ContentslistsavailableatScienceDirect
Comptes
Rendus
Palevol
ww w . sc i e n c e d i r e c t . c o m
General
Palaeontology,
Systematics
and
Evolution
(Vertebrate
Palaeontology)
Avian
fossil
assemblages
at
the
onset
of
the
LGM
in
the
eastern
Alps:
A
palaecological
contribution
from
the
Rio
Secco
Cave
(Italy)
Assemblages
fossiles
d’oiseaux
au
commencement
du
dernier
maximum
glaciaire
dans
les
Alpes
oreintales
:
une
contribution
paléo-écologique
de
la
grotte
du
Rio
Secco
(Italie)
Lisa
Carrera
a,
Marco
Pavia
b,
Matteo
Romandini
c,a,
Marco
Peresani
a,∗aUniversitàdiFerrara,DipartimentodiStudiUmanistici,SezionediPreistoriaeAntropologia,CorsoErcoleId’Este,32,44121Ferrara,
Italy
bUniversitàdegliStudidiTorino,MuseodiGeologiaePaleontologia,ViaValpergaCaluso,35,10125Torino,Italy cUniversitàdegliStudidiBologna,DipartimentodiBeniCulturali,ViadegliAriani,1,48121Ravenna,Italy
a
r
t
i
c
l
e
i
n
f
o
Articlehistory: Received9August2017
Acceptedafterrevision24October2017 Availableonline22February2018 Keywords:
Birds LatePleistocene LastGlacialMaximum Easternalps Italy Systematics Palaeoecology RioSeccocave
a
b
s
t
r
a
c
t
TheavianfossilassemblagesfromthelatePleistocenedepositsoftheRioSeccoCave (north-easternItaly)ispresentedherein.WestudiedthelayersthatdatebacktotheendofMIS3 andthebeginningofMIS2,whichalsocontainevidenceofGravettianfrequentationdated to33.5–30kacalBP.Thesystematicanalysisrevealedthepresenceof18speciesandother supraspecifictaxathatsupportedpalaeoenvironmentalreconstructions.Taxaindicatethat, attheonsetofLGM,sitesurroundingswerecharacterisedbyconiferormixedforests,open grasslands,slow-flowingwaterbodiesandmountainmeadowswithrockyoutcrops,as indicatedbyLagopusmuta.Today,thisenvironmentisfoundabovethetreeline(beyond 1500–2000m)andcannotbedetectednearthesite,locatedat580masl.Noteworthy,is alsothefindingofthesecondItalianlatePleistocenefossilrecordofPicuscanus.
©2017Acad ´emiedessciences.PublishedbyElsevierMassonSAS.Allrightsreserved.
Motsclés: Oiseaux
Pléistocènesupérieur Derniermaximumglaciaire Alpesorientales
Systématique Paléo-écologie GrotteduRioSecco
r
é
s
u
m
é
Lesassemblagesfossilesd’oiseauxdesdépôtsduPléistocènesupérieurdelagrottede RioSecco(Nord-Estdel’Italie) sontici présentés. Nousavons étudiélesniveauxqui remontentàlafinduMIS3etaudébutduMIS2,etquicontiennentaussidespreuves d’unefréquentationgravettiennedatéede33,5–30kacal.L’analysesystématiquerévèlela présencede18espècesetd’autrestaxonssupra-spécifiques,quipermettentdes reconsti-tutionspaléoenvironnementales.Lestaxonsindiquentqu’audébutduderniermaximum glaciaire,lesalentoursdu siteétaientcaractériséspardesforêtsmixtesoudesforêts
∗ Correspondingauthor.
E-mail addresses: [email protected] (L. Carrera), [email protected] (M. Pavia), [email protected] (M. Romandini), [email protected](M.Peresani).
https://doi.org/10.1016/j.crpv.2017.10.006
deconifères,desprairiesouvertes,descoursd’eauàfaibleintensitéetdesprairiesde mon-tagneàaffleurementsrocheux,commel’indiquelaprésencedeLagopusmuta.Actuellement, cetenvironnements’observeau-dessusdelalignedesarbres(au-dessusde1500–2000m) etnepeutêtretrouvéprèsdusite,localiséà580mau-dessusduniveaudelamer.Ànoter qu’ilestaussilelieudeladécouvertedusecondenregistrementfossileitalienauPléistocène supérieurdePicuscanus.
©2017Acad ´emiedessciences.Publi ´eparElsevierMassonSAS.Tousdroitsr ´eserv ´es.
1. Introduction
Geographic distributionandphenology ofbirds have changedmanytimesinthepast,duetoenvironmental, cli-matic,ecologicaland,recently,alsoanthropicfactors,in adynamicprocesswhichisstillunderway(Brudererand Salewski,2008;Gordo,2007;HolmandSvenning,2014; Jonzénet al., 2006).Asbird species areoften linkedto particularhabitats,especiallyconcerningvegetation,fossil avifaunasgiveimportantinformationsabout palaeoenvi-ronmentandpalaeoclimate.Someauthorsevenconsider birdsbetterpaleoenvironmentindicatorsthan micromam-mals(BedettiandPavia,2013;Serjeantson,2009).Despite this, contributionswhich dealwithavianfossil remains have so far been few, and this field seems to receive lessattentionthanthestudyofmammalfossilremains, maybebecauseinmanyfossilassemblagesthemammal remainsoutnumberthebirdremains,andalsobecauseof thescarcityofbirdskeletalcomparativecollections.
In this work, we present the systematic analysis of avifaunalremainsoftheRioSeccoCave,fromlayers dat-ing to the onset of MIS2. In Italy, several sites yielded avian remains from this period, but just a few in the areaof thenorthern belt ofthe exposed Great Adriatic Plain,wheretheRioSeccoCaveislocated(Fig.1).Some of them lack clear quantitative or stratigraphic indica-tions(Bonetal.,1991;Tyrberg,1998);amongsttheones provided withdatingsand a reliablestratigraphy, there areGrottadiFumane(CassoliandTagliacozzo,1994)and Riparo delBroion(Gurioliet al.,2006).Thelayers D1d-D1e of the Fumane cave deposit, which date back to 31,75–31,43ka 14C BP (Higham et al., 2009), provided, amongtheotherbirdtaxa,Buboscandiacusremains.The presenceofthisspeciesindicatesacolderclimate.Riparo del Broionyielded from layers dated 30–32 14C ka BP (ABA), birdremains that indicatethe presenceof slow-flowingwaterbodies.During theLastGlacialMaximum, birdremainsfromSandaljaII(southernIstria,Croatia)also indicatecolderclimateconditions,likeLagopuslagopusand Buboscandiacus (Tyrberg, 1998).Similartaxa related to colderclimaticconditions,includingBuboscandiacus,have alsobeenfoundintheCovolodiTreneinthesameperiod (RomandiniandNannini,2012;Tonon,1977).Duringthe coldphasesoftheLGM,theItalianpeninsula,asotherparts ofMediterraneanEurope(SpainandBalkans)represented refugia for northern species, which moved southwards afterglacialexpansions(Pellegrinoetal.,2014;Sanchez Marco,2004).Furthermore,thenumberofbirdtaxafrom openenvironments(steppe,alpine meadows,etc.), sug-gestsanexpansionofthesehabitatsincoldstages.Even
severalLateglacialsitesof north-easternItaly and Croa-tiayieldedavianremainsthathavebeenstudiedandhave providednumericalortaxonomicdataGrottadiVejaE,in theVenetoregion;RiparoSoman,RiparoDalmerieRiparo Cogola intheTrentino region;GrotteVerdidi Pradisin theFriuliregion;SandaljaIIeLjubicevapecinainCroazia (Fioreetal.,2015;GalaandTagliacozzo,2009,2010;Oros Srˇsenetal.,2014;Tonon,1983).Ljubicevapecina,in south-ernIstria,providedLagopuslagopusremains,indicatingthe permanenceinthisarea,intheearlystagesofLateglacial,of speciesofcoldenvironments(OrosSrˇsenetal.,2014).The aviantaxaidentifiedfromtheothersites,whosedeposits datebacktotheendofLateglacial,attheHolocene bound-ary,mostlycorrespondtothecurrentavifaunaofthearea.
2. Siteandstratigraphy
TheRioSeccoCaveislocatedatanelevationof580m aslonthePradisPlateauintheeasternpartoftheCarnic PreAlps(Fig.2),anorographicsystemoftheeasternAlps dissectedfromnorth–southandwest–eastorientedvalleys thatseparatemountainswithpeaks2000–2300masl.The PradisPlateauis6sqkm wideandisenclosedonthree sidesbymountainspeakingfrom1148mto1369mand tothesouthitisboundedbythefoothillsdissectedbythe Cosastream.Theplateaurisesfrom530mto590mand facesthesouthtothepresent-dayFriulianplain,which istheuppermostbeltofalargeplain,theGreatAdriatic Plain,whichemergedduringthelatePleistoceneandhad itsmaximalsouthernexpansionduringtheLGM(Antonioli andVai,2004;Shackletonetal.,1984).Theshelter pro-tectsahorizontalgallery,whichisalmostcompletelyfilled withsediments.In2002,atest-pitrevealedthepresenceof prehistoricsettlements.Anarchaeologicalexcavationhas beencarriedoutsince2010(Peresanietal.,2014).
