UNIVERSITA’ DEGLI STUDI di ROMA TRE
Dipartimento di Scienze
Scuola Dottorale in Geologia dell’Ambiente e delle Risorse
Dottorato di Ricerca – Ciclo XXVI
MACROEVOLUTIONARY PATTERN IN CERATOPSIAN
DINOSAURS (DINOSAURIA: ORNITHISCHIA) AND
BIOMECHANICS: AN INTEGRATED APPROACH BY MEANS OF
GEOMETRIC MORPHOMETRICS AND FINITE ELEMENT
ANALYSIS
Advisors: Candidate:
Prof. Anastassios Kotsakis Leonardo Maiorino Paolo Piras, Ph.D.
Dipartimento di Scienze, Università degli studi “Roma Tre”
Center for Evolutionary Ecology, L.go San L. Murialdo 1, 00146 - Roma Andrew A. Farke, Ph.D.
Raymond M. Alf Museum of Paleontology, 1175 West Baseline Road, Claremont, CA, 91711-2199
Organic life, we are told, has developed gradually from the protozoon to the philosopher, and this development, we are assured, is indubitably an advance. Unfortunately it is the philosopher, not the protozoon, who gives us this assurance …
[La vita organica, ci dicono, si è evoluta gradualmente dal protozoo al filosofo, e questa evoluzione, ci assicurano,
rappresenta senza dubbio un progresso. Disgraziatamente, chi ce lo assicura è il filosofo, non il protozoo …]
Bertrand Russell, Mysticism and Logic and Other Essays (Misticismo e logica), 1918
Man in his arrogance thinks himself a great work, worthy the interposition of a deity, more humble and I believe true to consider him created from animals …
[L'uomo nella sua arroganza si crede un'opera grande, meritevole di una creazione divina. Più umile, io credo sia più giusto considerarlo discendente degli animali …]
CONTENTS
1 – Project Goals p. 1
2 – Taxonomy p. 3
3 – Ornithischia p. 12
4 – Ceratopsia p. 14
5 – Evolutionary Remarks: a summary p. 33
6 – Chapter 1: Cranial and Mandibular Shape Changes during the Evolution of
Ceratopsian Dinosaurs (Dinosauria: Ornithischia) p. 36
7 – Chapter 2: The Evolution of Squamosal Shape in Ceratopsid Dinosaurs
(Dinosauria, Ornithischia) p. 127
8 – Chapter 3: Is Torosaurus Triceratops? Geometric Morphometric Evidence of
Late Maastrichtian Ceratopsid Dinosaurs p. 145
9 – Chapter 4: Evaluating Sexual Dimorphism and Allometry in Protoceratops
andrewsi (Neoceratopsia, Protoceratopsidae) p. 170
10 – Chapter 5: Mechanical Performance of the Ceratopsian Lower Jaws: a new
Insight revealed by Finite Element Analysis p. 204
11 – Acknowledgements p. 278
12 – References p. 285
13 – Image References p. 327
14 – Visited Collections p. 329
15 – Appendix I: List of the material p. 332
16 – Appendix II: Detailed description of the synthetic phylogeny p. 341
17 – Table 1S p. 352
18 – Supplementary Data 1 (Table 1S; Table 2S; Figure 1S) p. 361 19 – File S1 (Table S1; Table S2; Table S3; Table S4) p. 370
PROJECT GOALS
Exploring the evolutionary history of Ceratopsia (Dinosauria: Ornithischia) is challenging. In the past century many contributions increased the knowledge on systematics, functional
morphology and allometry (e.g. Marsh, 1888; Brown and Schlaikjer, 1940; Langston, 1966; Farlow and Dodson, 1975; Maryańska and Osmólska, 1975; Dodson, 1976; Lehman, 1990; Dodson, 1993; Chinnery, 2004a; Dodson et al., 2004; You and Dodson, 2004; Sereno et al., 2010; Sampson and Loewen, 2010; Sampson et al., 2010, 2013) of ceratopsians but did not quantitatively analyze morphological variation within ceratopsian clades or at the species level, including
macroevolutionary patterns of skull shape by means of geometric morphometrics and biomechanics. Geometric morphometrics (GM) has been used previously, in a few publications, to study cranial or postcranial shape changes of ceratopsian dinosaurs. Chapman (1990) analyzed
ontogenetic changes in skulls of Bagaceratops rozhdestvenskyi and also explored sexual
dimorphism in skulls and lower jaws of Protoceratops andrewsi by means of RFTRA (Resistant-Fit Theta-Rho-Analysis). Dodson (1993) explored the overall evolutionary changes in skulls across Ceratopsia, also using RFTRA and UPGMA (Unweighted Pair Group Method using arithmetic Average). Chinnery (2004a) used linear measurements and shape data to explore the taxonomic differences in the ceratopsian postcranial skeleton and to compare growth changes with
evolutionary changes in postcranial skeletal anatomy.
Despite these contributions, most published papers did not examine in depth the evolutionary paleobiology of Ceratopsia. Moreover, new fossil material, including extensive collections from bone beds (e.g. Ryan et al., 2001; Ryan, 2007, Wu et al., 2007; Sampson et al., 2010, 2013; Farke et al., 2011; Ryan et al., 2012a,b), has dramatically increased the number of ceratopsian species, with some taxa now represented by dozens of specimens, allowing researchers to have access to a huge sample of skulls, lower jaws and postcrania. This therefore allows a quantitatively approach to the study of morphological variation in the skull and in the lower jaw of ceratopsians.
That being so, the major goals of this project are:
1) Investigate inter-specific variability in the skull, lower jaw and squamosal within Ceratopsia.
2) Investigate morphological integration between the skull and lower jaw in different species and clades.
3) Investigate the occurrence of phenotypic evolutionary shifts of cranial and mandibular shape and size across the ceratopsian phylogeny
4) Investigate diversity and disparity of Ceratopsia through time.
5) Investigate the mandibular structural behavior, in lateral view, by means of Finite Element Analysis.
The minor goals of the project are:
1) Explore if any sexual dimorphism occurs in the skull (dorsal and lateral view) and in lower jaw of Protoceratops andrewsi.
2) Investigate allometry and cranial shape changes within Triceratops spp. and Torosaurus spp. in order to assess the taxonomic validity of these taxa.
TAXONOMY
The species list used in this work is in accordance with that presented in most part of literature by Sereno (1986), You and Dodson (2004), Dodson et al. (2004), Sampson and Loewen (2010), Sampson et al. (2010, 2013), Sereno et al. (2010), Mallon et al. (2011), Fiorillo and
Tykoski, (2012) and Ryan et al. (2012a,b) among others. In the past decades several opinions have been published concerning the validity of some taxa (e.g. Mojoceratops perifania, Magnirostris dodsoni) (Makovicky, 2002; Makovicky and Norell, 2006; Longrich, 2010). In this work I do not make any systematic review of ceratopsian taxa, but instead I just follow the systematic suggestions proposed by several previous authors (see below). Because the synonymic list of the species is very long, only the valid species names are presented here.
