1
Università di Pisa
Facoltà di Scienze Matematiche Fisiche e
Naturali
Corso di Laurea Specialistica in Biologia Marina
Relatori: Dott. Gianni Bedini
Candidato: Silvia Livelli
Primo contributo allo studio del contingente
macroalgale del litorale pisano
Dott.ssa Giuseppina Pardi
Dott. Fabio Rindi
2
Indice
1. Introduzione... 3
1.1 Parco di Migliarino San Rossore e Massacciuccoli ... 3
1.2 I popolamenti macroalgali ... 5
1.2.1 Chlorophyta ... 8
1.2.2 Chlorophyta più diffuse nel Mar Mediterraneo ... 10
1.2.3 Heterokontophyta... 13
1.2.4 Heterokontophyta più diffuse nel Mar Mediterraneo ... 14
1.3 Scopo dello studio ... 17
2. Materiali e metodi ... 18
2.1 Disegno di campionamento ... 19
2.2 Metodo di campionamento ... 24
2.3 Identificazione e conservazione dei campioni ... 25
2.4 Elaborazione dei dati ... 27
3. Risultati ... 28
3.1 Lista floristica ... 28 3.2 Grafici nMDS ... 404. Discussioni e conclusioni ... 50
5. Bibliografia ... 62
6. Appendice ... 72
3
1. Introduzione
1.1 Parco di Migliarino San Rossore e Massacciuccoli
La costa toscana è sede di importanti studi ecologici, in particolar modo alla fine degli anni ‟80 i popolamenti marini del litorale di Calafuria, a sud di Livorno, sono stati oggetto dei primi studi sperimentali effettuati nel Mediterraneo (Maggiet al., 2009); mancavano però dati qualitativi sulla flora macroalgale presente nel
Parco di Migliarino San Rossore e Massaciuccoli lungo il litorale pisano.
L'area protetta, istituita nel 1979, non tutela la parte marina ad eccezione delle Secche della Meloria istituite nel 2010; inoltre, essendo l‟area interessata dallo sbocco dei fiumi Arno, Morto Nuovo e Serchio e in vicinanza di centri urbani nonché di impianti turistici, è soggetta a fonti di alterazione ambientale.
Il Parco di Migliarino San Rossore Massaciuccoli è stato istituito con Legge Regionale Toscana n. 61 del 13 dicembre 1979 (Ente Parco regionale Migliarino S. Rossore Massaciuccoli, 2011) e presenta un‟estensione di circa 23.000 ettari sviluppandosi lungo circa 32 Km. di costa compresa tra le città di Livorno e di Viareggio (Rete civica pisana, 2011).
Questo tratto di litorale è esclusivamente di natura sabbiosa e quindi inadatto ad ospitare macroalghe bentoniche, che trovano però un habitat soddisfacente sulle barriere artificiali poste in opera a partire dalla fine del XIX secolo per contrastare l'erosione costiera. Tali strutture sono in grado di fornire una larga varietà di ambienti necessari all‟insediamento e alla colonizzazione (Rindi et al., 2002) dei popolamenti vegetali bentonici.
Il tratto che si estende a Nord del delta del Fiume Arno è stato interessato, negli ultimi cento anni, da un costante fenomeno erosivo; questi eventi sono fondamentalmente dovuti ad un deficit di sedimenti, essendo l‟apporto solido del corso d‟acqua notevolmente diminuito nel tempo, fino a risultare insufficiente a compensare le perdite di materiale dovute all‟asportazione verso il largo da parte del moto ondoso (Provincia di Pisa, 2003).
4
Per cercare di contenere questo fenomeno sono state realizzate le barriere artificiali lungo tutto il tratto di costa tirrenica antistante alla zona di sviluppo del Parco di Migliarino San Rossore Massaciuccoli .
Le prime scogliere per la difesa del litorale furono costruite a Marina di Pisa nel 1898. Le barriere furono importanti nel contenere il fenomeno erosivo sulle coste di Marina di Pisa e determinarono un risultato positivo sul lobo sinistro dell‟Arno, a differenza del lobo destro che ancora oggi mostra un arretramento di circa 700 m rispetto all‟altro lobo del fiume (Provincia di Pisa, 2003).
Successivamente nel 1939 furono costruiti i pennelli di foce del Fiume Morto Nuovo che interruppero, almeno in parte, il trasporto litoraneo verso Nord, determinando così la stabilizzazione della riva meridionale rispetto alla foce del fiume, mentre sulla spiaggia settentrionale si innescarono dei processi erosivi (Provincia di Pisa, 2003) che determinarono la riduzione della stessa.
Fra il 1962 e il 1968 furono realizzate cinque scogliere parallele alla linea di costa nella località Il Gombo con scarsi effetti sull‟erosione. Negli anni successivi, infatti, il fenomeno si accentuò tanto da rendere necessario nel 1984 l‟allungamento delle quattro scogliere meridionali (Provincia di Pisa, 2003). Questo intervento generò un “effetto pennello” causando la formazione di quattro tomboli di sabbia che mettevano in collegamento le scogliere con la spiaggia circostante (Provincia di Pisa, 2003). A seguito però di alcuni intensi eventi meteo-marini nel settembre-ottobre 2005, la linea di riva subì un arretramento medio di circa dieci m. (Provincia di Pisa, 2003), esponendo maggiormente le scogliere al moto ondoso e quindi alla colonizzazione delle flore bentoniche algali.
Negli anni 2000-2001, attraverso l‟intervento “Protezione del litorale compreso fra la foce dell‟Arno ed il Gombo”, vennero realizzati nove pennelli perpendicolari alla costa: cinque pennelli di lunghezza pari a 75-80 m. per la maggior parte emergenti e con la funzione di interrompere fisicamente l‟evoluzione erosiva del litorale adiacente; gli altri quattro di lunghezza pari a 130 m. parzialmente sommersi e aventi il ruolo di contenere e intercettare il trasporto litoraneo (Provincia di Pisa, 2003).
5
Le opere costiere hanno favorito, nel tempo, l'insediamento di popolamenti macroalgali. Si può ipotizzare che le prime costruzioni realizzate a Marina di Pisa (1898) possano rappresentare dei siti con un maggior numero di specie rispetto alle strutture che sono state edificate in tempi più recenti (2000-2001) ; l‟età dei pennelli e delle barrire quindi, potrebbe determinare delle variazioni nella composizione della flora macroalgale presente.
1.2 I popolamenti macroalgali
La conoscenza della distribuzione e della struttura delle comunità bentoniche rappresenta uno strumento fondamentale per la gestione dell‟ambiente marino (Piazzi et al., 2009b). I popolamenti macroalgali, in particolare, sono stati ampiamente utilizzati per caratterizzare e monitorare gli ecosistemi costieri (Rindi & Guiry, 2003; Arevalo et al.,2007; Ballesteros et al. 2007). Le macroalghe rappresentano, infatti, la componente dominante nei popolamenti bentonici di costa rocciosa nei mari temperati, dove rivestono un ruolo fondamentale nel determinare la biomassa, la produzione e la diversità dei sistemi costieri. Inoltre rappresentano degli ottimi indicatori ecologici, dal momento che cambiamenti nella struttura di popolamenti macroalgali riflettono modificazioni ambientali sia naturali che legati alle attività umane (Piazzi et al., 2009b). Infatti, la distribuzione delle macroalghe è legata sia a fattori abiotici, come la luce, la salinità, il tipo di substrato e l‟idrodinamismo (Kautsky & Van der Maarel, 1990; Lüning, 1993; Andrew & Viejo, 1998), sia biotici, come la competizione con altri organismi bentonici e la predazione (Paine, 1990; Benedetti-Cecchi & Cinelli, 1995; Sala & Boudouresque, 1997), ma anche a fattori di disturbo antropici, quali l‟inquinamento, la degradazione degli habitat e l‟introduzione di specie alloctone (Verlaque & Fritayre, 1994; Chapman et al., 1995; Rodriguez-Prieto & Polo, 1996; Walker & Kengrick, 1998; Soltan et al., 2001; Piazzi & Balata 2008a,b). Lo studio della struttura dei popolamenti macroalgali rappresenta quindi un importante strumento per la caratterizzazione e il monitoraggio degli ecosistemi costieri (Piazzi et al., 2009b).
Bisogna ricordare che i primi studi sulla flora marina toscana risalgono al 1800 - inizio 1900, e la prima lista completa sulle alghe è stata pubblicata da Pichi nel
6
1880. Negli ultimi venticinque anni, tuttavia, molti studi condotti soprattutto dall‟Università di Pisa e di Firenze, hanno permesso di ottenere considerevoli dati sulla distribuzione, morfologia ed ecologia delle alghe toscane (Rindi et al., 2002; Piazzi et al., 2009b).
