definita “moderata”, alla luce di quanto detto sopra.
L’MDS degli aplotipi, (Figg. 3.3.A e B), ottenuto con le distanze genetiche di Tamura & Nei (1993) per ciascuna coppia di individui, non mette in evidenza particolari relazioni biogeografiche e ripartisce gli aplotipi in due gruppi principali separati in maniera netta, che corrispondono agli aplogruppi dei network (Figg. 3.1 e 3.2). Quando agli aplotipi sono stati sostituiti i centroidi dei singoli campioni, le località atlantiche vengono chiaramente separate da quelle mediterranee lungo l’asse verticale (Fig. 3.4.A). Con l’esclusione dei campioni atlantici, l’MDS ha invece evidenziato una netta separazione tra le località dell’Adriatico e le altre località mediterranee lungo l’asse orizzontale (Fig. 3.4.B). Questo dato suggerisce l’esistenza di un grado di divergenza genetica tra gli individui di P. lividus del Mare Adriatico e quelli del resto del Mediterraneo, come già ipotizzato nello studio pilota comprendente campioni di quattro località mediterranee e una atlantica (Barbieri et
al., 2005).
La F-statistica calcolata per tutte le possibili coppie di campioni (Tab. 3.2) ha rivelato valori elevati e significativamente maggiori di zero per tutti i confronti comprendenti il campione di Galway. Ciò è dovuto alla peculiarità di questo campione, che, come abbiamo visto, proviene da un impianto di acquacoltura. Il campione atlantico di Baiona non presenta invece nessun tipo di distinzione nei confronti degli altri, confermando per P. lividus, l’assenza di una marcata divergenza genetica tra i popolamenti dell’Oceano Atlantico e quelli del Mar Mediterraneo. Più
interessante può essere considerata la situazione del campione adriatico di Ancona, che presenta valori di FST significativamente maggiori di zero, in alcuni casi anche
molto elevati, nei confronti di quasi tutte le altre località. Inoltre, seppure in misura minore, anche altri due campioni adriatici, Brindisi e Mljet, presentano valori compatibili con un certo grado di isolamento, contribuendo all’ipotesi di un relativo isolamento, seppure blando, tra l’Adriatico ed il resto del Mediterraneo.
Il flusso genico calcolato con il metodo di Nei (1982) presenta valori piuttosto bassi quando si considera il totale dei campioni analizzati. Ancora una volta questa condizione è probabilmente da ascrivere al campione atlantico di Galway e rappresenta una sottostima del flusso genico reale in P. lividus. Anche la stima relativa al solo Atlantico presenta un valore troppo basso per essere giudicato attendibile ed è necessario ampliare il disegno di campionamento in questo bacino per ottenere stime affidabili. Per il Mediterraneo è possibile osservare come nel complesso il flusso genico risulti moderatamente alto; esso aumenta notevolmente se si considerano separatamente i bacini orientale ed occidentale del Mediterraneo. Il Mar Adriatico si configura ancora una volta come un caso particolare e, a dispetto della scarsa estensione, presenta, al suo interno, livelli di flusso genico decisamente più bassi delle altre aree considerate nell’analisi. Questo risultato conferma ulteriormente l’ipotesi dell’esistenza di barriere fisiche e/o ecologiche che influenzano negativamente la dispersione in quest’area.
4.3. Aspetti di demografia storica
Analizzando i risultati sulla diversità genetica all’interno dei campioni di P.
lividus ottenuti in questo lavoro e in Duran et al. (2004), è possibile fornire una lettura
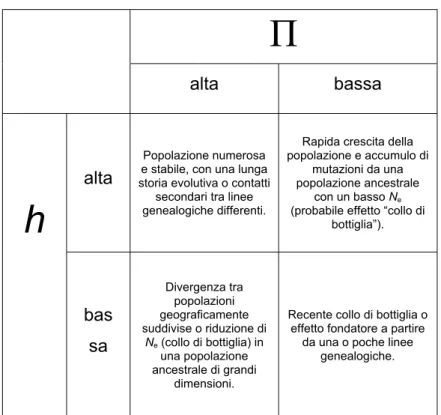
in chiave demografica della storia pregressa della specie. Seguendo le indicazioni suggerite da Grant & Bowen (1998), si possono ipotizzare quattro situazioni limite, derivanti dalle diverse combinazioni dei valori di diversità aplotipica e di quella nucleotidica (Fig. 4.1).
Π
alta bassa
alta
Popolazione numerosa e stabile, con una lunga storia evolutiva o contatti
secondari tra linee genealogiche differenti.