Ornithischia SEELEY, 1887
Marginocephalia SERENO, 1986 Ceratopsia MARSH, 1890
Yinlong downsi XU, FORSTER, CLARK ET MO, 2006
Ornithischia SEELEY, 1887
Marginocephalia SERENO, 1986 Ceratopsia MARSH, 1890
Chaoyangsauridae ZHAO, 1983
Chaoyangsaurus youngi ZHAO, CHENG ET XU, 1999
Xuanhuaceratops niei ZHAO, CHENG, XU ET MAKOVICKY, 2006
Ornithischia SEELEY, 1887
Marginocephalia SERENO, 1986 Ceratopsia MARSH, 1890
Psittacosauridae OSBORN, 1923 (a)
Hongshanosaurus houi YOU, XU et WANG, 2003 Psittacosaurus gobiensis SERENO, ZHAO ET TAN, 2010 Psittacosaurus lujiatunensis ZHOU, GAO, FOX ET CHEN, 2006 Psittacosaurus major SERENO, ZHAO, BROWN ET TAN, 2007
Psittacosaurus meileyingensis SERENO, CHAO, CHENG ET RAO, 1988 Psittacosaurus mongoliensis OSBORN, 1923 (a)
Psittacosaurus neimongoliensis RUSSELL ET ZHAO, 1996
Psittacosaurus sibiricus AVERIANOV, VORONKEVICH, LESCHCHINSKLY ET FAYNGERTZ,
2006
Psittacosaurus sinensis YOUNG, 1958
Psittacosaurus xinjangensis SERENO ET CHAO, 1988
Ornithischia SEELEY, 1887
Marginocephalia SERENO, 1986 Ceratopsia MARSH, 1890
Neoceratopsia SERENO, 1986
Liaoceratops yanzigouensis XU, MAKOVICKY, WANG, NORELL ET YOU, 2002 Yamaceratops dorngobiensis MAKOVICKY ET NORELL, 2006
Auroraceratops rugosus YOU, LI, JI, LAMANNA ET DODSON, 2005 Archaeoceratops oshimai DONG ET AZUMA, 1997
Archaeoceratops yujingziensis YOU, TANOUE ET DODSON, 2010 Helioceratops brachygnathus JIN, CHEN, ZAN ET GODEFROIT, 2009 Koreaceratops hwaseongensis LEE, RYAN ET KOBAYASHI, 2011
Marginocephalia SERENO, 1986 Ceratopsia MARSH, 1890
Neoceratopsia SERENO, 1986 Leptoceratopsidae NOPCSA, 1923
Asiaceratops salsopaludalis NESSOV, KAZNYSHKINA ET CHEREPANOV, 1989 Cerasinops hodgskissi CHINNERY ET HORNER, 2007
Gryphoceratops morrisoni RYAN, EVANS, CURRIE, BROWN ET BRINKMAN, 2012 Montanoceratops cerorhynchus (BROWN ET SCHLAIKJER, 1942)
Leptoceratops gracilis BROWN, 1914
Prenoceratops pieganensis CHINNERY, 2004 (b) Udanoceratops tschizhovi KURZANOV, 1992
Unescoceratops koppelhusae RYAN, EVANS, CURRIE, BROWN ET BRINKMAN, 2012 Zhuchengceratops inexpectus XU, WANG, ZHAO, SULLIVAN ET CHEN, 2010
Ornithischia SEELEY, 1887
Marginocephalia SERENO, 1986 Ceratopsia MARSH, 1890
Neoceratopsia SERENO, 1986
Graciliceratops mongoliensis SERENO, 2000
Ornithischia SEELEY, 1887
Marginocephalia SERENO, 1986 Ceratopsia MARSH, 1890
Neoceratopsia SERENO, 1986 Coronosauria SERENO, 1986
Ajkaceratops kozmai ÖSI, BUTLER, ET WEISHAMPEL, 2010
Bagaceratops rozhdestvenskyi MARYAŃSKA ET OSMÓLSKA, 1975 Protoceratops andrewsi GRANGER ET GREGORY, 1923
Protoceratops hellenikorhinus LAMBERT, GODEFROIT, LI, SHANG ET DONG, 2001 Magnirostris dodsoni YOU ET DONG, 2003
Ornithischia SEELEY, 1887 Marginocephalia SERENO, 1986 Ceratopsia MARSH, 1890 Neoceratopsia SERENO, 1986 Coronosauria SERENO, 1986 Ceratopsoidea HAY, 1902
Turanoceratops tardabilis NESSOV, KAZNYSHKINA ET CHEREPANOV, 1989 Zuniceratops christopheri WOLFE ET KIRKLAND, 1998
Ornithischia SEELEY, 1887 Marginocephalia SERENO, 1986 Ceratopsia MARSH, 1890 Neoceratopsia SERENO, 1986 Coronosauria SERENO, 1986 Ceratopsoidea HAY, 1902 Ceratopsidae MARSH, 1888 Centrosaurinae LAMBE, 1915
Achelousaurus horneri SAMPSON, 1995 Albertaceratops nesmoi RYAN, 2007 Avaceratops lammersi DODSON, 1986
Centrosaurus apertus LAMBE, 1904
Coronosaurus brinkmani (RYAN ET RUSSELL, 2005) Diabloceratops eatoni KIRKLAND ET DEBLIEUX, 2010 Einiosaurus procurvicornis SAMPSON, 1995
Nasutoceratops titusi, SAMPSON, LUND, LOEWEN, FARKE ET CLAYTON, 2013 Pachyrhinosaurus canadensis STERNBERG, 1950
Pachyrhinosaurus lakustai CURRIE, LANGSTON ET TANKE, 2008 Pachyrhinosaurus perotorum FIORILLO ET TYKOSKI, 2012 Rubeosaurus ovatus (GILMORE, 1930)
Sinoceratops zhuchengensis XU, WANG, ZHAO ET LI, 2010
Spinops sternbergorum FARKE, RYAN, BARRETT, TANKE, BRAMAN, LOEWEN ET
GRAHAM, 2011
Styracosaurus albertensis LAMBE, 1913
Xenoceratops foremostensis RYAN, EVANS ET SHEPHERD, 2012
Ornithischia SEELEY, 1887 Marginocephalia SERENO, 1986 Ceratopsia MARSH, 1890 Neoceratopsia SERENO, 1986 Coronosauria SERENO, 1986 Ceratopsoidea HAY, 1902 Ceratopsidae MARSH, 1888 Chasmosaurinae LAMBE, 1915
Agujaceratops mariscalensis (LEHMAN, 1989) Anchiceratops ornatus BROWN, 1914
Bravoceratops polyphemus WICK ET LEHMAN, 2013 Chasmosaurus belli LAMBE, 1902
Chasmosaurus russelli STERNBERG, 1940
Coahuilaceratops magnacuerna LOEWEN, SAMPSON, LUND, FARKE,
AGUILLÓN-MARTÍNEZ,DE LEON, RODRÍGUEZ-DE LA ROSA, GETTY ET EBERTH, 2010 Eotriceratops xerinsularis WU, BRINKMAN, EBERTH ET BRAMAN, 2007
Kosmoceratops richardsoni SAMPSON, LOEWEN, FARKE, ROBERTS, FORSTER, SMITH ET TITUS, 2010
Mojoceratops perifania LONGRICH, 2010 Nedoceratops hatcheri (LULL, 1905)
Ojoceratops fowleri SULLIVAN ET LUCAS, 2010 Pentaceratops sternbergi OSBORN, 1923 (b)
Tatankaceratops sacrisonorum OTT ET LARSON, 2010 Torosaurus latus MARSH, 1891
Torosaurus utahensis (GILMORE, 1946) Triceratops horridus MARSH, 1889 Triceratops prorsus MARSH, 1890
Utahceratops gettyi SAMPSON, LOEWEN, FARKE, ROBERTS, FORSTER, SMITH ET TITUS,
2010
Vagaceratops irvinensis (HOLMES, FORSTER, RYAN ET SHEPHERD, 2001)
In the last decades, many contributions have reviewed select taxa and assigned them as nomina dubia. Kulceratops kulensis NESSOV, 1995 was considered as nomen dubium by You and Dodson (2004), as well as Brachyceratops montanensis GILMORE, 1914, considered as nomen dubium by Sampson et al. (1997), McDonald (2011) and Farke et al. (2011). Ceratops montanus
MARSH, 1888 was considered as nomen dubium by Dodson et al. (2004). Breviceratops kozlowskii MARIAŃSKA et OSMÓLSKA, 1975, Bainoceratops efremovi TERESCHENKO et ALIFANOV, 2003, Platyceratops tatarinovi ALIFANOV, 2003, Lamaceratops tereschenkoi ALIFANOV, 2003 and Magnirostris dodsoni YOU et DONG, 2003 all have been considered as junior synonyms of Bagaceratops by Sereno (2000), You and Dodson (2004) and Makovicky and Norell (2006).
However Magnirostris dodsoni is considered a valid taxon by Kirkland and DeBlieux (2010), Ösi et al. (2010) and Sampson et al. (2013). I follow these latter suggestions. In this work I consider Magnirostris dodsoni a valid taxon, and Ceratops montanus, Kulceratops kulensis, Brachyceratops montanensis, Breviceratops kozlowskii, Bainoceratops efremovi, Platyceratops tatarinovi,
Lamaceratops tereschenkoi as invalid taxa. Moreover I do not use the family Bagaceratopsidae, previously proposed by Alifanov (2003). Thus, Bagaceratops is considered a member of
Protoceratopsidae (You and Dong, 2003; Brusatte, 2012), supported by phylogenetic analysis. Turanoceratops tardabilis, of middle-late Turonian of Uzbekistan, has been described by Nessov and colleagues (1989). The history of this taxon is rather controversial. It was first described as a member of “Monocloniinae” and later ascribed to Centrosaurinae (Nessov, 1995), suggesting an Asian origin of the clade Ceratopsidae. However the material is fragmentary and the first
description was incomplete. Recently some authors accepted its phylogenetic position within Ceratopsidae (Chinnery and Weishampel, 1998; Sereno, 2000), others considered this taxon as nomen dubium (You and Dodson, 2004; Dodson et al., 2004). In recent literature, Sues and Averianov (2009a) redescribed the original material with additional remains, providing new
information on the phylogenetic position of this Asian taxon. The review of the material, along with cladistic analyses, confirmed the previous description of the holotype as a member of Ceratopsidae (Sues and Averianov, 2009a) and later as a basal member within Centrosaurinae (Sues and
Averianov, 2009b). However, Farke and colleagues (2009a) performed a new cladistic analysis (character-taxon matrix by Dodson et al., 2004), recovering Turanoceratops as sister to
Ceratopsidae. I follow this latter suggestion in this work, even if the phylogenetic position of Turanoceratops is still problematic due to the poor nature of the material.
In 2010, Ryan and colleagues described a new chasmosaurine taxon, Medusaceratops lokii, from the upper part of the Judith River Formation, Montana, lithologically equivalent to the Oldman Formation of Canada. It is clearly a centrosaurine based on the morphology of the
parietal-squamosal contact (Longrich, 2013). The bonebed site is located approximately at the same level of the holotype of Albertaceratops nesmoi. The holotype of Medusaceratops lokii is represented by a parietal with ornamentation and it resembles the frill morphology of Albertaceratops nesmoi. Usually the epiparietals are important for making taxonomic determination at both the subfamilies (i.e. Chasmosaurinae and Centrosaurinae). However, considering that the parietal features of Medusaceratops are similar to those of Albertaceratops, and given that these taxa have the same stratigraphic range, I consider Medusaceratops a possible synonym of Albertaceratops. Therefore, I do not consider this taxon in the synthetic phylogeny used in this project and in the performed analyses. Recently, Longrich (2013) discussed the frill morphology of Medusaceratops, and also suggested that this taxon could be Albertaceratops.
In 2011, Longrich reassessed the taxonomic status of a chasmosaurine from the upper Fruitland Formation/lower Kirkland Formation of New Mexico, late Campanian in age, previously assigned to Pentaceratops sternbergi (Lehman, 1998). Longrich considered this specimen different from other ceratopsids in having a combination of several characters and a unique autapomorphy as an extreme enlargement of the premaxillary fossa. Longrich named this taxon Titanoceratops ouranos. Wick and Lehman (2013) considered this taxon as a junior synonym of Pentaceratops sternbergii. I follow this latter taxonomic consideration in this work.
In a recent paper Longrich (2013) described a new taxon of the Judith River Formation, from Montana, middle Campanian in age. He named this new chasmosaurine Judiceratops tigris. Unfortunately the material is fragmentary and not well preserved, so I do not feel confident to
consider this taxon as valid (Farke, pers. comm.). Thus I do not include it in the phylogeny or in other analyses in this work.