The sedimentarybody of thecave includes4 strati-graphicMacro-Units,partiallyaffectedbybioturbation:
• Macro-Unit 1: includes US2 (Unità Stratigrafica-Stratigraphic Unit 2) and US3. Contains historical settlements and Neolithic/Bronze Age archaeological material;
• Macro-UnitBR1:includesUS4andUS6andis character-izedbythepresenceof angular stonesandfragments of karst limestone pavement originated by vault col-lapses. US4is composedof blocks and stones witha loamyfinefractionandcontainsrarelithics.US6isathin anddiscontinuouslayerwithorganicmatterand micro-charcoal,rarebones,lithicimplementsandtwohearths.
Fig.1. MapofthenorthernAdriaticregionwiththepositionofRioSeccoCaveintheblacksquare. Fig.1. CartedelarégionAdriatiquenord,aveclapositiondelagrottedeRioSeccodansl’encart.
Fig.2. ViewofthePradisPlateauwiththepositionofRioSeccoCaveinthegorge,markedwiththearrow.Inthebackground,thealluvialplainwiththe TagliamentoRiverinthecenter.
Fig.2. VueduplateaudePradis,aveclapositiondelagrottedeRioSeccodanslagorge,indiquéeparlaflèche.Àl’arrière-plan,laplainealluviale,avecla rivièreTagliamentoaucentre.
Apreliminaryanalysisofmammalremainsrevealedthe presenceofCapraibex,Rupicaprarupicapra,Bos primige-nius/BisonpriscusandCastorfiber.Humanmodifications includecut-marksandcombustiononcapridshafts,ona marmotclavicleandonaCastorfiberscapula(Peresani etal.,2014).Charredwoodfromoneofthesehearths yieldedanageof33,480–30,020ycalBP,assessingthe placementofUS6intheearlyGravettianperiod(Talamo etal.,2014).Macro-UnitBR1probablycoversatimespan ofafewmillenniabetweentheendofMIS3andthe begin-ningofMIS2,inacontextofprogressivelyharsherclimate thatpeakedduringtheLGM(Clarketal.,2009;Monegato etal.,2017);
• Macro-UnitBR2:isamassivestone-supportedbreccia.It includesUS4BRandadiscontinuoussandylayeratthe base,labeledUS4sabbie.ThisMacro-Unitis archaeologi-callysterile;
• Macro-Unit BIO1: is composed of stones and loamy finefractionandcontainssomeMousterian archaeolog-ical material.It includes thelayers US5top,US7,US5, US8 and US9. The age of these deposits ranges from 47,940–45,840cal BP(US5top)tomore than 49,000y BP(lowerlevelofUS7)(Talamoetal.,2014).The mam-mal remains from these layers belong to ungulates (Cervuselaphus,Capreoluscapreolus,cf.Alcesalces, Rup-icaprarupicapra,Capraibex,Susscrofa,Bosprimigenius/
Bisonpriscus),carnivores(Ursusspelaeus,Martesmartes, Mustelaerminea,Melesmeles,Vulpesvulpes,Canislupus) and to Erinaceus europaeus and Lepus europaeus. The remainsofCervuselaphusandUrsusspelaeusshowhuman modifications(Peresanietal.,2014;Romandinietal.,in press).Amongthebirdremains,aclawofAquila chrysae-tosshowcut-marks(Romandinietal.,2014).
3. Materialsandmethods
Inthiswork,wepresentthesystematicanalysisof194 bird remains comingfromMacro-Unit BR1 and Macro-UnitBR2.Allsedimentswereexcavatedin50×50cmor 33×33cm squares and in some cases (e.g., US4), they wereremovedinarbitrarylevels.Birdremainshavebeen recovered after sediments wet sieving. The amount of processed sediment isapproximately57 m3 (39 m3 for
BR1and 18m3 forBR2).Thefossilmaterialisstored in
the Sezione diScienze Preistoriche e Antropologiche of theDipartimentodiStudiUmanisticioftheUniversityof Ferrara.Thetaxonomicdeterminationshavebeenbasedon comparisonswithtwomodernbirdskeletalcomparative collections: the “Marco Pavia Ornithological Collection” (MPOC),storedattheDipartimentodiScienzedellaTerra oftheTorinoUniversity,and theone heldatSezionedi ScienzePreistoricheeAntropologicheoftheDipartimento di Studi Umanistici of the University of Ferrara. Some specific contributions on the osteological features of single avian orders and families have also been very helpfulforthetaxonomicdeterminations (Cuisin, 1989; Erbersdobler,1968;Janossy,1983;Kraft,1972;Otto,1981; Schmidt-Burger, 1982; Tomek& Bochenski, 2000). Tax-onomyfollowsDelHoyoetal.(2014,2016).Osteological terminologyfollowsBaumelandWitmer(1993).
4. Taxonomicaldeterminationandecological background
Atotalof166remainshavebeenassignedto31bird taxa (Table1).Amongthese,severalremains havebeen attributedtospecies-leveltaxaandaredescribedindetail hereafter, while other remains have beenattributed to supraspecific taxa, because of their fragmentation or becauseofthelackofdiagnosticfeatures.Thelatterare notdescribedindetail.Finally,28remainsareleftasAves.
ClassAves OrderGalliformes FamilyPhasianidae GenusCoturnix
CoturnixcoturnixLinnaeus,1758
Material.Oneproximalrightcoracoid(RS320)fromBR1. Descriptionandcomparisons.Coturnixcoturnixbones differfromthoseofallotherwesternPalearcticGalliformes insize,beingclearlysmallerthantheotherspecies(Kraft, 1972).
Remarks. Coturnix coturnix currently breeds in most of Europeand winters in sub-saharan Africa(Del Hoyo etal.,2014).InItaly,itisabreeding,regularmigrantand
Table1
ListofbirdtaxafromRioSeccoCave.Thenumbersindicatetheidentified remainsforeachtaxon.
Tableau1
Listedestaxonsd’oiseaudelaGrottedeRioSecco.Lesnumérosindiquent lesrestesidentifiéspourchaquetaxon.
Taxa BR1 BR2 Coturnixcoturnix 1 Perdixperdix 1 1 Lagopusmuta 3 1 Tetraourogallus 1 2 Lyrurustetrix 6 1
Tetraourogallus/Lyrurustetrix 1 1
Galliformesindet. 10 2 Crexcrex 2 Fulicaatra 1 Rallidaeindet. 3 Asiootus 2 Asioflammeus 2 Strigidaeindet. 1 Buteobuteo 2 Picuscanus 1 Falcosp. 1 Picapica 1 Nucifragacaryocacactes 1 Corvidaeindet. 8 3 Paridaeindet. 1 Alaudidaeindet. 1 cf.Turdusviscivorus 2 Turdussp. 2 1 Erithacusrubecula 1 1 Muscicapidaeindet. 1 Prunellacollaris 1 Pyrrhulapyrrhula 1 1 Loxiacurvirostra 1 Fringillacoelebs/montifringilla 1 Emberizidaeindet. 1 Passeriformesindet. 60 31 Avesindet. 23 5 Totalremains 139 55
rarewinteringspecies.Itbreedsinopenareaswithlow
vegetation (Brichetti and Fracasso, 2004). This species
isreported inthePalearcticsince theearlyPleistocene, and becomes abundant in the fossil record of the late PleistoceneofEurope(Tyrberg,1998,2017).
GenusPerdix
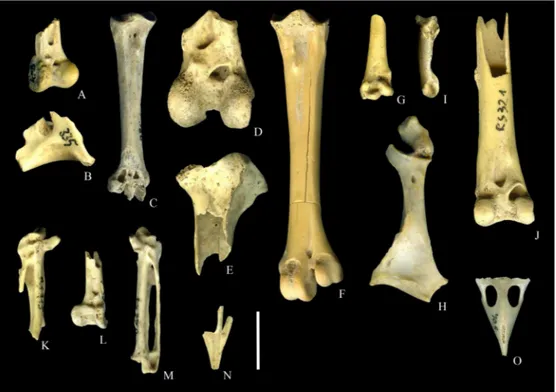
PerdixperdixLinnaeus,1758(Fig.3A)
Material.Onedistalrighttibiotarsus(RS403)fromBR1 andonedistalrightulna(RS375)fromBR2.
Descriptionandcomparisons.Thedistaltibiotarsus dif-fersfromtheoneofthegeneraAlectoris,LagopusandBonasa inthewidthoftheepiphysis(smallerthaninLagopus lago-pus,AlectorisrufaandAlectorisgreca)andintheextentof theponssupratendineum,whichisclearlywiderthanin BonasabonasiaandLagopusmuta.Theulnais distinguish-ablefromthatoftheabove-mentionedgenerainbeingless widedistally(Kraft,1972).
