3. Genus Allium L.
3.1 Generalities
Description
The genus Allium contains herbs, perennial, scapose, from tunicate bulbs, with onion odor and taste. (McNeal & Jacobsen, 2003). The bulbs are solitary or clustered, dividing at base, or on rhizomes, reforming annually; the outer coats are generally brown or gray, smooth, fibrous, or with cellular reticulation (generally important in identification); the inner coats are membranous. The leaves are generally withering from tip by anthesis, usually persistent, 1–12, basal; the blade is usually linear, terete, channeled, or flat, generally straight or ± falcate, generally not petiolate. The scape is usually persistent, terete or flattened. The inflorescences are umbellate, flowering centripetally (centrifugally in A. schoenoprasum), sometimes replaced totally or partially by bulbils, subtended by spathe bracts; the bracts are conspicuous, ± fused, usually 3 veined, equaling pedicel except in some introduced species, membranous. The flowers are erect, with 6 tepals, in 2 similar whorls, ± distinct, petal-like, usually becoming dry and persisting; the stamens are 6, epipetalous; the filaments are generally broad at base, fused into ring, linear, generally glabrous; the anthers and pollen are variously colored; the ovary is superior, 3-lobed, sometimes crested with processes, 3-locular, usually 2 ovules per locule, crest processes 3 or 6, smooth; style 1; stigma capitate to ± 3-lobed; the pedicel is usually erect or spreading. The fruits is capsular with loculicidal dehiscence. The seeds are black, obovoid, finely cellular-reticulate, with smooth or minutely roughened cells, with 1–8 papillae, generally without caruncle. x = 7, 8, 9.
Distribution
The genus Allium is the largest in the family Alliaceaea (species ca. 550) and it’s mainly distribuited in the Northern Hemisphere. More than 110 species occur in Europe, most of them around the Mediterranean; other centers in Asia and North America. Generally the genus is richly represented in regions that are seasonally dry (Dahlgren et al.1985).
Systematic framing
As with many other genera in the Liliaceae sensu lato, the genus Allium has been segregated into a separate family, Alliaceae, by most recent authors (Dahlgren et al., 1985; Rahn, 1998; Takhtajan, 1997; Thorne, 1992). The Alliaceae are included in the order Asparagales, superorder Liliiflorae. Old World species of Allium are generally placed in subgenera and sections. Attempts to treat New World species similarly have gained less acceptance. Traub (1972) recognized subg. Amerallium, encompassing all of the x = 7 North American members of the genus. Hanelt (1992) placed the Old World x = 7 species also in that subgenus, which includes all but three of the North American species (A. schoenoprasum, A. tricoccum, and A. victorialis), which have x = 8. These latter species have been placed in subg. Rhizirideum (Hanelt, 1992). Resolution of the problematic subgeneric and sectional relationships among Old and New World species require much more extensive molecular and phylogenetic analysis of the genus.
3.2 Allium ionicum Brullo & Tzanoud.
3.2.1 Generality
Allium ionicum Brullo & Tzanoud. (fig. 9) is a recently discovered species (Brullo & Tzanoudakis, 1994). It’s a Greek endemic unit distribuited in some of the Ionian Islands (Lefkas, Kefalonia and Ithaky). This species is common in the islands over mentioned as a member of the phrygana community. It seems to prefer the loose plant association occurring on sloopes, stony ground and semi-disused olive-groves.
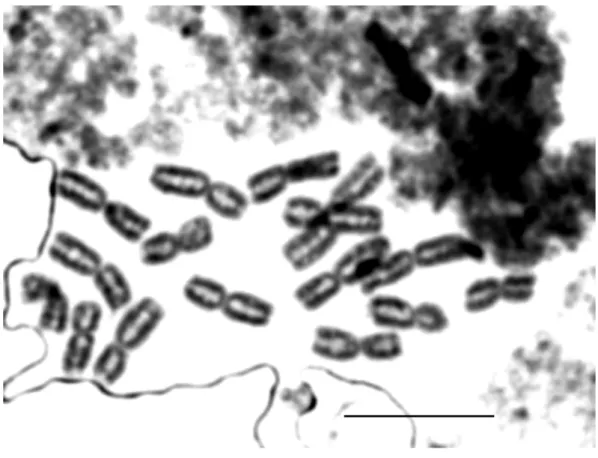
Concerning the karyological aspect, Allium ionicum is a diploid species with 2n=16.
Brullo & Tzanoudakis (1994) reported the karyotype from Lefkas
and Ithaky material, both of them consisting of 12 metacentric and 4 submetacentric chromosomes.