In 2013, Hedrick and Dodson, explored the 3D morphology of Lujiatun psittacosaurids (Psittacosaurus major, P. lujiatunensis and Hongshanosaurus houi) by means of geometric
morphometrics. The authors hypothesized that the three Lujiatun psittacosaurid species are only one species, P. lujiatunensis. Nevertheless, when investigating a 3D structure such as a psittacosaurid skull, the taphonomic distortion represents a main concern in these kind of approaches and studies. It introduces an error in the shape under investigation. In the several presented Principal Component Analyses, or “taphomorphospaces”, P. major, P. lujiatunensis and P. sp. do not show a clear
morphological separation, suggesting similar cranial morphologies (as expected for specimens pertaining to the same genus), but H. houi is more distantly related. However, as seen in the work, distortion represents an unsolvable problem and it affects the original cranial shape variation within psittacosaurids. Moreover the absence of multivariate test to assess difference in shape and the small sample size (some species are represented by few specimens) do not permit a careful comparison among species. Therefore I do not follow the Hedrick and Dodson’s indications. P. major, P. lujiatunensis and H. houi are considered all valid species in this work.
In 2003, Rich and Vickers-Rich described ceratopsian material from Australia, early Aptian in age. These authors considered this material a new ceratopsian dinosaur and it was named
Serendipaceratops arthurcclarkei, based on a partial left ulna. It was the first time that a ceratopsian dinosaur was described in Australia and therefore from Gondwana. Usually ceratopsians were restricted to Laurasia. However, recently, Agnolin and colleagues (2010) reviewed this material and reassessed this taxon as Genasauria indet. and S. arthurcclarkei as nomen nudum. I follow this latter suggestion in this work.
Figure 1. Saurischia and Ornithischia pelves
ORNITHISCHIA
The group Dinosauria OWEN, 1842 includes two distinct clades: Saurischia SEELEY, 1888, and Ornithischia SEELEY, 1888. Saurischia are characterized by a reptile-like pelvis and Ornithischia are characterized by a superficially bird-like pelvis (Figure 1).
Ornithischia is a stem-based clade ,defined as all dinosaurs more closely related to Triceratops than to
Tyrannosaurus (Weishampel, 2004). Since the early discoveries (Owen, 1842; Marsh, 1877a,b; Brown, 1908), nearly 180 valid genera have been described and five major clades have been
defined: Ankylosauria, Stegosauria, Ornithopoda, Pachycephalosauria and Ceratopsia. The early ornithischians are known from the Carnian of Argentina (i.e., Pisanosaurus mertii) and from the Norian of South Africa (i.e. Eocursor parvus) (Butler et al., 2007a, 2010; Sereno, 2012). They were all small bipedal herbivorous dinosaurs with adaptations for fast-running (Brusatte, 2012).
Ornithischia appear to be scarce during the Late Triassic and uncommon in the Early and Middle Jurassic and, at last, much more abundant in the Late Jurassic and Cretaceous, with the maximum known diversity reached in the Campanian stage (Sereno, 1997; Weishampel, 2004; Butler et al., 2010), with several quadrupedal herbivorous groups diversified in a wide range of odd morphologies and sizes (Butler et al., 2008).
In the last decades many researches focused their efforts on the phylogeny of Ornithischia (Norman, 1984; Maryańska and Osmólska, 1985; Sereno, 1986, 1997, 1999; Butler et al., 2007a, 2008) but few studies (Sereno, 1999; Butler et al., 2008) analyzed the ornithischian phylogeny using a robust character matrix for cladistic purpose. Uncertainty still remains on the phylogenetic
positions of basal taxa. Pisanosaurus is considered the oldest ornithischian, known from South America (Butler et al., 2010; Brusatte, 2012), but the position of Lesothosaurus diagnosticus, from the Pliensbachian of South Africa, is still uncertain. Sereno (1986, 1999, 2012) considered this taxon as a basal ornithischian; on the other hand, Butler et al. (2008, 2010) considered it as a basal thyreophoran. Heterodontosaurids (e.g., Heterodontosaurus tucki from the Pliensbachian of South Africa, and Fruitadens haagarorum from the Tithonian of Colorado) were placed as a more derived ornithischian group by Sereno (1986, 1999, 2012), close to Ornithopoda. In contrast Butler et al. (2008, 2010) argued that heterodontosaurids shared many features with basal ornithischians, so they considered heterodontosaurids as the basal clade of Ornithischia. A different opinion is stated by Xu et al. (2006), considering heterodontosaurids as a clade more closely related to Ceratopsia.
The clade Genasauria SERENO, 1986 comprises the five major ornithischian clades (Sereno, 1986, 1999; Butler et al., 2008; Brusatte, 2012). Stegosauria MARSH, 1877a and Ankylosauria OSBORN, 1923a constitute the clade Thyreophora NOPCSA, 1915, while Ornithopoda MARSH, 1881, Pachycephalosauria MARYAŃSKA ET OSMÓLSKA, 1974 and Ceratopsia MARSH, 1890 constitute the clade Cerapoda SERENO, 1986. Finally, Ceratopsia and Pachycephalosauria constitute the clade Marginocephalia SERENO, 1986 (Figure 2).
CERATOPSIA
Ceratopsian dinosaurs were a dominant group of herbivores in terrestrial ecosystems during Cretaceous in Asia and North America (Dodson et al., 2004). Ceratopsia had one of the highest rates of speciation in non-avian dinosaurs (Dodson, 1993, 1996; Dodson et al., 2004; Sampson and Loewen, 2010), represented by several species described from complete or nearly complete skulls and skeletons (Marsh, 1891; Colbert, 1948; Currie and Dodson, 1984; Dodson, 1986, 1996;
Sampson, 1995; Sampson et al., 1997; Ryan and Russell, 2001; You and Dodson, 2004; Dodson et al., 2004; Sampson and Loewen, 2010; Sampson et al., 2010, 2013; Farke et al., 2011; Fiorillo and Tykoski, 2012; Ryan et al., 2012a,b). In a time span of 50–60 million years, ceratopsians diversified from small (<2 m body length) bipedal species to huge (>9 m long) quadrupedal forms (Dodson, 1993).
Ceratopsia is a clade defined as all Marginocephalia closer to Triceratops than to
Pachycephalosaurus (You and Dodson, 2004). The autapomorphies of this clade are: 1) triangular-shaped head in dorsal view; 2) a narrow anterior beak (rostral bone); 3) enlarged premaxilla; 4) high external naris separated from the premaxilla ventral border by a flat area; 5) well-developed lateral flaring of the jugal; 6) wide dorso-ventral length of the infraorbital ramus of the jugal; 7) contact of palatal extension of the maxillae rostral to the choana; and 8) broad symphysis with a strong union between dentary and predentary (Sereno, 1986; You and Dodson, 2004).
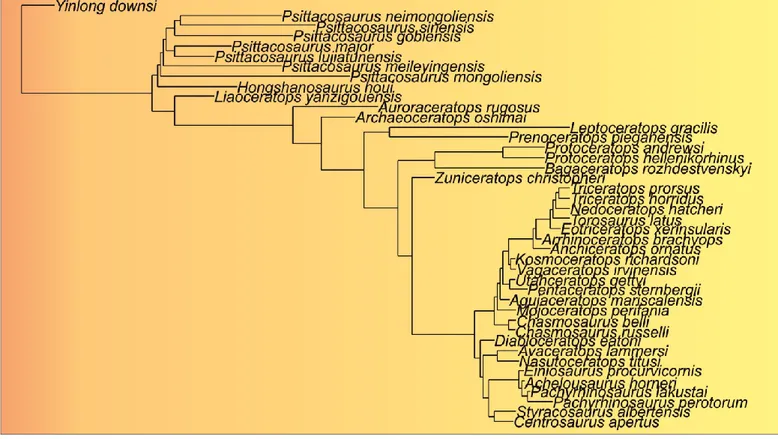
Ceratopsia comprises four well-defined families: Psittacosauridae OSBORN, 1923a,
Leptoceratopsidae NOPCSA, 1923, Protoceratopsidae GRANGER ET GREGORY, 1923, and Ceratopsidae MARSH, 1888.
The origin of this clade begins during the Late Jurassic (Oxfordian, 163.5÷157.3 Ma) in Asia, with the first representative, Yinlong downsi, from the Shishugou Formation, Xinjiang region, China (Figure 3) (Xu et al., 2006). It was a small bipedal ceratopsian (total body length = 1.2 m) characterized by a distinct fossa along the midline of the frontals, trapezoidal quadratojugal longer than deep, sharp ridges and grooves on the anterior surface of the proximal half of the paroccipital
process, a long basipterygoid process oriented posteroventrally, slit-like carotid canal laterally bordered by a lamina, a prominent tubercle on the posteroventral surangular and a vertical wear facet with a basal shelf on the premaxillary teeth (Xu et al., 2006).
Chaoyangsaurus youngi and Xuanhuaceratops niei represent the early ceratopsians after Yinlong, both taxa from the end of the Jurassic of China (Zhao et al., 1999, 2006). C. youngi is represented by a partial skull and fragmentary postcranial material from the Tuchengzi Formation, Liaoning Province, China (Tithonian, 152.1÷145 Ma) (Zhao et al., 1999, Weishampel et al., 2004, Butler et al., 2008). This taxon is distinguished by the following autapomorphies: quadratojugal overlaps posterior side of the quadrate shaft; quadrate slopes strongly anteriorly; ridge present between the planar lateral and ventral surfaces of the angular (Zhao et al., 1999; Butler et al., 2008). X. niei, the sister taxon of C. youngi, is represented by cranial and postcranial fragments of four specimens, from the Houcheng Formation, Hebei Province, China, Tithonian/Berriasian in age (Swisher et al., 2002, Zhao et al., 2006). Zhao et al. (2006) proposed the family Chaoyangsauridae for these two basal taxa.
