5.1 Introduzione
In questo capitolo verranno illustrati i “materiali” utilizzati nell’esecuzione degli esperimenti e verranno descritte le varie analisi realizzate sulle reti neuronali in vitro.
5.2 Tipi cellulari utilizzati
In una prima fase sono stati utilizzati i neuroblastomi SH-SY5Y, derivanti da linee cellulari tumorali retiniche umane, ottenute dalla “Bank of biological material Interlab Cell Line Collection (ICLC)” di Genova.
La scelta di questo tipo di cellule è stata dettata da esigenze pratiche, trattandosi di cellule facilmente coltivabili e di facile reperibilità, e anche da esigenze economiche.
Sfruttando tali tipi cellulari è stato possibile imparare a “maneggiare” le cellule, mettere a punto la miglior metodologia che consentisse di acquisire immagini delle stesse regioni della piastra di coltura e, a partire dalle immagini ottenute, sviluppare un algoritmo efficace per lo studio della connettività.
In un secondo momento si è passati allo studio, più complesso, di neuroni embrionali, forniti dalla dottoressa Francesca Vaglini (Istituto di Farmacologia, Scuola di Medicina, Università di Pisa).
Per entrambi i tipi cellulari le immagini sono state ottenute attraverso una telecamera collegata al microscopio ottico (Olympus), con ingrandimento 10 X.
5.2.1 Neuroblastomi SH-SY5Y
Gli SH-SY5Y sono una linea cellulare di neuroblastoma, un tumore maligno embrionario specifico del bambino, che prende origine dal tessuto simpatico, da cui fisiologicamente prendono origine la midollare del surrene ed i gangli del sistema nervoso simpatico. Tali cellule rappresentano un ottimo modello simil-nervoso, sono capaci di proliferare in modo continuo e possono essere differenziate con una semplice procedura.
La differenziazione è un processo di graduale specializzazione, durante il quale le cellule acquistano o esaltano la capacità di compiere una funzione specifica. Le cellule differenziate conservano lo stesso genoma della cellula iniziale, solo che restringono le potenzialità del loro patrimonio genetico per svolgere la funzione specifica.
In molti casi le cellule tumorali hanno perso la loro specificità, ma questa può essere indotta in vitro con varie molecole e nel caso dei neuroblastomi SH-SY5Y, viene usato l’acido retinico, un componente attivo della vitamina A (figura 5.1).
L'acido retinoico sembra partecipare alla maturazione embrionale e numerosi studi hanno dimostrato la sua capacità di indurre la differenziazione di alcune linee cellulari, tra cui anche la SH-SY5Y, osservabile sia come produzione di neuriti, che come espressione della proteina marker neuronale GAP-43. [1, 2, 3, 4, 5]
I recettori per l’acido retinoico fanno parte di una famiglia di recettori nucleari che comprende anche quelli per la vitamina D. L’acido retinoico penetrato nella cellula viene portato al recettore tramite specifiche proteine cellulari di trasporto ed, una volta legatosi, determina la regolazione dell’espressione di alcuni geni.
5.2.2 Colture degli SH-SY5Y
I neuroblastomi sono stati seminati in una fiasca e si è usato come mezzo di coltura il DMEM. Il terreno completo ha la seguente composizione:
• 93 % di DMEM semplice; • 5 % di FBS;
• 1 % di antibiotici (penicillina / streptomicina); • 1 % di L-glutammina.
Sono stati sottoposti a trattamento con RA 10-5 M, in quantità pari a 1 μl per ml di terreno, per 7 giorni.
