SEZIONE I
1. TTV
1.1 LA SCOPERTA
I molti tentativi, svolti in questi ultimi decenni, al fine di identificare gli agenti responsabili delle epatopatie di origine sconosciuta, hanno condotto nel 1997 all’identificazione di un nuovo agente virale, grazie anche allo sviluppo di nuove tecniche di indagine diagnostica e di biologia molecolare.
Il nuovo agente virale, scoperto da Nishizawa e collaboratori (Nishizawa et al., 1997) nel siero di un paziente con epatite post-trasfusionale non-A non-G, fu chiamato TT (TTV) dalle iniziali di tale soggetto. La tecnica che ha reso possibile l’identificazione del nuovo virus è stata l’analisi di rappresentazione differenziale (RDA): questa tecnica, sviluppata da Lisitsyn nel 1993 (Lisitsyn et Wigler, 1993), consente di evidenziare sequenze di DNA selettivamente espresse nei tessuti patologici quando confrontate con sequenze derivanti dai tessuti sani degli stessi soggetti. Nel caso specifico di TTV il materiale genetico fu estratto dal siero al momento in cui il paziente presentava un picco nei livelli di transaminasi (ALT), e successivamente confrontato con il DNA dello stesso individuo al momento che i livelli di ALT risultavano normali. L’analisi delle sequenze esclusive, individuate nel paziente al momento del picco, con quelle presenti in banca dati, ha portato all’isolamento di un
clone di circa 500 nucleotidi, denominato N22, che non aveva omologia con alcuna delle sequenze fino ad allora note e per questo fu considerato il prototipo di un nuovo virus a DNA. Studi condotti da Okamoto e collaboratori (Okamoto et al., 1998b) hanno portato alla caratterizzazione, tramite metodiche di clonaggio e sequenziamento, di un genoma virale, apparentemente completo, di 3.739 nucleotidi (nt), denominato TA278. Le piccole dimensioni, alcune peculiarità della sequenza nucleotidica e l’assenza di envelope fecero supporre l’appartenenza del virus alla famiglia Parvoviridae. Nel 1999, però, Miyata e collaboratori, evidenziando nell’isolato TA278 l’assenza delle strutture palindromiche terminali tipiche dei parvovirus, identificarono una nuova regione di 113 nucleotidi ricca in guanina e citosina (GC) e, mediante PCR inversa, riuscirono a dimostrare la natura circolare del genoma del virus. Questa scoperta, insieme ad alcune proprietà strutturali e molecolari che il virus mostrava condividere con i membri della famiglia Circoviridae, in particolare con il virus dell’anemia del pollo (CAV), alimentarono la convinzione che TTV rappresentasse il primo circovirus dell’uomo (Miyata et al., 1999).
In seguito, sono state isolate numerose specie virali con caratteristiche strutturali ed elementi genomici simili a TTV, ma con variabilità genetica elevata: tra questi gli isolati PMV (Hallet et al., 2000), SANBAN (Hijikata et al., 1999) e YONBAN (Yakahashi et al.,
2000a) aventi meno del 50% di omologia con il prototipo TA278. Questo quadro di specie virali altamente eterogenee dal punto di vista genetico divenne ancora più complesso con la scoperta di un altro gruppo di virus con genoma a singolo filamento di DNA circolare con proprietà intermedie tra TTV e CAV (Takahashi et al., 2000b), denominato inizialmente TLMV (TTV-Like-Mini-Virus), a sottolineare le dimensioni più piccole del genoma che risultava 2,9 Kb, contro le 3,8 Kb tipiche di TTV.
Dalla scoperta del virus ad oggi, sono stati effettuati molti studi per definire le sue proprietà molecolari e per determinare il suo reale ruolo patogeno. L’iniziale isolamento di TTV nel siero di un paziente con epatite criptogenetica ha fatto supporre che questo virus avesse un ruolo nel causare malattie del fegato, ma successivi studi volti a verificare questa ipotesi hanno spesso fornito risultati discordanti. Molti altri studi hanno preso in esame la possibilità che TTV potesse essere coinvolto in patologie extraepatiche, tuttavia in nessun caso è stato possibile arrivare ad una conclusione certa.
L’elevata prevalenza di TTV nella popolazione generale ha perfino suggerito che il virus potesse rappresentare un semplice commensale apotogeno dell’uomo (Griffiths, 1999; Mushahwar, 2000). Tuttavia, nell’attesa di una possibile dimostrazione del suo potenziale patogeno TTV può essere annoverato, insieme ad altri
agenti di recente scoperta nella categoria dei virus “orfani” attualmente non associati ad alcuna malattia.
1.2 TASSONOMIA E NOMENCLATURA
La classificazione di TTV ha subito nel corso degli anni numerose modifiche, ed ancora oggi l’inquadramento tassonomico del virus rimane incerto.
Per la ridotta dimensione della particella virale, l’assenza di
envelope, e la presenza di un DNA lineare a singolo filamento, TTV è
stato inizialmente associato alla famiglia Parvoviridae (Okamoto et al., 1998b). La successiva scoperta della regione ricca in GC, che dimostrava la circolarità del genoma (Miyata et al., 1999) ed altre caratteristiche strutturali e molecolari del virus, hanno condotto a sostenere la possibile appartenenza del virus alla famiglia
Circoviridae. I membri di questa famiglia sono infatti caratterizzati
dalla presenza di un genoma circolare a singolo filamento di DNA, mostrano un capside icosaedrico probabilmente costituito da un’unica proteina strutturale e sono privi di envelope (Todd et al., 1990). I circovirus animali classificati all’interno di questa famiglia sono tre: il virus della malattia del becco e delle penne dei pappagalli (BFDV), il circovirus porcino (PCV) ed il virus dell’anemia del pollo (CAV). In realtà tali agenti presentano tra di loro e con TTV notevoli differenze, sopratutto a livello della sequenza nucleotidica e quello
che sembra avvicinarsi maggiormente a TTV è CAV. Il loro genoma è infatti paragonabile sia per le dimensioni, 3.200 nucleotidi circa per CAV e 3.800 nucleotidi circa per TTV, sia per quanto riguarda la suddivisione del genoma in quanto possiedono una regione codificante con almeno 3 cornici di lettura aperte (Open Reading Frame, ORF) parzialmente sovrapposte (Iwata et al., 1998) ed una regione non tradotta UTR (Untranslated Region) ricca in GC con notevole omologia tra i due virus (Miyata et al., 1999). Nonostante queste omologie, numerose caratteristiche di TTV rimangono al momento uniche, tanto che Mushahwar e collaboratori suggerirono di considerare il virus come unico rappresentante di una nuova famiglia indicata con il nome di Circinoviridae (Mashahwar et al., 1999). In seguito alla scoperta del TT-like mini virus (TLMV), virus con caratteristiche intermedie tra TTV e CAV, è stato proposto di classificare i tre virus in una nuova famiglia detta Paracircoviridae (Takahashi et al., 2000b). Nell’attesa di una classificazione definitiva dal Comitato Internazionale per la Tassonomia dei Virus (CITV) è stato istituito un gruppo di studio che ha proposto di inserire TTV e TLMV in un nuovo genere detto Anellovirus (dal latino “anello”). Anche gli acronimi di TTV e TLMV sono stati modificati: Torqueteno virus (dal latino “catena sottile”) per TTV e Torqueteno mini virus (TTMV) per TLMV (Hino, 2002; Todd et al., 2002).
Tuttavia, il numero di virus simili a TTV sembra destinato a crescere. Recentemente, infatti, Jones e collaboratori hanno identificato, nel plasma di due pazienti con sindromi d’infezione acuta ad eziologia sconosciuta, due nuove sequenze virali. Dal confronto in banca dati e dalle analisi filogenetiche è emersa una somiglianza con i virus del gruppo TT seppur con divergenze genetiche elevate. Le nuove sequenze virali sono risultate avere dimensioni rispettivamente di 2.249 nt e di 2.635 nt, collocandosi perciò tra TTV e TTMV. I due nuovi virus sono stati, quindi, classificati nel genere Anellovirus in quanto possiedono un genoma circolare a singolo filamento con polarità negativa e presentano, come TTV e TTMV, una regione genomica codificante, suddivisa in più ORF parzialmente sovrapposte, e una regione non tradotta UTR. I nomi assegnati a tali virus sono Small Anellovirus 1 (SAV1) e Small Anellovirus 2 (SAV2). I due SAV mostrano tra loro un’omologia del 54% e con TTV e TTMV un’omologia rispettivamente del 32% e 35%. A causa delle loro dimensioni, della loro organizzazione genomica e della distanza filogenetica dagli altri anellovirus, è stato proposto che SAV1 e SAV2 rappresentino un terzo distinto gruppo di Anellovirus (Jones et al., 2005)..