CERATOPSIA – EARLY CRETACEOUS
The Early Cretaceous was mainly dominated by psittacosaurids. Psittacosauridae includes two genera, Psittacosaurus and Hongshanosaurus, with ten valid species, H. houi, P. mongoliensis, P. neimongoliensis, P. sinensis, P. major, P. xinjangensis, P. meileyingensis, P. lujiatunensis, P. gobiensis and P. sibiricus (Osborn, 1923a; Young, 1958; Sereno and Chao, 1988; Russell and Zhao, 1996; You et al., 2003; Lucas, 2006; Averianov et al., 2006; Zhou et al., 2006; Sereno et al., 2007, 2010; Sereno, 2010). Psittacosaurids were distributed in central Asia, ranging from eastern and western China and Mongolia to southern Siberia (You and Dodson, 2004) from the Valanginian (Wang et al., 1998; Zhou et al., 2003; Sun et al., 2011) until the Albian stage (You and Dodson, 2004; Lucas, 2006; Sereno, 2010).
Psittacosaurus (Figure 4) possesses an incipient frill, a short snout, high position of the naris, tall rostrum resembling a parrot’s
beak and reduction of the functional digits of the manus (Figure 5) (Dodson et al., 2004;
Lucas, 2006). Hongshanosaurus differs from Psittacosaurus by having a preorbital portion about half of basal skull length, and an elliptical external naris as well as an orbit and lower temporal fenestra with the major axis oriented dorsocaudally (You and Xu, 2005). They were small-bodied herbivores, bipedal and occasionally quadrupedal, which lived in semiarid and fluviolacustrine environments (You and Dodson, 2004).
Neoceratopsia SERENO, 1986 is the sister group to Psittacosauridae (Sereno, 1986; You and Dodson, 2004). This clade is defined as all ceratopsians closer to Triceratops than to
Psittacosaurus. The synapomorphies of this clade include: 1) head very large relative to body size, 2) rostral and predentary sharply keeled, 3) quadrate slopes rostrally, 4) parietal frill prominent, confluent with the supratemporal fossa, 5) exoccipitals exclude basioccipitals from the foramen magnum, 6) cervicals 1–3 fused and 7) ischium gently decurved (Sereno, 1986, You and Dodson, 2004). No horncores occur in basal neoceratopsians (You and Dodson, 2004).
Liaoceratops yanzigouensis is the first representative of Neoceratopsia (Xu et al., 2002). It was an herbivore dinosaur widespread in China, Liaoning Province, during the Hauterivian stage (~130 Ma)(You et al., 2007; Sun et al., 2011). It was comparable in size to other basal ceratopsians such as
Psittacosaurus (Xu et al., 2002) and possesses a number of primitive ceratopsian characters, such as a flat rather than a transversely expanded quadratojugal, an unkeeled rostral, the absence of an
Figure 5. Skull and skeleton of Psittacosaurus (from Lucas, 2006)
Figure 6. Holotype skull of Liaoceratops yanzigouensis
epijugal, and weakly developed primary ridges and oblique occlusion angles of the maxillary teeth (Figure 6)(You and Dodson, 2004).
Yamaceratops dorngobiensis is a basal neoceratopsian, close to Liaoceratops (Makovicky and Norell, 2006). This taxon was a small herbivore animal which lived in the Gobi Desert, Mongolia, during the Barremian stage (~128 Ma) (Shuvalov, 2000; see Jerzykiewicz and Russell, 1991 and Khand et al., 2000 for a different opinion). It was characterized by an unkeeled rostral, rostral patterned with anastomosing ridges and grooves, subtemporal part of jugal lobate and deeper than suborbital section, evidence of epijugal ossification (which appeared early in the history of Ceratopsia), angular with two tubercles along ventral edge, surangular forming vertical wall to glenoid and all cheek teeth with distinct, offset primary ridges (Makovicky and Norell, 2006). Anatomical features suggest intermediate morphology between psittacosaurids and the more derived neoceratopsians.
Auroraceratops rugosus represents a basal neoceratopsian from China (Gansu Province) of the Early Cretaceous (Albian) (Tang et al., 2001; You et al., 2005). It was a small short-faced
neoceratopsian (Figure 7), with a flat and wide skull, different from other ceratopsians in having a greater width of the nasals, a well developed rugosity of the jugal, dentary and surangular, and inflated, striated premaxillary teeth among others (You et al., 2005). Archaeoceratops is another representative of neoceratopsian dinosaurs from the Albian of China (Gansu Province) (Tang et al., 2001; You
and Dodson, 2003), the same formation as Auroraceratops rugosus (Xinminbao Group). Archaeoceratops includes two
Figure 7. Life restoration of Auroraceratops rugosus
species: A. oshimai and A. yujingziensis (Dong and Azuma, 1997; You et al., 2010). It appears to have been bipedal and quite small (about 1 meter long), with a comparatively large head.
Archaeoceratops oshimai (Figure 8) had no nasal and orbital horns, possessing only a small bony frill projecting from the back of its head. The naris is small and low, the orbit is large, the rostral is small and keeled, and the premaxilla still possesses teeth (You and Dodson, 2003, 2004).
Archaeoceratops yujingziensis differs from A. oshimai in having a laterally deflected rostral end of the maxilla, a spoon-shaped premaxillary beak with premaxillary teeth, striations on the enameled premaxillary teeth and no primary ridge on the maxillary teeth among others (You et al., 2010).
Helioceratops brachygnathus was a small neoceratopsian dinosaur from Jilin province, China, Cenomanian/Albian in age (Jin et al., 2009, 2010). Unfortunately the holotype material is really fragmentary, but it provides new evidence of the history of Neoceratopsia in Asia. The phylogenetic position is rather controversial. The phylogenetic analysis performed by Jin et al. (2009) indicates a more derived position than Archaeoceratops, in a polytomy with
Auroraceratops, Yamaceratops and Coronosauria. Nevertheless, in an incoming paper on systematics and cladistics of a new taxon from Montana (Farke et al., in prep.), Helioceratops represents the sister taxon of Archaeoceratops spp.
A new basal neoceratopsian was discovered on the Korean peninsula in 2008 (Lee et al., 2011). This new taxon is named Koreaceratops
hwaseongensis (Figure 9), found in Tando Basin, mid-west Korea, in Albian strata (~103 Ma). The holotype comprises nearly a complete tail with both ischia, partial distal hind limbs and feet. The autapomorphies concern the shape of the
astragalus and the very tall caudal neural spines, probably related to an adaptation for swimming
(Gregory and Mook, 1925; Brown and Schlaikjer, 1940; Tereschenko, 2008; Lee et al., 2011). Figure 9. Life restoration of Koreaceratops hwaseongensis
Koreaceratops occupies a phylogenetic position intermediate between Helioceratops +
Archaeoceratops and Leptoceratopsidae + more derived ceratopsians (Lee et al., 2011, Ryan et al., 2012a; Farke et al., in prep.).
CERATOPSIA – LATE CRETACEOUS
Ceratopsians are one of the most abundant groups of dinosaurs in the Late Cretaceous, particularly in the Campanian and Maastrichtian (83.6–66 Ma) of North America (Ott, 2007). Three well-represented clades occurred in Asia and North America during the Late Cretaceous, including Leptoceratopsidae, Protoceratopsidae and Ceratopsidae. These latter two families are members of Coronosauria (Sereno, 1986; Makovicky, 2001, 2002, 2010; Xu et al., 2002; Makovicky and Norell, 2006; Chinnery and Horner, 2007; Lee et al., 2011).
Members of this group are characterized by an elongated preorbital portion of the skull, an oval antorbital fossa, a triangular and caudolaterally directed supratemporal fenestra, parietal fenestrae, an expanded frill and the development of the syncervical, where atlas, axis, and several proximal cervical vertebrae fused together to support the enlarged head (Sereno, 1986; You and Dodson, 2004; You et al., 2005).
Leptoceratopsidae includes taxa closer to Leptoceratops gracilis than to Triceratops
horridus (Makovicky, 2001). This clade comprises Asian and North American taxa, known mainly in the Santonian, Campanian and Maastrichtian stages, except for Asiaceratops salsopaludalis (Nessov et al., 1989) which occurred in the Cenomanian. In the last decade, new leptoceratopsid discoveries supplemented knowledge of the fossil record and the evolution of this group (Chinnery, 2004b; Ott, 2007; Chinnery and Horner, 2007; Makovicky, 2010; Miyashita et al., 2010; Xu et al., 2010a; Ryan et al., 2012a). Asiaceratops salsopaludalis represents a basal leptoceratopsid
(Makovicky, 2010; Ryan et al., 2012a). The holotype is a fragmentary skeleton. All previously performed phylogenetic analyses (Makovicky, 2010; Ryan et al., 2012a; Farke et al., in prep.) have not had a good resolution, but its basal position within Leptoceratopsidae is accepted so far.
However some authors, in previous reviews, have been considered Asiaceratops salsopaludalis as nomen dubium (Sereno, 2000; Makovicky, 2002; You and Dodson, 2004). The fossil material represents a small herbivore leptoceratopsid dinosaur from the Cenomanian of Uzbekistan (100.5– 93.9 Ma).