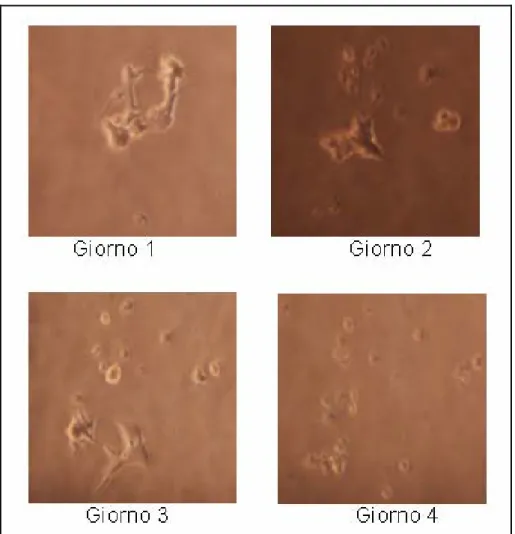
L’acido e il terreno di coltura sono stati rinnovati ogni tre giorni. Durante il processo di differenziazione sono state scattate delle foto che hanno dimostrato come le cellule a distanza di pochi giorni l’uno dall’altro si siano ben differenziate e abbiano formato dei lunghi prolungamenti (figura 5.2, 5.3 e 5.4).
Figura 5.2 Trattamento con RA: giorno 1
Figura 5.4 Trattamento con RA: giorno 5
Le cellule differenziate sono state tripsinizzate con le normali procedure e seminate in una nuova piastra.
Nei giorni successivi sono state scattate le foto, sempre nella stessa regione, grazie all’uso di apposite grigliette (figure 5.5 e 5.6).
Tali foto si sono dimostrate molto utili per testare la validità dell’algoritmo e scegliere passo passo gli operatori di elaborazione delle immagini più efficaci per i nostri fini.
Valutare l’andamento della connettività nel tempo è stato però impossibile, in quanto le cellule, nonostante la somministrazione dell’acido retinico, hanno continuato a replicarsi molto rapidamente, rendendo impossibile il riconoscimento di una stessa cellula da un giorno all’altro e quindi un’analisi a nodi costanti.
Figura 5.5 Cellule differenziate: giorno 1
5.2.3 Neuroni primari
Le cellule utilizzate in questa tesi sono neuroni embrionali di topo al tredicesimo giorno di gestazione provenienti dall’area del mesencefalo.
La preparazione delle colture neuronali da animali embrionali presenta numerosi vantaggi:
9 i neuroni sono meno suscettibili al danno durante la dissociazione quando i loro soma sono ancora piccoli e quando non hanno ancora formato grandi alberi assonici e dendritici e non sono altamente innervati;
9 i neuroni sono meno dipendenti dalle cellule target per il supporto trofico al primo stage del loro sviluppo;
9 tra i neuroni primari, quelli embrionali sono quelli che possono essere mantenuti in coltura in vitro più a lungo.
I neuroni sono stati prelevati dal topo in quanto il topo è il principale organismo modello per la sperimentazione nella genetica dei mammiferi e il surrogato dell’uomo studiato più intensivamente, grazie soprattutto alla consistenza genetica e al modesto costo. [6, 7]
Il topo è separato dagli esseri umani soltanto da circa 100 milioni di anni di evoluzione. Il suo genoma ha le stesse dimensioni del nostro ed esiste una corrispondenza quasi uno a uno tra fra geni umani e di topo. Le nostre proteine sono identiche all’80–90% nella loro sequenza amminoacidica e grossi blocchi di somiglianza di sequenza nucleotidica sono evidenti quando si confrontano le sequenze regolatrici di DNA.
Inoltre, grazie alla loro ingegnosità e perseveranza, i biologi dello sviluppo hanno trovato nuove strade per accedere all’embrione precoce di topo senza ucciderlo e per generare topi su misura con mutazioni in qualunque gene scelto. Le colture nervose dai mutanti neurologici forniscono la preziosa opportunità di analizzare gli effetti di specifici geni sullo sviluppo neurale e sulla funzione. [8, 9]
I neuroni derivano da due ceppi di topi diversi: SVI e CD1.