The karyotype from Lefkas is characterized by the longest metacentric pair being microsatellited and 2 satellited submetacentric pairs with nuclear organizers of different dimensions proximal to the telomere region of the smaller arm.
The Ithaky population shows a very similar karyogram. The only important difference is the absence of the satellites on the metacentric chromosome pair (Brullo & Tzanoudakis, 1994) Also there are some differences in the position of the satellited chromosome pairs.
A similar karyotype from Lefkas is reported by Kriemadi, Bareka, & Kamari (2002), confirming the variation in the morphology of the satellited chromosomes.
Fig. 9: Allium ionicum from Kefalonia; inflorescence in bud.
In this case the satellites are observed on the long arms of the third in size chromosome pair and on the short arms of the submetacentric chromosome pair, which, however, are not always visible. Moreover, the shortest metacentric chromosome pair bears spherical and always visible satellites, in addition to an eterozygosity in the size of their short arms. The chromosomes vary in size between 7.0 and 12.8 µm. (Kriemadi, Bareka & Kamari, 2002).
3.2.2 Results
The laboratory investigation has allowed to determinate, for both of the samples of Allium ionicum here studied, the chromosome number of 2n=16, and the karyotypes showed in the microphotographs and esemplified by the idiogrammatic formula reported in fig.10 and 11.
10 µm
2n=2x=16: 4m + 2m-SAT + 2sm-SAT + 6m + 2m-SAT
Fig. 10: Allium ionicum Brullo & Tzanoud. E98. Microphotograph of the metaphasic plate (up); relative idiogram and idiogrammatic formula (down).
10 µm
2n=2x=16: 4m + 2sm-SAT + 2m + 2m-SAT + 4m + 2m-SAT
Fig. 11: Allium ionicum Brullo & Tzanoud., K455: Microphotograph of the metaphasic plate (up); relative idiogram and idiogrammatic formula (down).
3.2.3 Discussion
The two populations studied are from one of the Ionian Islands included in the area of distribution of the species, Kefalonia, in which any karyologycal investigations don’t result until now. Most of the characteristics are in common for both of the two populations investigated, that come from different altitudes of the same island.
The karyotype formula is given for the two units:
E98: 2n=2x=16: 4m + 2m-SAT + 2sm-SAT + 6m + 2m-SAT
K455: 2n=2x=16: 4m + 2sm-SAT + 2m + 2m-SAT + 4m + 2m-SAT
The diploid karyotype with 2n=16 is confirmed, and it consists of 14 metacentric and 2 submetacentric chromosome pairs, confirming also the general prevalency of the metacentric pairs.
The submetacentric chromosome pair is satellited on the short arms for all the specimens studied, in agreement with the karyotype given for Lefkas and Ithaky.
The shortest chromosome pair bears macro-satellites well visible in both of the entities, in agree with Kriemadi & al. (2002). Brullo & Tzanoudakis (1994) show these satellites on the preceeding chromosome pair in size. The only remarkable difference between the two entities studied is the position of two satellited chromosome pairs.
Concerning the entity E98, satellites are observed on the long arms of the third in size chromosome pair, in agreement with Kriemadi & al. (2002).
The entity K455 shows satellites on the long arms too, but in the fifth chromosome pair. Moreover they have bigger size.
Also the position of the satellited submetacentric chromosome pair is variable; it’s the fourth in the unit E98, in agreement with Kriemadi & al. (2002), and it’s the third in K455.
These observations suggest the hypothesis of a similar karyotype of Allium
ionicum in the three Ionian Islands that constitute its area of distribuition, with some variations in the position of the satellited chromosome pairs and in the size of the satellites. These differences are probably originated by karyotype rearrangements. Moreover, the variations are observed not only between different islands, but also in different populations of the same island, as in the case studied from Kefalonia. In this case the two populations come from different altitudes; the entity E98 growed at 650 m and the entity K455 near to the sea level, at 90 m.
The variability in satellited chromosome pair position, and consequently the variation in the position of the secondary nuclear constriction, could be correlate to the variability in the localization of the heterochromatic bands as verified by Vosa (1976) for example in the paniculatum group, the same to which refers Allium
ionicum.
Furthermore, this variation can be observed in two samples coming from different altitudes, again in according to another research of Vosa (1996) about the correlation between heterochromatic segments (composed of highly ripetitive DNA of AT- and/or GC-nucleotide base composition), and altitude. In particular, he has studied some populations of Allium pulchellum at different altitude of Monti Pisani (Tuscany); it resulted an increase in the amount of heterochromatin in higher altitude populations, suggesting an important adaptive role of heterochromatin enabling
3.3 Allium paniculatum L.
3.3.1 Generalitiy
Allium paniculatum L. belongs to the paniculatum group, a natural group included
in the Section Codonoprasum (Rchb.) Endl. All the species of the paniculatum group are closely related and clearly distinguished from other species of Allium because of the many common characters, both morphological and karyotypical (Levan, 1937).