Il macrofitobenthos della Liguria è stato oggetto anch‟esso di numerosi studi nel 1800 inizio 1900 (tra gli autori: Ardissone & Strafforello, 1877; Preda, 1904, 1917). Dati recenti riguardanti questa regione sono scarsi, e spesso sono stati pubblicati in letteratura grigia (Mangialajo et al., 2004). Nel 2000 risultavano solo 116 taxa di Rhodophyta, Phaeophyta e Chlorophyta (Furnari et al. 2003). Successivamente in questa regione sono stati condotti altri studi, ed in particolar modo in un lavoro che si è svolto nel 2004 i taxa presenti erano 243 (Mangialajo
et al., 2004), questo dato evidenzierebbe quindi un aumento della biodiversità
nella regione Liguria. Questo fatto potrebbe essere correlato alla presenza di siti ad elevato interesse naturalistico che sono presenti nella regione; le Aree Marine Protette istituite infatti sono quattro: Portofino, Cinque Terre, Isola di Bergeggi e il Santuario per i Mammiferi Marini (Parks.it, 2011). Molti dei dati inediti derivano da studi effettuati nell‟ambito del Programma Afrodite, ideato dall‟ICRAM e realizzato dal CoNISMa, che aveva come scopo principale quello di effettuare un “punto zero” in tutte le Aree Marine Protette italiane e di valutare l‟effetto della protezione (Mangialajo et al., 2004).
È parso pertanto utile determinare da un punto di vista floristico la componente macroalgale bentonica del litorale pisano e in particolar modo le specie che si sono insediate nel Parco di Migliarino San Rossore Massaciuccoli.
Le macroalghe o alghe pluricellulari sono diffuse maggiormente nel dominio bentonico dove si trovano, almeno in una fase del loro ciclo biologico, attaccate ad un qualsiasi tipo di substrato duro (es. rocce, conchiglie … ecc.) oppure molle (es. sabbia). Il presente studio si è basato sull‟identificazione delle macroalghe caratterizzate dalla presenza di un tallo, cioè di uno pseudotessuto; questa struttura le differenzia dalle più complesse piante terrestri costituite invece, da veri e propri tessuti formati da cellule separate fra loro da setti trasversali. Le tallofite mancano quindi di radici, fusti e foglie e presentano clorofilla a come principale pigmento fotosintetico. Un‟altra loro caratteristica è l‟assenza di cellule sterili che avvolgono le cellule riproduttive (Lee, 1999).
7
La riproduzione può essere sia asessuale, sia sessuale. La prima non presenta fusione di gameti e può avvenire per frammentazione, cioè attraverso la rottura del tallo, seguita da accrescimento dei frammenti fino alla formazione di nuovi talli, oppure attraverso le zoospore. Queste ultime sono cellule mobili che si formano all‟interno di normali cellule vegetative o in cellule specializzate definite sporangi. Il processo dal quale si sviluppano le zoospore viene definito zoosporogenesi. In alcuni casi i gameti, che non si sono fusi a formare lo zigote, sono in grado di germinare partenogeneticamente per formare una nuova alga identica alla parentale. La riproduzione sessuale che prevede la fusione dei gameti e quindi la formazione di uno zigote si compie in tre modalità distinte: isogama quando si fondono insieme gameti simili fisiologicamente e morfologicamente e non si può parlare di gameti femminili e maschili ma solo di gameti più (+) e meno (-), quindi a polarità opposta. Il secondo tipo di riproduzione è definito anisogama se i gameti che si fondono presentano delle differenze fisiologiche e/o morfologiche, nel quale il gamete femminile è più grande e meno mobile di quello maschile. L‟ultimo tipo che si riscontra è l‟oogama che prevede la fusione di una grande oocellula immobile con uno spermatozoide di dimensioni minori e mobile (ad eccezione nelle Rhodophyta, dove gli spermi sono immobili); è il tipo di riproduzione sessuale più evoluto nel quale i gameti si sviluppano in strutture specializzate definite gametangi: l‟oocellula si origina entro l‟oogonio mentre gli spermatozoidi dentro l‟anteridio. I gameti si formano in un processo chiamato gametogenesi. Nelle specie omotalliche o monoiche i gameti di tipo diverso si formano sullo stesso tallo, mentre nelle specie eterotalliche o dioiche i gameti diversi sono prodotti su talli differenti.
L'esistenza di ogni organismo, quindi anche dei vegetali, si svolge secondo una sequenza ciclica di stadi ed eventi biologici caratteristica della specie di appartenenza. Queste sequenze costituiscono i cosiddetti cicli biologici. Nelle alghe possiamo trovate cinque differenti cicli biologici: ciclo aplonte che è considerato essere il ciclo più primitivo infatti la fase diploide è limitata al solo zigote, mentre tutto il resto del ciclo è condotto in condizioni di aploidia. Lo zigote è originato dall'unione di due cellule riproduttive aploidi, i gameti; la meiosi dello zigote porta alla formazione di quattro cellule aploidi definite meiospore. In condizioni ambientali sfavorevoli le meiospore restano quiescenti mentre il miglioramento delle condizioni ambientali porta alla germinazione di
8
quest‟ultime che, in seguito a ripetute mitosi, porteranno allo sviluppo di un individuo completo ed indipendente caratterizzato da un patrimonio genetico aploide. Il ciclo si chiude con la riproduzione dell'individuo aploide che, in questo momento, produrrà cellule generative (gameti), dalla cui unione si otterrà un nuovo zigote.
Nel ciclo diplonte la fase diploide è rappresentata dall'individuo che si sviluppa per mitosi dallo zigote; dall'unione di due cellule riproduttive aploidi rappresentate dai gameti, prodotte per meiosi in organi sessuali specializzati dell'organismo adulto che ha raggiunto la maturità sessuale, si origina lo zigote. La riproduzione avviene con la fusione dei gameti provenienti da organismi diversi della stessa specie (maschi e femmine). La fusione dei gameti aploidi porterà alla formazione di un nuovo individuo diploide, che presenta parte del corredo genetico di entrambi i genitori. La formazione del nuovo zigote diploide porterà all'inizio di un nuovo ciclo vitale.
Nel ciclo aplodiplonte è prevista un‟alternanza di generazioni: lo sporofito che rappresentata la generazione o l‟individuo che produce le spore e quindi la fase diploide del ciclo biologico e il gametofito che invece costituisce la generazione o l‟individuo dal quale originano i gameti e quindi la fase aploide. Le generazioni che si alternano possono essere isomorfiche o omologhe se il gametofito e lo sporofito sono strutturalmente identici mentre eteromorfiche o antitetiche se si alternano un piccolo tallo aploide con un grande tallo diploide o viceversa. Queste caratteristiche sono da tenere in considerazione durante l‟identificazione delle singole specie perché individui apparentemente diversi potrebbero in realtà appartenere alla stessa specie. Sono nel caso delle alghe rosse vi è un‟alternanza di tre generazioni e per questo il ciclo biologico è chiamato tifasico o trimetagenetico. Questo particolare ciclo prevede l‟alternanza di un gametofito aploide, un carposporofito diploide e un tetrasporofito diploide.
1.2.1 Chlorophyta
Le Chlorophyta o alghe verdi sono principalmente di acqua dolce e solo il 10 % sono marine (Smith, 1955). Questo phylum comprende 11 classi, 39 ordini, 115
9
famiglie, per un totale di 4294 specie (Guiry M.D. & Guiry G.M., 2011). Le Chlorophyta comprendono alghe unicellulari e pluricellulari, queste ultime oggetto di studio in questo lavoro. Le specie marine che si possono trovare nelle acque calde tropicali o semi-tropicali risultano essere simili a quelle ritrovate in tutto il mondo. Per le alghe marine che vivono invece nelle acque fredde, si possono notare delle differenze a livello dei due emisferi a causa delle acque calde che si trovano in prossimità dell‟equatore e che fungono da barriera naturale geografica per l‟evoluzione di nuovi generi e specie (Lee, 1999).