Figura 5.7 Topo CD1
5.2.4 Preparazione delle colture mesencefaliche
La preparazione delle colture mesencefaliche è una procedura molto delicata e relativamente complessa, che ha richiesto differenti passi. [10]
I cervelli prelevati dall’embrione sono stati messi in una soluzione bilanciata osmoticamente al pH fisiologico. Una soluzione di sale bilanciata (phosphate buffered saline: PBS) consiste in una semplice miscela di sali quali Na, K, Mg, Ca, Cl, PO4 e HCO3 alle
concentrazioni approssimativamente del fluido extracellulare, insieme con glucosio.
Il mesencefalo è stato sezionato sotto controllo microscopico e piazzato in un mezzo nutriente composto da F12 e dal terreno di coltura MEM, in rapporto1/1, supplementato con 2 mM di glutamina, 10% di Nu-serum e 3.33 mM di glucosio. Tutti i componenti del mezzo, a parte il Nu-serum sono stati comprati dalla Sigma.
Dopo la dissezione, il tessuto ottenuto è stato disperso in una sospensione di singole cellule. La dissociazione è stata realizzata sia con trattamento enzimatico che meccanico (triturazione e filtrazione). Il tessuto è stato meccanicamente disperso, usando delle pipette Pasteur, centrifugato a 100 g per tre minuti e risospeso nel mezzo di coltura.
Le cellule sono state contate con un emocitometro e piastrate, a diverse densità, in piastre multiwell da 6 pozzetti, ognuno dal diametro di 2,4 cm.
La superficie di ogni pozzetto era stata preventivamente ricoperta con 15 µg/ml di poli-D-lisina in soluzione acquosa per 1 ora a 37°C, e sciacquata tre volte con acqua sterile.
Le cellule sono state mantenute a 37°C in atmosfera contenente il 5% di CO2.
Per prevenire la crescita delle cellule gliali e ottenere delle colture neuronali virtualmente pure, dopo 72 ore dalla piastratura è stata aggiunta citosina arabinoside o Ara-C alla concentrazione di 10 µM.
L’Ara-C, usato generalmente come farmaco antitumorale, è un antimetabolita, ovvero una sostanza la cui struttura chimica è simile ad altri metaboliti normali presenti nell’organismo. Questa
analogia strutturale comporta o il blocco di un sistema enzimatico di cui il metabolita fisiologico funziona da substrato, o la sintesi di un prodotto inattivo o con diversa funzione. Si tratta più precisamente di un antipirimidinico: inibisce a vari livelli la sintesi delle pirimidine (citosina e timidina) costituenti del DNA.
5.2.5 Risposta dei neuroni primari in coltura
Si sono considerate cinque differenti densità di semina: • 10000 cellule/cm2;
• 50000 cellule/cm2; • 100000 cellule/cm2; • 250000 cellule/cm2; • 500000 cellule/cm2.
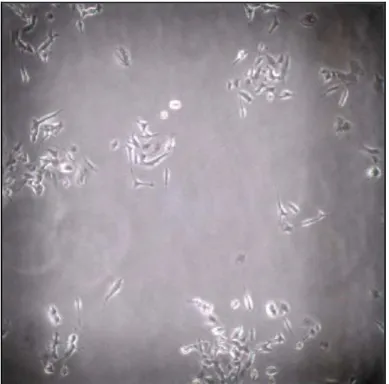
Si è visto però che, per densità di semina inferiori a 100000 cellule/cm2, i neuroni non sono in grado di sopravvivere per più di qualche giorno e di formare una rete neurale in vitro (figure 5.8 5.9, 5.10, 5.11, 5.12 e 5.13 ).
Figura 5.8 Neuroni di topo SVI
Figura 5.9 Neuroni di topo SVI
Figura 5.10 Neuroni di topo SVI
Figura 5.11 Neuroni di topo CD1
Figura 5.12 Neuroni di topo CD1
Figura 5.13 Neuroni di topo CD1
5.3 Preparazione delle griglie
Per poter fotografare sempre le stessa area della piastra, ovvero le stesse cellule a intervalli di tempo differenti, sono state preparate, usando il programma CorelDraw, delle apposite griglie micrometriche.