The group is distributed in Mediterranean and Irano-Turanian countries; in particular, the species A. paniculatum is diffused in Europe, northwards to north central France and eastwards to Slovakia and central Russia, as affirmed in Flora Europaea (1980).
With regards to the chromosome number, the group is composed by a polyploid series, with a basic chromosome number x=8. The formation of the species in nature in the paniculatum group consisted, in a large measure, in the rise of polyploid forms by crossing two forms of closely related diploid species (Levan, 1937).
The species Allium paniculatum L. was indicated as diploid by Levan (1937), while other authors reported variable levels of ploidy. Concerning the Greek territory, Tzanoudakis & Vosa (1986) indicated 2n=16, 24, 32, 40 for populations from the Greek mainland and Crete, and Karavokyrou & Tzanoudakis (1991) observed the chromosome number 2n=16, 32 in east Aegean plants. For Kalkidiki is
Fig. 12. Allium paniculatum L.; inflorescence and particular of the single flowers.
reported 2n=16 (Löve, 1979). Additionally, from Morocco has been reported 2n=16, and from Tunisia 2n=32 (Arends et al., 1979). For Madeira a tetraploid number is given (Dalgaard, 1986), and for Yugoslavia a diploid karyotype is indicated (Löve, 1980). From Italian material resulted 2n=40 (Marchi et al., 1974) and 2n=16 (Vosa, 1976). Ruiz Rejon (1986) gave 2n=16 for the Iberian Peninsula, observing also 0-2 B-chromosomes.
The chromosome complement is similar in all the forms of Allium paniculatum group; indeed only one type of chromosome occurs in the entire group, that presents two arms of approximately equal arms. As affirmed by Levan (1937), terminally or subterminally inserted chromosomes, of which at least one occurs in every chromosome set of Allium species, are entirely absent in the paniculatum group. Within the idiogram rather slight differences in size can be seen among the chromosomes. According to Levan at any rate is not possible to identify any special chromosome morphology, more than the metacentric one, in any species of the
paniculatum group. He affirmed also the general absence of satellites.
According to Flora Europaea (1980) Allium paniculatum L. includes four subspecies: ssp. paniculatum , ssp. villosulum, ssp. fuscum, ssp. euboicum.
The karyological description of the subspecies has been reported by Özhatay (1990) from Turkey. The ssp. paniculatum is described to have four pairs of metacentric and submetacentric chromosomes without any satellites and secondary constrictions. The ssp. fuscum was described as composed by five sets of metacentric and three sets of acrocentric; the ssp. villosulum consisted of eigth pairs of metacentric and one chromosome of the smallest pair with a satellite was observed. Any karyological description hasn’t been found about the ssp. euboicum, Greek endemic of the Evvia Island, east Greece.
10 µm
2n = 2x = 16: 16 m
Fig. 13 Allium paniculatum L., E 59: microphotograph of a somatic
metaphasic plate (up) and its corresponding idiogram and idiogrammatic formula (down). Scale bar = 10 µm.
3.3.3 Discussion
The present investigation has given a diploid chromosome number 2n=16 for the population of Allium paniculatum L. from Lefkas, Ionian Islands (fig. 13). This result confirms the basic chromosome number x=8 reported by several authors over mentioned from different countries.
Additionally, this basic number is the most common for the genus in the Greek territory, and especially in North and South Greek mainland, included the Ionian Islands, while the other two basic numbers of Allium, x=7 and x=11, are relatively more abundant in Crete and Kiklades.