Remarks.Perdixperdixisasedentaryspecies,whichis spreadacrossmostofEurope,partlybecauseof reintroduc-tions,alsoforhuntingpurposes(DelHoyoetal.,2014).In Italy,itiscurrentlyasedentaryandrarebreedingspecies, whichwasmuchmorewidespreadinhistoricaltimes.It livesinopengrasslandsandshrublands,preferablyhilly (Brichetti andFracasso,2004).Perdixperdixis knownin
Fig.3.BirdremainsfromthelatePleistocenedepositsofRioSeccocave.A.Perdixperdixdistalrighttibiotarsus(RS403),cranialview.B.Lagopusmuta distalleftcoracoid(RS335),dorsalview;C.Lagopusmutarighttarsometatarsus(RS380),dorsalview;D.Tetraourogallusdistalrighttibiotarsus(RS412) cranialview;E.Lyrurustetrixproximallefttibiotarsus(RS352),cranialview.F.Lyrurustetrixlefttarsometatarsus(RS324),dorsalview.G.Crexcrexdistal lefthumerus(RS452),cranialview.H.Fulicaatrarightcoracoid(RS378),dorsalview.I.AsioflammeusleftphalanxIofdigit1(RS455),lateralview.J.Buteo buteodistalrighttibiotarsus(RS321),cranialview.K.Picuscanusproximalleftcarpometacarpus(RS315),ventralview.L.Nucifragacaryocatactesdistalleft tibiotarsus(RS325),cranialview.M.cf.Turdusviscivorusrightcarpometacarpus(RS356),ventralview.N.Prunellacollarismaxilla(RS390),ventralview. O.Loxiacurvirostramaxilla(RS504),dorsalview.Thescalebarrepresentsonecentimetre.
Fig.3. Restesd’oiseaudesdépôtsPléistocènesupérieurdelagrottedeRioSecco.A.Tibiotarsusdroitdistal(RS403)dePerdixperdix,vuecrâniale.B.Coracoïde gauchedistal(RS335)deLagopusmuta,vuedorsale.C.Tarsometatarsusdroit(RS380)deLagopusmuta,vuedorsale.D.Tibiotarsusdroitdistal(RS412)de Tetraourogallus,vuecrâniale.E.Tibiotarsusgauchedistal(RS352)deLyrurustetris,vuecrâniale.F.Tarsometatarsusgauche(RS324)deLyurustetrix,vue dorsale.G.Humérusgauchedistal(RS452)deCrexcrex,vuecrânienne.H.Coracoïdedroit(RS378)deFulicaarta,vuedorsale.I.PhalangegaucheIdudoigt1 (RS455)d’Asioflammeus,vuelatérale.J.Tibiotarsusdroitdistal(RS321)deButeobuteo,vuecrâniale.K.Carpometacarpusgaucheproximal(RS315)dePicus canus,vueventrale.L.Tibiotarsusgauchedistal(RS325)deNucifragacaryocatactes,vuecrâniale.M.Carpometacarpusdroit(RS356)decf.Turdusviscivorus, vueventrale.N.Maxillaire(RS504)dePrunellacollaris,vueventrale.O.Maxillaire(RS504)deLoxiacurvirostra,vuedorsale.Labarred’échellereprésente 1cm.
thePalearcticsincetheearlyPleistocene,andiscommonly reportedinthelatePleistoceneofEurope,includingItaly (Tyrberg,1998,2017).
GenusLagopus
LagopusmutaMontin,1781(Fig.3BandC)
Material.Onecranialportionofaleftscapula(RS348), onedistalleftcoracoid(RS335)andoneshaftofarightulna (RS368)fromBR1andonerighttarsometatarsus(RS380) fromBR2.
Descriptionandcomparisons.Onaverage,Lagopusmuta isslightlysmallerthanLagopuslagopus.Thedistal cora-coidisdimensionallycompatiblewithbothspecies,butthe angulusmedialis,whichismorehook-shapedinLagopus mutathaninLagopuslagopus,allowsustoruleoutthislast species.Thetarsometatarsusdiffersfromtheoneof Lago-puslagopusintheclearlysmallerlength(Kraft,1972).We tentativelyreferthecranialportionofthescapulaandthe ulnashafttoL.muta(eveniftheseelementsarenot diag-nosticenoughtodistinguishitfromL.lagopus),onthebasis
ofahighmorphologicalcorrespondencewiththeformer species.
Remarks.Lagopusmuta,amostlysedentaryspecies,is distributedacrosstherockytundraareasofhighlatitudes and in the mountain areas of the mid-latitudes of the boreal hemisphere (Del Hoyo et al., 2014). In Italy, it currently lives in theAlpsas a sedentaryand breeding species; here, it prefers meadows with some creeping shrubsandrockygrounds,abovethetreeline.Itismostly observedat heightsbetween2,300and 2,700masl,but with a minimum of 1,550 and a maximum of 2,900m asl,asthisspeciesmakesaltitudinalseasonalmovements (Brichetti andFracasso, 2004).Thisspecies is knownin thePalearcticsince themiddle Pleistoceneand is often reportedfromlatePleistocenelocalities.Inthecoldphases of the Pleistocene, it was much more widespread,also at lower heights than today (Tyrberg, 1998, 2017). It currentlysurvivesonthemountainareasofmid-latitudes as a“glacial relict”(Holmand Svenning,2014; Tyrberg, 1991).
GenusTetrao
TetraourogallusLinnaeus,1758(Fig.3D)
Material.Onedistalrighttibiotarsus(RS412)fromBR1, one sternum fragment(RS387)and one distal posterior phalanx(RS506)fromBR2.
Description and comparisons. The size of the distal tibiotarsusfragmentandthesternumfragment,whichare bigger thanthat ofotherlargeGalliformes asPhasianus colchicus and Lyrurustetrix, makes theseremains easily identifiable asTetraourogallus (Erbersdobler, 1968).We tentativelyrefertheposteriorphalanx,whichshows fea-turestypicalofGalliformes(roundeddistalepiphysisand curvedandshortoutline),toTetraourogallusonthebasis ofitssize.
Remarks.Tetraourogallus,amostlysedentaryspecies,is spreadacrossthenorthernEurasiaandpartofthemedium latitudesofEurope(DelHoyoetal.,2014).InItaly,where it iscurrentlya sedentaryand breedingspecies,it lives in conifer or mixed forests of the central-eastern Alps (BrichettiandFracasso,2004).Inthehighlatitudesofthe borealhemisphere,itisobservedatlowerheights(Cramp, 1998).AlsoduringthelatePleistocene,asthefossilrecord indicates, thisspecies wasspread atlowerheights than today (Holm and Svenning, 2014). Tetrao urogallus is reportedinthePalearcticsincethemiddlePleistoceneand is oftenrecordedin thelatePleistocene(Tyrberg,1998, 2017).
GenusLyrurus
LyrurustetrixLinnaeus,1758(Fig.3EandF)
Material.Onefurculafragment(RS345),onecranial por-tion of a left scapula (RS337), onedistal rightcoracoid (RS405),oneproximallefttibiotarsus(RS352),one prox-imalrighttibiotarsus(RS369)andonelefttarsometatarsus (RS324)fromBR1.Oneproximalrightfemur(RS486)from BR2.
Descriptionandcomparisons.Thisspeciesislargerthan thoseinthegeneraAlectoris,Lagopus,BonasaandPerdix, butsmallerthaninTetraourogallus.Thebonesdifferfrom this speciesandfromPhasianuscolchicusalsoinseveral morphologicalfeatures (Erbersdobler, 1968).In thecase oftheproximalfemur,thesizeoftheepiphysisallowsus toruleoutTetraourogallus(Erbersdobler,1968)andthe extent andshape of thepneumatic foramina underthe cristatrochanteris,togetherwiththeoutlineofthe epiph-ysisinproximalview,ruleoutPhasianuscolchicus.Thetwo proximaltibiotarsicanbeeasilyseparatedfromTetrao uro-gallusbysize(Erbersdobler,1968);theseelementsdiffer fromPhasianuscolchicusintheprotrusionintheproximal directionofthecristacnemialiscranialis,whichislower inLyrurustetrix.Thecranialportionofthescapulacannot belongtoTetraourogallusonthebasisofthesizeanddiffers fromPhasianuscolchicusinthepresenceofthepneumatic foramenbetweenthefaciesarticularishumeralisandthe acromion.Thetotallengthofthetarsometatarsusallows ustoruleoutbothTetraourogallusandPhasianuscolchicus (Erbersdobler,1968).Thefurculadiffersfromtheoneof Phasianuscolchicusintheprotrusionofthehypocleideum, whichislesspronouncedinLyrurustetrix,whileTetrao uro-galluscanberuledoutonthebasisofthesize.Inthecaseof thedistalcoracoid,thedistanceoftheposteriorprocessus
lateralisfromtheapexmedialisallowsustoruleoutboth Tetraourogallus(inwhichitisgreater)andPhasianus colchi-cus(in which it is smaller). In these species, thefacies articularissternalisofthecoracoidsalsoshowsadifferent outlineinsternalview,morecurvedinPhasianus colchi-custhaninLyrurustetrixandstraighterinTetraourogallus (Erbersdobler,1968).