5.3.1 CorelDraw
CorelDraw è uno dei più rinomati programmi di grafica vettoriale, inserito in una suite di grafica professionale che comprende, oltre a CorelDraw, altri programmi come CorelPhotoPaint, per la grafica pittorica, CorelCapture, per la cattura delle immagini a video, Coreltrace , per il tracciamento delle immagini bitmap, e tanti altri. CorelDraw deve molto della sua popolarità alla sua estrema semplicità di utilizzo, alla totale personalizzazione dell'interfaccia ma soprattutto alla sua flessibilità ed intuitività che rende la realizzazione di qualsiasi progetto una vera passeggiata.
Consente di creare e modificare le immagini vettoriali.
La grafica vettoriale è la grafica orientata agli oggetti. Ogni oggetto è composto da singoli elementi, ognuno dei quali è un elemento a sé stante, indipendente da tutti gli altri e con le sue proprietà (colore di riempimento, colore di contorno, spessore del contorno ecc..). Ogni elemento trova le proprie posizione e forma grazie ad una serie di coordinate e impostazioni matematiche.
Un'immagine vettoriale si può spostare e modificare, si può ingrandire o rimpicciolire a piacimento, mantenendo inalterate chiarezza e definizione.
Si pensi ad un disegno rappresentante una casa: in questo caso la porta, la finestra, i vetri e tutto quello che concorre a formare l'immagine saranno oggetti divisi l'uno dall'altro, ognuno con le proprie caratteristiche ed ognuno di essi si potrà ridimensionare, ricolorare, cancellare senza intaccare gli altri elementi che compongono l'immagine.
Inoltre le immagini vettoriali richiedono poca memoria per essere eseguite.
5.3.2 Realizzazione delle griglie
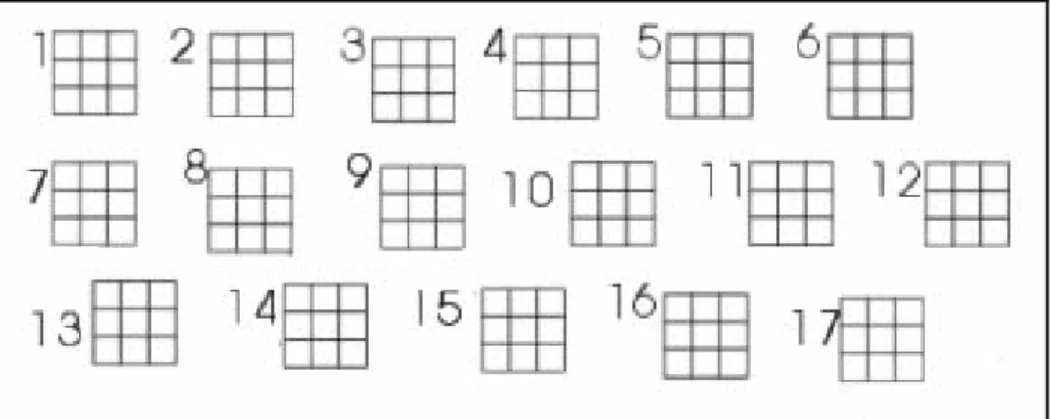
Prendendo ispirazione dalla camera di Burker, si è scelto di disegnare una serie di quadrati, ciascuno dei quali è stato suddiviso, attraverso due linee orizzontali e due verticali, in nove quadratini minori e quindi in nove aree.
Ciascun quadrato principale è inoltre stato numerato, così da poterlo identificare in maniera univoca(figura 5.14).
Tali griglie sono state stampate su lucido e poi incollate sotto la piastra di coltura, contenente le cellule aderite sulla superficie inferiore.
