INDICE
RIASSUNTO
31.
INTRODUZIONE
51.1 Biologia ed ecologia di Aristeus antennatus 6
1.1.1 Inquadramento sistematico e riconoscimento 6
1.1.2 Distribuzione geografica 8
1.1.3 Riproduzione e sviluppo larvale 9
1.1.4 Alimentazione 11
1.1.5 Cenni sulla pesca di A.antennatus 11
1.1.6 Variabilità genetica all’interno della specie 14
1.2 Il DNA mitocondriale 17
1.3 Il Flusso genico 21
1.4 La filogeografia 22
2.
MATERIALI E METODI
232.1 Siti di campionamento e trattamento dei campioni 23
2.2 Le analisi genetiche 25
2.2.1 Estrazione DNA 25
2.2.2 Estazione chimica – protocollo “fenolo/cloroformio” 26
2.3 Sintesi di oligonucleotidi 29
2.4 Reazione di amplificazione e sequenziamento 31
2.5 Trattamento statistico dei dati 35
2.5.1. Allineamento delle sequenze 35
2.5.2. Stime di diversità genetica all’interno dei campioni 35 2.5.3. Stime di diversità genetica tra i campioni 36
3.
RISULTATI
413.1. Diversità genetica all’interno dei campioni 41
3.2. Diversità genetica tra i campioni 43
3.2.1. F-Statistica 43
3.2.2. Distanza genetica 45
3.2.3. Analisi della varianza molecolare (AMOVA) 46
3.2.4. Network 48
3.2.5. Stime di flusso genico 52
3.2.6. Isolamento da distanza 53
3.3. Aspetti di demografia storica 55
4.
DISCUSSIONE
604.1. Diversità genetica all’interno dei campioni 60
4.2. Struttura genetica 62
4.3. Aspetti di demografia storica 65
4.4. Gestione delle risorse 67
5.
CONCLUSIONI
696.
BIBLIOGRAFIA
71RIASSUNTO
Aristeus antennatus (Risso, 1816) è un crostaceo decapode appartenente alla
superfamiglia dei Penaeidae e alla famiglia degli Aristeidae. Questo gambero batiale (conosciuto anche come gambero viola o gambero imperiale) è distribuito prevalentemente su fondali fangosi, tra i 300 ed i 3300 metri di profondità, ed è una delle specie bersaglio più importanti della pesca profonda nell’intero Mediterraneo. Il suo prelievo viene effettuato mediante rete a strascico, a profondità generalmente comprese tra i 300 e gli 800 m.
Il presente studio, svolto presso il Laboratorio di Ittiologia genetica dell’Università di Gerona (Spagna), ha l’obiettivo di ottenere informazioni sulla struttura genetica della specie. Sono stati presi in considerazioni campioni provenienti da tre differenti zone (59 individui per il Mar Catalano, 44 per il Mar Ligure e 40 per il Mar Tirreno meridionale). L’analisi della diversità genetica di A. antennatus è stata effettuata utilizzando due marcatori molecolari mitocondriali: il gene per la subunità ribosomale 16S (16SrDNA) e quello per la subunità I della citocromo ossidasi (COI). Dopo aver estratto il DNA dal muscolo dell’addome, i geni in questione vengono amplificati mediante reazione a catena della polimerasi (PCR), utilizzando primer specifici. Successivamente vengono sequenziati. Le sequenze nucleotidiche ottenute sono state utilizzate per stimare la diversità genetica all’interno e tra i campioni, e per l’applicazione di analisi filogeografiche. Sono stati rilevati valori di diversità aplotipica medio-alti (COI: 0.34<h<0.59; 16SrDNA: 0.36<h<0.57) e di diversità nucleotidica bassi (COI: 0.0016<π<0.0038; 16SrDNA: 0.0008<π<0.0013). I pattern ottenuti con il median-joining network, la distribuzione “mismatch” e i test R2 per le
A.antennatus nell’area di studio. Le stime di flusso genico hanno fornito valori molto
elevati (Nm>10), suggerendo che la dispersione larvale è efficace anche a grandi distanze. I risultati sulla moderata strutturazione genetica della specie sono in accordo con un precedente studio effettuato con l’elettroforesi degli alloenzimi.