After Asiaceratops, the more derived taxa within Leptoceratopsidae are:
Montanoceratops cerorhynchus (St. Mary River Formation, Montana, and Horseshoe Canyon Formation, Alberta), Cerasinops hodgskissi (Two Medicine Formation, Montana), Prenoceratops pieganensis (Two Medicine Formation, Montana, and the Oldman Formation, Alberta),
Udanoceratops tschizhovi (Djadokhta Formation, Mongolia), Leptoceratops gracilis (Scollard Formation, Alberta, Lance Formation, Wyoming, and Hell Creek Formation, Montana),
Zhuchengceratops inexpectus (from Wangshi Group, Shandong Province, China), Gryphoceratops morrisoni (Milk River Formation, Alberta) and Unescoceratops koppelhusae (Dinosaur Park Formation, Alberta) (Brown, 1914; Brown and Schlaikjer, 1942; Sternberg, 1951; Ostrom, 1978; Kurzanov, 1992; Chinnery and Weishampel, 1998; Ryan and Currie, 1998; Makovicky, 2001; Chinnery, 2004b; Chinnery and Horner, 2007; Makovicky, 2010; Miyashita et al., 2010; Ott, 2007; Xu et al., 2010a; Ryan et al., 2012a). All leptoceratopsids were quadrupedal dinosaurs, with a total body length of about 2 meters (except for U. tschizhovi with a probable body length of 4 m, and Z. inexpectus with a mandibular length of 50 cm), characterized by a lack of horns, robust jaws equipped with highly specialized large teeth, and an extremely short frill (Figure 10) (Dodson, 1996; Makovicky, 2002).
Cerasinops hodgskissi (middle-early Campanian, 80÷76.5 Ma) has two premaxillary teeth, an horizontal ridge on the jugal, a uniquely-shaped quadratojugal, short but high frill, a
narrow coronoid process, and very short and gracile forelimbs, among others, suggesting a possible bipedal locomotion (Chinnery and Horner, 2007). Montanoceratops cerorhynchus (early Maastrichtian) possesses autapomorphies such as a caudo-ventral ridge along the squamosal border, supraoccipital depression, a straight ventral border of the dentary, and the extreme elongation of some of mid-caudal neural spines (Ostrom, 1978; Makovicky, 2010). Prenoceratops
pieganensis (middle Campanian, ~76 Ma) possesses a caudally oriented naris, rostral position of pterygoid-maxilla contact, narrow postorbital bar at the frontal-postorbital contact, wide and triangular jugal, gracile quadratojugal and surangular and a reduced articular, among others (Chinnery, 2004b). Leptoceratops gracilis (latest
Maastrichtian) lacks horns and possesses a larger lacrimal, a rugose boss near the tip of coronoid Figure 11. Skull of
Leptoceratops gracilis (CMN 8887)
(not a leptoceratopsid)
process, and a dorsal face of postorbital which forms wide shelf bordering the anterior end of supratemporal fenestra (Figure 11) (Makovicky, 2002; You and Dodson, 2004). Udanoceratops tschizhovi (Campanian) is characterized by
abbreviated nasals, enlarged naris with kinked ventral border, and a dentary as deep as long (Figure 12) (Kurzanov, 1992; Sereno, 2000; Makovicky, 2002). Zhuchengceratops inexpectus (Late Campanian) is characterized by having a mandible that is extremely deep and transversely thin, an anterior end of the dentary deeper than the coronoid region, surangular
with additional fossa immediately anterior to glenoid fossa, and middle cervical ribs bifurcated due to presence of prominent accessory dorsal process among others (Xu et al., 2010a). Unescoceratops koppelhusae (middle Campanian, ~75.5 Ma) differs from all other leptoceratopsids in having a gently sloping posterior portion of the ventral margin of the dentary, a prominent chin-like flange projecting below the ventral margin and dentary teeth from midpoint of the tooth row with rounded profile (Ryan et al., 2012a). Gryphoceratops morrisoni (latest Santonian) possesses a relatively deeper jaw and a posterior border of the predentary symphysial contact which merges with the ventral margin (Ryan et al., 2012a). U. koppelhusae and G. morrisoni represent the more derived leptoceratopsids (Ryan et al., 2012a).
Graciliceratops mongoliensis represents the sister taxon of Coronosauria (Ryan et al., 2012a; Farke et al., in prep). This taxon has known a controversial history. Bohlin (1953) erected a new species, Microceratops gobiensis, based on a more prominent ridge in the dentary teeth, but this feature is shared in other neoceratopsians. Maryanska and Osmólska (1975) referred a partial skull and skeleton (ZPAL MgD-I/156), from the Gobi Desert, Mongolia, to M. gobiensis, however no autapomorphies were diagnosed for this taxon and the revision was based on symplesiomorphic features (Sereno, 2000; Makovicky, 2002). Sereno (2000), after a revision of ZPAL MgD-I/156,
Figure 12. Life restoration of Udanoceratops tschizhovi
established this taxon as the holotype of Graciliceratops mongoliensis, different from other
neoceratopsians in having a very slender median and posterior parietal frill margin, and a high tibio-femoral ratio (1.2:1). However, all these anatomical features are shared within the ontogenetic changes seen in Protoceratops andrewsi. Makovicky (2002), after a review of the fossil material, established this taxon as nomen dubium, even if, in recent scientific contributions, the status of this neoceratopsian dinosaur as a valid taxon is still accepted (You and Dodson, 2004; Butler and Sullivan, 2009; Ryan et al., 2012a; Farke et al., in prep.).
Protoceratopsidae includes 1 European taxon and 4 Asian taxa. Ajkaceratops kozmai, recently described by Ösi and colleagues (2010) (Csehbánya Formation, western Hungary,
Santonian in age), resembles Bagaceratops rozhdestvenskyi in cranial morphology. Recent cladistic analyses (Ryan et al., 2012a; Farke et al., in prep.) placed
these two species as sister taxa. The autapomorphies of this taxon include 1) the part of-- the premaxilla ventral to the external naris and the accessory fenestra is
dorsoventrally shallow relative to its rostro-caudal length, 2) the caudolateral process of the premaxilla is curved along its length becoming nearly horizontal
caudally, and 3) the buccal margin of the predentary is sharp and not beveled. Moreover, this protoceratopsid possesses a large oval accessory antorbital fenestra, between the premaxilla and maxilla, a condition also seen in Bagaceratops rozhdestvenskyi, Magnirostris dodsoni, the non-ceratopsid coronosaur Zuniceratops christopheri and the basal centrosaurine Diabloceratops eatoni (Kirkland and DeBlieux, 2010; Ösi et al., 2010). Ajkaceratops kozmai represents the result of a dispersal event during the early Late Cretaceous from Asia across the Tethys Ocean. Maryanska and Osmólska (1975) described a nearly complete skull (MgD-I/126) as a new taxon from the red beds of the Barun Goyot Formation, Mongolia (middle Campanian) and named it Bagaceratops rozhdestvenskyi (Figures 13 and 14).
Figure 13. Bagaceratops skull (from Maryanska and Osmólska, 1975)
It possesses a short parieto-squamosal frill, accessory antorbital fenestra between the premaxilla and the maxilla, a short preorbital portion of snout, a maxillary diastema, ten maxillary teeth, a long anterior
edentulous portion of the upper jaw, a deeply bifurcated posterior end of the splenial, and a shallow mandible with straight ventral margin (Maryanska and Osmólska, 1975; Makovicky, 2002; You and Dodson, 2004). Protoceratopsidae is the most commonly accepted clade (at family level), in the literature, which includes Protoceratops
andrewsi, Protoceratops hellenikorhinus, Bagaceratops rozhdestvenskyi, Magnirostris dodsoni and Ajkaceratops kozmai (You and Dodson, 2003, 2004; You and Dong, 2003; Makovicky and Norell, 2006; Ösi et al., 2010; Brusatte, 2012).
The genus Protoceratops includes two species: P. andrewsi and P. hellenikorhinus (Granger and Gregory, 1923; Lambert et al., 2001). P. andrewsi is represented by tens of skulls and
skeletons, associated with egg nests, in ontogenetic series, discovered during the Third Central Asiatic Expedition in the Gobi Desert, Djadokhta Formation (upper Campanian), Mongolia, in 1922 (Figures 15 and 16). After ninety years, over than one hundred specimens of P. andrewsi have been dug by several expeditions in Mongolia (Watabe
et al., 1996; Makovicky, 2002; Handa et al., 2012). P. andrewsi autapomorphies are: 1) a triangular rostral with a straight posterior border (in lateral view) and 2) parietal with small process projecting posteriorly developed on the anterior border of parietal fenestra (Makovicky, 2002).
P. hellenikorhinus is larger than P. andrewsi and has some distinct differences such as the absence of premaxillary teeth, two small separated nasal horns, a strong fronto-parietal depression, and a forwardly recurved frill with reentrant of the parietals along the midline (Lambert et al., 2001;
Figure 15. Skeleton of Protoceratops andrewsi
Figure 14. Life restoration of Bagaceratops
You and Dodson, 2004). Magnirostris dodsoni is a large neoceratopsian from the Late Cretaceous of Gobi Desert, which has a geographic and
stratigraphic distribution overlapping that of Protoceratops and Bagaceratops. Magnirostris possesses an elongate and robust rostral and incipient supraorbital horns, and shares with Bagaceratops and Ajkaceratops an additional antorbital fenestra.
In the Turonian (93.9–89.8 Ma), Ceratopsoidea appeared (originating in North America or Asia) with the best-known representative,
Zuniceratops christopheri, from the Moreno Hill Formation of New Mexico (Wolfe and Kirkland, 1998). The Superfamily Ceratopsoidea is a clade (Sereno, 2000) characterized by an enlarged naris, nasal horn core, expanded frill, tall cervical neural spines and a short postpubic process
(Sereno, 1986). Ceratopsoidea comprises Zuniceratops christopheri, Turanoceratops tardabilis from Uzbekistan, and Ceratopsidae from North America and Asia. Z. christopheri (Figures 17 and 18) is the oldest non-ceratopsid ceratopsian in North America, exhibiting brow horns (indicating them as a plesiomorphic condition for Ceratopsidae), absence of nasal horn, not-robust coronoid process in the dentary, as seen in Triceratops juveniles (Goodwin et al., 2006), single-rooted teeth, parietal fenestra and an expanded frill without ornamentations resembling the Protoceratops morphology (Wolfe et al., 2010).