L’osservazione al microscopio delle piastre ha permesso di identificare le zone ottimali per l’acquisizione delle immagini.
Si sono scelte quelle regioni la cui area corrispondeva all’area racchiusa da uno dei quadratini minori di uno dei quadrati principali.
Ricordando la posizione del quadratino scelto all’interno del quadrato principale (alto a destra, alto a sinistra, ecc..) e il numero di questo ultimo, ogni volta è stato possibile ritrovare la stessa area e fotografarla.
5.4 Analisi delle reti neurali
Per comprendere come una rete neuronale funzionante possa costituirsi a partire da singole entità e studiarne l’evoluzione e la variazione della connettività, fino al raggiungimento di una configurazione stabile, sono state scattate, ad intervalli di 24 ore, delle foto della medesima area della piastra in cui le cellule sono state fatte crescere.
A partire dalle immagini, per ognuna di essa, è stato possibile realizzare differenti tipi di analisi.
Per poter far ciò, le reti neuronali sono state modellate matematicamente come dei grafi in cui i nodi sono il centro di un aggregato di neuroni e gli archi sono le sinapsi.
¾ tutti i nodi sono identici; ¾ tutti gli archi sono identici;
¾ la direzionalità degli archi è trascurata;
¾ il numero dei nodi è costante nel tempo e, tra di essi, variano solo le proprietà di raggiungibilità e connettività e la distanza euclidea..
5.4.1 Analisi della raggiungibilita’
Per studiare l’evoluzione della matrice di raggiungibilità nel tempo, è stato realizzato un algoritmo in MATLAB che, a partire da un’immagine della rete, consente di ottenere automaticamente la matrice di raggiungibilità e il corrispondente grafo.
L’algoritmo consiste di differenti fasi.
FASE 1: rilevamento dei nodi
L’immagine acquisita dalla telecamera viene letta dal programma e in seguito viene visualizzata sullo schermo.
I nodi vengono individuati ciccandoci sopra con il tasto sinistro del mouse e per ognuno vengono mostrati il numero che lo identifica e le coordinate spaziali. (figura 5.15).
1 347 133 2 456 166 3 192 235 4 368 298 5 528 338 6 236 322 7 299 357 8 247 413 9 344 529 10 149 349 11 430 434 12 519 230 13 600 176 14 711 144 15 670 298 16 616 429
Figura 5.15 Esempio di identificazione dei nodi
Viene inoltre costruita una matrice costituita da un numero di righe pari al numero dei nodi e da due colonne; la riga iesima di tale matrice riporta le coordinate (x, y) del nodo i.
FASE 2: ricerca degli oggetti di interesse (cellule)
Tale fase consiste nell’elaborare e modificare l’immagine sorgente, in modo da ottenere un’immagine binaria chiamata maschera, in cui sono evidenziati i neuroni.
La ricerca degli oggetti di interesse avviene in differenti passi, descritti di seguito e riassunti in figura 5.16.
Figura 5.16 Passi della segmentazione dell’immagine originale
¾ Passo 1: trasformazione dell’immagine RGB in un’immagine a livelli di grigio.
Tale conversione è fondamentale in quanto gli operatori di elaborazione utilizzati possono essere applicati solamente alle immagini a livelli di grigio (figura 5.17).
Figura 5.17 Immagine a livelli di grigio
¾ Passo 2: rilevamento dei bordi.
Per il rilevamento dei bordi si è scelto di utilizzare un operatore di bordo basato sul gradiente: l’operatore di Sobel. Tale scelta è legata essenzialmente alla semplicità e all’estrema versatilità di tale operatore e al fatto che, dopo diverse prove e confronti con altri operatori, esso è risultato essere il più efficace ai nostri scopi.
L’immagine del gradiente è sogliata in modo da selezionare i pixels caratterizzati da significativi cambiamenti di intensità, che vengono messi a bianco, mentre gli altri sono neri (figura 5.18).