1.
INTRODUZIONE
Il presente lavoro, svolto nell’ambito del Progetto I+D (2006-2008) del Ministerio de Educaciòn y Ciencia (Spagna) “Gambas de aguas profundas del Mar Mediterraneo:
identificaciòn genética de las poblaciones mediante anàlisis de ADN e implicancia para la gestiòn de las pesquerias.”, ha come obiettivo dare delucidazioni riguardo la
differenziazione genetica tra le popolazioni del gambero A.antennatus nel Mar Mediterraneo Occidentale. Rifacendosi ad un precedente studio che rilevava la mancanza di strutturazione genica nella specie e l’assenza di una suddivisione geografica, riscontrata invece con le analisi morfologiche, si è voluto utilizzare tecniche più sensibili e diversi marcatori molecolari per comprendere se le differenze morfologiche osservate sono attribuibili a fattori non genetici ma ambientali.
1.1.Biologia ed ecologia di Aristeus antennatus
1.1.1 Inquadramento sistematico e riconoscimento
Fig.1.1. Esemplare di Aristeus Antennatus (Risso, 1816). Fonte: Internet. PHYLUM: Arthropoda CLASSE: Crustacea SUBCLASSE: Malacostraca ORDINE: Decapoda SUBORDORDINE: Natantia FAMILIA: Aristeidae GENERE: Aristeus SPECIE: A. antennatus
Il gambero imperiale, Aristeus antennatus, è stato descritto agli inizi del diciannovesimo secolo nel Mar Ligure da Risso, insieme ad Aristaeomorpha foliacea. Le due specie costituiscono gli unici rappresentanti della famiglia Aristeidae che si possono incontrare nel Mar Mediterraneo.
Fig.1.2. A) Fotografia di un campione di A.antennatus B) Illustrazione che evidenzia le caratteristiche morfologiche esterne
A. antennatus è un crostaceo di dimensioni medie attorno ai 12 cm, max 20-22 cm., con corpo compresso lateralmente, costituito da una parte anteriore (cefalotorace) ed una posteriore segmentata (addome). Il cefalotorace è ricoperto di una robusta corazza dotata di spine (carapace) ed è provvisto di 13 paia di appendici: un paio di antennule, uno di antenne, uno di mandibole, due paia di mascelle, cinque paia di arti per la locomozione (pereiopodi), di cui quattro terminano con una piccola pinza. Il cefalotorace è seguito dalla regione posteriore o addome, che è costituita da sei segmenti articolati, lisci ed intersecati longitudinalmente da una piega, di cui i primi cinque sono muniti ciascuno di un paio di appendici per il nuoto (pleopodi) ed il sesto è formato da appendici a lamelle (uropodi) e termina con un ventaglio (telson). L’esoscheletro leggero e i lunghi pleopodi suggeriscono che la specie sia un abile nuotatore (Cau et al., 2002 ). Il carapace è armato da
Spine rostrali pleopodi p e re op od i telson
un rostro munito nella parte superiore di tre denti. Il rostro presenta dimorfismo sessuale (più lungo nelle femmine e nei giovani), particolare che permette l’identificazione del sesso. I grossi occhi sono localizzati lateralmente su un peduncolo sotto il rostro e sormontano l'apparato boccale. La colorazione del corpo è rosso-chiara o rosea, con sfumature violacee nella parte superiore del carapace e lungo le giunture dei segmenti dell'addome.
Aristeus antennatus viene spesso commercializzato insieme con un'altra specie
denominata gambero rosso (Aristaeomorpha foliacea). I caratteri che permettono la distinzione delle due specie sono:
• il numero di denti nella parte superiore del rostro (tre in A. antennatus e cinque-sei in A. foliacea) (Fig. 1.3);
• la colorazione, più scura nel gambero rosso che possiede anche pereopodi più lunghi e sottili.
.