Dal momento della scoperta del prototipo TA278 per la classificazione di TTV e dei virus ad esso correlati sono state avanzate diverse proposte, influenzate spesso dalla regione
genomica utilizzata per le analisi filogenetiche. Infatti i primi studi sono stati condotti scegliendo come bersaglio per l’amplificazione la regione N22 dell’ORF1: tali analisi hanno permesso l’identificazione di 23 genotipi che sono stati inizialmente riuniti in quattro principali raggruppamenti filogenetici (Okamoto et al., 1999; Biagini et al., 1999).
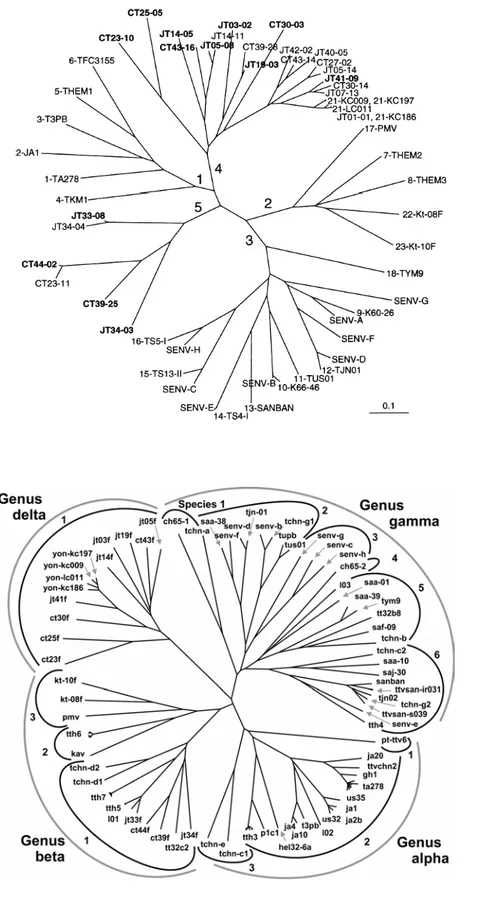
Successive analisi di sequenza condotte sulla regione non tradotta (Untranslated Region UTR) e sull’intero genoma hanno portato all’identificazione di più di 40 genotipi e 70 sottotipi. Nel 2002 Peng e collaboratori proposero una nuova classificazione per tutti gli isolati fino a quel momento identificati (72): essi venivano suddivisi in 5 principali genogruppi, identificati con i numeri da 1 a 5, sulla base di divergenze nucleotidiche almeno del 50% (Peng et al., 2002; figura I.2A). In base a questa classificazione il genogruppo 1 è rappresentato dal prototipo TTV (isolato TA278) di genotipo 1 ed include altri cinque genotipi aggiuntivi, dal 2 al 6 (Okamoto et al., 1999).
Il genogruppo 2 comprende gli isolati TTV di genotipo 7, 8, 22, 23 ed anche il 17, che è rappresentato dall’isolato PMV (Hallett et al., 2000). Il genogruppo 3, la cui classificazione risulta molto complessa per la numerosità degli isolati che comprende, è composto dai genotipi dal 9 al 16 (l’isolato prototipo è il SANBAN), dai genotipi 18,19 (SENV-F) e 20 (SENV-G) e da altri sette genotipi di SENV, da
A ad H. Gli ultimi due genogruppi, il 4 e il 5, comprendono rispettivamente, nove genotipi (l’isolato prototipo è YONBAN) e quattro nuovi isolati il cui prototipo è JT33 (Peng et al., 2002).
Una variabilità simile, se non superiore a quella di TTV, è stata dimostrata anche per TTMV che è attualmente distinto in 4 genogruppi, separati da distanze genetiche superiori al 40% (Biagini et al., 2001b; Gallian et al., 2002).
In un recente studio condotto da Jelcic e collaboratori, effettuato sulla base di un’analisi filogenetica nel quale sono state confrontate le sequenze di 24 nuovi isolati con quelle di 79 isolati già presenti in banca dati, è stato proposto di inserire i ceppi di TTV in una nuova famiglia denominata Anelloviridae (Jelcic et al., 2004). Il CITV attualmente non si è ancora pronunciato su tale proposta.
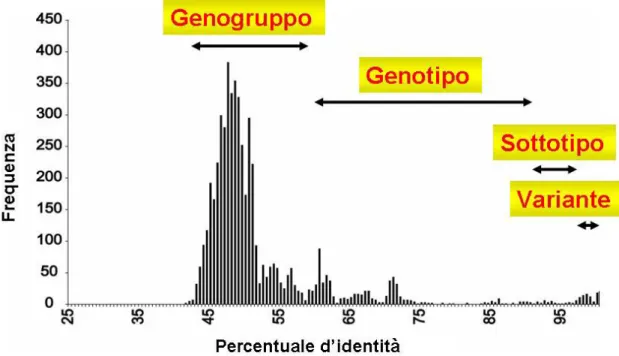
Secondo questa nuova classificazione, il genogruppi 1 verrebbe trasformato nel genere α e i genogruppi 3 e 4 rispettivamente nei generi γ e δ, mentre i genogruppi 2 e 5 sarebbero raggruppati nel genere β. In base a questo nuovo criterio di classificazione, gruppi di TTV che presentano un’identità minore del 50% costituiscono generi separati, all’interno di questi gli isolati hanno tra loro un’omologia di sequenza compresa tra il 50% e 60%. La specie viene definita come un gruppo di isolati di TTV che mostrano tra loro un’identità di sequenza superiore al 60%. Si parla invece di tipi e sottotipi virali se l’identità di sequenza supera rispettivamente il 90% e il 96%; infine
gli isolati che presentano tra loro omologie del 96% vengono definiti come varianti di uno stesso sottotipo (figura I.1B; figura I.2).
B
B
Figura I.1. Relazioni filogenetiche tra i vari isolati di TTV. In (A) i
gruppi definiti da Peng e collaboratori (2002); in (B) i generi definiti da Jelcic e collaboratori (2004).
Figura I.2: Classificazione genetica dei vari isolati di TTV in base alla
proposta di Jelcic. Sull’asse delle ascisse è riportata la distribuzione di frequenza delle percentuali d’identità ottenute dal confronto delle sequenze nucleotidiche di 103 isolati di TTV (3200 nt) (Jelcic et al., 2004).
1.3 LA PARTICELLA VIRALE
Le informazioni circa le proprietà chimicho-fisiche e le caratteristiche morfologiche della particella virale sono generalmente riferite al genotipo 1 di TTV, che è stato il primo ad essere individuato e di conseguenza ad essere studiato più approfonditamente. Tuttavia le attuali conoscenze sul virus, porterebbero ad escludere differenze morfologiche significative tra i vari genotipi.
Purtroppo non si dispone ancora di un sistema di coltura cellulare in
vitro capace di supportare la replicazione di TTV, per cui le
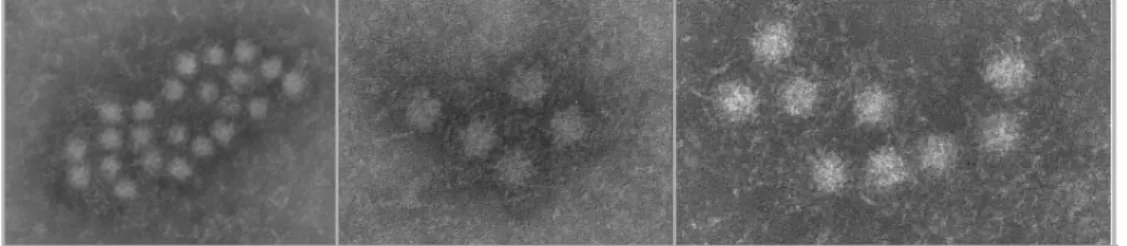
informazioni sulle caratteristiche strutturali del virus derivano, per lo più, da studi condotti con particelle virali purificate da plasma o da feci. Itoh e collaboratori hanno visualizzato la particella virale al microscopio elettronico, dopo incubazione di sovranatante fecale, contenente TTV di genotipo 1, con le γ-globuline purificate da plasma umano contenente anticorpi specifici per il medesimo genotipo (figura I.3, Itoh et al., 2000). TTV è apparso di forma sferica con capside a probabile simmetria icosaedrica e privo di pericapside. Il diametro del virus è risultato compreso tra 30-32 nm, come già era stato precedentemente determinato mediante l’uso di filtri di policarbonato a porosità decrescente (Mushahwar et al., 1999). La resistenza ai detergenti come il Tween 80 (Okamoto et al., 1998a), ai solventi e al calore secco (62°C per 96 h) hanno confermato l’assenza di un involucro lipidico esterno, assenza ulteriormente
avvalorata dal ritrovamento di TTV nella bile di soggetti infetti (Ukita et al., 1999) dove, per la presenza di acidi biliari, si possono rilevare solo virus privi di envelope. TTV, quindi, appare molto stabile e resistente come i membri della famiglia Circoviridae (Simmonds et al., 1998; Takayama et al., 1999a; Yokozaki et al., 1999b). Alcuni lavori riportano l’esistenza nel sangue di particelle virali complessate ad immunoglobuline (Itoh et al., 2000); per il resto non si conoscono altre forme circolanti del virus.