Remarks.Lyrurustetrixismostlysedentary.In north-eastern Europe and Russia, where this species is more widespread,itmostlylivesinlowlandhabitats,transitional betweenwoodland andopenheath,bog andsteppe;in themountainareasofthemid-latitudes,liketheAlps,it survivesasa“glacialrelict”(Cramp,1998;DelHoyoetal., 2014).In Italy, itis currently a sedentaryand breeding speciesintheAlps,whereitlivesinconiferordeciduous forest,alsoinareaswithsparsevegetationornearopen areas (Brichetti and Fracasso, 2004). Pleistocene fossil recordssuggestthatthisspecies,duringcoldphases,was muchmorewidespreadthantoday(HolmandSvenning, 2014).LyrurustetrixisknownsincetheearlyPleistoceneof thePalearcticandisquiteabundantinthelatePleistocene avianfossilrecordofEurope(Tyrberg,Tyrberg,2017).
OrderGruiformes FamilyRallidae GenusCrex
CrexcrexLinnaeus,1758(Fig.3G)
Material.Onedistallefthumerus(RS452)andonedistal rightulna(RS399)fromBR1.
Descriptionandcomparisons.Thebonefeaturesof Ral-lusaquaticusareextremelysimilartothoseofCrexcrex, but,asinthecaseofthedistalhumerusandthedistalulna, Rallusaquaticuscanberuledoutbecauseofitssmallersize. Remarks.CrexcrexcurrentlybreedsinmostofEurope andwintersinsub-saharanAfrica(DelHoyoetal.,2014). InItaly,itisabreeding(onlyinmountainareasof north-eastern Italy), regular migrant and irregular wintering species.Itbreedsinopenorsemi-openenvironments,like meadowswithtallgrassandmoist grasslands(Brichetti andFracasso,2004).CrexcrexisknowninthePalearctic fossilrecord sincethe earlyPleistocene(Tyrberg,1998; Tyrberg,2017).
GenusFulica
FulicaatraLinnaeus,1758(Fig.3H)
Material.Onerightcoracoid(RS378)fromBR2. Descriptionandcomparisons.WithintheRallidae fam-ily, the size of this coracoid is intermediate between PorphyrioporphyrioandGallinulachloropusandthusthis remaincanbedeterminedasFulicaatra.
Remarks.Fulicaatraisdistributed acrossEuropeand Asia.PartofthepopulationwintersinAfricaand south-easternAsia(DelHoyoetal.,2014).InItaly,itiscurrently asedentaryandbreeding,regularmigrantandwintering species.Thisspeciesinhabitsstillorslow-flowingwaters (BrichettiandFracasso,2004).Itisaspeciesthatispresent inthePalearcticfossilrecordsincetheearlyPleistocene (Tyrberg,1998,2017).
OrderStrigiformes FamilyStrigidae
GenusAsio
AsiootusLinnaeus,1758
Material.OnephalanxIIIofdigit2(RS331)andone inde-terminateungualphalanx(RS350)fromBR1.
Descriptionandcomparisons.Thesetwo ungual pha-langes have been ascribed to Asio otus because their proximal epiphysis, in proximal view, are less round-shapedwithrespecttotheproximalepiphysisoftheungual phalangesofAsioflammeus,themostsimilarspeciesfrom anosteologicalpointofview.
Remarks. Asio otus is widespread across the mid-latitudes of the northern hemisphere (Del Hoyo et al., 2014).In Italy,it currentlyis asedentaryand breeding, regularmigrantand winteringspecies. Itprefers wood-landssurroundedbyopenareas,usedforhuntingpurposes (BrichettiandFracasso,2006).Asiootusisreportedinthe PalearcticsincethemiddlePleistoceneandbecomesmore abundantinthelatePleistocene(Tyrberg,1998,2017).
AsioflammeusPontoppidan,1763(Fig.3,I)
Material.OneleftphalanxIofdigit1(RS455)andone phalanxIIofdigit1(RS398)fromBR1.
Descriptionandcomparisons.TheleftphalanxIofdigit 1 shows a processus that extends in the ventral side of theproximal part of thebone. This is a typical fea-tureoftheIstphalanxofthedigit1ofStrigidae.Among medium-sizedStrigidae,itshows greatermorphological correspondencewithAsioflammeusandAsiootus.The spec-imenisattributedtoAsioflammeusbecauseofthestouter outlineand aflatter proximalepiphysis, inlateralview, withrespecttoAsiootus.ThephalanxIIofdigit1(anungual phalanx)hasbeenascribedtothisspeciesbecausethe pro-cessuspresentintheproximalpartoftheboneiswider inproximalviewthaninAsiootusandextendstoamore ventraldirection,inlateralview,thaninAsiootus.
Remarks.Asio flammeusbreeds inthehighand mid-latitudes of the northern hemisphere and winters in southern North America, southern Asia and Africa (Del Hoyoetal.,2014).InItaly,itiscurrentlyaregularmigrant andwinteringspecies.Itpreferswetlands,meadows, grass-landsandsteppes,oftenobservedalsoinmountainareas (BrichettiandFracasso,2006).Asioflammeusisknownin thePalearcticsincethemiddlePleistocene(Tyrberg,1998; Tyrberg,2017).
OrderAccipitriformes FamilyAccipitridae GenusButeo
ButeobuteoLinnaeus,1758(Fig.3J)
Material.Onesternumfragment(rostralarea)(RS323) andonedistalrighttibiotarsus(RS321)fromBR1.
Description and comparisons.This tibiotarsus differs fromtibiotarsiofotherraptorsofsimilarsize(likeAccipiter gentilis,Pernisapivorus,Circusaeruginosus,Milvusmilvus, MilvusmigransandAquilapennata)especiallyinthe cen-tralpositionofthecanalisextensoriusandofthesulcus extensorium,witha 45◦ inclined ponssupratendineum. ButeolagopusandButeorufinustibiotarsiaresimilartothis remaininosteologicalfeatures,butareruledoutbecause oftheirlargersize,especiallyconsideringthatthewidthof thedistalepiphysisofthistibiotarsusisshorterthanthe
rangeindicatedforButeobuteobySchmidt-Burger(1982). Moreover,thisremainisslightlysmallerthanalltheButeo buteotibiotarsiweusedforcomparisonandfromthe lit-erature.ThesternumfragmenthasbeenascribedtoButeo buteoonthebasisofthemorphologicalcorrespondencein thegeneraloutlineofthesulcusarticulariscoracoideusand ofthedorsallabrum,andalsointheshapeoftherostrum spina externa(Otto, 1981).Even thesternum fragment lookssmallerthanthespecimensinthecollection.
Remarks.ButeobuteoiswidespreadacrossEuropeand western Asia, withpart of the populationwintering in sub-saharan Africa(Del Hoyoet al.,2014).InItaly, it is currentlyasedentaryandbreeding,regularmigrantand wintering species.It lives ina varietyof different habi-tats(BrichettiandFracasso,2003).Thisspeciesisreported inthePalearcticfossilrecordsincethemiddlePleistocene (Tyrberg,1998,2017).
OrderPiciformes FamilyPicidae
GenusPicus
PicuscanusGmelin,1788(Fig.3K)
Material.Oneproximalleftcarpometacarpus(RS315) fromBR1.
Description and comparisons. This carpometacarpus, similarinosteologicalfeaturesbothtoPicuscanusandPicus viridis,isslightlysmallerthantheoneofP.viridis.Moreover, theprocessusintermetacarpalisisinamoreproximal posi-tionandthetrochleacarpalisisslightlylesspointedthan inP.viridis.TheotherPalearcticspeciesofPiciformesare ruledoutonthebasisofthesize.
Remarks. Picus canus is distributed across the mid-latitudesofEurasia(DelHoyoetal.,2014).InItaly,where it lives in the eastern Alps, it is currently a sedentary and breeding, irregularmigrantand irregular wintering species. It inhabits conifer, deciduous or mixed forests (BrichettiandFracasso,2007).Picuscanusisknowninthe PalearcticfossilrecordsincethemiddlePleistocene.Inthe latePleistocene,asthefossilrecordattests,itwasmostly spreadincentralandeasternEurope.TheRioSeccoCave remain representsthe third Italian fossilrecord of this species (Pavia and Insacco, 2013; Tyrberg, 1998, 2017). ThefirstcomesfromthelatePleistocenedepositsofthe Caverna delle Arene Candide, in Liguria (Cassoli, 1980), whilethesecond,acompleteulnadubitativelyreferredto P.canus,comesfromthemiddlePleistocenedepositsofthe Comisoarea,inSicily(PaviaandInsacco,2013).
OrderPasseriformes FamilyCorvidae
GenusPica
PicapicaLinnaeus,1758
Material.Onecranialportionofarightscapula(RS338) fromBR1.
Descriptionandcomparisons.Slightdifferencesinthe osteologicalcharacteristicsdistinguishthisscapulafrom thoseofotherCorvidsofthesamesize,likethespeciesof thegenusPyrrhocoraxandCorvusmonedula.Indetail,inthis remaintheacromionisclearlyhigherthantheventralpart
ofthetuberculumcoracoideum:thisisatypicalfeatureof Picapicascapulae(TomekandBochenski,2000).
Remarks. Pica pica is widespread across Europe and mostofAsia(DelHoyoetal.,2016).InItaly,whereitisvery abundantandwidespread,itisasedentaryandbreeding, irregularmigrantandirregularwinteringspecies.Itlives ina varietyofopenandwoodedhabitats (Brichetti and Fracasso,2011).PicapicaisknowninthePalearcticsince theearlyPleistocene.ItislargelypresentintheEuropean fossilrecordofthelatePleistocene(Tyrberg,1998,2017).