Fig.1.3. Differenze nella morfologia del rostro in A. foliacea e A. atennatus Fonte: internet (ftp://ftp.fao.org/)
1.1.2. Distribuzione geografica
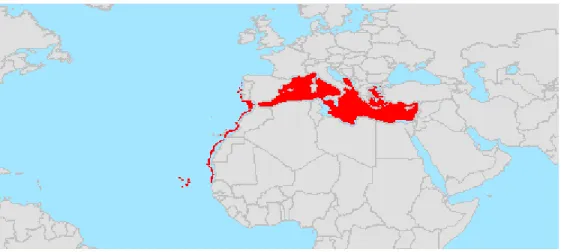
Aristeus antennatus è diffuso nell'intero bacino del Mediterraneo, ad eccezione
dell’alto e medio Adriatico ed è presente anche nell’Oceano Atlantico, dal Portogallo al Marocco, fino alle Isole di Capo Verde (Fig. 1.4). Si distribuisce su fondali fangosi tra i
300 ed i 3300 metri di profondità (Sardà et al., 2003). A. antennatus è una specie gregaria che vive in gruppi numerosi. Gli individui di questa specie effettuano spostamenti verticali da profondità minori (circa 200 m), dove sono presenti durante la notte, verso profondità maggiori (circa 800 m), dove sono presenti durante il giorno, suggerendo un comportamento bento–pelagico (Relini, 1981; Bianchini et al., 1998). La specie effettua, inoltre, migrazioni stagionali, stazionando in acque meno profonde nei periodi più freddi.
Fig 1.4. Distribuzione geografica di A. antennatus (www.fao.org)
1.1.3. Riproduzione e sviluppo larvale
A. antennatus è una specie gonocorica, le cui femmine raggiungono la maturità
sessuale in estate e producono notevoli quantità di uova. Alla schiusa le larve sono piatte, con lunghi arti e nuotano in superficie conducendo vita pelagica, per poi mutare più volte fino a diventare forme giovanili e insediarsi sul fondo. Qui adottano stile di vita bentonico e raggiungono, attraverso una serie di mute, il completo sviluppo. Lo sviluppo larvale è caratterizzato dalla presenza di quattro stadi larvali: nauplio, zoëa, mysis e post-larva. Lo stadio naupliare è caratterizzato dal possesso di tre paia di appendici che hanno una
funzione natatoria (Fig. 1.5). Il nauplio è privo di apparato boccale, non si alimenta, ma si accresce a carico delle riserve nutritive accumulate nel corpo. Nello stadio di zöea si ha la comparsa di occhi peduncolati, di un breve rostro, degli uropodi e delle mandibole che permettono alla larva di nutrirsi di fitoplancton (Fig. 1.6 A). Si passa poi allo stadio di mysis, in cui appaiono i pleopodi (Fig. 1.6 B) e cambia l’alimentazione che, da fitoplanctonica, diventa zooplanctonica. Con il passaggio a post-larva i pleopodi assumono in pieno la loro funzione natatoria ed i pereiopodi quella ambulacrale (Fig. 1.6 C). Dopo qualche mese di residenza nelle zone superficiali, gli adulti, che iniziano a maturare sessualmente, migrano nuovamente verso zone più profonde, completando il ciclo vitale. La durata media della vita per questa specie è stimata intorno agli 8-9 anni (Orsi Relini & Relini, 1998; Cau, 2002)
Fig.1.5. Sviluppo larvale: A) Nauplio precoce; B) Nauplio più avanzato; C) nauplio visto al microscopio c
Fig. 1.6. Sviluppo larvale A Prima protozoea B Prima mysis C Prima postlarva
1.1.4. Alimentazione
Il comportamento alimentare di A. antennatus è di tipo eurifago. Esso si nutre di detrito inorganico ed organico (piccoli organismi animali o vegetali morti od in decomposizione), ma assume anche comportamento predatorio. La dieta comprende, infatti, sia prede bentoniche, come policheti, echinodermi, piccoli bivalvi, gasteropodi e crostacei appartenenti a vari gruppi, sia organismi euribati del micro-necton, particolarmente crostacei eufausiacei e decapodi (Relini & Orsi Relini, 1987).