Le proteine di TTV non sono state ancora ben caratterizzate, tuttavia, per analogia con CAV, si ritiene che il capside icosaedrico sia costituito da molte unità di un’unica proteina strutturale; inoltre non sono stati identificati, al momento, né carboidrati né lipidi virali.
Figura I.3. Visualizzazione al microscopio elettronico di particelle di
TTV, in forma di aggregati in seguito ad incubazione di sovranatante fecali con γ–globuline umane specifiche per il genotipo 1a (Itoh et al., 2000).
1.4 ORGANIZZAZIONE DEL GENOMA E
PROTEINE CODIFICATE
La caratterizzazione del genoma di TTV è stata eseguita sottoponendo l’acido nucleico, estratto da plasma, all’azione di diversi enzimi: la sensibilità alla Dnasi I ed alla nucleasi Mung-bean e la resistenza alla Rnasi A ed all’enzima di restrizione NdeI ne hanno dimostrato la natura a singolo filamento di DNA (Okamoto et al., 1998b; Mushahwar et al., 1999). Attraverso esperimenti di ibridazione con molecole di RNA a polarità positiva e successiva digestione è stato possibile dimostrare che il DNA virale presenta una polarità negativa (Mushahwar et al., 1999). Ulteriori informazioni sull’organizzazione genomica derivano dall’analisi delle sequenze. L’amplificazione della molecola in tutta la sua lunghezza, circa 3,8 Kb, è stata ottenuta mediante clonaggio e PCR inversa condotta a partire dalla regione N22 (Mushahwar et al., 1999).
Il sequenziamento del 90% del genoma aveva fatto inizialmente ipotizzare che il DNA del virus fosse lineare (Okamoto et al., 1998b), supposizione smentita dalla successiva identificazione di una regione di circa 100 nucleotidi ricca in GC (Miyata et al., 1999): il genoma di TTV è quindi costituito da un singolo filamento di DNA circolare chiuso.
L’analisi filogenetiche di tutte le sequenze complete (o quasi) di TTV e TTMV ha rilevato una notevole somiglianza tra i due gruppi di virus
nell’organizzazione genomica anche se l’ampiezza della loro sequenza nucleotidica differisce di circa 1 Kb. Le differenze nelle dimensioni del genoma fra i virus del gruppo TTV sono risultate invece più modeste (Kamahora et al., 2000; Okamoto et al., 2000; Muljono et al., 2001). Le informazioni riguardanti la struttura del genoma di TTV si basano essenzialmente sull’analisi delle sequenze dei diversi isolati e sul relativo confronto con i virus meglio caratterizzati della famiglia Circoviridae.
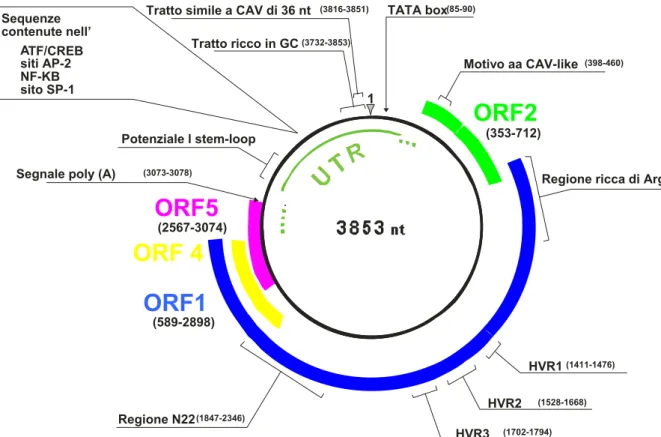
Il genoma è costituito per due terzi (2,6 Kb) da una regione codificante ORF e per un terzo da una regione UTR non tradotta ma ricca di elementi regolatori (Kamada et al., 2004; Suzuki et al., 2004; figura I.4). Sul filamento complementare al genoma, la regione espressa comprende da due a sei ORF parzialmente sovrapposte, potenzialmente codificanti con cornici di lettura differenti. Il gene ORF1 di TTV, localizzato tra i nucleotidi 589 e 2.898 sul ceppo TA278, codifica per un prodotto proteico di circa 770 aminoacidi, (675 in TTMV), ritenuto l'equivalente della proteina capsidica di CAV denominata VP-1. La proteina tradotta dall'ORF1 di TTV oltre a svolgere probabili funzioni strutturali, potrebbe essere coinvolta nel meccanismo replicativo: in queste proteine è stato identificato infatti un tratto di circa 100 aminoacidi molto ricco in arginina il quale, per la sua natura idrofilica, potrebbe avere un ruolo nel mediare l’interazione con il DNA (Shoya et al., 1998; Vihinene-Ranta et al.,
1997); tale dominio si ritrova anche nella regione aminoterminale di VP1 di CAV (Okamoto et al., 1998b).
Inoltre, all’estremità C-terminale del gene ORF1, sono stati individuati due motivi aminoacidici tipici delle proteine Rep replicasi utilizzate da virus a DNA che utilizzano il meccanismo a cerchio rotante. Queste proteine sono caratterizzate dalla presenza di almeno quattro motivi aminoacidici che risultano essere molto conservati tra i circovirus animali e vegetali. Inoltre, nella porzione centrale dell’ORF1, TTV presenta tre regioni ipervariabili dette HVR-1, HVR-2 e HVR-3 (Hight variable region) e potenziali siti di glicosilazione che, variabili in numero e in localizzazione nei differenti isolati, potrebbero modificare le proprietà biologiche ed antigeniche della proteina (Hijikata et al., 1999). Il gene ORF2, compreso tra i nucleotidi 353 e 712 del ceppo TA278, sembra codificare per una proteina strutturale di circa 120 aminoacidi in TTV (100 in TTMV), probabilmente implicata nella replicazione virale. A tal proposito è stata dimostrata un’omologia di sequenza fra questa proteina, in particolare quella codificata dall’isolato SANBAN, e la proteina VP-2 di CAV che ha una doppia funzione di fosfatasi, tirosin- e serin/treonina fosfatasi ed è implicata nei meccanismi di trasmissione intracellulare dei segnali durante la replicazione del virus (Peters et al., 2002). Dall’analisi della sequenza di molti isolati di TTV, il gene ORF-2 risulta spesso costituito da due piccole ORF, denominate a e
b, la prima delle quali appare molto conservata fra i diversi genotipi e le cui capacità codificanti non sono ancora note ( Tanaka et al., 2000b; Kakkola et al., 2002).
TATA box (85-90)
Tratto simile a CAV di 36 nt (3816-3851)
Sequenze contenute nel
A
l’
TF/CRE Tratto ricco in GC (3732-3853)
Figura I.4: Organizzazione genomica di TTV. Le posizioni si
riferiscono all’isolato TA278.
1
Regione ricca di Arg Motivo aa CAV-like (398-460) NF-KB B siti AP-2 sito SP-1
ORF2
(353-712) (3073-3078)Segnale poly (A)
Potenziale l stem-loop
ORF5
(2567-3074)ORF 4
ORF1
(589-2898) HVR1 (1411-1476) HVR2 (1528-1668) (1702-1794) HVR3 Regione N22 (1847-2346)Esperimenti in cui sono state transfettate cellule COS1 con un vettore contenente un costrutto del genoma linearizzato di TTV (genotipo 1), hanno dimostrato la presenza di altre ORF e condotto all’identificazione di tre distinti mRNA di 3,0, 1,2 e 1,0 Kb (Kamahora et al., 2000); i tre mRNA, che presentano polarità antigenomica ed hanno il sito di poliadenilazione al 3’ e il sito cap al 5’, sono stati ritrovati anche in cellule del midollo osseo infettate da TTV (Okamoto et al., 2000d; 2001). Il meccanismo di splicing ipotizzato per la produzione degli mRNA mediante l’utilizzo di diversi schemi di lettura, agirebbe sulle regioni ORF conosciute ma anche su ORF minori di nuova identificazione, dette ORF4 e ORF5, localizzate vicino alla porzione C-terminale dell’ORF1 e ad essa sovrapposte. Dall’analisi dei potenziali splicing, è emerso che il messaggero più lungo conterebbe per intero il gene ORF1, mentre i due mRNA più corti risulterebbero da porzioni dell’ORF2 con l’ORF4 e l’ORF5 (figura I.5).I prodotti proteici derivanti dall’ORF3 (o ORF2-4) e dalla ORF2-5 potrebbero essere implicati nella replicazione del virus o nei meccanismi di regolazione nella trascrizione delle proteine cellulari. In particolare l’ORF-3 sembra codificare per due proteine (p38 e p41) dotate di un differente grado di fosforilazione e per questo simili alla proteina NS5A del virus dell’epatite C (HCV; Asabe et al., 2001). Inoltre, in uno studio condotto da Kooistra e collaboratori, basato sulla considerazione che alcune sequenze nucleotidiche di TTV
mostravano similitudini con il gene dell’apoptina di CAV, è stato dimostrato che l’ORF3 codifica anche per una proteina di 105 aa in grado di provocare l’apoptosi di linee cellulari derivate da carcinoma epatocellulare umano (Kooistra et al., 2004).