GenusNucifraga
NucifragacaryocatactesLinnaeus,1758(Fig.3L) Material.Onedistallefttibiotarsus(RS325)fromBR1. Descriptionandcomparisons.Thewidthofthedistal epiphysisofthetibiotarsusruleoutallCorvidaeapartfrom NucifragacaryocatactesandPicapica(Tomekand Bochen-ski,2000).Theshapeof thecanalisextensoriusandthe obliquityoftheponssupratendineusallowustoruleout Picapica.
Remarks.Nucifraga caryocatactesis spreadacrossthe highandmid-latitudesofEurasia(DelHoyoetal.,2016). In Italy, where it only lives in the Alps, is a sedentary and breeding,irregular migrantand irregular wintering species.Itinhabitsconiferforests(BrichettiandFracasso, 2011). This species is reported in the Palearctic fossil recordsincethemiddlePleistocene(Tyrberg,1998,2017).
FamilyTurdidae GenusTurdus
cf.TurdusviscivorusLinnaeus,1758(Fig.3M) Material.Tworightcarpometacarpi(RS356andRS411) fromBR1.
Descriptionandcomparisons.Thetwocarpometacarpi, clearlybelongingtothefamilyTurdidaeonthebasisoftheir osteological features, are tentatively referred to Turdus viscivorus for thelargesize and for a high morphologi-calcorrespondence.Nevertheless,thetotallengthofboth the specimens islonger thanany Turdusviscivorus car-pometacarpusofthecollection(RS411length:25.8mm; RS356length:25.4mm).IntheEuropeanbirdfossilrecord ofthePleistocene,severalspecimensofaTurdidaebigger thanTurdusviscivorushavebeenreported.Someofthem (onlyhumeriandcranialremains)haveevenbeenassigned toanewspecies,Meridiocichlasalotti(Louchart,2004). Nev-ertheless,consideringthatcarpometacarpiarenothighly diagnosticbones(especiallyforPasseriformes),and con-sideringtheabsenceintheliteratureofthesizerangeof carpometacarpiof Turdusviscivorus,we preferto tenta-tivelyreferRS356andRS411tocf.Turdusviscivorusinstead, ratherthantootherlessprobabletaxalikeZootheradauma (abigasianTurdidae,rarevagrantinEurope)or Meridioci-chlasalotti(Louchart,2004).
Remarks. Turdus viscivorus is distributed across the highandmid-latitudesofEuropeand westernAsia(Del Hoyoetal.,2016).InItaly,itisasedentaryandbreeding, regular migrant and wintering species. It lives in open woodedenvironmentsinmountainorhillyareas,whereit prefersclearingsusedforfeedingpurposes(Brichettiand Fracasso,2008).Turdusviscivorousisknowninthefossil
recordofthePalearcticsincethemiddlePleistocene.Itis commonlyreportedinEuropeanlatePleistocenelocalities (Tyrberg,1998,2017).
FamilyMuscicapidae GenusErithacus
ErithacusrubeculaLinnaeus,1758
Material.Onelefthumerus(RS363)fromBR1andone proximalrighthumerus(RS434)fromBR2.
Descriptionandcomparisons.Thehumeriofthe gen-eraLusciniaandErithacusshowsomedifferencesfromthe generalosteologicalfeatures ofthehumeriof theother membersofthefamilytheybelong,theMuscicapidae.The lattershow,intheproximalhumerus,2fossae,separated byamedialbar,whileLusciniaandErithacushaveonlythe pneumo-anconeanfossawelldeveloped, witha shallow tricipitalis fossa.The humerusof Erithacus rubecula dif-fersfromLusciniamegarhynchosandLuscinialusciniainthe smallersizeandfromLusciniasvecicainthenarrower prox-imalepiphysisandintheabsenceofarimwhich,fromthe tricipitalisfossa,continuesupuntiltheventralpartofthe bone(Janossy,1983).
Remarks. Erithacus rubecula is widespread across Europe and northern Africa(Del Hoyo et al., 2016). In Italy,itiscurrentlyasedentaryandpartialbreeding, reg-ularmigrantandwinteringspecies.Itlivesinavarietyof woodedenvironments(BrichettiandFracasso,2008).This speciesisreportedinthePalearcticfossilrecordsincethe middlePleistoceneandisscarcelyrepresentedintheItalian Pleistocenefossillocalities(Tyrberg,1998;Tyrberg,2017).
FamilyPrunellidae GenusPrunella
PrunellacollarisScopoli,1769(Fig.3N) Material.Onemaxilla(RS390)fromBR1.
Description and comparisons. Thegeneral outlinein dorsalviewandtheslightcurvingoftheosnasalein cor-respondencewiththenosecavity inlateralview,make thisremainascribabletothefamilyPrunellidae.Withinthe family,itssizerulesoutPrunellamodularis,whichisclearly smaller(Cuisin,1989;Janossy,1983).
Remarks.Prunellacollarisisspreadacrossthe Mediter-ranean basin and central Asia (Del Hoyo et al., 2016). InItaly,itiscurrentlyasedentaryandbreeding,regular migrantandwinteringspecies.Duringthebreedingseason it lives in mountain areas withlow cliffs, rocky slopes and alpine meadows,above thetree line(Brichetti and Fracasso,2007).Thisspeciessurvivesin mountainareas ofmid-latitudesasa“glacialrelict”(Tyrberg,1991).This speciesisknowninthePalearcticfossilrecordsincethe middlePleistocene(Tyrberg,1998,2017).
FamilyFringillidae GenusPyrrhula
PyrrhulapyrrhulaLinnaeus,1758
Material.Onelefthumerus(RS411)fromBR1andone proximallefthumerus(RS500)fromBR2.
Description and comparisons. The left complete humerusisascribedtoPyrrhulapyrrhulaonthebasisofthe
totallengthandthemorphologyoftheproximalepiphysis, withthemedialbarextendingmoreintothedirectionof dorsalside,afeaturethatallowsustoseparatethisspecies fromtheotherFringillidaeofthesamesize(Janossy,1983). TheproximalhumerusisreferredtoPyrrhulapyrrhulaon thebasisofthesizeandoftheabove-mentioned morphol-ogyoftheproximalepiphysis.
Remarks.Pyrrhula pyrrhulaisspread acrosshighand mid-latitudes of Eurasia (Brichetti and Fracasso, 2013; DelHoyoetal.,2016).InItaly,itiscurrentlyasedentary and breeding,regular migrantand wintering species. It inhabits conifer,deciduousand mixed forests(Brichetti and Fracasso, 2013). Pyrrhula pyrrhula is known in the Palearctic fossil record since the middle Pleistocene (Tyrberg,1998,2017).
GenusLoxia
LoxiacurvirostraLinnaeus,1758(Fig.3O) Material.Onemaxilla(RS504)fromBR2.
Descriptionandcomparisons.Theoutlineand morphol-ogyofthebill,withcrossedtips,isthemosttypicalfeature ofthisspecies,thatmakesitunmistakable(Cuisin,1989). BetweentheotherspeciesofthegenusLoxia,Loxia pyty-opsittacusand Loxiascotica can beruled outbecauseof theirmorerobustbillandagreatersizethanL.curvirostra. LoxialeucopteradiffersfromL.curvirostrainbeingslightly smallerandintheslenderbill.
Remarks.Loxiacurvirostraisdistributedacross temper-ateandsubarcticareasofthenorthernhemisphere(Del Hoyoetal.,2016;BrichettiandFracasso,2013).InItaly,it iscurrentlyasedentaryandbreeding,regularmigrantand winteringspecies,thatlivesinconiferforestsofmountain areas(BrichettiandFracasso,2013).Thisspeciesisreported in thePalearcticfossilrecord since the late Pleistocene (Tyrberg,1998,2017).