1.1.5. Cenni sulla pesca di A. antennatus
A. antennatus rappresenta una risorsa economicamente importante che caratterizza l’attività di pesca di molte marinerie mediterranee. La sua pesca, attuata con lo strascico di profondità, è spesso associata a quella del gambero rosso (Aristaeomorpha foliacea); le due specie costituiscono la categoria commerciale dei “gamberoni”. Il loro sfruttamento intensivo nel bacino mediterraneo occidentale è iniziato alla fine degli anni ‘50, grazie alla possibilità di congelare il pescato direttamente a bordo e all’introduzione di antiossidanti quali conservanti del pescato A livello internazionale e comunitario è considerato prioritario lo sfruttamento sostenibile delle risorse della pesca; in particolare, una delle problematiche prioritarie individuate dalla UE, è lo studio dello stato degli stock mediterranei di gamberi di profondità. Nonostante il gambero imperiale sia sottoposto ad un’elevata pressione di pesca, si ritiene che questa risorsa sia soggetta ad un tasso di prelievo sostenibile. Infatti, la possibilità di mantenere tali livelli di sfruttamento, sembrerebbe dovuta alla distribuzione estremamente ampia della specie sui fondali batiali (Bianchini & Ragonese, 1994) e alla vulnerabilità degli stock di A. antennatus che è solo “parziale”, poiché solo la frazione più superficiale dello stock è accessibile alla pesca commerciale (Demetre & Lleonard, 1993; Demestre & Martin, 1993; Sardà et al., 1994). I rendimenti di pesca sul gambero imperiale sono tuttavia soggetti a variazioni stagionali, annuali e a fluttuazioni geografiche. Registrazioni storiche, ma locali, indicano che in alcune zone queste risorse presentano notevoli fluttuazioni in termini di abbondanza dello stock. Ad oggi non si conosce precisamente lo stato degli stock di A. antennatus nel Mediterraneo; sono state effettuate valutazioni a livello regionale per, ma non esistono informazioni sullo stato generale.
Un incremento della taglia di prima cattura, mediante l’adozione di una maglia più selettiva, permetterebbe di ottenere in futuro incrementi del rendimento, portando quindi ad una condizione di sfruttamento meno gravosa senza ripercussioni economiche (Spedicato et
al., 1995; Ragonese & Bianchini, 1996). Inoltre viene consigliato un periodo di fermo
biologico, nel periodo fine estate – inizio autunno, per ridurre la pressione di pesca sulle reclute, più vulnerabili durante tale periodo (D’ Onghia et al., 1997).
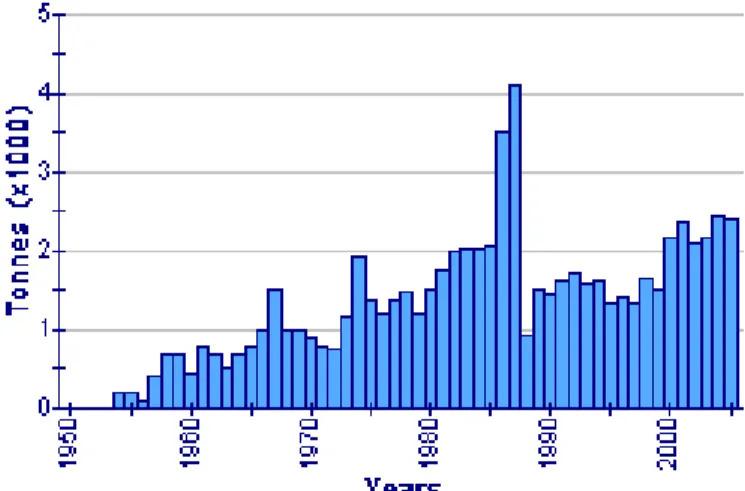
Fig. 1.7. Dati FAO relativi alle catture complessive globali di A. antennatus (in tonnellate) dal 1950 al 2005.
Sebbene negliultimi vent’anni siano state raccolte numerose informazioni su questa specie, molti aspetti bio-ecologici restano ancora sconosciuti. In particolare, si conosce molto poco sul reclutamento e non si riesce ancora a fornire un’adeguata spiegazione alle continue fluttuazioni di abbondanza che si registrano nelle varie zone di pesca. In realtà quasi tutte le conoscenze finora acquisite si riferiscono all’intervallo batimetrico 400-800 m, mentre, indagini realizzate nel Mediterraneo occidentale, al largo delle coste spagnole, hanno evidenziato, non soltanto la presenza di questo gambero fino a 3300 m di profondità, ma anche un possibile reclutamento al di sotto degli 800 m (Sardà et al., 2004). Pertanto, la
reale distribuzione batimetrica di questa specie è significativamente maggiore di quella in cui si realizza lo sfruttamento.