Altre ORF possono essere dedotte dalla sequenza nucleotidica di alcuni isolati di TTV, ma il loro significato funzionale rimane al momento sconosciuto (Yokayama et al., 2002).
Studi recenti di trasfezione, in cui sono stati inseriti costrutti del genotipo 6 di TTV in cellule di rene umano immortalizzate (cellule 293), hanno evidenziato, mediante Western blotting, l’espressione di almeno 6 proteine, generate da 3 distinti mRNA con differenti strategie di traduzione a partire da 2 diversi codoni d’inizio, O1AUG (nt 581) e O2AUG (nt 354). Questi ultimi, ottenuti con splicing alternativi, mostravano dimensioni pressoché identiche (2,8, 1,2, e 1,0 kb) agli mRNA precedentemente identificati in vitro nelle cellule COS1 e nel midollo osseo. E’ stato inoltre possibile stabilire, utilizzando adeguati sistemi di marcatura, la localizzazione sub-cellulare delle 6 proteine: sono stati osservati tre patterns di espressione secondo cui due proteine derivanti dall’ORF1 ed ORF2 risultavano avere una localizzazione principalmente citoplasmatica, due proteine (ORF2/2 ed ORF 2/3) erano localizzate esclusivamente nel nucleo, mentre altre due (ORF1/1 ed ORF1/2) erano distribuite tra citoplasma e nucleo. Le proteine ORF1 ed ORF2, in base a
questo studio, sarebbero tradotte dall’mRNA di 2,8 kb a partire, rispettivamente, dalle triplette d’inizio O1AUG e O2AUG; le proteine ORF2/2 ed ORF2/3 deriverebbero invece dallo splicing degli mRNA rispettivamente di 1,2 e 1,0 kb partendo da O2AUG; anche ORF1/1 ed ORF1/2 risultano derivare dagli stessi RNA messaggeri di 1,2 e 1,0 kb ma utilizzando come codone d’inizio traduzione O1AUG (Qiu et al., 2005).
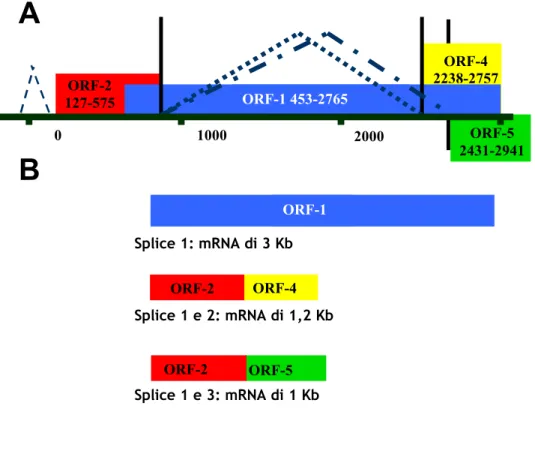
ORF-1 Splice 1: mRNA di 3 Kb ORF-2 ORF-4 ORF-5 ORF-2 Splice 1 e 2: mRNA di 1,2 Kb Splice 1 e 3: mRNA di 1 Kb 0 1000 2000 ORF-2 127-575 ORF-1 453-2765 ORF-5 2431-2941 ORF-4 2238-2757
B
A
Figura I.5: In A lo schema del trascritto primario con indicate le
regioni codificanti e i tre possibili splicing. In B lo schema dei tre trascritti secondari ottenuti dopo lo splicing.
La regione non codificante UTR, lunga circa 1,2 Kb, comprende un tratto di circa un centinaio di nucleotidi, con il 90% di basi nucleotidiche costituito da guanina e citosina. Al suo interno è inoltre presente una sequenza di 36 nucleotidi che possiede un alto grado di omologia con la stessa regione di CAV (Miyata et al., 1999). Alcuni tratti della UTR risultano estremamente conservati tra tutti i ceppi di TTV tanto da essere utilizzati come bersaglio per lo sviluppo di metodiche di amplificazione in grado di individuare i diversi genotipi del virus; tali regioni così conservate svolgono probabilmente funzioni di estrema importanza per la vitalità del virus. A questo proposito è stato dimostrato che in tutta la regione UTR, e in particolar modo nel tratto ricco in GC, si possono formare complesse strutture secondarie caratterizzate da tipici motivi ad ansa notoriamente associati a funzioni regolatorie (Hijikata et al., 1999; Miyata et al., 1999; Mushahwar et al., 1999). Mediante saggi di espressione in vitro (Noteborn et al., 1994), nella regione UTR di CAV sono state evidenziate, in aggiunta al sito di poliadenilazione a valle di VP-1 e al sito di trascrizione TATA BOX a monte di VP-2, alcune sequenze ripetute invertite che costituiscono elementi di attivazione della trascrizione, ed altre regioni di legame per specifici fattori di trascrizione in grado di stimolare la proliferazione cellulare (SP-1, NF-Kb, AP-2). Tali motivi sono stati ritrovati anche nell’UTR di TTV, compresa una regione palindromica, situata a valle del sito di
poliadenilazione, altamente conservata fra i diversi isolati del virus (fino al 90%).
Esperimenti di mutagenesi hanno evidenziato la presenza di alcuni elementi regolatori negativi e positivi (figura I.6) la cui attività, probabilmente, dipende dal tipo di cellula infettante (Suzuki et al., 2004). Studi recenti hanno inoltre identificato la localizzazione del sito di inizio trascrizione dei 3 RNA messaggeri osservati, che risulta collocarsi 113 nt a valle rispetto al promotore basale, costituito dalla TATA BOX (Kamada et al., 2004).
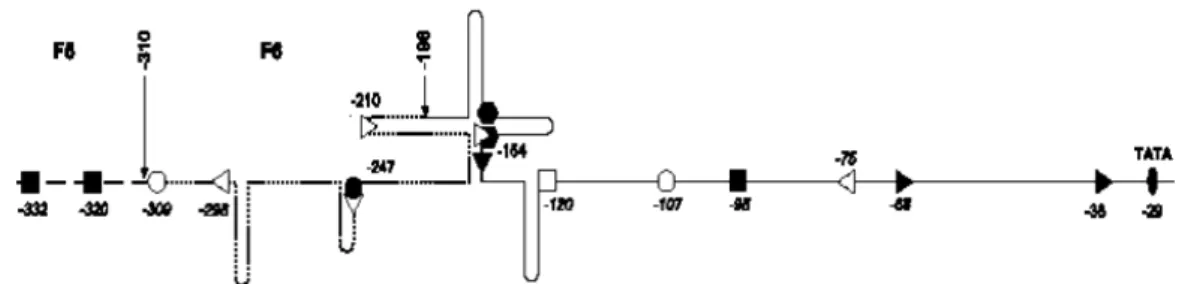
Figura I.6: Promotori ed enhancer di TTV in relazione alla struttura
terziaria dell’UTR. Siti conservati per il legame dei fattori di trascrizione: cerchio vuoto, CREB; cerchio pieno. E2F; quadrato vuoto c-Ets, quadrato pieno, USF; esagono SP1; le posizioni sono indicate rispetto al sito di inizio trascrizione (+1 nt). Modificata da Kamada et al., 2004.
1.5 ETEROGENEITÀ GENETICA
TTV e TTMV mostrano una straordinaria eterogeneità genetica, peraltro evento molto inconsueto fra i virus a DNA; l’identificazione delle numerose specie virali altamente divergenti ma correlate a TTV e la rapidità con cui ultimamente vengono isolati nuovi ceppi virali del gruppo TT ne rappresentano una chiara testimonianza.