The result obtained can be placed in the general distribution pattern of Allium cytotypes in Greece, investigated by Tzanoudakis and Vosa (1986); from their study
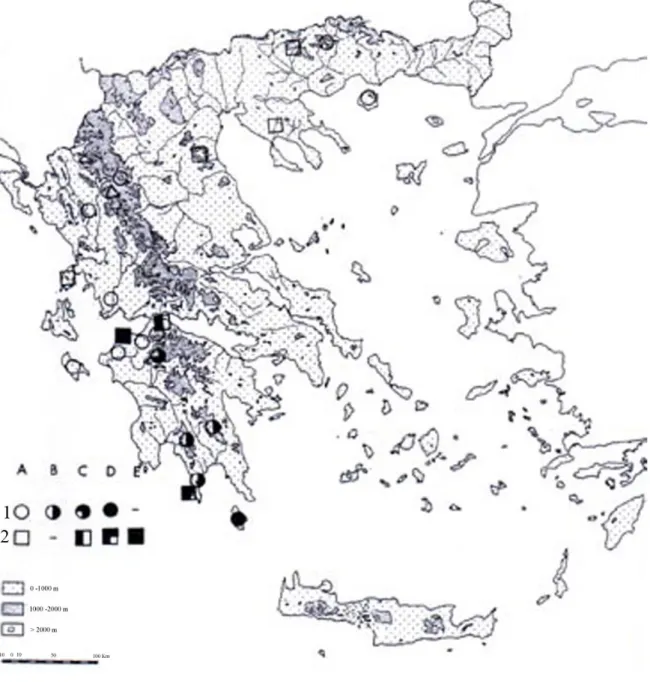
A. paniculatum L. resulted included in the third group of their cytologycal classification of Greek Allium. The group III includes seven Greek species with x=8 in which diploid and polyploid, even higher than 3x population, were found. The distribution pattern of cytotypes resulted clear in continental Greece, where most of the widespread species of the group III concentrate their polyploid cytotypes in the Southern part, mainly in the Peloponnese (see fig.14).
It’s to point out that Lefkas is not actually an island, being connected to the north-west Greek mainland by a narrow link. Consequently, the result obtained from the present study can be placed in the study cited, which observed for the north mainland’s population of the third group a diploid chromosome number. In particular also a population of Allium paniculatum L. from Lefkas has given the same result.
These observations are useful in order to estabilish plants migration routes; the distribution pattern of several species of the group III in Greece, included Allium
paniculatum L., suggests a north-south immigration, being the Peloponnese a “cul de
sac” of many species (Turril, 1929) in which migrations stopped and gave rise to polyploid forms. Anyway, the possibility of an east-west moviment cannot be excluded, as affirmed by Tzanoudakis and Vosa (1986); many of the species of this cytologycal group, e.g. A. paniculatum L., also occur in Asia Minor and their Peloponnesian populations or their ancestors could have arrived directly through the central Aegean migration routes.
investigated it results composed of metacentric chromosomes with length slightly decreasing in the idiogram, confirming the general uniformity in chromosome morphology as affirmed by Levan (1937) for all the taxa included in the paniculatum group.
Fig. 14. Distribution pattern of cytotypes in Allium guttatum (1) and Allium paniculatum (2). Note that the polyploid populations occurs only in south Greece. A: 2x, B: 2x, 3x, C: 3x, D:4x, E: 5x.
3.4
Allium guttatum Steven ssp. sardoum (Moris) Stearn
3.4.1 Generality
Allium guttatum ssp. sardoum (= A. margaritaceum Sm., non Moench, A. sardoum Moris, A. gadinatum Pérez Lara ex Willk., A. confusum Halácsy) is one of
the three subspecies of Allium guttatum Steven. The other two subspecies are ssp.
guttatum and ssp. dalmaticum, as reported in Flora Europaea (1980).
The species is distributed in Southern Europe and it grows mainly in dry places. In particular the ssp. sardoum grows from Portugal to Turkey and it’s one of the most common and abundant taxon in Greece.
Concerning the karyological aspect, the chromosome numbers 16, 32 and 48 are mentioned for Allium guttatum ssp. sardoum in Flora Europaea (1980), pointing out
32 for this subspecies which he mentions from Attica, Kiklades, Makedonia. Özhatay (1990, 2002) reported a diploid chromosome number for Turkey. Tzanoudakis (1985) studied many populations belonging to this taxon from Greece; most of them resulted diploid, and triploid 2n=3x=24 were also found as new chromosome record. It must be observed that triploid or mixed triploid and diploid populations were found only in the mountains of Peloponnese. All the population of central and northern Greece were exclusively diploid, with only one exception. In two populations a supernumerary (B) chromosome has been observed in the metaphase plates. In both cases the supernumerary chromosome was easily visible, even if small in size, with more or less equal arms. Also Cheshmedziev (1971) mentioned 0-3 B-chromosomes in A. guttatum.
With regard to the chromosome morphology, in all the Greek populations of
Allium guttatum ssp. sardoum studied by Tzanoudakis (1985) the haploid
chromosome complement x=8 included six metacentric or nearly metacentric chromosomes and two satellited chromosomes.
Following the nomenclature of Tzanoudakis (1983), that classifies four types of satellited chromosome pair (A, B, C, D) for the relative position of the centromere and of the nucleolar constriction, one of the satellited chromosome resulted of type B, that means metacentric and submetacentric chromosomes with the nucleolar constriction in a ± median region of the chromosome arm.