5. Discussion
Thebirdtaxarecognizedinthisworkallbelongtothe extantItalianavifauna(BrichettiandFracasso,2015).Taxa likeLyrurustetrix,Tetraourogallus,Picuscanus,Nucifraga caryocatactes,Pyrrhulapyrrhula,Loxiacurvirostraindicate thepresence,inthevicinityofthesite,ofmainlyconifer ormixedforests.TaxalikeLagopusmutaePrunellacollaris suggestthepresenceofopenareaswithlow vegetation androckygrounds,abovethetreeline.Lagopusmutais absenttodayfromtheareaofthesite.Indetail,thisspecies hasbeenobserved12timesinFriuli-VeneziaGiuliainthe periodbetween2006–2011,butatalmost30kmfurther fromthesiteandathigherheights(Guzzonetal.,2013). Duringthetimespanrepresentedinthesedimentsinthis study,thesite,which islocatedat580masl,was prob-ablysurroundedbyaforestenvironment,ifweconsider thattheminimumheightreachedbythetreelineduring LGM,intheorientalPrealps,was7–800masl(Ravazzietal., 2007).Anyway,assumingthatthecatchment/huntingarea ofthepredatorresponsiblefortheaccumulationofbird remains(being ita nocturnal raptor,adiurnal raptor,a carnivoreoraman)ishardlywiderthan30kmfromthe site(Serjeantson,2009),thepresenceofLagopusmutaat
theRioSeccoCaveindicatesthatmountainopen environ-mentswere locatedclosertothesite, and therefore,as expected,thetreelinewaslocatedatlowerheightsthan today.Thepresenceoftheseopenandrockyenvironments is furthermoreconfirmedby Capra ibex,Rupicapra rupi-capraandMarmotamarmota(Peresanietal.,2014).Perdix perdixsuggeststhepresenceofopenareaswithlow vegeta-tion,whileTurdusviscivorusindicatesthepresenceofopen woodedareas.Stillorslow-flowingwaterbodieshadtobe foundnearthesite,asthepresenceofFulicaatraindicates. ThisisconfirmedbytheremainsofCastorfiber(Peresani etal.,2014).Coturnixcoturnix,Crexcrex,Asiootus,Asio flam-meus,ButeobuteoandErithacusrubecula,speciesthattoday haveamigratorybehaviour,couldhavedeceasedalso dur-ingmigration.Inthisphasetheycanbefoundinseveral environments,therefore,thesetaxaarenotusefulfor pale-oenvironmentreconstructionpurposes.Pica picadoesn’t giveanypalaeoecologicalindicationasitcurrentlylivesin avarietyofdifferentenvironments.
The taxa identified do not suggest any significant environmental change between the investigated layers, showing a prevalence, both in BR1 and BR2, of species typicalofforestenvironments,intermsofthenumberof identifiedspecimensandalsoofthenumberoftaxa iden-tified.
ThefindingofL.mutaremainsatlowerheightswith respecttoitscurrentaltitudinaldistributionsuggests,as expected,thepresenceofacolderclimatethanthatofthe present,aswearedealingwiththeonsetoftheLGM. How-ever,cautionisrequiredwheninferringclimateoscillations onthebasisofchangesinfossilavifaunadistribution pat-terns.Itmustbetakenintoaccountthatthesechangescan beduetoavarietyofdifferentfactorsthatarenot neces-sarilylinkedtotemperatureshifts,suchasvegetationand foodavailability,seasonality,precipitationtrends, fluctu-ationsofthepopulations,interactionswithotherspecies (potentialcompetitors),changesintheecologyandhabits ofthespecies,andevenecologicalpressureduetohuman activities.Onlystronglatitudinaloraltitudinalshiftsina birdspeciesdistribution,possiblysupportedbyother evi-dencesandbyattestedclimatetrends,shouldbeusedasa climateproxy(Bedetti&Pavia,2001;Bedetti&Pavia,2007; Finlayson,2011;Tyrberg,2010).
Ofparticularinterestisthefinding,amongthebirdfossil remainsofthissite,ofacarpometacarpusofP.canus.This remainrepresentsthesecondItalianlatePleistocenefossil recordofthisspeciesandthethirdforItaly.Thisrecordfalls insideitscurrentItaliandistribution,whiletheotherItalian fossilrecordsfalloutsideitscurrentdistribution(Cassoli, 1980;PaviaandInsacco,2013;Tyrberg,1998,2017).
Theresultsofthetaphonomicanalysisindicatethatthe responsiblesfortheaccumulationofthebirdremainsinthe deposit werenocturnalraptors, carnivoresandhumans. Somenocturnalraptorspeciesfrequentcavesandrocky cliffs,forbreedingorrestingpurposes,andpreyonother birds.Thebonesoftheirpreys,ejectedinthepellets,can show traces ofdigestion (rounding and thinningof the articularandbrokenends,lookinglikeaslightcorrosion), or,often,theyshownotracesofdigestionatall(Andrews, 1990; Bochenski&Tomek,1997; Rufàetal.,2017; Ser-jeantson, 2009). Diurnal raptors or carnivore digestion
usuallyproducesmoreseveredamageonthesurfaceofthe bonesbecauseoftheirstrongerdigestiveacids(Bochenski etal.,1997,1998;Lloverasetal.,2014,2016;Serjeantson, 2009). Furthermore, the small bones contained in owl pelletsaremoreprotectedfrompost-depositional tapho-nomicmodificationsastheyarewrappedinamatrixoffur andfeathers(Serjeantson,2009).Thepercentageofslightly corrodedbirdremainsthatwedetectedintheassemblage (11,9%ofthetotal,n=23)andtheamountofwellpreserved smallPasseriformesandsmallrodentremains,supportthe hypothesisofnocturnalraptorsasaccumulatorsofatleast partofthebirdbones.Inthisavianfossilassemblage,we identifiedtwospeciesofStrigiformes,AsiootusandAsio flammeus.Thesespecies,however,donotusuallyfrequent caves orrockycliffs(Brichetti&Fracasso, 2006;Cramp, 1998),thereforetheyshouldnotbeconsideredthe respon-siblefor theaccumulationofthebird bones.Bubobubo couldmostlikelybethecausativeagent,especially con-sideringthatsomeoftheslightlycorrodedbird remains belongtoLyrurustetrixandTetraourogallus(thespeciesof thegenusAsiofeedonlyonsmallerbirds).Moreover,Bubo buboisreportedtopreyondiurnalandnocturnalraptors (Brichetti&Fracasso,2006;Cramp,1998;Mikkola,1976): thiscouldexplainthepresenceofAsiootusandAsio flam-meusamongthebirdremains.Thehomerange(i.e.hunting area)ofapairofB.buboisreportedtobeupto20km2,even
ifsomestudiesreportthattheirterritoriesmaybelarger (Cramp,1998;Martínezetal.,2003).Therefore,assuming B.buboastheaccumulator,thebirdspeciesitbroughttothe siterepresenttheenvironmentwithinarangeof approxi-mately2,5kmaroundthesite.
Wecanassessthatcarnivoreshavealsobeen respon-siblefor theaccumulation of partofthe bird bones,as carnivore damages (pits, punctures, scores) and strong digestivedamagehavebeendetected,respectively,on2.1% (n=4)and1%(n=2)ofthebirdremains.Unfortunately,we donothaveenoughdatafromthetaxonomicalanalysisof themammalremainsfromtheselayers,yet,toinferwhich carnivorespeciescouldhavebeenthepossible accumula-tor.
Amongtheothernaturalmodificationsdetected, man-ganesedioxidestainingwasthemostcommon(20.6%ofthe total,n=40),followedbyrootmarks(2.6%,n=5),rodent’s gnawing(2.1%,n=4),trampling(2.1%,n=4),concretions (1.5%,n=3)andexfoliation(0.5%,n=1).Someofthebird remainsshowhumanmodification,thatwillbepresented indetailinaforthcomingcontribution.
ThepollenrecordfromAzzanoDecimocore(whichis locatedabout40kmfarfromtheRio SeccoCave,inthe Friulianplain)showsanincreaseofxerophyticplantsfor thetimeintervalconsideredinthiswork,confirmingthe expansionofopenanddryenvironmentscausedbyharsher climaticconditions(Pinietal.,2009).
6. Conclusions
ThesystematicanalysisofthebirdremainsoftheRio SeccoCaveallowedpalaeoenvironmentalreconstructions. AttheonsetoftheLGM,thesurroundingofthecavewas characterized,onthebasisoftheaviantaxa,bythe pres-enceofdifferentenvironments,suchasconiferormixed
forests,openareaswithlowvegetationandrockygrounds, open grasslands, open wooded areas and slow-flowing waterbodies.Harshconditionswithincreaseofxerophytic plants,inthetimeintervalinvestigated,arealsoindicated fromtheAzzanoDecimopollencoreintheFriulianplain about40km south ofRio SeccoCave (Pini etal.,2009). ThismosaicofhabitatsinthenorthernbeltoftheGreat AdriaticPlain,thatguaranteedtheavailabilityofseveral different resources, probably attracted early Gravettian hunter-gathererswhovisited thesiteabout30 kayBP. ThepresenceofL.mutaatlowheightsalsosuggeststhe presenceofaclimatecolderthantodayduringthetime spaninvestigated.Besides,thefindingofthesecondItalian latePleistocenefossilrecordofP.canusconfirmsthe long-standingpresenceofthisspeciesinthemountainareasof north-easternItaly.
This contribution represents a further step in the knowledgeofpaleornithocenosis,palaeoenvironmentand palaeoclimateofthelatePleistoceneintheeasternAlps. Moreover,thisisthefirstsystematicanalysisofanavian fossilassemblagefromtheareaofFriuli-VeneziaGiulia,for aperiodolderthantheLateglacial(Tonon,1983)and,at thesametime,supportedbydatingsandareliable stratig-raphy.Furtheranalysesoffossilavifaunalassemblageswill givenewinsightsaboutthepalaeoecologyand paleobio-geographyofbirds,theenvironmentsofthepast,andeven abouthuman-birdinteractions inPrehistory(Blascoand Peresani,2016).