Tale variabilità genetica riguarda principalmente la regione genomica codificante di TTV, ed è massima nella zona centrale della ORF1 (si osserva soltanto il 32% di omologia fra l’isolato SANBAN e il prototipo TA278) dove sono stati identificati almeno tre segmenti ipervariabili. Queste regioni, denominate HVR-1, HVR-2 e HVR-3 codificanti rispettivamente per 22, 47 e 31 aminoacidi nel caso del genotipo 1, presentano codoni multipli di inserzione o delezione ed una diversità genetica a livello aminoacidico superiore al 70% (Erker et al., 1999; Hijikata et al., 1999; Takahashi et al., 1998b). In uno studio condotto da Nishizawa e collaboratori nel quale è stata analizzata, in un periodo compreso tra tre e otto anni, la sequenza dei prodotti di PCR dedotti dall’ORF1 di due soggetti, è stato osservato che le regioni HVR presentano un grado maggiore di variabilità in soggetti con infezione cronica e che tale variabilità diviene particolarmente pronunciata durante la fase acuta dell’infezione, suggerendo un probabile coinvolgimento di queste regioni nei meccanismi utilizzati dal virus per sfuggire alla risposta
immunitaria. Si ritiene inoltre che le HVR siano implicate nella formazione della quasispecie di TTV, come del resto si verifica con l’HVR di HCV e la V3 di HIV (Nishizawa et al., 1999).
Tuttavia, la stessa variabilità all’interno delle HVR, non è stata riscontrata per tutti gli isolati di TTV: in un periodo di osservazione compreso tra tre e otto anni le sequenze degli isolati di genotipi 12 e 13 non hanno mostrato particolari variazioni (Ukita et al., 2000).
Un ulteriore meccanismo ipotizzato per spiegare l’elevata eterogeneità di TTV è rappresentato dall’utilizzo preferenziale, da parte del virus, di polimerasi cellulari con scarsa capacità di “correzione di bozze” durante la replicazione. Sebbene tale processo non sia stato ancora dimostrato, è avvalorato dal fatto che la frequenza con cui avvengono le sostituzioni nucleotidiche in alcune regioni del genoma (7,3 x 10-4 per sito per anno nelle HVRs di
SENV) risulta simile a quella di virus ad RNA, cioè circa 10 volte maggiore di quella di altri virus a DNA, come ad esempio, il virus dell’epatite B (HBV; Umemura et al., 2001b).
Un’altra ipotesi avanzata per spiegare l’alto grado di eterogeneità genetica è la possibilità che tra isolati diversi o addirittura all’interno di uno stesso ceppo virale, si verifichino eventi multipli di ricombinazione (Worobey, 2000; Biagini et al., 2001b; Manni et al., 2002). Sono stati identificati, infatti, 19 siti di ricombinazione omologa, di cui 13 appartenenti alla regione UTR (Worobey, 2000;
Biagini et al., 2001b); tale regione quindi potrebbe avere un ruolo importante nell’amplificazione dell’eterogeneità genetica (Erker et al., 1999; Hijikata et al., 1999). La ricombinazione, oltre che dalle caratteristiche del genoma, potrebbe essere facilitata dalla presenza di diversi isolati virali di TTV che spesso circolano contemporaneamente nel sangue o infettano lo stesso distretto corporeo dell’ospite (Worobey, 2000; Niel et al., 2000; White et al., 2000). Anche per TTMV è stata dimostrata la contemporanea presenza di ben 15 differenti isolati nel sangue di un soggetto immunocompromesso (Vasconcelos et al., 2002).
Tuttavia c’è da sottolineare, come emerso da un recente lavoro, che non vi è ancora concordanza di opinioni circa l’ipotesi della ricombinazione come causa primaria della elevata eterogeneità genetica di TTV e TTMV (Jelcic et al., 2004).
Alcuni autori hanno ipotizzato che gli eventi di ricombinazione possano coinvolgere, pure se con frequenza minore, virus animali correlati: molti ceppi simili a quelli identificati nell’uomo sono stati isolati anche in primati non umani ed in altre specie animali, come cavalli, maiali, capre e galline (Leary et al., 1999; Gibbs et Weiller, 1999; Martelli et al., 2006). Sebbene non sia ancora noto quale importanza possa rivestire, ai fini epidemiologici, la presenza dei genomi TTV-simili in questi animali, è ipotizzabile che questi ospiti
mediante il passaggio interspecie possano contribuire all’evoluzione genetica del virus.
Infine anche la considerazione che TTV sia un virus antico, che circola nella popolazione umana da migliaia di anni, può giustificare il grado di variabilità genetica che potrebbe essersi accumulata nel corso di lunghi periodi (Hijikata et al., 1999; Prescott et al., 1999).
1.6 REPLICAZIONE
I componenti della famiglia del genere Anellovirus adottano per replicare il proprio genoma, delle modalità che al momento rimangono ancora tutte da chiarire; è possibile solo formulare delle ipotesi basandosi sulle analogie condivise da tali virus con alcuni membri della famiglia Circoviridae ed in particolare con CAV e il circovirus porcino di tipo 2 (PCV-2). La mancanza di sequenze codificanti per DNA polimerasi o per retrotrascrittasi nel genoma dei circovirus animali, suggerisce che la loro replicazione è strettamente dipendente da enzimi di origine cellulare. Inoltre, l’assenza di sequenze capaci di alterare il ciclo cellulare della cellula ospite (Todd et al., 2001) e la capacità dei circovirus di propagarsi nelle cellule linfoidi attivate, lasciano supporre che il loro processo di replicazione dipenda dallo stato di attivazione della cellula, e in particolare da proteine prodotte durante la fase S del ciclo cellulare (Fields et al., 1996). Anche per TTV è stata dimostrata la necessità di una replicazione in cellule in attiva divisione come quelle del fegato, del midollo osseo, (Okamoto et al., 2000d; Okamoto e Mayumi, 2001) e del sistema linfoide stimolate (Maggi et al., 2001; Mariscal et al., 2001); ciò induce a pensare che il processo replicativo avvenga, almeno in parte, nel nucleo mentre non si dispone di nessuna informazione sulle modalità di assemblaggio del virione. Inoltre, considerando l’elevata eterogeneità genetica che contraddistingue i
virus del gruppo TT (Umemura et al., 2001b), è probabile che la funzione di replicasi sia svolta da polimerasi di origine cellulare con scarsa attività di “correzione di bozze”. Resta ancora da chiarire anche le modalità con cui TTV viene rilasciato dalla cellula ospite anche se tuttavia la mancanza di un involucro lipoproteico porta ad ipotizzare che la liberazione dei virioni maturi avvenga per lisi cellulare.
Per la replicazione del genoma di TTV, in virtù delle somiglianze con i circovirus e in analogia ai virus vegetali e batterici con DNA monocatenario circolare, è stato proposto un meccanismo a cerchio rotante che porterebbe alla formazione di un intermedio replicativo a doppio filamento circolare. Tale ipotesi è rafforzata dall’individuazione di alcuni motivi aminoacidici tipici delle proteine Rep all’interno della regione ORF1 del virus; tali proteine vengono utilizzate dai virus a DNA che si replicano mediante il meccanismo suddetto (Erker et al., 1999); questa ipotesi è ulteriormente avvalorata dal ritrovamento nelle cellule del fegato e del midollo osseo dell’intermedio replicativo di TTV a doppio filamento di DNA (Okamoto et al., 2000d). Nonostante ciò, la mancanza della sequenza nonanucleotidica indispensabile per le interazioni iniziali durante la replicazione a cerchio rotante, fa dubitare sul reale utilizzo di TTV di questo modello replicativo (Erker et al., 1999).
È stata inoltre indagata l’eventuale capacità di TTV di replicarsi attraverso un intermedio ad RNA (come si verifica per il virus dell’epatite B, HBV), ma non avendo trovato nel suo genoma alcun motivo codificante per una retrotrascrittasi virale, si ritiene che tale possibilità sia improbabile (Nishizawa et al., 1999).
Il ritrovamento del genoma di TTV in un elevato numero di campioni diversi, ha evidenziato la capacità da parte del virus di infettare un ampio spettro di tipi cellulari; ciò fa presupporre che i recettori utilizzati da TTV durante la fase di riconoscimento cellulare siano probabilmente molecole distribuite a livello di differenti tipi di tessuti, sebbene non sia escluso l’uso di specifici corecettori da parte dei diversi genogruppi e/o genotipi del virus. In questa ottica è importante sottolineare la presenza di porzioni genomiche ipervaribili (HVR) localizzate nella parte centrale della regione codificante ORF1, le quali potrebbero indirizzare il tropismo cellulare di TTV (Takahashi et al., 2002).