Figura 5.18 Applicazione dell’operatore di Sobel e sogliatura
¾ Passo 3: dilatazione.
L’immagine binaria ottenuta è dilatata per delineare meglio i contorni e ripristinarne la continuità, eliminando eventuali gaps (figura 5.19).
Si sono utilizzati elementi strutturali verticali seguiti da elementi strutturali orizzontali.
Figura 5.19 Immagine dilatata
¾ Passo 4: riempimento interior gaps.
All’ interno degli oggetti segmentati ci sono ancora piccole cavità, che vengono riempite per mezzo dell’operazione morfologica di “Interior Fill” (figura 5.20).
Figura 5.20 Applicazione dell’operatore morfologico ”Interior Fill”
¾ Passo 5: erosione.
Gli oggetti segmentati vengono rifiniti erodendo l'immagine 2 volte (figura 5.21).
L’erosione ripristina le dimensioni originali degli oggetti, mantenendo uniti quelli molto vicini.
Figura 5.21 Applicazione dell’operatore morfologico erosione
A questo punto si ottiene un’immagine binaria, in cui le strutture di interesse, ovvero i neuroni con tutte le loro diramazioni, sono bianche, mentre lo sfondo è nero.
FASE 3: divisione in regioni
In base alla connettività dei vari pixels, l’immagine è divisa in regioni o oggetti: tutti i pixel 8-connessi tra loro appartengono ad una stessa regione.
A ciascun oggetto identificato viene inoltre assegnato un determinato colore (figura 5.22).
Figura 5.22 Divisione dell’immagine in regioni
FASE 4: costruzione della matrice di raggiungibilità
Due nodi sono raggiungibili se appartengono allo stesso oggetto, ovvero se i pixel che li rappresentano hanno gli stessi valori di R, G, B.
Per ogni nodo i viene realizzato un confronto tra i suoi valori di R, G, B e quelli di ogni altro generico nodo j e, se tali valori sono uguali, viene messo 1 nella posizione i,j della matrice di raggiungibilità, diversamente 0.
FASE 5: costruzione del grafo di raggiungibilità
Una volta realizzata la matrice di raggiungibilità viene disegnato il grafo di raggiungibilità, in cui esiste una freccia tra il nodo i e il nodo j se l’elemento i,j della matrice è pari a 1 (figura 5.23).
Node 1 Node 2 Node 3 Node 4 Node 5 Node 6 Node 7 Node 8 Node 9
Node 10 Node 11 Node 12 Node 13 Node 14 Node 15
Node 16
Figura 5.23 Grafo di raggiungibilità
5.4.2 Studio delle distanze
Dopo aver individuato i vari nodi, il programma realizzato in Matlab consente anche di calcolare la distanza euclidea tra di essi. Viene costruita una matrice quadrata di dimensione pari al numero dei nodi e l’elemento i,j di tale matrice rappresenta la distanza tra il nodo i e il nodo j.
E’ chiaro che la matrice delle distanze è una matrice simmetrica.
5.4.3 Costruzione del grafo e analisi della connettività
Dati i nodi, viene costruita la matrice delle adiacenze e quindi, attraverso opportuni comandi di Matlab, il grafo.
Dalla matrice delle adiacenze è inoltre ricavata la connettività o grado di ogni nodo. Come detto nel capitolo 3, il grado di un nodo è definito come il numero totale di connessioni con altri nodi e fornisce un’idea dell’importanza di un nodo.
La connettività del nodo i si ottiene sommando gli elementi della iesima colonna (o riga) della matrice delle adiacenze.
Viene costruito, sempre attraverso Matlab, un vettore riga di dimensione pari al numero dei nodi, il cui elemento i rappresenta la connettività del nodo i. Da tale vettore è possibile costruire l’istogramma: per ogni valore di connettività (asse delle ascisse) viene riportato il numero di nodi che hanno quel determinato valore (asse delle ordinate).