Rapida crescita della popolazione e accumulo di
mutazioni da una popolazione ancestrale
con un basso Ne
(probabile effetto “collo di bottiglia”).
h
bas sa Divergenza tra popolazioni geograficamente suddivise o riduzione di Ne (collo di bottiglia) in una popolazione ancestrale di grandi dimensioni.Recente collo di bottiglia o effetto fondatore a partire
da una o poche linee genealogiche.
Fig. 4.1. Situazioni limite derivanti da diverse combinazioni di valori di diversità aplotipica e diversità nucleotidica. Modificato da Grant & Bowen (1998).
Seguendo questa linea teorica, i dati rilevati nel presente lavoro portano a ritenere che, sia per le singole località, sia per l’intera area di studio, gli odierni popolamenti di P. lividus derivino da un processo di espansione demografica prodottasi in seguito ad una riduzione storica del numero di individui. Una simile dinamica è in grado di lasciare un’impronta dal punto di vista genetico: l'attuale situazione potrebbe essere stata raggiunta per accumulo di mutazioni puntiformi a partire da una ristretta popolazione efficace, e quindi da un numero relativamente piccolo di aplotipi ancestrali. I dati relativi ad una possibile espansione in un passato relativamente recente possono trovare conferma nella disposizione degli aplotipi nei network. I motivi che possono avere prodotto la riduzione delle popolazioni di P.
lividus in passato, o, in ultima analisi, di fenomeni locali di
estinzione-ricolonizzazione, sono sconosciuti (Barnes et al., 2001). A tale proposito risulta importante segnalare l’estesa moria di individui di P. lividus che si verificò negli anni ’80 e che è noto per la maggior parte dell’arcipelago toscano e delle coste toscane continentali. Sulla natura di questa moria è possibile escludere con una certa affidabilità l’effetto dell’inquinamento antropico, visto che il fenomeno si è verificato anche sulle coste delle isole e dell’Argentario, che erano esenti da ogni forma di
impatto (G. Cognetti, comunicazione personale). Non è da escludere l’effetto di un agente patogeno, come dimostrato nel caso di Zostera marina, soggetta all’infezione del fungo Labyrinthula macrocystis (Davidson & Hughes, 1998); in questi casi si assiste generalmente allo sviluppo di forme di resistenza che consentono il ripopolamento (G. Cognetti, comunicazione personale).
I network ottenuti per P. lividus mostrano una disposizione piuttosto disordinata degli aplotipi, e, parallelamente a quanto riportato da Duran et al. (2004), si può notare come alcuni aplotipi siano condivisi in qualche caso da individui provenienti da località geograficamente molto distanti. D’altro canto, il ritrovamento di sequenze di mtDNA identiche in località molto distanti è di fatto comune per invertebrati marini con dimensioni efficaci delle popolazioni relativamente grandi e con elevato potenziale di dispersione e quindi flusso genico (Avise, 1994, 2000; Palumbi, 1996). I network inoltre mostrano una netta separazione in due aplogruppi, separati da 11 mutazioni. I due gruppi differiscono per composizione: uno, che contiene il maggior numero di individui, è costituito da aplotipi relativi ad animali campionati nell’intera area considerata, che suggerisce l’assenza di barriere alla migrazione dei gameti e delle larve, in accordo con alti livelli di flusso genico. Nell’altro gruppo, numericamente inferiore, sono invece assenti aplotipi di provenienza adriatica o atlantica (Fig. 3.2). Questo risultato suggerisce che l’elevato potenziale per la dispersione contribuisce ad una diffusione vastissima di alcuni aplotipi e quindi ad una scarsa differenziazione tra campioni di località anche molto distanti. La dispersione su larga scala, però, non è sufficiente a omogeneizzare completamente la distribuzione degli aplotipi, in quanto, seppur moderata, è stata rilevata strutturazione genetica. La presenza di aplotipi ai quali sono connessi altriaplotipi affini ai primi, cioè separati da uno o due mutazioni (“star phylogeny”), nell’intera area considerata, potrebbe non solo essere il riflesso diretto di elevato flusso genico al momento attuale, ma anche essere frutto di una espansione demografica risalente al passato recente della specie. Una successiva riduzione dei livelli di flusso genico e trasporto larvale potrebbe essere sufficiente per favorire un successivo differenziamento genetico su scala locale, come suggerito da Palumbi et al. (1997) per specie di echinoidi tropicali del genere Echinometra. Non si può escludere, comunque, l’errore di campionamento, cioè l’assenza di individui adriatici e atlantici