1.7 METODI PER LA RIVELAZIONE
DIAGNOSTICA DI TTV
Le tecniche di laboratorio utilizzate per l’identificazione di TTV e TTMV, mancando sistemi di coltivazione e metodi sierologici, sono sostanzialmente basate sulla ricerca e l’amplificazione molecolare degli acidi nucleici virali. La notevole variabilità genetica che caratterizza tali virus, ha reso critica la scelta della regione genomica da amplificare. La regione che è stata inizialmente utilizzata come bersaglio dell’amplificazione è la porzione N22 del gene ORF1 (PCR N22); ma tale regione, non essendo conservata fra tutti gli isolati di TTV, si è rilevata adatta solo per l’identificazione dei genotipi dall’1 al 6 (Muljiono et al., 2001). Le conoscenze sempre più approfondite del genoma hanno successivamente permesso di individuare un tratto dell’acido nucleico virale molto più conservato tra tutti gli isolati di TTV; questo tratto appartenente alla regione UTR ha permesso di amplificare con buona sensibilità tutti i genotipi conosciuti di TTV (PCR UTR, o universale) (Okamoto e Mayumi, 2001). Gli isolati virali, una volta identificati e quantificati con Real-Time PCR, possono essere successivamente caratterizzati mediante PCR genogruppo-specifiche dedotte da diverse regioni del genoma (Maggi et al., 2001c).
Anche per identificare l’infezione da TTMV le metodiche disponibili si basano sull’amplificazione di una porzione dell’UTR (Biagini et al., 2001a).
L’utilizzo della PCR UTR ha notevolmente incrementato la percentuale dei soggetti risultati positivi per TTV: si è passati infatti dal 23 al 92% nello studio di Takahashi (Takahashi et al., 1998a), dal 9 al 50% nello studio di Irving (Irving et al., 1999) e dal 20 al 95% in quello di Itoh (Itoh et al., 1999b). Tuttavia, un quadro definitivo sulla reale prevalenza della diffusione di TTV si potrà avere solo con lo sviluppo di test sierologici che consentono l’identificazione di forme pregresse d’infezione. Nei pochi studi effettuati, sono stati eseguiti saggi di immunoblotting o di immunoprecipitazione combinata alla PCR; come antigeni sono state utilizzate particelle virali intere o frammenti della proteina ORF1 o ORF2 espressi in batteri. Attraverso un saggio di immunoprecipitazione, nel siero di alcuni pazienti sono state rilevate IgG umane dirette contro il virus ed è stata quantificata la percentuale di TTV immunocomplessato; inoltre, utilizzando come antigene il genoma di TTV estratto da campioni fecali sono state rilevate anche le IgM (Nishizawa et al., 1999; Tsuda et al., 1999; Handa et al., 2000; Ott et al., 2000). Tali metodiche, però, non hanno fornito risultati riproducibili, come mostrato dai valori di prevalenza molto variabili emersi nei diversi studi (dal 10% al 98% dei soggetti sani sono risultati positivi per anticorpi anti-TTV di tipo G; Handa et
al., 2000; Ott et al., 2000; Tsuda et al., 2001; Kakkola et al., 2002). Probabilmente il tipo di antigene utilizzato è in grado di influenzare notevolmente la prestazione del saggio sierologico.
La messa a punto di metodi sierologici sensibili e specifici permetterebbe di individuare tutti quei soggetti che sono venuti a contatto con il virus in precedenza e che al momento dell’indagine sono risultati negativi per TTV; la risoluzione spontanea dell’infezione da TTV sembra infatti un evento possibile (Koboyashi et al., 1999; Matsumoto et al., 1999; Nishizawa et al., 1997).
1.8 EPIDEMIOLOGIA
L’aspetto sicuramente più interessante dell’epidemiologia di TTV è la sua elevata distribuzione in tutte le popolazioni del mondo indipendentemente dall’area geografica, dall’origine etnica e dallo sviluppo socio-economico.
I primi studi di prevalenza, in cui come bersaglio di amplificazione nelle reazioni di PCR era stata utilizzata la regione ORF1, avevano segnalato una discreta diffusione del virus, anche se con differenti percentuali di positività per TTV a seconda dell’area geografica considerata (Niel et al.,1999). Nei paesi industrializzati, Stati Uniti e Nord Europa, la frequenza di TTV risultava molto bassa. Al contrario, nelle popolazioni africane e sud americane si evidenziarono percentuali di positività assai elevate, mentre in Asia il virus era mediamente prevalente. Furono, inoltre, rilevati livelli di viremia particolarmente elevati in popolazioni isolate, ovvero limitatamente in contatto con le restanti regioni del mondo (Prescott e Simmonds, 1998). In tutti questi studi non fu osservata alcuna associazione della prevalenza di TTV con lo status clinico dei soggetti, né con altre caratteristiche della popolazione analizzata.
Con l’introduzione della PCR universale è stata dimostrata una diffusione del virus ancora più sorprendente di quanto ritenuto in precedenza; TTV è stato infatti ritrovato in circa 2/3 della popolazione mondiale, con variazioni non significative fra le differenti aree
geografiche (Biagini et al., 2000; Biagini et al., 2001a; Mastubara et al., 2001; Niel e Lampe, 2001).
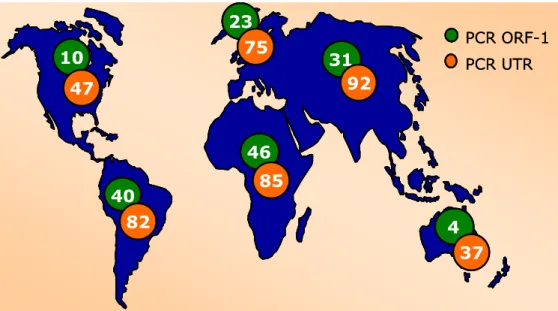
In base a questi studi TTV è risultato particolarmente diffuso in Asia con una prevalenza superiore al 90%, al contrario è risultato essere meno diffuso (circa 40%) in Nord America e in Oceania (figura I.7).
40 46 85 31 23 10 PCR ORF-1PCR UTR 92 82 75 4 37 47 40 40 46 46 85 85 31 31 23 23 10 10 PCR ORF-1PCR UTR 92 92 82 82 75 75 4 4 37 37 47 47
Figura I.7: Prevalenza di TTV nella popolazione. Le percentuali
indicate si riferiscono ai dati ottenuti amplificando regioni dell’ORF1 (verde) e dell’UTR (rosso), da Bendinelli et al., 2001.
È importante precisare che l’uso della PCR permette di rilevare solo le forme attive dell’infezione, sia acute che croniche, ma non evidenzia i casi di infezione pregressa: è quindi ipotizzabile che i livelli di prevalenza siano ancora più elevati di quelli attualmente osservati. L’isolamento di sequenze di TTV in diversi tessuti anche in assenza di viremia, confermerebbe tale ipotesi (Maggi et al., 2001c; Pollicino et al., 2002).
I dati disponibili per TTMV sono simili a quelli ottenuti per TTV (Biagini et al., 2000; 2001a). La distribuzione di entrambi i gruppi di virus non appare essere influenzata dal sesso dei soggetti esaminati, mentre, secondo alcuni autori, sembra che la frequenza degli individui TTV positivi aumenti con l’età (Oguchi et al., 1999; Yamada-Osaki et al., 1998); sebbene già nella prima infanzia sia stato possibile evidenziare un‘ampia diffusione del virus (Davidson et al., 1999; Hsieh et al., 1999; Vasconcelos et al., 2002; Maggi et al., 2003a), i livelli di prevalenza tendono ad aumentare nel tempo raggiungendo l’apice nell’età adulta (Chen et al., 1999; Davidson et al., 1999; Hsieh et al., 1999; Maggi et al., 1999).
L’elevata prevalenza di TTV e TTMV nella popolazione mondiale fa ipotizzare che tali virus si diffondano attraverso molteplici vie di trasmissione. L’alta percentuale di positività osservata in numerosi studi fra i soggetti esposti a sangue o agli emoderivati (emofiliaci, emodializzati, politransfusi, talassemici, e tossicodipendenti) ha reso
probabile l’ipotesi che TTV e TTMV possono essere facilmente trasmessi attraverso la via parenterale. Il ritrovamento di sequenze virali in campioni di feci di soggetti viremici ha suggerito che tali virus possono essere trasmessi anche per via oro-fecale. La determinazione di TTV in neonati ha inoltre fatto supporre l’esistenza di una via di trasmissione materna-fetale del virus sebbene in un primo momento questa ipotesi fosse stata smentita (Okamura et al., 1999; Hsieh et al., 1999; Yokazaki et al., 1999a). Al contrario il ritrovamento di TTV in più del 50% dei campioni di sangue cordale esaminati ha confermato questa ipotesi,suggerendo peraltro che le madri possano trasmettere l’infezione al figlio attraverso la placenta (Morrica et al., 2000); il genoma del virus è stato inoltre ritrovato in campioni di latte materno: questo spiega l’esistenza di neonati positivi con madri negative e di bambini che presentano isolati virali diversi da quelli identificati nel sangue materno (Davidson et al., 1999).