Le cellule delle alghe verdi presentano pareti cellulari costituite da cellulosa, che risulta quindi essere il principale polisaccaride presente, tuttavia nelle Caulerpales si possono trovare anche xilano o mannosio (Huizing et al., 1979). Sono alghe eucariotiche, ma si differenziano dal resto delle alghe sempre di tipo eucariotico, per la formazione di amido come prodotto di riserva direttamente nel cloroplasto grazie alla presenza nello stesso, di un pirenoide. Nel genere Caulerpa è invece l‟amiloplasto che produce amido sotto forma di granuli (Lee, 1999). I cloroplasti sono circondati solo da un involucro costituito da una doppia membrana; i tilacoidi sono raggruppati in bande di 3-5 tilacoidi e mancano i grana. Nelle alghe verdi si possono trovare dei pigmenti fotosintetici simili a quelli delle piante superiori, in quanto sono presenti clorofilla a e b e la luteina come carotenoide essenziale. I carotenoidi si formano nel cloroplasto e a causa di una deficienza di azoto si possono accumulare fuori dallo stesso e cioè nel citoplasma, per questa ragione queste alghe si possono anche colorare di rosso-arancione. I carotenoidi extra-plastidici sono elementi che spesso si riscontrano negli zigoti e quindi anche nelle strutture riproduttive sessuali (Lee, 1999). Le Chlorophyta presentano riproduzione asessuale e sessuale. La riproduzione asessuale avviene per frammentazione, mediante zoospore ma può avvenire anche attraverso la formazione di aplanospore se si tratta di cellule immobili, oppure mediante autospore se vengono prodotte spore identiche alle cellule madri. La riproduzione sessuale segue tutte e tre le modalità: isogama, anisogama e oogama; nelle specie isogame e anisogame i gameti possono essere prodotti anche dalle cellule vegetative, mentre nel caso della riproduzione oogama i gameti si formano in gametangi specializzati. Le Chlorophyta sviluppano i seguenti cicli biologici: aplonte, diplonte e aplodiplonte.
10
1.2.2 Chlorophyta più diffuse nel Mar Mediterraneo
Tra le macroalghe più diffuse in Mediterraneo troviamo l‟ordine Ulvales (Lee, 1999) con rappresentati generalmente marini. Presenta cellule uninucleate con il cloroplasto addossato alla parete. In natura, queste alghe, presentano un tallo distromatico composto da due strati di cellule oppure da un cilindro cavo con una sola parete di cellule. Il genere Ulva è un‟alga perenne e la sua proliferazione avviene in primavera. La divisione cellulare può avvenire in qualsiasi parte del tallo, ma tutte le divisioni avvengono lungo il piano perpendicolare di tutta la sua superficie. Presenta alternanza di generazioni isomorfe, con un gametofito dal quale si originano gameti biflagellati e uno sporofito che forma zoospore quadriflagellate. Le aree riproduttive si formano vicino al margine del tallo e sono ben evidenti perché la porzione fertile cambia di colorazione e da verde, ne assume una verde olivastra marrone. Il rilascio dei gameti e delle zoospore è controllato dai cicli lunari e corrisponde ai minimi o ai massimi di marea secondo le specie. Ulva è normalmente un genere marino tuttavia è possibile trovarla anche in acque salmastre, particolarmente negli estuari. Cresce su rocce nella zona intertidale e durante i mesi freddi quest‟alga colonizza ampie aree verticali mentre nei mesi caldi la sua superficie di colonizzazione si fa più ristretta, rimanendo il genere che occupa la posizione più bassa lungo la zona intertidale; in questo modo le fronde sono meno esposte e quindi meno soggette al disseccamento. È un‟alga opportunista, capace di rapida colonizzazione e crescita quando le condizioni sono favorevoli. Questo si verifica principalmente per il rapido tasso di crescita e la capacità di utilizzare nutrienti disponibili nelle acque. A causa di questa capacità
Ulva ha proliferato in molte aree sottoposte a un arricchimento di nutrienti di
origine antropica.
L‟ordine Cladophorales include individui filamentosi che presentano cellule multinulcleate e i talli possono essere ramificati o non ramificati. Il genere
Cladophora colonizza habitat sia marini sia di acque dolci e può rappresentare
una macroalga ubiquitaria nelle acque dolci di tutto il mondo. Il suo ciclo vitale è di tipo aplodiplonte con alternanza di generazioni isomorfiche. È un‟alga bentonica che può determinare un substrato utile all‟ancoraggio di molti epifiti.
11
Molto presente in Mediterraneo è anche l‟ordine Dasycladales che vive nelle acque dei mari tropicali e subtropicali. Queste macroalghe sono caratterizzate da talli molto spesso calcificati, a simmetria radiale con asse eretto e ramificazioni. Un‟altra peculiarità è la presenza di talli vegetativi uninucleati, che risultano multinucleati solo prima della riproduzione e i gameti si formano in cisti all‟interno di gametangi specializzati. Sono considerate dei “fossili viventi” e secondo la definizione di Stanley (1979) sono definiti come organismi che includono cladi ancora esistenti sopravvissuti per molti tempi geologici con una bassa diversità numerica e che hanno subito dei piccoli cambiamenti evolutivi, mostrando tutt‟oggi dei caratteri morfologici primitivi. Il genere Acetabularia ne è un esempio e vive in acque marine calde e porta ramificazioni verticillate. A maturità il tallo unicellulare produce dei raggi gametangiali, dove migrano i nuclei secondari prodotti per meiosi dal nucleo primario. Attorno ai nuclei secondari si formano dei setti che isolano diverse cisti di resistenza che in seguito si accrescono mentre i nuclei subiscono diverse divisioni. Infine i raggi gametangiali si staccano e alcune settimane dopo le cisti di resistenza, fortemente calcificate, germinano emettendo degli isogameti bi flagellati.
Un altro ordine esteso nel mar Mediterraneo è Caulerpales che include alghe marine di acque calde. I talli sono multinucleati e i nuclei non sono isolati da parete (talli cenocitici) eccetto durante la riproduzione. Presentano due carotenoidi specifici: sifonoxantina e sifoneina. Il genere Derbesia è un‟alga filamentosa che è inclusa in quest‟ordine. Si trova in acque temperate e tropicali, dove colonizza le zone ombreggiate a livello della zona intertidale e i suoi substrati sono di origine rocciosa. Il suo ciclo vitale include l‟alternanza di uno sporofito filamentoso e di un gametofito vescicolare (stadio di Halicystis) che era stato considerato inizialmente come unaalga indipendente. Lo stadio gametofitico si trova in profondità e spesso è epifita di alghe coralline. Sono alghe eterotalliche che producono un solo tipo di gameti. L‟individuo maschile presenta gametangi verde olivastro mentre per quello femminile sono di colore nero. Il rilascio di gameti è indotto dalla luce che causa un incremento della pressione di turgore e la conseguente apertura di un poro nella parete del gametangio. Lo zigote porta i nuclei maschili e femminili non fusi determinando la condizione di eterocariosi che viene mantenuta durante tutto lo sviluppo dello sporofito. Lo sporofito produce sporangi ellissoidali su corti rami laterali e dalla meiosi si originano delle
12
zoospore multiflagellate che producono i gametofiti eterotallici; negli sporangi avviene la cariogamia cioè la fusione dei nuclei genitoriali. Il genere Bryopsis fa sempre parte dell‟ordine Caulerpales (Lee, 1999): si trova principalmente in zone ombreggiate e predilige ambienti riparati dal moto ondoso. È possibile trovata come un singolo individuo oppure come raggruppamento di alcuni individui. Il tallo è costituito da un asse principale dal quale si sviluppano delle ramificazioni erette laterali. Il ciclo cellulare è aplodiplonte includendo l‟alternanza di due generazioni; il gametofito presenta le pareti cellulari che contengono una grande quantità di xilani mentre le pareti cellulari della fase sporofitica sono invece composte da mannano come principale componente fibrillare. La riproduzione asessuale avviene attraverso frammentazione, aplanospore e l‟estrusione del protoplasto, mentre la riproduzione sessuale coinvolge anisogameti biflagellati. La produzione di gameti non segue la morte dell‟intero tallo. Bryopsis comprende alcune specie che, nella stagione autunnale, si possono trovare come epifiti delle alghe coralline.
Codium è un altro genere che appartiene all‟ordine Caulerpales (Lee, 1999) e
colonizza gli ambienti marini di acque temperate e tropicali, dal limite di bassa marea fino a 70 m. di profondità. Il tallo presenta due porzioni: una prostata aderente al substrato e un‟eretta costituita da strutture cilindriche ramificate dicotomicamente. Le ramificazioni presentano una medulla centrale che si origina dall‟intersecarsi di vari filamenti; questi ultimi portano alla loro estremità delle porzioni espanse denominate utricoli, i quali vanno a circondare la medulla. La riproduzione sessuale è di tipo anisogama e dagli utricoli si originano i gametangi che rilasciano una sostanza gelatinosa con i gameti. Il genere Halimeda si trova in ambienti poco illuminati e può contribuire in modo decisivo alla calcificazione delle aree lagunari. Infatti presenta un tallo composto da segmenti calcificati separati da nodi non calcificati, anche se la deposizione di calcare è inferiore a quella osservata nelle Rhodophyta Corallinales. Presenta dei filamenti cenocitici sulla cui superficie si formano utricoli che rilasciano zoospore flagellate, ancora oggi è sconosciuto il ruolo di queste strutture asessuali.