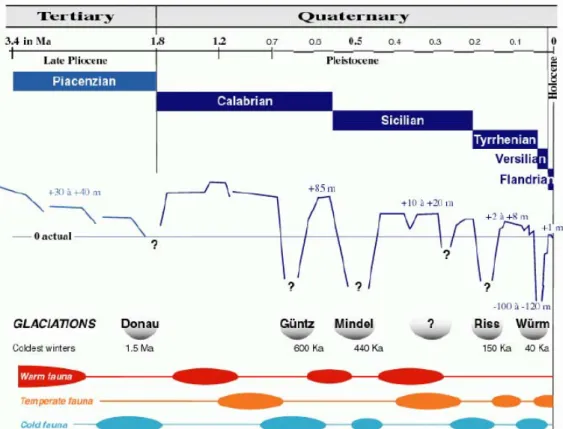
nell’aplogruppo più piccolo potrebbe venir meno aumentando il numero di individui analizzati per campione. Nelle Figg. 3.1 e 3.2 sono riconoscibili alcune “star phylogeny”, in cui al centro si ritrova un aplotipo ancestrale più abbondante, mentre gli aplotipi di più recente derivazione si connettono al primo tramite rami corti. Le “star phylogeny” evidenziano graficamente quello che esprimono numericamente alti valori di diversità aplotipica e bassi valori di diversità nucleotidica. L’alta diversità aplotipica è riconoscibile nell’elevato numero di aplotipi (rappresentati graficamente come cerchi e ovali rispettivamente nelle Figg. 3.1 e 3.2), mentre la bassa diversità nucleotidica è riscontrabile nel bassissimo numero di nodi, e quindi di mutazioni, che separano questi aplotipi nel grafico. Facendo riferimento alla classificazione proposta da Avise (2000), è possibile evidenziare nei network ottenuti una disposizione riconducibile nel complesso alla categoria II. L’interpretazione che Avise (2000) dà a tale pattern è peraltro compatibile con le caratteristiche biologiche di P. lividus. Tale pattern può essere interpretato come il risultato dell’erosione genetica di tutti gli aplotipi intermedi, oppure come caratteristico di popolazioni storicamente separate e che sono poi state soggette a contatto secondario. La possibilità in P. lividus di una separazione in popolazioni isolate con conseguente differenziamento genetico non ha riscontri certi. Un’ipotesi che potrebbe essere avanzata a tale riguardo è che durante il Pleistocene, periodo caratterizzato dall’alternanza di lunghe glaciazioni e brevi periodi interglaciali, il periodico susseguirsi di drastici cambiamenti climatici e, in particolare le fluttuazioni del livello del mare (Fig. 4.2), abbiano promosso fenomeni di estinzione-ricolonizzazione e/o isolamento-contatto secondario tra popolamenti differenti, analogamente a quanto rilevato per le faune bentoniche profonde (Emig & Geistdoerfer, 2004).
Fig. 4.2. Dati relativi al Mar Mediterraneo da 3.4 milioni di anni fa ad oggi: la linea spezzata in blu indica il livello delle acque, gli ellissi in bianco le principali glaciazioni, gli ellissi in rosso, arancio e azzurro rappresentano rispettivamente la presenza di faune “calde”, “temperate” e “fredde”. Da Emig & Geistdoerfer (2004).
L’ipotesi di un’espansione demografica pleistocenica in P. lividus, stimata in circa 130000-300000 anni fa, è stata avanzata da Duran et al. (2004). Nel presente lavoro l’ipotesi dell’espansione demografica è confermata dai risultati ottenuti dalla distribuzione mismatch (Fig. 3.7) e confermata statisticamente dal test R2 (Tab. 3.8),
quando applicati all’intero insieme dei dati (con la sola omissione del campione di acquacoltura di Galway). Le analisi applicate ai vari sottoinsiemi, corrispondenti ai raggruppamenti biogeografici impiegati anche nell’AMOVA, hanno consentito di focalizzare l’attenzione sulle singole regioni. La distribuzione mismatch ha mostrato un andamento bimodale per il totale dei dati e per i vari sottoinsiemi di dati considerati nelle analisi successive. Da rilevare che l’andamento bimodale della distribuzione mismatch rafforza l’interpretazione dei network come riconducibili alla categoria II di Avise (2000). L’Adriatico si discosta dai restanti raggruppamenti biogeografici e mostra una distribuzione osservata della distribuzione mismatch che è molto vicina a quella attesa, fedele quindi al modello dell’espansione demografica.
come la significatività rilevata per alcuni sottoinsiemi di dati fosse un effetto condizionato dalla presenza dei campioni dell’Adriatico; in realtà è per l’Adriatico che si riscontrano i valori più consistenti con l’ipotesi di espansione demografica e il maggior grado di significatività (P < 0.001), mentre per il resto le analisi non mettono in evidenza casi di accordo con il modello di espansione demografica.