The other representative of Ceratopsoidea is Turanoceratops tardabilis from the Bissekty Formation, Uzbekistan, of middle-late Turonian age (Nessov et al., 1989). Turanoceratops tardabilis possesses supraorbital horns situated rostrodorsal to the orbit, a shallow supracranial
Figure 17. Skeleton of Zuniceratops christopheri
Figure 16. Life restoration of Protoceratops andrewsi
cavity, a frontal excluded from the orbital margin, double-rooted teeth, and a reduced subsidiary ridge on the teeth (Sues and Averianov, 2009b). Although recent cladistic analyses placed this taxon within Centrosaurinae (Sues and Averianov, 2009b), other contributions (Farke et al., 2009a) considered this taxon as the sister to Ceratopsidae.
Ceratopsidae, the true horned dinosaurs, represents the most speciose clade in the
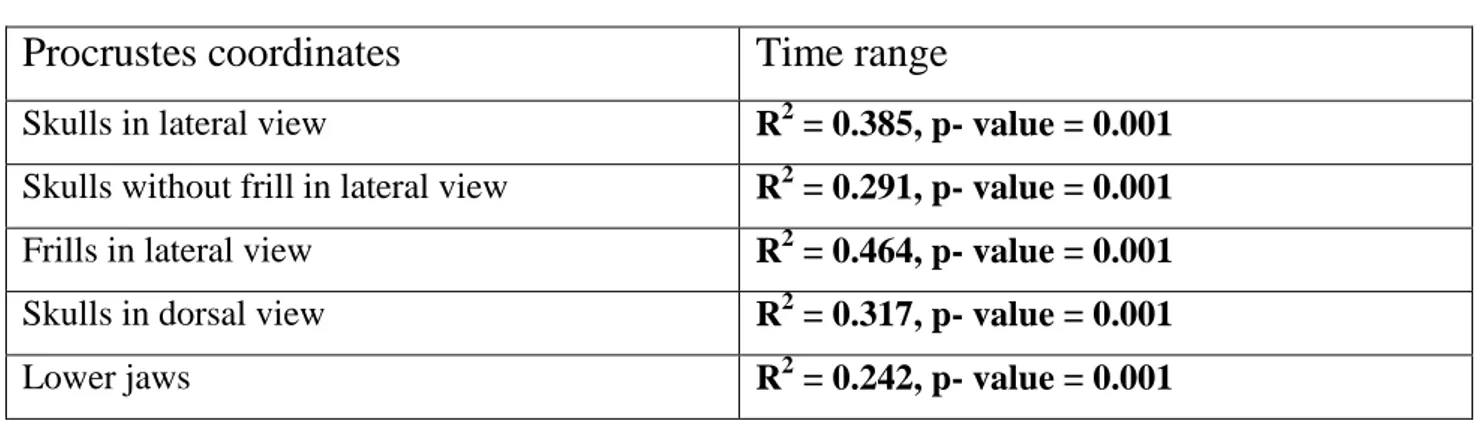
evolutionary history of Ceratopsia (Figure 19). This clade originated in North America between 90– 80 Ma (Sampson et al., 2013) when the elevated global sea levels (Western Interior Seaway) subdivided the North America continent into two distinct landmasses: an eastern region,
Appalachia, and a western region, Laramidia (Figure 20). Ceratopsid dinosaurs were part of the typical fauna of Laramidia (Sampson et al., 2010, 2013). The evolutionary radiation of ceratopsids is apparently restricted both temporally and geographically (Campanian and Maastrichtian; 83.6–66 Ma) to the Western Interior Basin of North America, with only one exception, the centrosaurine Sinoceratops zhuchengensis from the Latest Cretaceous of eastern China (Xu et al., 2010b).
Figure 18. Life restoration of Zuniceratops christopheri
In the last decades, tens of new genera and species have been formally described (e.g., Sampson, 1995; Holmes et al., 2001; Wu et al., 2007; Farke et al., 2011; Fiorillo and Tykoski, 2012; Kirkland and DeBlieux, 2010; Longrich, 2010, 2011, 2013; Ryan et al., 2012b; Sampson et al., 2010, 2013; Wick and Lehman, 2013), greatly increasing the knowledge of disparity and diversity within Ceratopsidae. Ceratopsids were huge quadrupedal (4–8 m body length),
herbivorous ornithischians recognizable by their
diagnostic bizarre cranial ornamentation, including elaborate horns and frill morphologies, and possessing skulls longer than 2.5 m in some taxa (Dodson, 1993; Dodson et al., 2004; Sampson and Loewen, 2010). Horned dinosaurs were characterized in having hypertrophied narial regions, complex dental batteries (double-rooted teeth), more than two replacement teeth in each vertical series, antorbital fenestra reduced in size, nasal and postorbital horn-cores, a reduced lacrimal,
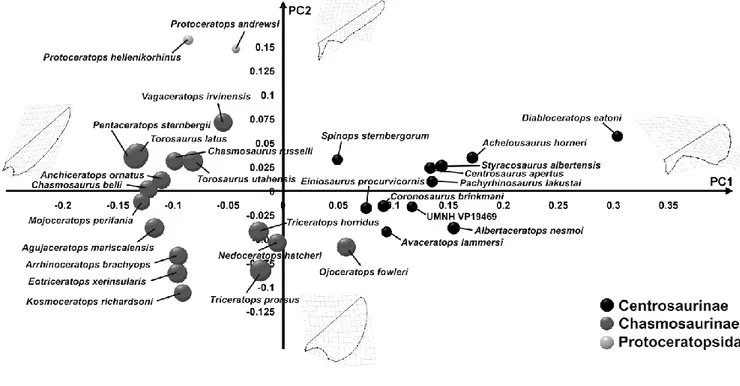
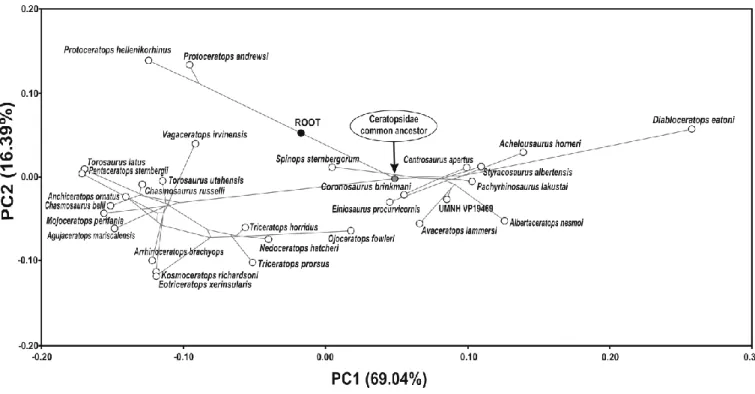
Figure 19. Cranial disparity within Ceratopsidae (Chasmosaurinae in red, Centrosaurinae in light blue)
expanded parieto-squamosal frill with a variable number of epoccipitals on the margin, short and broad sternum, and a large supracetabular process, among others (see Dodson et al., 2004 for more details; Sampson and Loewen, 2010).
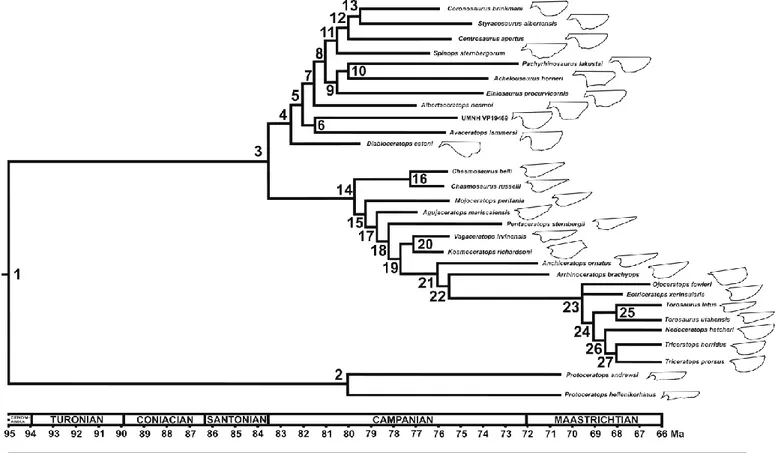
Two morphologically distinct subfamilies, Chasmosaurinae and Centrosaurinae, constitute Ceratopsidae. Chasmosaurinae comprises 19 genera and 22 species (most of these taxa are
monospecific), including problematic and controversial taxa as Tatankaceratops sacrisonorum, Titanoceratops ouranos, Judiceratops- tigris and Nedoceratops hatcheri (Ott and Larson, 2010; Farke, 2011; Scannella and Horner, 2011, Longrich and Field, 2012; Longrich, 2013; Wick and Lehman, 2013).
They were widespread from Alberta to Mexico and their time distribution was restricted to the middle Campanian and Maastrichtian (78/77 to 66 Ma).
LARAMIDIA
APPALACHIA WESTERN
INTERIOR
SEAWAY
Figure 20. Paleogeography of North America during the late Campanian (~75 Ma). Modified after Blakey, 2011
Autapomorphies of chasmosaurines include a long, low facial region, inter-premaxillary fossae, small nasal horn core, presence of postfrontal foramen, frontoparietal fontanelle walled mostly by the postorbital bones, frontal bones reduced, small prefrontal bones, elongated frill bearing triangular episquamosal bones and a strongly curved ischium, among others (Lehman, 1990; Dodson et al., 2004; Maidment and Barrett, 2011).
Centrosaurinae comprises 15 genera and 17 species (most of genera are monospecific, as seen in chasmosaurines) including one controversial taxon, Medusaceratops lokii, previously assigned to Chasmosaurinae (Ryan et al., 2010) and recently reassessed as a centrosaurine and a possible synonym of Albertaceratops nesmoi (Longrich, 2013). Centrosaurines were widespread from Alaska to Utah and restricted to middle Campanian–Maastrichtian stages (80–68 Ma). A unique exception is represented by an Asian centrosaurine, Sinoceratops zhuchengensis, from the late Campanian/early Maastrichtian of China (Xu et al., 2010b). This taxon, close to Xenoceratops foremostensis (Sampson et al., 2013), is evidence of a dispersal event following to a land
connection between North America and Asia during the Campanian (Russell, 1995; Xu et al., 2010b; Sampson and Loewen, 2010).