The other satellited chromosome pair usually resulted of type D, that means chromosomes of any type with a nucleolar constriction in one of the arms in a very proximal position to the centromere region; it is symbolized as (D). Variations were observed with regard to the position of the nucleolar constriction in the satellited chromosomes, especially in that of type D; so in some populations karyotypes with both of the satellited chromosome pair of type B were present.
It’s pointed out that the classification of nucleolar chromosomes of Tzanoudakis (1983) is based on the Ved Brat’s classification (1965), that indicated six types of satellited chromosomes in the genus Allium. Tzanoudakis (1983) proposed a new classification during his investigation on some Allium taxa of Greece, because of the great variation observed with regard to the position of the centromere and of the nucleolar constriction of the satellited chromosomes; consequently the appearance of satellited chromosomes with transitional characteristic suggested to unit some of the
Ved Brat’s types. For example, the A. scorodoprasum (III) and A. sativum (IV) types, have been united in the (B) type of Tzanoudakis; the centromere position is median-submedian in both the two cases but the nucleolar constriction is proximal to the centromere in the A. sativum type and median-submedian in the A. scorodoprasum type.
3.4.2 Results
10 µm
2n = 2x = 16: 8 m + 2 m (B) + 2 m + 2 m (B) + 2 m
Fig. 16 Allium guttatum Steven ssp. sardoum (Moris) Stearn, K18: microphotograph of a somatic metaphasic plate (up) and its corresponding idiogram with the idiogrammatic formula (down). Scale bar = 10 µm.
3.4.3 Discussion
The present investigation on a population of Allium guttatum ssp. sardoum from Zakynthos, Ionian Islands, gives a diploid chromosome number, 2n=16 (fig. 16). This result agrees with the several authors mentioned above, confirming the basic number x=8. It can support the hypothesis of Tzanoudakis (1985), that observed almost exclusively diploid populations of the taxon in object in central Greece and in the lowland of Peloponnese, being Zakynthos very close to Peloponnese. Furthermore, after wider studies on Greek Allium, Tzanoudakis and Vosa (1986) could describe a distribution pattern of cytotypes in Greece, as already explained in the previous paragraph for Allium paniculatum. This unit has been included in the third group likewise Allium guttatum.
In this context, the fact of a north-south gradient of rising polyploidy levels in Greek Allium can be observed also for the species in object: diploid in north Greece, north Peloponnese, and Zakinthos (from which comes also the specimen in object), mixed diploid and triploid toward the southern mountains of Peloponnese, tetraploid in Kithira, southernmost island of Peloponnese (see fig 14). Consequently, the present study can be placed as a support in this distribution pattern.
Concerning the karyotype morphology, the idiogrammatic formula obtained from the present study is:
2n = 2x = 16: 8 m + 2 m (B) + 2 m + 2 m (B) + 2 m
This karyotype complement is in agreement with Tzanoudakis (1985) for his investigation on the Greek populations, being formed of six metacentric chromosome pairs, and two satellited pairs. Instead, some light differences can be observed in respect to the Turkish populations studied by Özhatay (1990), that reported two pairs of metacentric and six pairs of submetacentric, four of which with secondary constriction close to the centromere. Anyway some of the metacentric pairs indicated in the present study are tendent to a submetacentric morphology, and it can explain the difference noted. Additionally, the estimation of the parameters in these particular type of satellited chromosomes can be done in various ways, taking or not into account the size of the satellite for the calculation of the r-index.
agreement with the other Greek populations investigated.
With regard to the satellites observed, their position on the fifth and the seventh chromosome pairs is verified in this study, if the size of the satellites is included in the calculations of the chromosome lenght. In this way the resulting positions of satellited pairs is placed in the little range of variability deduced from the other investigations made on Greek plants; Tzanoudakis (1985) indicated them in the fifth and eigthth position, and in sixth and eigthth position for material from Cyprus (1999).
The nucleolar constriction of both of the metacentric or submetacentric satellited chromosome pairs in the metaphasic plates here studied results to be in a ± median region of the chromosome arm; consequently, following the classification of Tzanoudakis (1983), both of the satellited pairs in object are of (B) type.
In respect to the Ved Brat’s classification (1965), they can considered of the A.
scorodoprasum (III) type. Anyway, further investigations are needed in order to
observe the continuous variability, pointed out by Tzanoudakis, among some of the satellited types.
At last, focusing on the metaphasic plates, a small supernumerary chromosome with equal arms can be observed, in agreement with some authors over mentioned.