Un altro ordine presente in Mediterraneo è Siphonocladales che include organismi marini tropicali che vivono in acque basse e calde. I talli multicellulari presentano cellule multinucleate e la riproduzione sessuale avviene per isogamia. Valonia è un genere che sviluppa un tallo vescicolato con una porzione superiore costituita
13
da una grande cellula multinucleata e una basale nella quale, per divisioni seguite da cellularizzazione, si differenzia un rizoide. La riproduzione asessuale avviene tramite la produzione di aplanospore oppure in conseguenza al rilascio di zoospore biflagellate; la fusione dei gameti non è mai stata osservata.
1.2.3 Heterokontophyta
Le Phaeophyceae, o alghe brune, appartengono al phylum delle Heterokontophyta. La classe comprende 19 ordini, 53 famiglie e 1754 specie (Guiry M.D. & Guiry G.M., 2011). Sono soprattutto alghe multicellulari che si trovano prevalentemente in ambiente marino a parte quattro generi (Heribaudiella, Pleurocladia, Bodanella, Sphacelaria) che prediligono invece le acque dolci. Molte delle Phaeophyceae colonizzano la zona dell‟intertidale e sopra di questa e sono molto abbondanti nelle regioni che presentano acque fredde, in particolar modo nell‟Emisfero Nord; l‟abbondanza è però quantitativa e non è nel numero di specie, infatti, in questo emisfero svolgono un ruolo importante nella costruzione di veri e propri habitat simili a foreste sottomarine; gli individui appartenenti all‟ordine Laminariales sono considerati le alghe costruttrici più importanti di questi ambienti potendo raggiungere anche 60 m. di lunghezza. Un altro esempio di ambiente unico costituito da una notevole biomassa di alghe brune in acque però tropicali è dovuto al genere Sargassum che vive nel mar dei Sargassi. Le Phaeophyceae appartenenti invece all‟ordine
Fucales sono sfruttate come alimento nella dieta umana. Le alghe brune devono il
loro nome alla presenza di una gran quantità del carotenoide fucoxantina nei loro cloroplasti. Questi ultimi contengono anche clorofilla a, c1 e c2 e sono circondati
da quattro membrane: due di natura lipoproteica e due di reticolo endoplasmatico. Nel cloroplasto si formano i fisodi o vescicole di fucosano che contengono dei composti fenolici sottoforma di un fluido acido e trasparente. Queste vescicole si ossidano facilmente in aria originando il pigmento ficofeina di colore marrone o nero. Il prodotto di riserva è rappresentato da una vescicola di laminarina che si trova vicina al pirenoide ed esterna al cloroplasto. La parete cellulare è formata da due componenti: la fibrillare rappresentata dalla cellulosa e l‟amorfa che comprende acido alginico e fucoidina. Le alghe che appartengono a questa classe sono filamentose, pseudoparenchimatose o parenchimatose poiché presentano dei
14
pori o dei plasmodesmi che collegano le cellule, in questo modo i protoplasmi di due cellule adiacenti sono continui. I plasmodesmi sono raggruppati nei gruppi più evoluti oppure separati in quelli più primitivi. I talli parenchimatici possono presentare rizoide, cauloide e filloide laminare; nel cauloide si sviluppano le cellule cribrose, con placche cribrose che presentano dei pori e il callosio può essere anche assente come ad esempio nel genere Laminaria; mancano però di cellule compagne che collaborano al trasposto attivo di fotosintati (principalmente mannitolo). I pori delle placche cribrose si sono evoluti dai plasmodesmi. I talli sono complessi e presentano vari tipi di accrescimento. Le alghe brune comprendono anche specie calcificate come nel caso del genere Ladina, nel quale si possono osservare dei cristalli di aragonite disposti in bande concentriche. I cicli biologici che interessano questa classe sono diploe e aplodiplonte. Nel ciclo aplodiplonte la meiosi avviene negli sporangi uniloculari e i gameti si formano sui gametangi del gametofito, ma non sempre è osservata la meiosi e la fusione dei gameti. Alcune specie oltre a produrre lo sporofito ed il gametofito, originano un tallo filamentoso microscopico detto pletismotallo che può produrre spore di vario tipo e dalle quali derivano altri pletismotalli, sporofiti o gametofiti. Durante la riproduzione sessuale tutte le attività cellulari sono controllate da ormoni sessuali, che mediano in particolare l‟emissione esplosiva degli spermatozoidi dagli anteridi e l‟attrazione degli spermatozoidi verso i gameti femminili. Gli ormoni sono differenti a seconda del genere considerato.
1.2.4 Heterokontophyta più diffuse nel Mar Mediterraneo
Nel Mediterraneo troviamo Ectocarpales, un noto ordine che presenta dei talli a filamenti liberi; il genere Ectocarpus è caratterizzato dal possedere un tallo eterotrico rappresentato dall‟insieme di un sistema strisciante e da uno eretto. Il ciclo è aplodiplonte con alternanza di generazioni isomorfiche; lo sporofito produce sporangi uni o pluriloculari, questi ultimi sporangi possono rilasciare zoospore. La cellula madre degli sporangi uniloculari è sferica e presenta un nucleo di grandi dimensioni. I cloroplasti e i nuclei degli sporangi uniloculari si dividono meioticamente e un nucleo ed un cloroplasto si possono associare e formare il rivestimento di zoozpore aploidi che producono talli gametofitici. Gli sporangi pluriloculari sono ramificazioni laterali che possono contenere fino a 660
15
cellule in seguito a ripetute divisioni mitotiche, che successivamente germinano dando origine ad un tallo sporofitico.
Un altro ordine molto conosciuto nel mar mediterraneo è Cutleriales che presenta dei talli appiattiti ad accrescimento tricotallico. Il ciclo biologico è aplodiplonte con alternanza di generazioni isomorfiche ed eteromorfi che. Il genere Cutleria vive in acque calde dell‟emisfero boreale e presenta un gametofito eretto, laminare a ramificazione dicotomica. La riproduzione è anisogama e i gameti maschili e femminili vengono prodotti su talli diversi. In seguito alla gamia, lo zigote inizia a diversi per formare lo sporofito che consiste in una struttura colonnare circondata da un‟espansione laminare aderente al substrato, sulla quale sono portati gli sporangi uniloculati. In questo genere è possibile osservare anche la partenogenesi, in quanto i gameti femminili non fecondati danno origine a dei nuovi gametofiti.
Un altro ordine diffuso nel mar Mediterraneo è Sphacelariales; queste macroalghe di colore marrone-olivastro presentano dei talli con una struttura polisifonata a ramificazione regolare e ad accrescimento di tipo apicale. La riproduzione asessuale avviene per mezzo di propaguli mentre quella sessuale è di tipo isogama ed anisogama. Il genere Dictyota è caratterizzato dal possedere talli eretti, laminari e parenchimatici. Si trova nei mari caldi di tutto il mondo ed è molto frequente anche in Mediterraneo. Presenta un‟alternanza di generazioni isomorfiche e la riproduzione asessuale avviene attraverso la formazione ed il rilascio di aplanospore, inizialmente contenute in sporangi uniloculari, isolati o raggruppati in sori. La riproduzione sessuale è oogama con un‟oosfera per oogonio e 650-1500 spermatozoidi per anteridio.
Fucales è un ordine che presenta sempre dei talli parenchimatici e riproduzione
sessuale oogama ma il ciclo biologico è di tipo diplonte. I gametofiti possono essere omotallici o eteritallici, con produzione di gameti in speciali strutture chiamate concettacoli; questi ultimo possono essere dispersi sulla superficie del tallo oppure raggruppati in ricettacoli. Anche questo ordine è molto diffuso nel Mediterraneo, oltre che nei mari di tutto il mondo. Il genere Fucus sviluppa un tallo ramificato dicotomicamente; le ramificazioni sono appiattite e mostrano una venatura centrale sporgente e ali laterali con criptoblasti che rappresentano delle depressioni con dei peli sterili che facilitano l‟assorbimento di nutrienti.
16
Presentano epidermide, cortex e medulla oltra all‟essere caratterizzati dall‟avere cisti aeree all‟apice dell ramificazioni che ne consentono il galleggiamento.
Caulerpa racemosa var. cylindracea (Sonder) Verlaque, Huisman et
Boudouresque è una Chlophyta considerata una tra le alghe invasive in Mediterraneo (Boudouresque & Verlaque, 2002); la specie è apparsa nel Bacino tra gli anni ‟80 e ‟90 dello scorso secolo ed ha rapidamente colonizzato ampie aree costiere, mostrando preoccupanti caratteristiche di invasività (Piazzi et
al.,2005; Piazzi & Balata, 2009). Caulerpa racemosa è un‟alga verde originaria
dell‟Australia e presenta una struttura stolonifera che le consente di accrescersi rapidamente su tutti i tipi di substrati e di coprire la maggior parte degli organismi bentonici mediterranei, causandone il soffocamento (Piazzi et al. 2001). Gli stoloni di C. racemosa possono costituire spessi strati che determinano la presenza di sedimento ridotto portando alla morte degli organismi inferiori (Piazzi et al., 2009a). La colonizzazione di un‟area da parte dell‟alga può causare la scomparsa di molte specie mediterranee, importanti cambiamenti nella struttura dei popolamenti bentonici e un‟erosione della biodiversità (Piazzi et al., 2009a). C.
racemosa, nonostante l‟alto grado di adattabilità alle differenti condizioni
ambientali, è tuttavia una specie fotofila che raggiunge il massimo sviluppo nelle aree più superficiali (Capiomont et al., 2005). L‟invasione di questa specie può causare una riduzione della biodiversità, danneggiando le specie sensibili e portando ad una dominanza delle specie tolleranti (Piazzi et al., 2009a).