Sardà et al. (2003) suggeriscono che nel Mediterraneo la distribuzione di questa specie si può suddividere in due intervalli di profondità, con caratteristiche ben differenziate. Tra i 400 e gli 800 m, gli individui sono sottoposti ad un’alta pressione di pesca e la popolazione è composta prevalentemente da femmine adulte di grandi dimensioni (Tobar & Sardà, 1987; Demestre & Martin, 1993; Martinez-Baños, 1997). Tra gli 800 e i 3300 m di profondità, gli organismi dispongono di un “rifugio”, dove non c’è attività di pesca (“stock vergine”). In questa fascia batimetrica la densità diminuisce, gli organismi sono di piccola taglia e la sex-ratio cambia, con una proporzione 2:1 a favore dei maschi.
Questa specie si adatta all’energia disponibile alle differenti profondità, modificando la struttura della popolazione, la densità, la taglia degli individui ed anche concentrando il potenziale riproduttivo e il reclutamento a uno specifico intervallo di profondità (Sardà et al., 2002). Infatti, gli individui dei fondali soggetti a pesca, ricevono un input energetico (carbonio organico) cinque volte maggiore rispetto a quelli degli stock vergini. Questo però non influisce sul ritmo endogeno riproduttivo né sul tasso di crescita, che si mantengono a spese di una piccola taglia e di una minor densità. Quindi questa specie ha una grande plasticità adattativa, modificando il proprio ciclo vitale ai differenti habitat nei quali l’apporto energetico diminuisce con la profondità. Alcuni parametri come la riproduzione e la crescita possono essere simili alle diverse profondità, altri parametri come densità, dimensione media o sex-ratio possono variare notevolmente (Sardà et al., 2002).
1.1.6. Variabilità genetica all’interno della specie
La variabilità genetica rappresenta la base necessaria per i futuri cambiamenti evolutivi delle specie ed i livelli di diversità genetica nelle popolazioni possono fornire le prove di eventi evolutivi avvenuti in tempi passati più o meno recenti (Weir, 1990). Le modalità di ingresso di nuova variabilità genetica in una popolazione sono ritenute classicamente la mutazione, che agisce su scale di tempo relativamente grandi, ed il flusso genico, che può permettere l’ingresso di nuovi geni mediante l’incrocio degli individui nativi con gli immigranti (Hartl & Clark, 1993). Le popolazioni naturali di una determinata specie, che appartengono a regioni geograficamente distinte, presentano quasi sempre differenze nella distribuzione dei geni al loro interno, ciò indica la presenza di strutturazione genetica in quella specie. La struttura genetica attuale delle specie è il risultato di processi storici e delle forze di tipo microevolutivo (Hartl & Clark, 1993). La struttura genetica delle specie e, quindi, i meccanismi evolutivi e le strategie adattative da esse adottate, possono essere conosciuti analizzando la distribuzione dei geni all’interno delle popolazioni. Nonostante esistano vari metodi per valutare la struttura genetica di specie, questi forniranno informazioni solo su un piccolo campione, sia perché il numero di individui analizzati rispetto alla taglia della popolazione sarà limitato, sia perché l’informazione sarà limitata a quella fornita dai marcatori genetici utilizzati (Procaccini & Maltagliati, 2003). I dati ottenuti saranno quindi soggetti ad errore di campionamento per quanto riguarda i parametri di variabilità genetica in senso assoluto, ma saranno estremamente utili per analisi comparative tra le popolazioni. Le stime del grado di relazione dei geni, tra individui e tra popolazioni, possono essere valutate grazie all’esistenza di vari modelli teorici semplificativi che si fondano su assunzioni che
approssimano le caratteristiche genetiche delle popolazioni naturali. Il classico modello di Hardy-Weinberg, per esempio, è basato su una popolazione di taglia infinita e panmittica, con tasso di mutazione pari a zero ed assenza di fenomeni come migrazione e selezione. Una popolazione con caratteristiche simili è ovviamente inesistente in natura, ciò nonostante il modello di Hardy-Weinberg è tutt’oggi di grande utilità.