Acknowledgements
FinancialsupporttotheRioSeccoResearchProjectis providedbytheAdministrationoftheClauzetto Munic-ipalityand theFriuli VeneziaGiuliaRegionanda group ofpublicinstitutions(EcomuseodelleDolomiti Friulane “Lis Aganis”, BIM Tagliamento Consortium, Pordenone Province), Foundations (Fondazione CRUP) and private companies(FriulovestBanca).M.Peresanistructuredthe researchprojectand directedthefieldwork;M. Roman-dini coordinatedthefieldwork.TheRio Seccoprojectis alsoco-designedbytheNeanderthalMuseum(A.Pastoors &G.C. Weniger)and theUniversitat Rovira yVirgili at Tarragona(M.Vaquero).Thisstudyhasreceivedfunding fromtheTorinoUniversityex60%Grants2016and2017. WearegratefultoProf.Benedetto Salaforhishelpand precioussuggestions,tothreeanonymous reviewersfor constructivesuggestionsandtoLeahMasciaforrevision oftheEnglishtext.Authors’contributions:M.Pe.andM.R. plannedtheresearch;L.C.,M.Pa.andM.R.analyzedthe data;L.C.,M.Pa.,M.R.andM.Pe.wrotethepaper.
References
Andrews,P.,1990.Owls,cavesandfossils.Naturalhistorymuseum pub-lications,London,pp.231.
Antonioli,F.,Vai,G.B.,2004.ClimexMapsItalyandexplanatorynotes. In:Margottini,C.,Vai,G.B.(Eds.),ProjectsponsoredbyENEA.32nd InternationalGeologicalCongress,Florence,Italy.
Baumel,J.J.,Witmer,L.M.,1993.Osteologia.In:Baumel,J.J.(Ed.), Hand-bookofAvianAnatomy:NominaAnatomicaAvium.,SecondEdition. PublicationsoftheNuttallOrnithologicalClub,23,CambridgeMA,pp. 45–132.
Bedetti,C.,Pavia,M.,2001.Climateandenvironmentalchangesandfaunal turnover:theavifaunacasehistory.Internationalconferenceon pale-obiogeographyandpaleoecology.Placenza,Abstractbook,pp.34–35. Bedetti,C.,Pavia,M., 2007. Reinterpretationof theLate Pleistocene IngaranoCavedepositbasedonthefossilbirdassociation(Apulia, southeasternItaly).RivistaItalianadiPaleontologiaeStratigrafia113 (3),487–507[Res.Paleontol.Stratigra.].
Bedetti, C., Pavia, M., 2013. Early Pleistocene birds from Pirro Nord (Puglia, southern Italy). Palaeontographica Abteilung A-Paläozoologie-Stratigraphie298.,pp.31–53.
Blasco,R.,Peresani,M.,2016.Human-birdinteractionsinprehistory.Quat. Internat.421,1–5.
Bochenski,Z.,Tomek,T.,1997.Preservationofbirdbones:erosionversus digestionbyowls.Int.J.Osteoarchaeol7,372–387.
Bochenski,Z.,Korovin,V.,Nekrasov,A.E.,Tomek,T.,1997.Fragmentation ofbirdbonesinfoodremainsofimperialeagles(Aquilaheliaca).Int.J. Osteoarchaeol.7,165–171.
Bochenski,Z.,Huhtala,K.,Jussila,P.,Pulliainen,E.,Tornberg,R.,Tunkkari, P.S.,1998.DamagetobirdbonesinpelletsofgyrfalconFalcorusticolus. J.Archaeol.Sci.25,425–433.
Bon,M.,Piccoli,G.,Sala,B.,1991.IgiacimentiquaternaridiVertebrati fossilinell’Italianord-orientale.MemoriediScienzeGeologiche43, 185–231.
Brichetti,P.,Fracasso,G.,2003.Ornitologiaitaliana.Vol.1: Gaviidae-Falconidae.Identificazione,distribuzione,consistenzaemovimenti degliUccelliitaliani.AlbertoPerdisaEditore,463p.
Brichetti,P.,Fracasso,G.,2004.Ornitologiaitaliana.Vol.2: Tetraonidae-Scolopacidae.Identificazione,distribuzione,consistenzaemovimenti degliUccelliitaliani.AlbertoPerdisaEditore,396p.
Brichetti,P.,Fracasso,G.,2006.OrnitologiaItaliana.Vol.3: Stercorariidae-Caprimulgidae.Identificazione,distribuzione,consistenza e movi-mentidegliUccelliitaliani.AlbertoPerdisaEditore,437p.
Brichetti,P.,Fracasso,G.,2007.Ornitologiaitaliana.Vol.4: Apodidae-Prunellidae.Identificazione,distribuzione,consistenzaemovimenti degliUccelliitaliani.AlbertoPerdisaEditore,441p.
Brichetti,P.,Fracasso,G.,2008.Ornitologiaitaliana.Vol.5: Turdidae-Cisticolidae.Identificazione,distribuzione,consistenzaemovimenti degliUccelliitaliani.AlbertoPerdisaEditore,429p.
Brichetti,P.,Fracasso, G.,2011. Ornitologiaitaliana. Vol.7: Paridae-Corvidae.Identificazione, distribuzione, consistenza e movimenti degliUccelliitaliani.AlbertoPerdisaEditore,493p.
Brichetti,P.,Fracasso,G.,2013.Ornitologiaitaliana.Vol.8: Sturnidae-Fringillidae,Identificazione,distribuzione,consistenzaemovimenti degliUccelliitaliani.AlbertoPerdisaEditore,464p.
Brichetti,P.,Fracasso,G.,2015.Check-listdegliuccelliitalianiaggiornata al2014.RivistaItalianadiOrnitologia.Res.Ornithol.85,31–50. Bruderer,B.,Salewski,V.,2008.Evolutionofbirdmigrationina
biogeo-graphicalcontext.J.Biogeography35(11),1951–1959.
Cassoli,P.F.,1980.L’avifaunadelPleistocenesuperioredelleArene Can-dide(Liguria).Memoriedell’IstitutoItalianodiPaleontologiaUmana Roma3,pp.155–234.
Cassoli,P.F.,Tagliacozzo,A.,1994.Considerazionipaleontologiche, pale-oecologicheearcheozoologichesuimacromammiferiegliuccellidei livellidelPleistocenesuperioredelRiparodiFumane(VR)(scavi 1988-91).BollettinodelMuseoCivicodiStoriaNaturalediVerona 18(1991).,pp.349–445.
Clark,P.U.,Dyke,A.S.,Shakun,J.D.,Carlson,A.E.,Clark,J.,Wohlfarth,B., Mitrovica,J.X.,Hostetler,S.W.,McCabe,A.M.,2009.TheLastGlacial Maximum.Science325,710–714.
Cramp,S.,1998.ThecompletebirdsofthewesternPalearcticonCD-ROM. Optimedia.OxfordUniversityPress.
Cuisin,J.,1989.L’identifcationdescranesdePassereaux(Passeriformes: Aves).PhDdissertation.UniversityofBoulogne,France,340p. DelHoyo,J.,Collar,N.J.,Christie,D.A.,Elliot,A.,Fishpool,L.D.C.,2014.
Illus-tratedchecklistofthebirdsoftheworld.Vol.1:non-passerines.Lynx Éditions,Barcelona,903p.
DelHoyo,J.,Collar,N.J.,Christie,D.A.,Elliott,A.,Fishpool,L.D.C.,Boesman, P.,Kirwan,G.M.,2016.Illustratedchecklistofthebirdsoftheworld. Vol.1:non-passerines.LynxÉditions,Barcelona,1013p.
Erbersdobler,K.,1968.Vergleichend morphologischeUntersuchungen anEinzelknochendespostcranialSkelettsinMitteleuropa vorkom-mendermittelgrosserHuhnervogel.UnpublishedPhDDissertation, Ludwig-Maximilians-Universitat,München,153p.
Finlayson,C.,2011.Aviansurvivors.Thehistoryandbiogeographyof palearcticbirds.T&ADPoyser,London,pp.304.
Fiore,I.,Gala,M.,Tagliacozzo,A.,2015.L’originedellacacciaagliuccelli nellapenisolaitaliana.In:Preistoriadelcibo.L’ambientefontedi risorsealimentari.Sessione2.LRiunioneScientificaIstitutoItaliano PreistoriaeProtostoria.
Gala,M.,Tagliacozzo,A.,2009.Aspettiecologiciedeconomicidell’avifauna delTardoglacialeitaliano.In:Atti6◦ConvegnoNazionaledi Archeozo-ologia.,pp.79–88.
Gala,M.,Tagliacozzo,A.,2010.TheavifaunafromLateGlacial archaeolog-icalsitesinItaly:atentativesynthesis.In:Prummel,W.,Zeiler,J.T., Brinkhuizen,D.C.(Eds.),BirdsinArchaeology:proceedingsofthe6th MeetingoftheICAZBirdWorkingGroupinGroningen.,12.Barkhuis, 205p.
Gordo,O.,2007.Whyarebirdmigrationdates shifting?Areviewof weatherandclimateeffectsonavianmigratoryphenology.Climate Res35(1–2),37–58.
Gurioli,F.,Cappato,N.,DeStefani,M.,Tagliacozzo,A.,2006. Consider-azioniPaleontologiche,PaleoecologicheeArcheozoologichedeilivelli delPaleoliticosuperioredelRiparodelBroion(ColliBerici,Vicenza). In:AttidelConvegnoNazionalediArcheozoologia,Rovereto,5.,pp. 47–56.