17
1. 3 Scopo
Il lavoro trattato si è svolto lungo il litorale pisano che si estende dalla foce del canale Scolmatore d‟Arno a sud al fosso della Bufalina a nord per una lunghezza di 26 Km, interamente compresi nel tratto che va dal porto di Livorno a quella di Viareggio (Provincia di Pisa, 2003), in particolar modo si è concentrato sul censimento della flora macroalgale presente nel Parco di Migliarino San Rossore Massaciuccoli.
L‟aggiornamento delle liste delle specie dei mari italiani, della loro distribuzione e delle principali caratteristiche, assume un ruolo importante anche in relazione agli impegni del nostro Paese nell‟ambito delle convenzioni internazionali riguardanti la biodiversità ed in particolare la Convenzione di Rio del 1992 e di Barcellona del 1995 (protocollo SPA/BIO). Gli studi floristici rappresentano, inoltre, un importante contributo alla realizzazione ed implementazione della “Strategia nazionale sulla Biodiversità”.
Inoltre, la segnalazione della presenza delle specie nelle zone geografiche consente di avere un‟idea della loro distribuzione e di sapere quali aree necessitano di maggiori indagini per migliorarne le conoscenze (Furnari et
al.,2010) oltre a poter monitorare la tipologia e lo stato di colonizzazione dei
popolamenti macroalgali presenti nell‟area interessata.
È un lavoro di tipo descrittivo che può integrare le informazioni sulle flore algali che si possono desumere dai precedenti lavori riguardanti la costa toscana.
Con i dati ottenuti da questo studio preliminare condotto in Toscana si potranno eventualmente svolgere ulteriori esperimenti di tipo quantitativo sulla composizione della vegetazione bentonica della regione.
18
2. Materiali e metodi
È stata analizzata la composizione floristica del popolamento macroalgale presente lungo tutto il tratto considerato, dalla superficie alla profondità di un metro in tre periodi dell‟anno: estate, autunno, inverno. La scelta della profondità da considerare durante tutte le operazioni di raccolta del macrophytobentos non è stata casuale, ma imposta dalla localizzazione delle strutture artificiali sulle quali i popolamenti vegetali si sviluppano.
In particolar modo, infatti, i pennelli si originano direttamente dalla linea costa, essendo perpendicolari ad essa, quindi maggiore è la vicinanza alla riva, minore è la profondità alla quale le strutture si trovano collocate. Le barriere sono invece disposte parallelamente alla costa e sono posizionate ad una certa distanza dalla linea di costa; quindi in questo le strutture sono poste ad una profondità anche maggiore rispetto al metro considerato.
La scelta della profondità di campionamento di un metro è stata fatta per due motivi: 1- effettuare dei campionamenti che potessero essere paragonabili per entrambe le tipologie delle strutture artificiali; 2- in molti casi al di sotto della profondità di un metro non erano presenti macroalghe bentoniche ed il popolamento era costituito in maniera predominante dalla specie di Mytilus
galloprovincialis (Lamarck, 1819).
Durante tutte le operazioni di raccolta del materiale floristico la profondità era misurata attraverso un profondimetro.
È stato necessario un primo sopralluogo per definire i siti di campionamento in modo tale da prendere in esame tutto l‟arco costiero interessato e quindi procedere con la stesura di un disegno di campionamento.
19
2.1 Disegno di campionamento
Lo studio è stato realizzato in un tratto di costa tirrenica situata tra la Foce di Fiume Morto Nuovo, a Nord, e Tirrenia, a Sud e per un‟estensione totale di 10 Km circa.
L‟area in oggetto di studio è stata suddivisa in due zone prendendo in considerazione la presenza del fiume Arno: San Rossore che rappresenta il settore a Nord del fiume ed un settore a Sud del fiume rappresentato da Marina di Pisa. La scelta di tale suddivisione deriva da studi precedenti che hanno permesso di evidenziare la presenza sulle coste tirreniche di una corrente predominante avente direzione Sud-Nord (Millot, 1999).
Nella prima zona sono state identificate 4 località tra il limite settentrionale della costa interessata e il lato nord del delta dell‟ Arno definiti: foce Fiume Morto Nuovo, Bagni del Personale, Il Gombo e Bocca d‟ Arno lato Nord. La seconda zona, cioè Marina di Pisa, che si trova a sud rispetto al corso del fiume, è stata considerata come una unica località.
In ogni località sono presenti strutture artificiali definite come barriere, se parallele alla costa oppure pennelli, se perpendicolari alla linea di riva. Si è proceduto a prelevare le macroalghe bentoniche che si erano insediate su tali strutture seguendo questo schema: nel caso dei pennelli, sono stati identificati due siti, uno sul lato nord ed uno sul lato sud; anche nel caso delle barriere, si sono raccolte le alghe in due siti,uno sul lato esposto ed uno sul lato non esposto al moto ondoso.
Riassumendo i siti campionati in ogni zona sono: 17 a San Rossore e 13 a Marina di Pisa per un totale complessivo di 30 siti di rilevamento. Le figure 1 e 2 rappresentano i due settori nei quali si sono svolte le campagne di rilevamento. Queste carte sono state prodotte attraverso un sistema informativo computerizzato definito GIS (Geographic(al) Information System). Questo sistema consente l'acquisizione, la registrazione, l'analisi, la visualizzazione e la restituzione di informazioni derivanti da dati geografici (geo-riferiti). Sulle mappe i siti di campionamento sono indicati con una stella.
20
21
22
Fig.3: pennello a foce del fiume Morto Nuovo a San Rossore.
23
Fig. 5: pennello a Marina di Pisa.
24
Per valutare l‟andamento temporale della colonizzazione di macroalghe bentoniche sulle strutture artificiali presenti nel tratto di costa indagato sono state effettuate tre campagne di rilevamento in tre stagioni. In questo modo tutti i 30 siti sono stati replicati tre volte. In estate, nei mesi di giugno e luglio il campionamento è stato eseguito in 8 giornate; in autunno nei mesi di settembre, ottobre e novembre in 9 giornate e nei mesi di dicembre e gennaio in 9 giornate. In conclusione sono stati raccolti ed analizzati 90 campioni.
2.2 Metodo di campionamento
Il campionamento è stato effettuato nel piano infralitorale, mediante l‟utilizzo di un martello e di uno scalpello per prelevare direttamente dal substrato roccioso le macroalghe bentoniche. Una volta raccolto il materiale vegetale si è provveduto ad inserirlo in una busta di polietilene che veniva poi chiusa all‟estremità da un elastico per impedire che il materiale venisse disperso.
25
Successivamente si procedeva con l‟etichettatura di ogni busta indicando: luogo di raccolta, data e altre informazioni riguardanti il tipo di struttura dalla quale si erano raccolte le alghe (barriera parallela, pennello perpendicolare, sud e nord, esposta e non esposta).
Tutti i campioni raccolti sono stati conservati in acqua marina fino al raggiungimento del laboratorio, quindi asciugati ed infine congelati alla temperatura di -20°C
2.3 Identificazione e conservazione dei campioni
Il materiale, una volta scongelato, è stato esaminato in laboratorio attraverso l‟utilizzo di un binoculare, di un microscopio e di chiavi analitiche per identificare e definire la sistematica di tutte le alghe presenti in un determinato campione. Si è ottenuto una lista macroalgale per ogni sito che è stata appuntata su di un quaderno. Successivamente il campione identificato è stato posto in una boccetta contenente alcol oppure congelato per la sua conservazione.
Ogni boccetta è stata contrassegnata da un numero progressivo e messa in relazione alla lista di alghe determinate sul quaderno.
26
Per la nomenclatura delle specie è stato utilizzato il sito www.Algaebase.org, che costituisce il supporto più aggiornato nella determinazione tassonomica.