Infine viene calcolata anche la connettività media della rete, ottenuta come media aritmetica delle connettività di ogni nodo.
5.4.5 Test small-world
Per poter capire se una data rete neuronale in vitro appartiene alla classe degli small-worlds, sono stati calcolati il coefficiente di clustering e la lunghezza media di un cammino, così da poter effettuare il cosiddetto “test small-world”. [11]
Per poter classificare una rete come small-world essa deve soddisfare due condizioni:
1) il coefficiente di clustering della rete deve essere maggiore di quello di un grafo random con lo stesso numero di nodi e la stessa connettività media, ovvero la rete deve avere una elevata tendenza a formare gruppi locali molto ben connessi;
2) la lunghezza media di un cammino deve essere minore di quella di un grafo regolare con lo stesso numero di nodi e la stessa connettività media e maggiore di quella di un grafo random sempre con lo stesso numero di nodi e la stessa connettività media.
E’ stata inoltre graficata la distribuzione delle lunghezze del cammino: nell’asse delle ascisse ne sono riportati i possibili valori e
nell’asse delle ordinate il numero totale di coppie di nodi che hanno un determinato valore.
Bibliografia
1. V. Guarnirei, “Effetto sinergico dell’acido retinico e del deidroepiandrosterone sul differenziamento neuronale”- Dottorato di ricerca in Biochimica e Biotecnologia cellulare
2. Peter B. Simpson, Jamil I. Bacha, Emma L. Palfreyman, Anthony J. Woollacott, Ruth M. McKernan, Julie Kerby, - Retinoic Acid-Evoked Differentiation of Neuroblastoma Cells Predominates over Growth Factor Stimulation: An Automated Image Capture and Quantitation Approach to Neuritogenesis, Anal Biochem 2001 Nov 15; 298(2), pp. 163-169
3. D. Chambèry, B. de Gallé, S. Balbajko, - Retinoic acid stimulates IGF binding protein (IGFBP)-6 and depress 2 and IGFBP-4 in SK-N-SH human neuroblastoma cells, Journal of Endocrinology (1998) 159, 227-232
4. Elizabeth M. V. Jones-Villeneuve, Michael W. McBurney, Kem A. Rogers, Vitauts I. Kalnins, - Retinoic Acid Induces Embryonal Carcinoma Cells to Differentiate into Neurons and Glial Cells, The Journal of Cell Biology, volume 94, August 1982, 253-262
5. Tesi di laurea in Chimica e Tecnologia Farmaceutiche di Delia Venanzoni “Organizzazione e differenziazione di neuroblastomi SH-SY5Y su substrati polimerici microstrutturati: potenziali
applicazioni nel campo dell’ingegneria dei tessuti nervosa retinici”, 2002
6. Bruce Alberts, Alexander Johson, Julian Lewis, Martin Raff, Keith Roberts, Peter Walter, “Biologia molecolare della cellula”, Zanichelli, quarta edizione
7. Gary Bunker and Kimberly Goslin, editors, “Culturing Nerve Cells”, second edition, 1998
8. A. Messer and D. M. Smith, - In vitro behaviour of granule cells from Staggerer and Weaver mutants of mice, Brain Res. 130 (1977), p.p. 13-23
9. P. Kofugi, M. Hofer, K. J. Millen, et al., - Functional analysis of the weaver mutant GIRK2 K+ channel and rescue of weaver granule cells, Neurons 16 (1996), p.p. 941-952
10. Francesca Vaglini, Carla Perdini, Roberto Maggio, Giovanni U. Corsini, - Role of excitatory amino-acids in diethyldithiocarbamate-induced cell death in mesencephalic cultures Brain Research 674 (1995), 127-132
11. Orit Shefi, Ido Golding, Ronen Segev, Eshel Ben-Jacob, Amir Ayali, “Morphological characterization of in vitro neuronal networks”, Physical Review E 66, 021905 (2002)