La distanza fisica esistente fra gli organismi può influenzare il livello dei rapporti genetici esistenti fra essi ed agire come fattore gerarchizzante. I pattern di diversità genetica osservati variano in rapporto ai cicli vitali ed alla capacità di dispersione delle specie: generalmente, specie con alte capacità di dispersione, tendono ad essere meno geneticamente strutturate, grazie all’azione omogeneizzante del flusso genico (Procaccini & Maltagliati, 2003). È da notare, comunque, che recentemente sono state trovate eccezioni a questa regola (Jones et al., 1999;; Swearer et al., 1999,; Weber et al., 2000; Goldson et al., 2001; Luttikhuizen et al., 2003; Taylor & Hellberg, 2003; Becker et al., 2007).
Gli studi di genetica di popolazione riguardo ad A. antennatus sono ad oggi scarsi e forniscono esigue informazioni sulla struttura genetica della specie. Le uniche conoscenze ci vengono fornite da Sardà et al. (1998) che hanno comparato i risultati ottenuti con l’elettroforesi degli alloenzimi con quelli di tipo morfometrico, in organismi provenienti da diverse aree del Mediterraneo. Analisi morfometriche multifattoriali indicano che alcuni caratteri morfologici variano con la longitudine. Le analisi elettroforetiche di campioni di muscolo ed epatopancreas hanno identificato 15 loci alloenzimatici, di cui sono risultati 10 monomorfici e i rimanenti 5 mostravano polimorfismo da basso a moderato. I test per verificare l’eterogeneità delle frequenze alleliche hanno rivelato bassa diversità genetica tra i campioni provenienti dalle diverse aree geografiche (Sardà et al., 1998). Ciò risulta in accordo con quanto già ottenuto in precedenza in specie di peneidi (Lester, 1979; Nelson &
Hedgecok, 1980; Tam & Chu, 1993). I bassi valori dell’indice di fissazione di Wright (FST
= 0.017) e della distanza genetica di Nei (D < 0.002) rilevati in quel lavoro, evidenziano una strutturazione in popolazioni molto bassa. Ciò suggerisce che la dispersione in questa specie, dovuta soprattutto alle forme larvali, tende a mantenere omogeneità genetica anche tra regioni geograficamente distanti. Questa analisi genetica, in contrapposizione agli studi morfometrici, rivela assenza di suddivisione geografica tra gli individui della specie.
1.2. Il DNA mitocondriale
Quasi tutte le cellule eucariotiche contengono un genoma mitocondriale, in aggiunta al genoma nucleare. In assenza di mutazioni, ciascuno eredita un mtDNA identico a quello della madre. Nella seconda generazione e in quelle successive, solo i discendenti per linea
materna condividono lo stesso DNA mitocondriale (mtDNA). Il mtDNA è rappresentato da
una singola molecola circolare, costituita da circa 16000 paia di basi (bp), è un genoma estremamente compatto, ed è costituito principalmente da regioni codificanti prive di introni e non separate tra loro da sequenze interposte. In ogni cellula sono contenute numerose copie di mtDNA e generalmente queste copie sono identiche (omoplasmiche).
Dato il gran numero di mitocondri in una cellula, si può presentare il caso in cui una popolazione di mitocondri abbia una sequenza differente. Tale condizione, in cui si ha la presenza di basi differenti in una medesima posizione, è conosciuta come eteroplasmia.