Guzzon,C.,Kravos,K.,Parodi,R.,Sava,S.,Toller,M.,2013.Resoconto orni-tologicodelFriuliVeneziaGiulia.In:Anni2006-2011.MuseoFriulano diStoriaNaturale,Udine,pp.191.
Higham, T., Brock, F., Peresani, M., Broglio, A., Wood, R., Douka, K., 2009. Problems with radiocarbon dating the Middle to Upper Palaeolithic transition in Italy. Quat. Sci. Rev. 28 (13), 1257–1267.
Holm,S.R.,Svenning,J.C.,2014.180,000yearsofclimatechangeinEurope: avifaunal responses and vegetation implications.Plos one9 (4), e94021.
Janossy,D.,1983.HumeriofcentralEuropeansmallerPasseriformes. Frag-mentaMineralo.Palaeontol.11,85–112.
Jonzén,N.,Lindén,A.,Ergon,T.,Knudsen,E.,Vik,J.O.,Rubolini,D., Piacen-tini,D.,Brinch,C.,Spina,F.,Karlsson,L.,Stervander,M.,Andersson,A., Waldenström,J.,Lehikoinen,A.,Edvardsen,E.,Solvang,R.,Stenseth, N.C.,2006.Rapidadvanceofspringarrivaldatesinlong-distance migratorybirds.Science312(5782),1959–1961.
Kraft, E., 1972. Vergleichende morphologische Untersuchungen an Einzelknochen nord und mitteleuropaischer kleinerer Hunervo-gel.UnpublishedPhDDissertation,Ludwig-Maximilians-Universitat, München.,pp.195.
Lloveras,L.,Thomas,R.,Lourenc¸o,R.,Caro,J.,Dias,A.,2014. Understand-ingthetaphonomicsignatureofBonelli’seagle(Aquilafasciata).J. Archaeol.Sci.49,455–471.
Lloveras,L.,Thomas,R.,Cosso,A.,Pinyol,C.,Nadal,J.,2016.Whenwildcats feedonrabbits:anexperimentalstudytounderstandthetaphonomic signatureofEuropeanwildcats(Felissilvestrissilvestris).Archaeol. Anthropol.Sci.
Louchart,A.,2004.Anextinctlargethrush(Aves:Turdidae)fromtheLate QuaternaryofMediterraneanEurope.NeuesJahrb.Geol. Palaeontol.-Abh.233,275–296.
Martínez,J.A.,Serrano,D.,Zuberogoitia,I.,2003.Predictivemodelsof habi-tatpreferencesfortheEurasianeagleowlBubobubo:amultiscale approach.Ecography26(1),21–28.
Mikkola,H.,1976.Owlskillingandkilledbyotherowlsandraptorsin Europe.Br.Birds69,144–154.
Monegato,G.,Scardia,G.,Hajdas,I.,Rizzini,F.,Piccin,A.,2017.TheAlpine LGMintheborealice-sheetsgame.Sci.Rep.7(1),2078.
Oros Srˇsen,A.,Brajkovi ´c,D.,Radovi ´c,S.,MauchLenardi ´c,J.,Miracle, P.T., 2014. TheAvifauna of SouthernIstria (Croatia) during the latePleistocene:implicationsforthepalaeoecologyand biodiver-sityoftheNorthernAdriaticregion.Int.J.Osteoarchaeol.24(3), 289–299.
Otto,C.,1981.VergleichendmorphologischeUntersuchungenan Einzel-knocheninZentraleuropavorkommendermittelgrossenAccipitridae: 1,Schadel,Brustbein,SchultergurtelundVorderextremitat. Unpub-lishedPhDDissertation.Ludwig-Maximilians-Universitat,Munchen, 183p.
Pavia,M.,Insacco,G.,2013.Thefossilbirdassociationsfromtheearly middlePleistoceneoftheRagusaprovince(SESicily,Italy).Bollettino dellaSocietàPaleontologicaItaliana52(3),157–165.
Pellegrino,I.,Negri,A.,Cucco,M.,Mucci,N.,Pavia,M., ˇSálek,M.,Boano,G., Randi,E.,2014.PhylogeographyandPleistocenerefugiaoftheLittle OwlAthenenoctuainferredfrommtDNAsequencedata.Ibis156(3), 639–657.
Peresani,M.,Romandini,M.,Duches,R.,Jéquier,C.,Nannini,N.,Pastoors, A.,Picin,A.,Schmidt,I.,Vaquero,M.,Weniger,G.C.,2014.New evi-dencefortheMousterianandGravettianatRioSeccoCave,Italy.J. FieldArchaeol.39,401–416.
Pini,R.,Ravazzi,C.,Donegana,M.,2009.Pollenstratigraphy, vegeta-tionandclimatehistoryofthelast215kaintheAzzanoDecimo core(plainofFriuli,north-eastern Italy).Quat. Sci.Rev.28(13), 1268–1290.
Ravazzi,C.,Peresani,M.,Pini,R.,Vescovi,E.,2007.IlTardoglacialenelleAlpi italianeeinPianuraPadana.Evoluzionestratigrafica,storiadella veg-etazioneedelpopolamentoantropico.IlQuaternario20(2),163–184. Romandini,M.,Nannini,N.,2012.Chasseursépigravettiensdansle ter-ritoiredel’oursdescavernes:lecasduCovoloFortificatodiTrene (Vicenza,Italie).Anthropologie116(1),39–56.
Romandini,M.,Peresani,M.,Laroulandie,V.,Metz,L.,Pastoors,A.,Vaquero, M.,Slimak,M.,2014.Convergentevidenceofeagletalonsusedbylate neanderthalsinEurope:afurtherassessmentonsymbolism.PlosOne 9(7),e101278.
RomandiniM.,TerlatoG.,NanniniN.,TagliacozzoA.,PeresaniM.,(in press)Manandbear,aNeanderthalhistory:reconstructing uncom-monbehaviorsfromsouthernEuropeanzooarchaeologicalevidence. J.Archaeol.Sci.
Rufà,A.,Blasco,R.,Roger,T.,Rué,M.,Daujeard,C.,2017.Arallyingpointfor differentpredators:theavianrecordfromaLatePleistocenesequence ofGrottedesBarassesII(Balazuc,Ardèche,France).Archaeol. Anthro-pol.Sci.,1–18.
SanchezMarco,A.,2004.Avianzoogeographicalpatternsduringthe Qua-ternaryintheMediterraneanregionandpaleoclimaticinterpretation. Ardeola51(1),91–132.
Schmidt-Burger,P.,1982.Vergleichendmorphologische Untersuchun-genanEinzelknocheninZentraleuropavorkommendermittelgrossen Accipitridae:2,BekenundHinderextremitat.UnpublishedPhD Dis-sertation.Ludwig-Maximilians-Universitat,München,pp.122. Serjeantson,D.,2009.Birds.CambridgeManualsinArchaeology,
Cam-bridgeUniversityPress,Cambridge,514p.
Shackleton,J.C.,vanAndel,T.H.,Runnels,C.N.,1984.Coastal paleogeog-raphyofthecentral andwesternMediterranean duringthe last
125,000yearsanditsarchaeologicalimplications.J.FieldArchaeol. 11,307–315.
Talamo,S.,Peresani,M.,Romandini,M.,Duches,R.,Jéquier,C.,Nannini, N.,Pastoors,A.,Picin,A.,Vaquero,M.,Weniger,G.C.,Hublin,J.J.,2014. DetectinghumanpresenceattheborderofthenortheasternItalian Pre-Alps.14CDatingatRioSeccoCaveasexpressionofthefirst
Gravet-tianandthelateMousterianinthenorthernAdriaticregion.PlosOne 9(4),e95376.
Tomek,T.,Bochenski,Z.M.,2000.ThecomparativeosteologyofEuropean Corvids(Aves:Corvidae),withakeytotheidentificationoftheir skele-talelements.Instituteofsystematicsandevolutionofanimals,Polish academyofsciences,Krakow,Poland,102p.
Tonon,M.,1977.RinvenimentodiNycteascandiacanell’avifauna pleis-tocenica del Covolo di Trene nei Colli Berici (Vicenza). Annali dell’UniversitàdiFerraraIII(3),54–60.
Tonon, M., 1983. Rinvenimento di Cinclus cinclus (L.) nell’avifauna pleistocenica della grotta di Pradis (Pordenone). In: Atti del IV ConvegnodiSpeleologia,Friuli VeneziaGiulia. GEAP,Pordenone, pp.17–20.
Tyrberg,T.,1991.Arctic,montaneandsteppebirdsasglacialrelictsin WestPalearctic.Orn.Verh25,29–49.
Tyrberg,T.,1998.PleistocenebirdsofthePalearctic:acatalogue. Publica-tionsoftheNuttallOrnithologicalClubNo.27,Cambridge,720p. Tyrberg,T.,2010.Avifaunalresponsestowarmclimate:themessagefrom
lastinterglacialfaunas.Rec.Aust.Museum62(1),193–205. Tyrberg,T.,2017.Supplementto“PleistocenebirdsofthePalaearctic”at
http://web.telia.com/∼u11502098/pleistocene.html.(Accessedon10 July2017).