La trasmissione per via sessuale non è stata ancora chiarita ma, in alcuni studi, TTV e TTMV sono stati isolati dalle secrezioni vaginali, dal fluido spermatico e dalla saliva di alcuni soggetti (Gerner et al., 2000; Kazi et al., 2000; Inami et al., 2000; Liu et al., 2000; Martinez et al., 2000; Matsubara et al., 2000; Matsubara et al., 2001; Schroter et al., 2000; Stark et al., 2000; Biagini et al., 2001a; Chan et al.,
2001; Fornai et al., 2001; Krekulova et al., 2001; Pirovano et al., 2002a).
Uno studio recente effettuato su bambini di età inferiore ai due anni con malattie respiratorie acute ha indicato il tratto respiratorio come probabile sito di replicazione di TTV, e proposto la via aerea come un’ulteriore modalità di trasmissione del virus. I titoli virali determinati nelle secrezioni e nelle cellule nasali dei bambini esaminati risultavano infatti significativamente più alti se confrontati con i livelli osservati nei corrispondenti campioni di plasma (Maggi et al., 2003a). In precedenza erano già state individuate sequenze di TTV a livello dell’epitelio respiratorio, nei polmoni, nelle tonsille e nelle adenoidi, a sostegno dell’ipotesi dell’esistenza di una via di trasmissione aerea del virus (Pollicino et al., 2002).
Sequenze molto simili a quelle di TTV sono risultate frequenti in varie specie animali di allevamento e da compagnia come gatti e cani: tali ospiti potrebbero rappresentare dei serbatoi naturali di persistenza del virus, nonché possibili fonti di trasmissione interspecie dell’infezione (Leary et al., 1999; Okamoto et al., 2002).
Studi di prevalenza dei diversi genotipi di TTV e TTMV hanno evidenziato una maggiore diffusione di alcuni genotipi rispetto ad altri, come si evince da un lavoro recentemente pubblicato in cui i genotipi 1 e 3 sono stati ritrovati con maggior frequenza nella popolazione generale rispetto al 4 e 5, mentre il genogruppi 2
sembra essere quello più raro (Maggi et al., 2005). Da altri lavori è invece emerso un certo grado di variabilità nella distribuzione geografica dei vari isolati (Gallian et al., 2000; Mikuni et al., 2002) ad esempio, il genotipo 1 ed alcuni genotipi di SENV sono risultati più diffusi in Asia piuttosto che nei paesi occidentali con il 10% dei donatori giapponesi positivi per SENV-D e SENV-H rispetto al 2-3% dei donatori positivi statunitensi e italiani (Shibata et al., 2001; Umemura et al., 2001a). È stata anche dimostrata una differente prevalenza di alcuni genotipi di TTV fra paesi dello stesso continente: l’infezione con i genotipi 22 e 23 è risultata frequente in Indonesia, ma praticamente assente in Giappone (Muljono et al., 2001).
Rimane ancora da chiarire se i diversi genotipi vengano trasmessi con diversa efficienza; a tal proposito, in un recente studio sono stati identificati genotipi di TTV appartenenti al genogruppo 2 in soggetti HIV e HBV positivi, ma non in donatori di sangue (Devalle e Niel 2004); inoltre studi condotti da Pirovano e collaboratori hanno evidenziato una maggiore prevalenza di SENV-A e SENV-B in soggetti tossicodipendenti piuttosto che in individui infettati per via sessuale (Pirovano et al., 2002b).
1.9 INTERAZIONI CON L’OSPITE INFETTATO
I numerosi studi condotti su TTV hanno dimostrato che una volta acquisita l’infezione, il virus persiste per anni nel sangue della maggior parte dei soggetti infetti, (per almeno 22 anni in uno studio; Matsumoto et al., 1999) suggerendo che la cronicizzazione del virus è l’evenienza più comune. Alcuni lavori riportati in letteratura hanno evidenziato che più dell’80% degli individui infettati diviene portatore cronico del virus (Takayama et al., 1999b; Ali et al., 2002; Lefrère et al., 1999; Matsumoto et al., 1999). I meccanismi implicati nella persistenza sono attualmente sconosciuti, ma sembra da escludere che il virus rimanga latente nella cellula ospite in forma episomiale o integrata, fenomeno che risulta possibile per altri virus a DNA (Tanaka et al., 2000a; Yu et al., 2002). La persistenza di TTV potrebbe essere facilitata dall’incapacità del sistema immunitario dell’organismo ospite di controllare l’infezione. Gli anticorpi anti-TTV, sebbene siano prodotti durante l’infezione, non appaiono in grado, almeno nella maggior parte dei casi, né di eradicare l’infezione né di prevenire le superinfezioni; infatti, le infezioni miste con isolati eterologhi di TTV rappresentano un evento molto frequente, con percentuali del 65% rilevate nei bambini di età inferiore ai sette anni, e addirittura del 90% in soggetti adulti sani (Ukita et al., 2000; Saback et al., 2002; Maggi et al., 2005).La risposta umorale si manifesta circa 10-12 settimane dopo l’infezione, con la comparsa di IgM rilevabili nel sangue solo per 5-11 settimane, seguita intorno alla 15°-17° settimana,da una lenta produzione di IgG che solitamente persistono per anni (Tsuda et al., 2001). La capacità di questi anticorpi di neutralizzare il virus rimane ancora da dimostrare; dati contrastanti sono stati ottenuti soprattutto per le IgG, la cui presenza correla con la scomparsa o l’assenza della viremia plasmatica, mentre in altri casi non sembra aver prodotto alcun effetto sulla persistenza del virus. Studi condotti da Tsuda e collaboratori hanno evidenziato come la comparsa di immunocomplessi TTV-IgG sia seguita da una diminuzione del titolo virale (Tsuda et al., 1999). Al contrario, Nishizawa e collaboratori hanno dimostrato che, in pazienti cronicamente infetti, la maggior parte di TTV circola complessato ad IgG (Nishizawa et al., 1999). L’importanza della risposta cellulo-mediata nel controllo dell’infezione da TTV è un aspetto ancora da chiarire; si ritiene comunque che le cellule del sistema immunitario possano avere un ruolo essenziale sia nella persistenza che nella disseminazione virale ai vari tessuti. Un evento precoce dell’infezione da TTV è la comparsa del virus nel sangue, come più volte osservato in soggetti trasfusi ed anche in scimpanzè infettati sperimentalmente. In tutti questi casi, la viremia diveniva dimostrabile dopo 5-8 settimane dall’esposizione e spesso restava evidenziabile per tutto il periodo di osservazione (Mushahwar
et al., 1999; Tawara et al., 2001). I livelli plasmatici del virus nella fase cronica dell’infezione sono paragonabili a quelli di altri virus persistenti, come HCV, HBV e HIV ed oscillano fra 103 e 108 copie per ml di plasma (Pistello et al., 2001) mostrando ampie fluttuazioni nel tempo in alcuni soggetti ed una relativa stabilità in altri (Ball et al., 1999; Matsumoto et al., 1999). La cinetica del virus è stata indagata in un gruppo di pazienti HCV positivi cronicamente infettati da TTV misurando gli effetti dell’interferone (IFN) sulla viremia (Maggi et al., 2001c). Applicando un modello matematico simile a quello utilizzato per gli studi su HIV e HCV, è stato stimato che l’emivita di TTV nel plasma sia di circa 6 ore, che ogni giorno vengano prodotti almeno 1010 virioni e che più del 90% del virus plasmatico subisca un rinnovamento giornaliero (Maggi et al., 2001c; tabella I.1).
Tabella I.1: Dinamica della viremia plasmatica di TTV confrontata
con quelle di altri virus persistenti.
Virus Emivita del virione
N° minimo di virioni prodotti al
giorno
Referenza HBV 1,2 giorni > 1,7 x 10 13 Whalley et al. 2001
HCV 2,7 ore > 1,3 x 10 12 Neumann et al. 1998
HIV < 1 ora > 9,8 x 10 9 Ramratnam et al. 1999
Anche se la cronicizzazione dell’infezione è l’evenienza più comune, in letteratura non mancano casi in cui l’infezione sembra risolversi spontaneamente, talvolta anche dopo anni o mesi dal contagio (Lefrère et al., 1999; Matsumoto et al., 1999; Yuki et al., 1999). In realtà la scomparsa della viremia osservata in questi casi potrebbe non significare la completa eradicazione del virus; a conferma di ciò sono state infatti isolate sequenze di TTV in diversi tessuti anche in assenza di viremia (Maggi et al., 2001c; Pollicino et al., 2002). L’apparente risoluzione dell’infezione potrebbe dipendere da una ridotta immissione in circolo del virus dovuta a fasi di latenza, da una modificazione delle sequenze virali che non permettono più la rilevazione del DNA, o semplicemente da un aumento della sua eliminazione. L’andamento dell’infezione potrebbe inoltre differire a seconda del genotipo infettante; infatti in uno studio condotto da Otho e collaboratori è stata osservata la risoluzione spontanea dell’infezione in circa il 20% dei bambini infettati da TTV, ma con diverse percentuali a seconda del genotipo virale coinvolto nell’infezione (Otho et al., 2002). Un ulteriore studio ha evidenziato la scomparsa della viremia nel 77% di soggetti tossicodipendenti infettati con l’isolato SENV (Wilson et al., 2001). Tuttavia sono necessari ulteriori studi in tale direzione, prima di poter affermare con certezza che i genotipi di TTV possiedano una differente capacità di persistenza.