Un‟ altra importante fase del lavoro svolto è stata l‟allestimento di un erbario; l‟erbario è anch‟esso un metodo per la conservazione dei campioni raccolti. Per la sua creazione si procede in quest‟ordine: scongelare il materiale, prendere un foglio di carta e sistemarlo su di un sostegno di plastica. Immergere il sostegno di plastica con il foglio in un vaschetta contenete acqua; sistemare la specie da allestire sul foglio e trascinare il sostegno fuori dalla vaschetta cercando il più possibile di distendere l‟alga. Prendere il foglio sul quale si è appoggiata l‟alga e
Fig. 9: Ceramium ciliatum (J.Ellis) Ducluzeau
27
posizionarci sopra una retina, inserirlo in altri fogli asciutti e a questo punto porre il tutto sotta una pressa.
Tutti i campioni conservati con questi due metodi saranno presso l‟ Orto Botanico di Pisa con lo scopo di aumentarne le conoscenze.
2.4 Elaborazione dei dati
Un‟ analisi multivariata non-parametrica è stata effettuata sui dati di presenza-assenza della lista di specie raccolta nell‟area studiata. A tale scopo è stato utilizzato il programma PRIMER-5 (Clarke & Gorley, 2001).
La matrice di similarità tra campioni è stata calcolata utilizzando l‟indice di similarità di Bray-Curtis (Clarke, 1993) su dati non trasformati.
Non metric multidimensional scaling (nMDS) (Clark, 1993) è stato utilizzato per produrre una rappresentazione grafica bidimensionale dei siti analizzati.
28
3. Risultati
3.1 Lista floristica
Phylum Rhodophyta classe Bangiophyceae ordine Bangiales famiglia BangiaceaeBangia fuscopurpurea (Dillwyn) Lyngbye Porphyra leucosticta Thuret
ordine Goniotrichales
famiglia Goniotrichaceae
Stylonema alsidii (Zanardini) K.M.Drew classe Florideophyceae
ordine Bonnemaisoniales
famiglia Bonnemaisoniaceae
Asparagopsis armata (fase Falkenbergia rufolanosa) Harvey Bonnemaisonia hamifera Hariot
ordine Ceramiales
famiglia Callithamniaceae
Aglaothamnion sp.
Aglaothamnion tenuissimum (Bonnemaison) Feldmann-Mazoyer Callithamnion tetragonum (Withering) S.F.Gray
famiglia Ceramiaceae
Antithamnion amphigeneum A.J.K.Millar Centroceras clavulatum (C.Agardh) Montagne Ceramium ciliatum (J.Ellis) Ducluzeau
Ceramium diaphanum (Lightfoot) Roth Ceramium echionotum J.Agardh
Ceramium virgatum Roth
Pterothamnion crispum (Ducluzeau) Nägeli
29
famiglia Dasyaceae
Dasya corymbifera J.Agardh Dasya ocellata (Grateloup) Harvey Dasya rigidula (Kützing) Ardissone
Heterosiphonia crispella (C.Agardh) M.J.Wynne
famiglia Delesseriaceae
Myriogramme unistromaticum Boudouresque
famiglia Rhodomelaceae
Alsidium corallinum C.Agardh
Aphanocladia stichidiosa (Funk) Ardré Chondria dasyphylla (Woodward) C.Agardh Chondria sp.
Dipterosiphonia rigens (Shousboe ex C.Agardh) Falkenberg Herposiphonia secunda (C.Agardh) Ambronn
Herposiphonia secunda f. tenella (C.Agardh) Ambronn Herposiphonia sp.
Laurencia obtusa (Hudson) J.V.Lamouroux Osmundea pinnatifida (Hudson) Stackhouse Polysiphonia elongata (Hudson) Sprengel Polysiphonia furcellata (C.Agardh) Harvey Polysiphonia sertularioides (Grateloup) J.Agardh Polysiphonia spp.
Pterosiphonia ardreana Maggs & Hommersand Pterosiphonia pennata (C.Agardh) Sauvageau
Pterosiphonia pinnulata (Kützing) Maggs & Hommersand Symphyocladia marchantioides (Harvey) Falkenberg
famiglia Wrangeliaceae
Anotrichium furcellatum (J.Agardh) Baldock Compsothamnion thuyoides (Smith) Nägeli Griffithsia sp.
ordine Corallinales
famiglia Corallinaceae
Amphiroa cryptarthrodia Zanardini Corallina elongata J.Ellis & Solander
30
Corallina officinalis Linnaeus Jania longifurca Zanardini
Jania rubens (Linnaeus) J.V.Lamouroux
Jania squamata (Linnaeus) J.H.Kim, Guiry & H.-G.Choi
ordine Gelidiales
famiglia Gelidiaceae
Gelidium crinale var. corymbosum (Kützing) Feldmann & G.Hamel Gelidium pusillum (Stackhouse) Le Jolis
Gelidium spathulatum (Kützing) Bornet
famiglia Gelidiellaceae
Gelidiella sp.
famiglia Pterocladiaceae
Pterocladiella capillacea (S.G.Gmelin) Santelices & Hommersand
Pterocladiella melanoidea (Schousboe ex Bornet) Santelices & Hommersand
ordine Gigartinales
famiglia Caulacanthaceae
Feldmannophycus rayssiae (Feldmann & G.Feldmann) Augier &Boudouresque
famiglia Cystocloniaceae
Hypnea musciformis (Wulfen) J.V.Lamouroux Rhodophyllis divaricata (Stackhouse) Papenfuss
famiglia Gigartinaceae
Chondracanthus acicularis (Roth) Fredericq
famiglia Phyllophoraceae
Gymnogongrus griffithsiae (Turner) Martius
ordine Gracilariales
famiglia Gracilariaceae
Gracilaria sp.
Gracilariopsis longissima (S.G.Gmelin)M.Steentoft,L.M.Irvine&W.F.Farnham
ordine Halymeniales
famiglia Halymeniaceae
Grateloupia subpectinata Holmes Grateloupia turuturu Yamada
ordine Nemaliales
31
Nemalion helminthoides (Velley) Batters
ordine Plocamiales
famiglia Plocamiaceae
Plocamium cartilagineum (Linnaeus) P.S.Dixon
ordine Rhodymeniales
famiglia Faucheaceae
Gloiocladia furcata (C.Agardh) J.Agardh
famiglia Lomentariaceae
Lomentaria clavellosa (Turner) Gaillon Phylum Chlorophyta
classe Ulvophyceae ordine Bryopsidales
famiglia Bryopsidaceae
Bryopsis cupressina J.V.Lamouroux Bryopsis duplex De Notaris
Bryopsis hypnoides J.V.Lamouroux Bryopsis pennata J.V.Lamouroux Bryopsis plumosa (Hudson) C.Agardh
famiglia Caulerpaceae
Caulerpa racemosa var. cylindracea(Sonder)Verlaque,Huisman & Boudouresque
famiglia Codiaceae
Codium fragile (Suringar) Hariot
famiglia Derbesiaceae
Pedobesia simplex (Meneghini ex Kützing) M.J.Wynne & Leliaert
famiglia Udoteaceae
Pseudochlorodesmis furcellata (Zanardini) Børgesen
ordine Cladophorales
famiglia Cladophoraceae
Chaetomorpha ligustica (Kützing) Kützing Chaetomorpha aerea (Dillwyn) Kützing Chaetomorpha sp.
Cladophora albida (Nees) Kützing Cladophora dalmatica Kützing
32
Cladophora sp.
Rhizoclonium implexum (Dillwyn) Kützing
ordine Ulvales
famiglia Ulvaceae
Enteromorpha multiramosa Bliding Ulva clathrata (Roth) C.Agardh Ulva intestinalis Linnaeus Ulva linza Linnaeus Ulva rigida C.Agardh Ulva sp.
Phylum Heterokontophyta classe Phaeophyceae
ordine Scytosiphonales
famiglia Scytosiphonaceae
Colpomenia sinuosa (Mertens ex Roth) Derbès & Solier
ordine Dictyotales
famiglia Dictyotaceae
Dictyota dichotoma (Hudson) J.V.Lamouroux
ordine Sphacelariales
famiglia Sphacelariaceae
Sphacelaria fusca (Hudson) S.F.Gray
La figura 1 elenca le 92 specie identificate nell‟area di studio. Delle 92 specie trovate per 80 è stato possibile la determinazione fino al livello di specie mentre per le restanti 10 si è potuto arrivare alla determinazione del genere (Tab. 2). In particolar modo il numero di Rhodophyta risulta essere 66, il numero di Chlorophyta 23 mentre il numero di Heterokontophyta 3. All‟interno del phylum Rhodophyta sono state identificate 2 classi, 13 ordini e 25 famiglie; per il phylum Chlorophyta 1 classe, 3 ordini e 7 famiglie. Per quanto riguarda il phylum Heterokontophyta è stato possibile definire 1 classe, 3 ordini e altrettante famiglie.
33
Estate Autunno Inverno
Heterokontophyta 2 2 1
Rhodophyta 31 44 55
Chlorophyta 11 17 14
Fig.11: Numero di specie identificate all‟interno di ciascun phylum.