Autapomorphies of centrosaurines include a short and subcircular narial region, moderately elongated frill bearing squared squamosal bones, no inter-premaxillary fossae, finger-like processes of the nasal bones projecting into the naris, large nasal horn core, postfrontal fontanelle walled primarily by large frontal bones, large prefrontal bones, ulna with reduced olecranon process and a relatively straight ischium, among others (Lehman, 1990; Dodson et al., 2004; Sampson and Loewen, 2010).
Fossil discoveries during the last ten years highlighted new data on ceratopsid paleoecology. Ceratopsid bone beds (or mass death assemblage) are well documented in the Dinosaur Park
Formation (Alberta), Two Medicine Formation (Montana) and in the Aguja Formation (Texas) for Agujaceratops mariscalensis, Anchiceratops ornatus, Centrosaurus apertus, Chasmosaurus belli, Einiosaurus procurvicornis, Pachyrhinosaurus canadensis and Styracosaurus albertensis (Currie
and Dodson, 1984; Lehman, 1989; Rogers and Sampson, 1989; Horner et al., 1992; Sampson, 1995; Eberth, 1996; Sampson et al., 1997; Ryan et al., 2001). Some of these sites preserve hundreds or thousands specimens (juveniles, subadults and adults) with bone elements usually belonging to a single species (Dodson et al., 2004). All these data suggest that ceratopsids were gregarious animals, living in large groups at least part of the time (Hunt and Farke, 2010). Recent studies carried out on the biostratigraphy of Dinosaur Park Formation argued that some species likely were gregarious for a portion of the year and were more abundant in coastal habitats than inlands. This evidence suggests possible seasonal migrations of ceratopsid herds in order to seek food or for nesting (Brinkman et al., 1998; Dodson et al., 2004; Sampson and Loewen, 2010).
Regarding the function of horns and frills, several hypothesis have been formulated over the years. Traditionally, the most common idea has been that these structures were used in predator defense or thermoregulation (Wheeler, 1978; Bakker, 1986; Dodson et al., 2004; Hone et al., 2012). However the variety of horn and frill morphologies within ceratopsids and the thinness of the parieto-squamosal complex (~2mm) argued against these hypotheses (Dodson et al., 2004; Hone et al., 2012). These ornaments were mostly useless against predators. The consensus view in literature is that these structures functioned in intraspecific signaling and in male competition, using horns in combats, and in species recognition as well (Farlow and Dodson, 1975; Farke et al., 2009b; Padian and Horner, 2011).
The Late Cretaceous fossil record (in particular ceratopsids and hadrosaurids) and new stratigraphic radiometric data provide support for the hypothesis of a dinosaur provincialism for both Campanian and Maastrichtian stages (Lehman, 1997, 2001; Gates et al., 2010; Sampson and Loewen, 2010; Sampson et al., 2010, 2013): a northern (Alberta and Montana) and southern region (Colorado, Utah, New Mexico and Texas) of the Western Interior Basin, with two distinct biomes of flora and fauna. A broad fossil record along with robust stratigraphic constraints well defines the Campanian assemblages. Evidence suggests that rapid species turnover and low standing diversity characterized Ceratopsidae during the Campanian stage. Chasmosaurines and centrosaurines lived
in distinct ecological niches that persisted over approx one million of years (Sampson and Loewen, 2010; Mallon and Anderson, 2013). The presence of several factors, a high level of mate
competition within herds and the transgression–regression cycles of Western Interior Seaway, allowed the isolation of populations for extended time intervals (Horner et al., 1992; Sampson and Loewen, 2010). Therefore the occurrence of vicariance events followed a physical barrier formed around 77 Ma excluding north–south dispersals (Gates et al., 2010; Sampson et al., 2010). Natural selection but mainly sexual selection may have driven the evolutionary history of ceratopsids in northern and southern regions, allowing a rapid turnover along with low diversity of species (Sampson and Loewen, 2010).
Recent discoveries and descriptions of new ceratopsid taxa (e.g., Diabloceratops eatoni, Spinops sternbergorum, Utahceratops gettyi, Kosmoceratops richardsoni and Nasutoceratops titusi) provide additional important evidence for the dinosaur provincialism hypothesis (Kirkland and DeBlieux, 2010; Farke et al., 2011; Sampson et al., 2010, 2013).
EVOLUTIONARY REMARKS: a summary
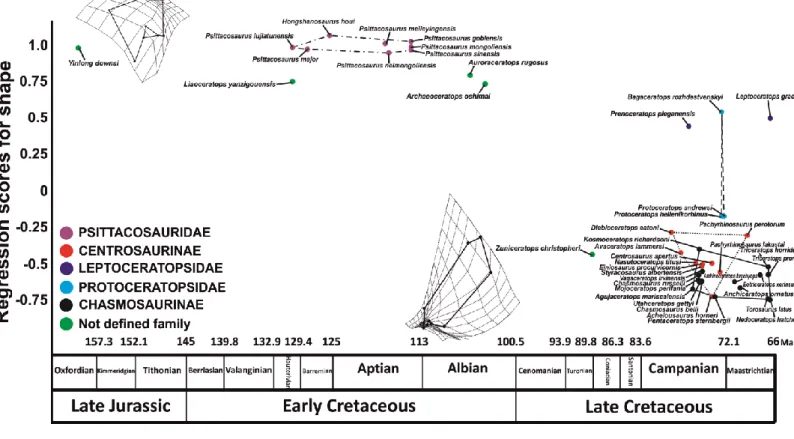
Ceratopsian dinosaurs originated in Asia during the Late Jurassic (Oxfordian) from small bipedal forms, 1.2–1.4 m in body length (Zhao et al., 1999; Xu et al., 2006), and evolved into derived quadrupedal forms of Late Cretaceous of North America (ceratopsids), 8–9 m in body length (Dodson, 1993; Dodson et al., 2004). The first part of ceratopsian history (Late Jurassic and Early Cretaceous) is characterized by bipedal and small taxa, conservative in size and similar in morphology (Sereno et al., 2010). Ceratopsians first shortened their snout, broadened their occipital region and elevated the posterior margin of the skull roof, then deepened their snout and mandible, and finally extended their frill posteriorly (Xu et al., 2006). All these cranial modifications were accompanied by an increasing of size along with postcranial changes. Ceratopsians seem to reflect the Cope-Depéret’s rule, a general tendency to increase in body size during the evolutionary history (Cope, 1885; Polly, 1998; Van Valkenburgh et al., 2004; Hone and Benton, 2005).
The progressive increase of body size involved several anatomical modifications in the postcranium: a development of skeletal robustness along with a progressive elongation of trunk and a shortening of tail and limbs to support more weight (Chinnery, 2004a; Dodson et al., 2004; Lee et al., 2011). Metatarsals became relatively shorter and wider while phalanges, including unguals, also became shorter and thicker bringing therefore a change of stance from the more upright posture of the basal neoceratopsians to a more sprawling one, especially in the forelimbs, within Ceratopsidae (Chinnery, 2004a; Lee et al., 2011). Quadrupedalism was restricted to Late Cretaceous
Coronosauria (Chinnery, 2004a; Senter, 2007; Tereschenko, 2008; Lee et al., 2011).
An important modification in the postcranium is the development of the syncervical, a complete fusion of the first four cervical vertebra to support a major cranial weight, or involved in buttress during combats or related to a feeding mechanism (Farlow and Dodson, 1975; Bakker, 1986; Ostrom and Wellnhofer, 1986; Dodson et al., 2004; Campione and Holmes, 2006; VanBuren, 2013). This anatomical feature is typical of neoceratopsians but no syncervical remains are recorded for basal members such as Liaoceratops, Yamaceratops, Auroraceratops and Archaeoceratops. It is
absent in the most basal members and in psittacosaurids (Campione and Holmes, 2006; VanBuren, 2013).
Two pairs of bone, medially and posteriorly fused parietals and antero-laterally placed squamosals, constitute the frill (Dodson and Currie, 1990; Sampson et al., 1997). In the early forms this structure served as a platform for the attachment of large jaw adductor musculature (Ostrom, 1966) and in more derived taxa probably acquired a display role or used in combats within Coronosauria (Sampson et al., 1997; Farke, 2004; Makovicky and Norell, 2006). The major modifications in the frill are marked within the clade Ceratopsidae: Chasmosaurinae, with simply adorned, elongate parieto-squamosal frills (Lambe, 1915) and Centrosaurinae, with relatively shorter, highly adorned frills (Lambe, 1915; Dodson, 1993; Ryan et al., 2001, Lund, 2010; Sampson and Loewen, 2010). These structures rapidly evolved probably partially under the influence of sexual selection.
Horns on the skull roof could have been affected by similar selection (Sampson et al., 1997; Sampson and Loewen, 2010). The presence of horns on the skull is known only in Coronosauria (Dodson et al., 2004). Within Coronosauria, the basal taxa (Protoceratops and Bagaceratops) possessed a nasal horn placed far caudal to the external naris and close to the orbit while within Ceratopsoidea, the nasal horns are placed dorsal to the external naris (Dodson et al., 2004). Within Centrosaurinae, basal members exhibited elongate supraorbital horns, short nasal horns and simple frill morphologies, whereas most derived taxa had short supraorbital horns or bosses (e.g.