L’eteroplasmia può derivare da nuove mutazioni insorte a livello del mtDNA, dal “pool” di mitocondri presenti nella cellula uovo da cui deriva l’individuo (Moritz, 1987), in alcune specie può essere, invece, la conseguenza di una trasmissione biparentale del mtDNA (Zouros, 1992). Molti studi dimostrano che il mtDNA è più soggetto a mutazione di quello nucleare, anche perché più esposto ad agenti mutageni: il mitocondrio è la sede della fosforilazione ossidativa ed è quindi a contatto con i radicali liberi dell’ossigeno prodotti durante alcune reazioni (Avise, 2000). Da sottolineare è anche il fatto che il mtDNA non è circondato da particolari membrane, ma si trova libero all’interno del mitocondrio, strettamente associato alla membrana mitocondriale interna (Wallace, 1994). Nonostante l’esposizione diretta ai contaminanti, il mtDNA possiede limitate capacità di riparazione rispetto al DNA nucleare. Inoltre va incontro a molti più cicli di replicazione, ed ha quindi
una maggiore probabilità di accumulare errori dovuti all’inserzione di basi scorrette durante i processi duplicativi, visto anche che la DNA-polimerasi mitocondriale non possiede l’attività esonucleasica 3’-5’. La frequenza di errore della DNA-polimerasi mitocondriale è da tre a cinque ordini di grandezza superiore rispetto agli errori commessi dalla DNA-polimerasi nucleare, che è provvista della subunità che svolge la funzione di correzione di bozze (Cooper et al., 1995). Rispetto al genoma nucleare, quello mitocondriale è molto meno esteso, però è presente in molte più copie per cellula, tanto che il numero di copie di mtDNA presente in un’unica cellula può essere dell’ordine delle migliaia. Dato questo enorme numero di geni mitocondriali presenti in ogni cellula, ci si potrebbe aspettare che, una mutazione che insorga in una singola molecola di mtDNA, abbia scarsa probabilità di essere fissata, e di conseguenza il tasso di mutazione sia molto basso; in realtà il genoma mitocondriale possiede un elevato indice di fissazione. Una possibile spiegazione per l’elevato tasso di fissazione è che si verifichi una sorta di collo di bottiglia nelle popolazioni di mtDNA, mediante il quale viene selezionata solo una parte delle molecole di mtDNA destinate ad essere successivamente duplicate (Lightowlers et al., 1997). La combinazione tra instabilità del mtDNA e un elevato indice di fissazione fa sì che il tasso di mutazione del mtDNA sia molto elevato: circa dieci volte maggiore di quello riscontrabile in sequenze equivalenti del genoma nucleare (Brown et al., 1979). Di norma le regioni codificanti hanno tassi di mutazione più bassi, mentre le regioni non codificanti presentano tassi di mutazione più alti. Il differente tasso di mutazione delle varie regioni permette di usare il mtDNA per l’analisi della diversità genetica sia a livello intra- che inter-specifico (Procaccini & Maltagliati, 2003). Il mtDNA è comunemente usato come marcatore molecolare per la sua versatilità ed affidabilità, sia nei vertebrati che negli invertebrati. Non bisogna, comunque, dimenticare alcuni inconvenienti che i marcatori mitocondriali possono
presentare, quali ad esempio, l’eteroplasmia, o la presenza di pseudogeni, cioè inserzioni di geni mitocondriali all’interno del genoma nucleare (Zhang & Hewitt, 1996). La presenza di geni mitocondriali nel genoma nucleare e la loro successiva amplificazione nelle analisi di studi filogeografici e di genetica di popolazioni possono alterare sostanzialmente i risultati e condurre a conclusioni errate. L’mtDNA è abbondantemente utilizzato per studi sulla struttura di popolazione, filogeografia, e sulle relazioni filogenetiche ai vari livelli tassonomici (Wilson et al., 1985; Moore, 1995; Avise, 2000).
L’uso di tecnologie avanzate nel sequenziare il mtDNA ha permesso di determinare sequenze complete di diversi taxa. Per l’ordine dei decapodi, il gruppo più diversificato dei crostacei, comprendenti più di 10 000 specie, con 120 famiglie e 1200 generi, vengono comunemente utilizzati la subunità I del citocromo c ossidasi (COI) e la subunità grande del RNA ribosomiale (16SrDNA), più di tutti gli altri geni mitocondriali, apparentemente per l’“universalità“ dei primer utilizzati nella reazione di sequenziamento (Folmer et al., 1994, Simon et al., 1994; Kambhampati & Smith, 1995). Esistono, però, poche sequenze di mtDNA complete di crostacei, tra cui quella di Penaeus monodon (GENBANK AF 217843) (Wilson et al., 2000) che fornisce le sequenze di riferimento per lo studio del genoma dei decapodi e quindi anche di A. antennatus.