Tab.3: la tabella indica il numero di specie diverse nei phyla che sono state trovate nelle tre stagioni.
Fig.12: Numero di specie identificate nelle tre stagioni campionate, suddivise nei tre phyla.
34
Dal grafico mostrato in figura 12 si può notare come il numero di Rhodophyta sia più alto in tutte le tre stagioni, mentre il numero di Heterokontophyta è sempre il più basso. Un‟altra considerazione riguarda l‟aumento del numero di Rhodophyta dall‟estate all‟inverno, a differenza delle Heterokontophyta che invece aumentano dall‟inverno verso l‟estate.
La tabella 4 mostra il numero di classi, ordini e famiglie che sono presenti all‟interno di ciascun phylum. Le Chlorophyta e le Heterokontophyta hanno lo stesso numero di classi (1) e lo stesso numero di ordini (3); il numero di famiglie invece è superiore nelle Chlorophyta. Per quanto riguarda le Rhodophyta, invece, è da notare l‟ elevato numero di ordini (13) e di famiglie (25).
Phylum Chlorophyta 1 classe 3 ordini 7 famiglie PhylumHeterokontophyta 1 classe 3 ordini 3 famiglie Phylum Rhodophyta 2 classi 13 ordini 25 famiglie
35
La figura 13 indica l‟aspetto corologico relativo alla componente macroalgale del litorale pisano. È possibile distinguere cinque gruppi corologici: Ampia distribuzione (specie cosmopolite e sub cosmopolite), Atlantico, Circumtropicale, Mediterraneo e Indo-Pacifico. Le specie identificate appartengono soprattutto di Ampia distribuzione (64%), seguono le specie atlantiche e le circumtropicali (5%) ed infine le specie provenienti dal settore Indo-Pacifico risultano essere le meno presenti (4%).
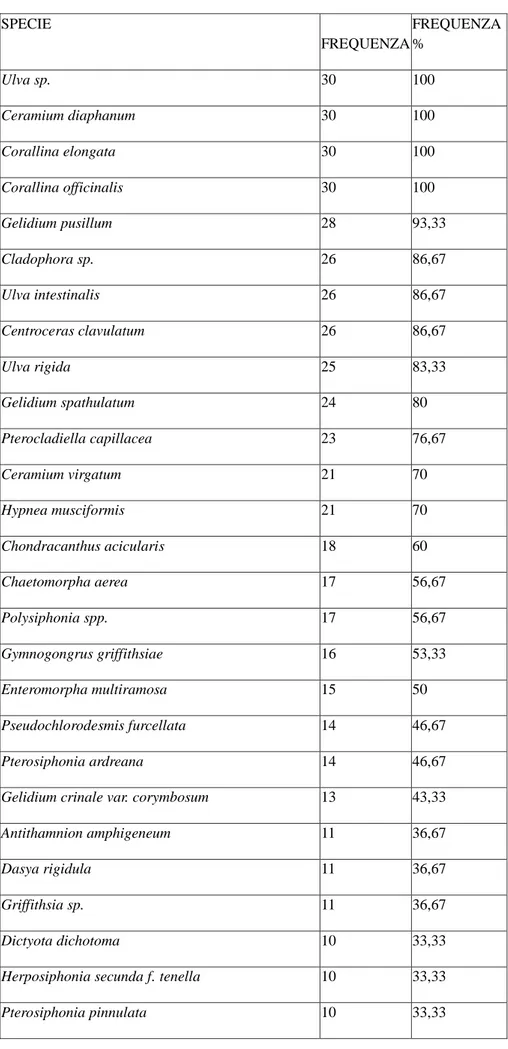
36 SPECIE FREQUENZA FREQUENZA % Ulva sp. 30 100 Ceramium diaphanum 30 100 Corallina elongata 30 100 Corallina officinalis 30 100 Gelidium pusillum 28 93,33 Cladophora sp. 26 86,67 Ulva intestinalis 26 86,67 Centroceras clavulatum 26 86,67 Ulva rigida 25 83,33 Gelidium spathulatum 24 80 Pterocladiella capillacea 23 76,67 Ceramium virgatum 21 70 Hypnea musciformis 21 70 Chondracanthus acicularis 18 60 Chaetomorpha aerea 17 56,67 Polysiphonia spp. 17 56,67 Gymnogongrus griffithsiae 16 53,33 Enteromorpha multiramosa 15 50 Pseudochlorodesmis furcellata 14 46,67 Pterosiphonia ardreana 14 46,67
Gelidium crinale var. corymbosum 13 43,33
Antithamnion amphigeneum 11 36,67
Dasya rigidula 11 36,67
Griffithsia sp. 11 36,67
Dictyota dichotoma 10 33,33
Herposiphonia secunda f. tenella 10 33,33
Pterosiphonia pinnulata 10 33,33
37
Caulerpa racemosa var. cylindracea 9 30
Jania longifurca 9 30
Bryopsis duplex 6 20
Bryopsis hypnoides 6 20
Chaetomorpha sp. 6 20
Anotrichium furcellatum 6 20
Asparagopsis armata (fase Falkenbergia
rufolanosa) 6 20 Heterosiphonia crispella 6 20 Jania rubens 6 20 Plocamium cartilagineum 6 20 Bryopsis pennata 5 16,67 Cladophora dalmatica 5 16,67 Dasya corymbifera 5 16,67 Dasya ocellata 5 16,67 Herposiphonia secunda 5 16,67 Lomentaria clavellosa 5 16,67 Myriogramme unistromaticum 5 16,67 Pterosiphonia pennata 5 16,67 Stylonema alsidii 5 16,67 Bryopsis cupressina 4 13,33 Cladophora albida 4 13,33 Aglaothamnion sp. 4 13,33 Gloiocladia furcata 4 13,33 Grateloupia subpectinata 4 13,33 Porphyra leucosticta 4 13,33 Rhodophyllis divaricata 4 13,33
Rhizoclonium riparium var. implexum 3 10
Aphanocladia stichidiosa 3 10
38 Ceramium ciliatum 3 10 Gracilariopsis longissima 3 10 Grateloupia turuturu 3 10 Jania squamata 3 10 Polysiphonia elongata 3 10 Polysiphonia sertularioides 3 10 Chaetomorpha ligustica 2 6,67 Colpomenia sinuosa 2 6,67 Aglaothamnion tenuissimum 2 6,67 Bangia fuscopurpurea 2 6,67 Ceramium echionotum 2 6,67 Chondria dasyphylla 2 6,67 Compsothamnion thuyoides 2 6,67 Gracilaria sp. 2 6,67 Herposiphonia sp. 2 6,67 Osmundea pinnatifida 2 6,67 Pterothamnion crispum 2 6,67 Bryopsis plumosa 1 3,33 Cladophora pellucida 1 3,33 Codium fragile 1 3,33 Pedobesia simplex 1 3,33 Ulva clathrata 1 3,33 Ulva linza 1 3,33 Sphacelaria fusca 1 3,33 Alsidium corallinum 1 3,33 Amphiroa cryptarthrodia 1 3,33
Bonnemaisonia hamifera (fase Trailliella intricata) 1 3,33
Chondria sp. 1 3,33
Dipterosiphonia rigens 1 3,33
39
Dalla tabella 5 è possibile individuare le specie che sono state rilevate in tutti i 30 siti in almeno una stagione. Le specie: Ceramium diaphanum, Corallina elongata,
Corallina officinalis ed il genere Ulva sp. sono risultate presenti in tutti i siti
campionati mentre le specie: Bryopsis plumosa, Cladophora pellucida, Codium
fragile, Pedobesia simplex, Ulva clathrata, Ulva linza, Sphacelaria fusca, Alsidium corallinum, Amphiroa cryptarthrodia, Bonnemaisonia hamifera (fase Trailliella intricata), Dipterosiphonia rigens, Feldmannophycus rayssiae, Laurencia obtusa, Nemalion helminthoides, Polysiphonia furcellata, Pterocladiella melanoidea, Symphyocladia marchantioides e i generi: Chondria sp. e Gelidiella sp. sono risultate presenti sono in uno dei 30 siti analizzati.
Gelidiella sp. 1 3,33 Laurencia obtusa 1 3,33 Nemalion helminthoides 1 3,33 Polysiphonia furcellata 1 3,33 Pterocladiella melanoidea 1 3,33 Symphyocladia marchantioides 1 3,33
40
3.2 Grafici nMDS
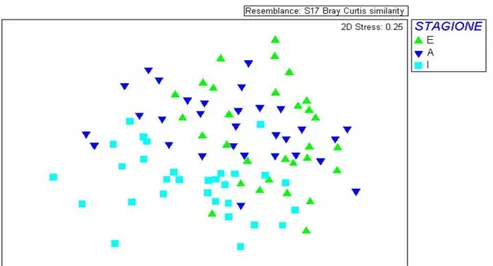
Il grafico MDS in figura 14 mostra l‟insieme dei 90 campioni suddivisi nelle tre campagne di rilevamenti floristici. Il valore di stress risulta elevato considerando l‟elevato numero di campioni analizzati. È possibile notare che i campioni invernali sono raggruppati e distaccati dai campioni che rappresentano le altre due stagioni.