Achelousaurus and Pachyrhinosaurus spp.), long nasal horns and more elaborated frill
ornamentations with the development of spikes and hooks (e.g., Centrosaurus, Styracosaurus and Rubeosaurus) (Dodson et al., 2004; Ryan et al., 2007; McDonald, 2011). On the other hand chasmosaurines, exhibited an inverse pattern. The basal taxa (e.g. Chasmosaurus spp.) bore short supraorbital horns, fairly elongate nasal horns and elaborated frill morphologies, as opposed to more derived chasmosaurines bearing long supraorbital horns, short nasal horns and simple frill morphologies (e.g., Triceratops) (Sampson and Loewen, 2010; Sampson et al., 2010, 2013).
Ceratopsoids such Zuniceratops and Turanoceratops could have been affected by the same selective pressures but better fossil material needs to improve the knowledge on the cranial
variation of these taxa.
Ceratopsians, along with the rest of non-avian dinosaurs, became extinct at the end of the Mesozoic era (66 Ma) (Dodson et al., 2004). Global climatic changes, during the Late Cretaceous, gave arise to environmental changes where dinosaurs did not succeed in adapting to the new global conditions (Brusatte, 2012). At the end of the Maastrichtian, ceratopsians already were a declining group (Dodson, 1996). A few taxa constituted the clade at that time (Eotriceratops, Ojoceratops, Nedoceratops, Torosaurus and Triceratops), centrosaurines were already extinct during the middle Maastrichtian (Sampson et al., 2013) while basal chasmosaurines disappeared at the beginning of Maastrichtian (Sampson et al., 2010), evidence that ceratopsians were already galloping along their “sunset boulevard”…
CHAPTER 1
CRANIAL AND MANDIBULAR SHAPE CHANGES DURING THE
EVOLUTION OF CERATOPSIAN DINOSAURS (DINOSAURIA:
ORNITHISCHIA)
INTRODUCTION
Understanding the evolution of phenotypic variation and the phylogenetic history of shape changes is one of the primary concerns in evolutionary biology since Darwin (Klingenberg and Gidaszewski, 2010; Piras et al., 2011). Geometric morphometrics (GM) is a suitable technique to explore morphological disparity in biological organisms. During the past twenty years GM has been used to investigate how evolutionary processes drive phenotypic variation among species through time and to better evaluate the phylogenetic relationships among related clades, allometry and mechanical performances in a phylogenetic context (Rohlf et al., 1996; Bacon, 2000; Monteiro, 2000; Chinnery, 2004a; Mitteroecker et al., 2005; Meloro et al., 2008; Marcolini et al., 2009; Adams and Nistri, 2010; Young et al., 2010; Baab et al., 2012; Bookstein, 2013; Piras et al., 2013). However, GM has been little involved in the world of dinosaur research so far. Only recently an increasing number of scientific contributions used this technique to investigate cranial shape variation within distinct groups of dinosaurs (Chapman, 1990; Dodson, 1993; Chinnery, 2004a; Brusatte et al., 2010; 2011; Campione and Evans, 2011; Foth and Rauhut, 2013; Hedrick and Dodson, 2013; Maiorino et al., 2013).
The herbivorous ceratopsians (Dinosauria, Ornithischia, Marginocephalia) represent a well-documented group of dinosaurs widespread in Laurasia and probably had one of the highest rates of speciation in non-avian dinosaurs (Dodson et al., 2004). Their evolutionary history lasted almost
100 million of years, from the early Upper Jurassic (Oxfordian) (Xu et al., 2006) until the end of the Mesozoic Era (Maastrichtian stage), when non-avian dinosaurs became extinct at the K/Pg
boundary (Dodson, 1996; Dodson et al., 2004). In the Late Jurassic and Early Cretaceous,
ceratopsians were represented by small basal taxa and mostly by psittacosaurids, whereas after the late Early Cretaceous this clade experienced a rapid radiation that occurred in Asia and, even more so, in North America. During this latter part of their history, the clade experienced both an increase in the number of species and in the variation of cranial ornamentations, including bizarre frill morphologies and development of nasal and supraorbital horns, typical of North American
ceratopsids, or the “true” horned dinosaurs of the Late Cretaceous (Chinnery, 2004a; Dodson et al., 2004; Sampson and Loewen, 2010). Thus, in a time span of 50–60 million years, ceratopsians diversified from small (<2 m body length) bipedal, occasionally quadrupedal, to huge (>9 m body length) quadrupedal forms (Sereno, 1990; Dodson, 1993, 1996; Chinnery, 2004a; Dodson et al., 2004).
Ceratopsia includes four major clades: Psittacosauridae, Protoceratopsidae,
Leptoceratopsidae, Ceratopsidae (Centrosaurinae and Chasmosaurinae), and several basal and derived taxa that are not part of distinct radiations [with the exception of a basal clade,
Chaoyangsauridae, which includes Chaoyangsaurus youngi and Xuanhuaceratops niei, from Late Jurassic of China (Zhao, 1983; Zhao et al., 1999, 2006)].
The oldest known ceratopsian is Yinlong downsi from the Oxfordian (Late Jurassic) of China. It was a small bipedal dinosaur (Xu et al., 2006). The “parrot-beaked” psittacosaurids constitute another one of the basal clades, restricted to central Asia (Siberia, China and Mongolia and possibly Thailand) during the Early Cretaceous. They were characterized in having a short skull, laterally pronounced jugal horn, short and deep snout and a scarcely developed frill (Sereno, 1990; You and Dodson, 2004; Sereno, 2010; Sereno et al., 2010). Psittacosauridae includes two genera (Psittacosaurus and Hongshanosaurus) and at least 10 valid species (You and Dodson, 2004; Sereno, 2010; Sereno et al., 2010).
Another basal clade, Leptoceratopsidae, includes quadrupedal taxa with total body lengths ranging from two to four meters. They are morphologically distinct from other clades in having a robust jaw bearing highly specialized large teeth, absence of horns and presence of a short frill (Makovicky, 2002; Xu et al., 2010a). Leptoceratopsids were widespread in central Asia (Uzbekistan and China) and, more frequently, in North America during the Late Cretaceous (Makovicky, 2002; You and Dodson, 2004; Ryan et al., 2012a).
Protoceratopsidae represents a clade that diversified during the Late Cretaceous in China and Mongolia (Lambert et al., 2001; You and Dodson, 2004; Handa et al., 2012). They were herbivorous quadrupedal dinosaurs characterized by a moderately caudodorsally developed frill, short and deep snout, prominent jugals and a nasal horn (Brown and Schlaikjer, 1940; Maryańska and Osmólska, 1975; Dodson, 1976, 1996; Lambert et al., 2001; You and Dodson, 2004; Handa et al., 2012). Dodson (1976) hypothesized sexual dimorphism in Protoceratops andrewsi, where the nasal height of the skull and frill width represented potential traits to distinguish males and females.
The clade Ceratopsidae comprises herbivorous, quadrupedal, large-bodied (5–8 m body length) ornithischian dinosaurs (Dodson et al., 2004). Their geographic range was almost entirely restricted to North America, particularly, the portion called Laramidia, formed when increasing global sea levels subdivided the North American continent in two distinct paleolandmasses: Appalachia in the East and Laramidia in the West (Sampson and Loewen, 2010; Sampson et al., 2010, 2013). Only one exception is known, the centrosaurine Sinoceratops zhuchengensis from the Latest Cretaceous of eastern China (Xu et al., 2010b). Ceratopsid evolutionary history began 90–85 million of years ago and continued up until K/Pg extinction (Dodson et al., 2004). Their rapid radiation is characterized by a variety of bizarre frill morphologies and elaborate nasal and
supraorbital horns, with some taxa having a skull exceeding 2.5 m in length (Dodson, 1993, 1996; Dodson et al., 2004).
Ceratopsidae comprises two morphologically distinct clades, Chasmosaurinae and Centrosaurinae. Chasmosaurines possess an elongated and low facial region, long supraorbital
horns, nasal horn, reduced frontals and a well caudodorsally expanded frill bearing a triangular squamosal. By contrast, centrosaurines possess a short and deep facial region, a prominent nasal process of the nasal bone, short or absent supraorbital horns (in more derived taxa), enlarged frontals, caudo-dorsally expanded frill bearing a squared squamosal (Lehman, 1990; Maidment and Barrett, 2011). The most derived centrosaurines (e.g., Pachyrhinosaurus, Achelousaurus) had a skull roof characterized by the presence of bosses (Sampson et al., 2013).
The clade Ceratopsia is characterized by several dramatic morphological changes involving firstly the cranium and secondarily the postcranium. During their evolutionary history, ceratopsians first shortened their snout, broadened their occipital region and elevated the posterior margin of the skull roof, then deepened their snout, enlarged the jugal, reduced the size of infratemporal fenestra and then shifted it ventrally, and finally developed brow and nasal horns, and extended their frill posteriorly, accompanied by an extreme elongation of squamosals and parietals, used as a platform for the large jaw adductor muscles (Dodson, 1993, 1996; Makovicky and Norell, 2006; Xu et al., 2006). Cranial changes were accompanied by several mandibular modifications. Basal members possessed a slender lower jaw bearing a short predentary, low and slender dentary with a short coronoid process and caudally elongated angular and surangular. Later, ceratopsians elongated their dentary and predentary, developed a long edentulous anterior jaw, reduced the angular and
surangular along with a dorsal elongation of the coronoid process, and displayed a progressive backward displacement of the tooth row (Ostrom, 1966; Tanoue et al., 2009a). The most derived clade (Ceratopsidae) had double-rooted dental batteries, whereas basal taxa had the primitive
condition of single-rooted functional teeth in one single row (You and Dodson, 2004; Dodson et al., 2004; Tanoue et al., 2009a, 2010).
All of these cranial changes were accompanied by an increase of size along with postcranial modifications: a development of skeletal robustness along with a progressive elongation of the trunk and a shortening of tail and limbs to support more weight (Chinnery, 2004a; Dodson et al., 2004; Lee et al., 2011). A significant modification in the neoceratopsian postcranium is the