Fig.1.8. Organizzazione generale del genoma mitocondriale di P. monodon (Wilson et al., 2000). Il gene COI viene indicato come COX1, l’srRNA come rns
1.3. Flusso genico
Si intende come flusso genico ogni scambio genetico fra popolazioni: quindi migrazione o dispersione dei gameti, embrioni, larve, adulti ed anche espansione di popolazioni in località precedentemente non occupate. Ovviamente affinché vi sia flusso genico non è sufficiente l’evento migrazionale: gli individui migranti devono anche riprodursi. Il flusso genico può considerarsi uno dei fattori microevolutivi più importanti che agiscono sulla struttura genetica di una specie, definendo il grado di connettività tra le popolazioni che la compongono. La migrazione degli individui da una popolazione a un’altra ha come risultato l’introduzione di nuovi alleli in una popolazione e/o il cambiamento delle frequenze alleliche. Nel caso in cui il fenomeno di migrazione sia ricorrente, il flusso genico rappresenta un elemento che diminuisce la diversità tra le popolazioni. Invece nel caso di flusso migratorio episodico che si verifica tra popolazioni normalmente separate, la migrazione viene considerata come sorgente di variabilità genetica. L’isolamento geografico ed il flusso genico tra popolazioni determinano il potenziale per la differenziazione genetica ed in ultima analisi, per la speciazione. Le stime di flusso genico possono essere ottenute tramite metodi diretti o indiretti (Slatkin, 1985): i primi consistono nei metodi di marcatura, rilascio e ricattura, utili per stimare gli spostamenti degli stadi adulti; le misure indirette sono, invece, basate sull’uso di parametri relativi alla struttura genetica della popolazione, permettendo di indagare gli eventi di flusso genico che hanno originato la variabilità genetica osservata.
1.4. La filogeografia
La filogeografia è la disciplina che riguarda i principi e i processi che governano la distribuzione geografica delle linee genealogiche, specialmente quelle all’interno delle specie, oppure quelle tra specie molto affini (Avise, 2000). L'analisi filogeografica contribuisce alla conoscenza della biodiversità e consente di ricostruire la storia delle popolazioni che si sono differenziate. La filogeografia è una disciplina che si posiziona a cavallo tra i fenomeni macroevolutivi (come la speciazione) e i processi microevolutivi (come flusso genico, mutazione e selezione naturale); essa tratta le componenti storiche e filogenetiche della distribuzione spaziale delle linee genealogiche. In altri termini, il tempo e lo spazio sono considerati in maniera unitaria ed in questo spazio ideale vengono riportate le linee genealogiche oggetto di studio (Avise, 2000). Nelle popolazioni naturali, la maggior parte degli aplotipi esistenti, sono presenti in molte copie identiche, originatesi per replicazione del DNA mitocondriale (mtDNA) e successiva trasmissione matrilineare. Quando una di queste copie subisce una mutazione, originando un nuovo aplotipo, è estremamente improbabile che le altre copie mutino a loro volta, determinando la scomparsa dell’aplotipo ancestrale; quindi ci si aspetta che l’aplotipo ancestrale sia ancora rappresentato nella popolazione, e possa essere campionato insieme agli aplotipi che ne derivano. I tradizionali metodi filogenetici sono basati su alberi che si dividono in maniera dicotomica, ma la persistenza dell’aplotipo ancestrale, contemporaneamente alla presenza di molti aplotipi da esso derivanti, produce ramificazioni multiple non rappresentabili come semplici biforcazioni. Detto ciò, si può prevedere che più antico sarà un aplotipo, maggiore sarà il numero di aplotipi derivati ad esso connessi. I processi evolutivi che intervengono a livello di popolazione, come la ricombinazione tra geni, l’ibridazione di differenti linee
genealogiche e l’omoplasia, generano relazioni di tipo reticolato che non sono ammesse dai metodi tradizionali e possono essere male interpretate (Posada & Crandall, 2001).