Fig.14: nMDS ottenuto analizzando tutti i campioni all‟interno di ogni stagione. I simboli indicano: E = estate, A = autunno, I = inverno.
Fig. 15: nMDS ottenuto analizzando i 90 campioni che si trovano a nord del fiume Arno e a sud del fiume Arno. I simboli indicano: N = nord Arno, S = sud Arno.
41
I grafici nMDS effettuati sui siti prelevati a nord del fiume Arno (Fig.16) e a sud del fiume Arno (Fig.17) nelle tre stagioni, non mettono in evidenza differenze stagionali di rilievo, dal momento che i siti si trovano mescolati in un unico gruppo. I siti utilizzati per la elaborazione dei grafici sono 13 per i rilevamenti a sud Arno e 17 a nord Arno. Per ogni stagione il numero dei rilevamenti considera entrambe i lati dei pennelli (lato nord e lato sud) e i lati esposto e non esposto delle barriere. Il valore di stress risulta essere elevato considerando l‟elevato numero di campioni analizzati.
Fig.16: nMDS ottenuto analizzando i siti campionati a nord del fiume Arno nelle tre stagioni. E = estate, A = autunno, I = inverno.
Fig.17: nMDS ottenuto analizzando tutti i siti campionati a sud del fiume Arno nelle tre stagioni. E = estate, A = autunno, I = inverno.
42
Fig.18: nMDS ottenuto analizzando i 30 siti campionati in inverno che si trovano a nord del fiume Arno e a sud del fiume Arno. I simboli indicano: N = nord Arno, S = sud Arno.
Fig.19: il grafico indica l‟insieme dei 30 siti campionati in estate che si trovano a nord del fiume Arno e a sud del fiume Arno. I simboli indicano: N = nord Arno, S = sud Arno.
Fig.20: nMDS ottenuto analizzando i 30 siti campionati in autunno che si trovano a nord del fiume Arno e a sud del fiume Arno. N = nord Arno, S = sud Arno.
43
Dal grafico in figura 15 è possibile notare la formazione di due raggruppamenti abbastanza evidenti, che separano i 90 campioni analizzati in base alla posizione geografica rispetto alla foce del fiume Arno. E‟ possibile notare due campioni (3I e 5I) a nord dell‟ Arno più vicini al raggruppamento dei campioni che si trovano a sud dell‟ Arno;. Il valore di stress risulta essere elevato considerando l‟elevato numero di campioni analizzati. I grafici presenti in fig.18, 19 e 20 indicano i 30 siti campionati in inverno, estate ed autunno rispettivamente. Per quanto riguarda la stagione autunnale il grafico mostra una separazione tra i campioni a Nord e a Sud del fiume Arno. Non è possibile notare la stessa situazione per quanto riguarda l‟estate e l‟inverno.
Dal grafico presente in figura 21 è possibile notare che i siti campionati sui pennelli e sulle barriere non si separano a formare raggruppamenti. Il valore di stress risulta essere elevato considerando l‟elevato numero di campioni analizzati.
Fig.21: nMDS ottenuto analizzando l‟insieme dei siti campionati distinguendo tra pennelli e barriere che sono state campionate nelle tre stagioni.
44
Fig.22: nMDS ottenuto analizzando il lato nord e sud dei pennelli durante l‟inverno.
Fig.23: nMDS ottenuto analizzando il lato nord e sud dei pennelli durante l‟estate.
Fig.24: nMDS ottenuto analizzando il lato nord e sud dei pennelli durante l‟autunno.
il grafico mostra i diversi lati campionati (lato nord e lato sud) delle strutture pennelli durante l’autunno.
45
Dai grafici presenti in figura 12, 13 e 14, che considerano i siti campionati sul lato nord e sud dei pennelli nelle tre stagioni, è possibile notare l‟assenza di raggruppamenti. Il grafico ottenuto mostra un valore di stress basso, questo indica una efficace rappresentazione grafica della similarità tra siti.
Fig.25: nMDS ottenuto analizzando i lati esposti e non esposti campionati delle barriere durante l‟inverno. ne = non esposto, e = esposto.
Fig.26: nMDS ottenuto analizzando i lati esposti e non esposti campionati delle barriere durante l‟estate. ne = non esposto, e = esposto.
46
Dai grafici mostrati in figura 25, 26 e 27 che considerano i siti campionati sul lato esposto e non esposto delle barriere nelle re stagioni, è possibile notare l‟assenza di raggruppamenti. Il grafico ottenuto mostra un valore di stress basso, questo indica una efficace rappresentazione grafica della similarità tra siti.
Fig.27: nMDS ottenuto analizzando i lati esposti e non esposti campionati delle barriere durante l‟autunno. ne = non esposto, e = esposto.
47 SPECIE NORD ARNO SUD ARNO Rhodophyta Aglaothamnion sp. • • Anotrichium furcellatum • • Antithamnion amphigeneum • •
Asparagopsis armata (fase Falkenbergia rufolanosa) • •
Callithamnion tetragonum • • Centroceras clavulatum • • Ceramium diaphanum • • Ceramium virgatum • • Chondracanthus acicularis • • Corallina elongata • • Corallina officinalis • • Dasya corymbifera • • Dasya rigidula • •
Gelidium crinale var. corymbosum • •
Gelidium pusillum • •
Gelidium spathulatum • •
Gymnogongrus griffithsiae • •
Herposiphonia secunda • •
Herposiphonia secunda f. tenella • •
Herposiphonia sp. • • Hypnea musciformis • • Jania longifurca • • Jania rubens • • Lomentaria clavellosa • • Polysiphonia sertularioides • • Polysiphonia spp. • • Pterocladiella capillacea • • Pterosiphonia ardreana • • Pterosiphonia pennata • • Pterosiphonia pinnulata • • Pterothamnion crispum • • Rhodophyllis divaricata • • Stylonema alsidii • • Alsidium corallinum • Amphiroa cryptarthrodia • Bangia fuscopurpurea • Dipterosiphonia rigens • Gelidiella sp. • Gracilaria sp. •
Tab 5: lista delle specie comuni ed esclusive presenti a Nord Arno e a Sud Arno divisa nei tre phyla trovati. Il simbolo • indica la presenza.
48 Gracilariopsis longissima • Grateloupia subpectinata • Grateloupia turuturu • Jania squamata • Porphyra leucosticta • Pterocladiella melanoidea • Aglaothamnion tenuissimum • Aphanocladia stichidiosa •
Bonnemaisonia hamifera (fase Trailliella intricata) •
Ceramium ciliatum • Ceramium echionotum • Chondria dasyphylla • Chondria sp. • Compsothamnion thuyoides • Dasya ocellata • Feldmannophycus rayssiae • Gloiocladia furcata • Griffithsia sp. • Heterosiphonia crispella • Laurencia obtusa • Myriogramme unistromaticum • Nemalion helminthoides • Osmundea pinnatifida • Plocamium cartilagineum • Polysiphonia elongata • Polysiphonia furcellata • Symphyocladia marchantioides • Heterokontophyta Dictyota dichotoma • • Colpomenia sinuosa • Sphacelaria fusca • Chlorophyta Bryopsis cupressina • • Bryopsis hypnoides • • Chaetomorpha ligustica • • Chaetomorpha aerea • • Cladophora dalmatica • • Cladophora sp. • • Enteromorpha multiramosa • • Pseudochlorodesmis furcellata • • Ulva intestinalis • • Ulva rigida • • Ulva sp. • •
49 Bryopsis plumosa • Chaetomorpha sp. • Cladophora albida • Cladophora pellucida • Ulva clathrata • Ulva linza • Bryopsis duplex • Bryopsis pennata •
Caulerpa racemosa var. cylindracea •
Codium fragile •
Pedobesia simplex •
Rhizoclonium implexum •
La tabella 5 permette di evidenziare le specie che sono presenti a Nord Arno e a Sud Arno e le specie che invece sono state trovate a Nord o a Sud del delta del fiume. È possibile notare che per quanto riguarda le Rhodophyta sono 33 le specie in comune, 12 le specie identificate a Nord Arno e 21 quelle trovate a Sud Arno. Per quanto riguarda le Heterokontophyta, invece, dalla tabella è possibile rilevare la presenza di un‟ unica alga in comune e di 2 specie rilevate a Sud. Nelle Chlorophyta le specie comuni sono in totale 11, 6 le specie presenti esclusivamente a Nord Arno e 6 le specie raccolte esclusivamente a Sud Arno.