4.4. Gestione
I risultati del presente lavoro non chiariscono definitivamente come possa essere applicato il concetto di popolazione a Paracentrotus lividus. Ciò rende assai difficile inferire sulla natura e sulla dimensione di eventuali stock genetici e complica notevolmente la realizzazione di efficaci piani di gestione e di regolamentazione del prelievo di questa risorsa. I dati ottenuti nel presente lavoro insieme a quelli ottenuti da Duran et al. (2004) consentono comunque di attestare per questa specie un generale stato di “buona salute genetica”, riscontrabile nell’elevata variabilità genetica dei campioni. Nonostante non siamo in possesso di dati storici sulla variabilità genetica di questa specie, dati gli elevati valori di variabilità genetica rilevati, si presume che l’erosione genetica dovuta al prelievo non sia avvenuta o sia comunque molto bassa.Segnali meno rassicuranti arrivano da altri settori: drammatiche oscillazioni nell’abbondanza di P. lividus sono note da tempo anche se ne sono ignote le cause (Barnes et al., 2001), fenomeni di mortalità di massa sono riportate per le coste Toscane negli anni ’80 a causa di un probabile agente patogeno (G. Cognetti, comunicazione personale) e nei primi anni del 2000 nelle acque antistanti alla provincia di Massa Carrara per le fioriture estive di dinoflagellati tossigeni (Sansoni et al., 2003). Si riscontra inoltre, in alcuni siti della Puglia soggetti a intensa attività di pesca, un sensibile aumento delle classi giovanili (Guidetti et al., 2004), che, come è noto, è una tipica conseguenza del sovrasfruttamento (overfishing). Fino ad alcuni decenni fa era diffusa l’idea che gli invertebrati marini non potessero essere soggetti a sovrasfruttamento per le loro caratteristiche biologiche peculiari: tassi di crescita relativamente rapidi, alta fecondità, e dispersione larvale su larghissima scala. Alcuni studi hanno comunque dimostrato come l’intenso prelievo dell’echinoide delle coste pacifiche nordamericane
Strongylocentrotus purpuratus possa portare a fenomeni di sovrasfruttamento (FAO,
1996; Pfister & Bradbury, 1996; Tegner et al., 1996). Gli effetti di una pesca sconsiderata di P. lividus non influirebbero solamente sulla specie interessata, ma potrebbero avere conseguenze più o meno indirette su tutte quelle che ne condividono l’habitat, innescando processi complessi, con la capacità di propagarsi all’interno dei vari livelli trofici e di modellare le comunità bentoniche, alterando processi biologici importanti (Guidetti et al., 2004). Tra l’altro la possibilità di una vicarianza ecologica da parte di A. lixula è a tutt’oggi dibattuta, e non si può arguire se gli eventuali effetti di un crollo demografico di P. lividus sugli ecosistemi bentonici possano essere bilanciati in qualche misura dall’attività di A. lixula (Benedetti Cecchi
et al., 1998; Bulleri et al., 1999; Guidetti et al., 2004).
La densità costante nei popolamenti di P. lividus, riscontrata in zone notoriamente sfruttate della Puglia (Guidetti et al., 2004), suggerisce una notevole capacità di resilienza in questa specie. Pe runo sfruttamento sostenibile di questa risorsa, oltre al rispetto delle attuali norme sulla pesca di P. lividus, che prevedono la taglia minima di cattura e la quota quotidiana pro capite, potrebbe essere utile l’introduzione di una rotazione delle aree di pesca, al fine di favorire il ripopolamento dei siti sfruttati (Guidetti et al., 2004). Un’altra possibile regolamentazione è quella che viene attuata in Sardegna nell’area marina protetta della Penisola del Sinis – Isola di Mal di Ventre: qui la pesca di P. lividus sostiene una piccola economia locale ed è ulteriormente limitata dal divieto di utilizzo di apparecchiature subacquee. La cattura può quindi avvenire solamente da natante per mezzo di pertiche o lunghi rastrelli o retini, preservando totalmente tutti gli individui che vivono sotto l’isobata dei 5 m (Baroli et al., 2006). Gli individui profondi contribuiscono ogni anno al reclutamento delle zone più superficiali intensamente sfruttate. Politiche di questo tipo, affiancate anche dai sempre più numerosi progetti di acquacoltura, potrebbero evitare il collasso della risorsa ed una conseguente crisi della pesca del riccio di mare edule e del suo indotto.