Il genoma di TTV è stato identificato in numerosi fluidi biologici tra cui bile, saliva, lacrime, seme, secrezioni vaginali, fluido duodenale, liquido sinoviale e fluido cerebrospinale (Deng et al., 2000; Biagini et al., 2001a; Tapiri et al., 2001; Maggi et al., 2001a; Fornai et al., 2001; Nakagawa et al., 2000; Inmai et al., 2000), ad indicare che il virus è disseminato in tutto l’organismo. Inoltre, sono stati ritrovati intermedi replicativi e trascritti virali in molti tessuti (polmone, milza, pancreas, rene, linfonodi, muscolo scheletrico e tiroide; Okamoto et al., 2001) a dimostrazione della capacità del virus di replicarsi in molti tessuti infettati (Okamoto et al., 2001) e non solo nelle cellule epatiche o in quelle del midollo osseo, come si pensava precedentemente (Okamoto et al., 1998b; Okamoto et al., 2000b; Okamoto et al., 2000d; Kikuchi et al., 2000; Lopez-Alcorocho et al., 2000; Rodriguez-Inigo et al., 2000; Comar et al., 2002). Questi risultati hanno fornito importanti indicazioni, ma la mancanza di sistemi di coltivazione idonei per TTV, non ha ancora permesso l’identificazione di tipi cellulari in grado di supportarne la replicazione. Tuttavia, recenti studi
in vitro hanno dimostrato che TTV è in grado di infettare i
linfo-monociti, ma la sua replicazione avviene solo quando questi ultimi sono opportunamente stimolati. Ciò suggerisce che la capacità di replicazione di TTV dipenda dallo stato di attivazione della cellula ospite, come del resto accade per molti altri virus a DNA (Maggi et al., 2001b; Mariscal et al., 2002).
Non è ancora chiaro se esista una sottopopolazione linfocitaria nella quale il virus preferenzialmente si replichi e/o persista, ma in alcuni lavori sono stati osservati quantità di virus maggiori a livello dei granulociti e dei linfociti CD19+ (Lopez-Alcorocho et al., 2000;
Takahashi et al., 2002; Yu et al., 2002; Zhong et al., 2002). Anche in questo caso è ipotizzabile che diversi genotipi di TTV abbiano un tropismo cellulare preferenziale, come suggerito dalla discordanza tra le sequenze individuate nei tessuti di un singolo paziente e quelle presenti in circolo (Okamoto et al., 2000c). Al contrario il recente ritrovamento di ben 24 isolati del virus nella milza di un paziente affetto da tumore sembra smentire le precedenti ipotesi (Jelcic et al., 2004).
Moen e collaboratori hanno infine indagato la possibilità che TTV e TTMV abbiano un ruolo come agenti immunomodulanti, similmente a quanto dimostrato per i circovirus animali. Per far luce su tale ipotesi sono stati arruolati tre gruppi di pazienti sottoposti a trattamenti immunosoppressivi diversi: i risultati hanno evidenziato cambiamenti nei titoli di TTV in base alle variazioni delle risposte di difesa fisiologiche dell’ospite (Moen et al., 2002b). In altri lavori, a conferma di quanto emerso negli studi condotti da Moen, è stata osservata una diminuzione della viremia basale in seguito a trapianti di midollo osseo o a trattamenti mielosoppressivi (Kanda et al., 1999b).
1.10 ASSOCIAZIONE CON LA MALATTIA
Dalla scoperta di TTV ad oggi sono stati compiuti numerosi studi per cercare di far luce sulla reale capacità patogenetica del nuovo virus. Nonostante i risultati di molti lavori lascino aperta la possibilità che l’infezione da TTV e TTMV possa avere un qualche significato clinico, non si è ancora arrivati ad una conclusione certa che definisca tale argomento (Naoumov, 2000).
Sulla base dell’elevata prevalenza di TTV e TTMV nella popolazione sana, della loro ubiquitarietà nell’ospite infettato e dell’esistenza di diverse modalità di trasmissione, alcuni autori si sono affrettati a ritenerli semplici commesali apotogeni dell’uomo (Griffiths, 1999; Mushahwar, 2000). Poiché TTV è stato identificato per la prima volta in pazienti con forme di epatite non-A non-G (Nishizawa et al., 1997), è stato proposto che il virus fosse la causa di epatiti croniche o acute ad eziologia sconosciuta. Tuttavia, studi successivi hanno sollevato molti dubbi su tale potenzialità patogenetica del virus, in quanto la presenza di TTV è stata dimostrata non solo in pazienti con epatite criptogenetica ma anche in soggetti privi di alterazioni epatiche o addirittura senza alcuna manifestazione patologica (Berg et al., 1999; Colombatto et al., 1999; He et al., 1999; Kanda et al.,1999; Nakano et al., 1999; Toniutto et al., 1999; Viazov et al., 1998). Inoltre, scimpanzè naturalmente o sperimentalmente infettati con TTV o con virus correlati non mostravano evidenze biochimiche o istologiche di
danno epatico (Mushahwar et al., 1999; Verschoor et al., 1999). Non è stata neppure dimostrata, in studi condotti su pazienti trasfusi e/o coinfettati con il virus dell’epatite B (HBV) o il virus dell’epatite C (HCV), alcuna correlazione tra l’acquisizione dell’infezione da TTV e la progressione dell’epatite e/o l’entità del danno epatico (Giménez-Barcons et al., 1999; Irving et al., 1999; Lefrère et al., 1999; Matsumoto et al., 1999; Oguchi et al., 1999; Berg et al., 1999; Gad et al., 2000; Kao et al., 2000). I risultati emersi da tutti questi studi hanno pertanto portato ad escludere un ruolo primario del virus del gruppo TT nella patologia epatica (Matsumoto et al., 1999; Kao et al., 2000; Shang et al., 2000; Ali et al., 2002). Comunque in letteratura sono presenti alcuni studi in cui è stata rilevata una possibile associazione tra danno epatico e TTV, come dimostrato dalle sporadiche correlazioni fra viremia ed andamento delle transaminasi in alcuni soggetti (Cleavinger et al., 2000; Forns et al., 1999; Fujiwara et al., 1998; Ikeda et al., 1999; Itoh et al., 1999a; Jiang et al., 1999; Kanda et al., 1999b; Lefrère et al., 1999; Luo et al., 1999; Naoumov et al., 1998).
Inoltre, in uno studio condotto su pazienti con epatite fulminante, è stata osservata una mortalità del 100% in soggetti TTV positivi e del 50% in quelli negativi (Tanaka et al., 1999): non è quindi da escludere un coinvolgimento occasionale di TTV nell’aggravamento di quadri patologici a carico del fegato.
In recenti lavori, data la presenza e la capacità replicativa di TTV e TTMV in molti tessuti diversi da quello epatico, è stata valutata la possibile associazione di tali virus con altri tipi di patologia. È emersa infatti un’interessante relazione tra l’infezione da TTMV e gravi patologie dell’apparato renale (Gallian et al., 2002); è stato inoltre evidenziato, in un esperimento condotto in vitro, un possibile ruolo di TTV nel provocare alterazioni in cellule renali simili a quelle osservate nella sindrome nefrosica (Yokoyama et al., 2002). In questo lavoro topi transgenici per varie porzioni dell’ORF1 mostravano segni patologici solo a livello del rene, tanto più gravi quanto maggiore era l’espressione del transgene, a conferma che il danno renale era indotto dalla proteina codificata. Un ulteriore studio, condotto su un gruppo di pazienti sottoposti a trapianto renale, ha sottolineato la persistenza di isolati del genogruppo 1 in questo organo (Szladek et al., 2003).
Al contrario, non è emersa alcuna relazione tra TTV ed anemia aplastica, crioglobulinemia, lupus eritematoso, psoriasi (Kikuchi et al., 2000; Cacoub et al., 2003; Maggi et al., 1999) e nemmeno con la presenza di infezione da Helicobacter pilori (Toyoda et al., 2000). Parziali associazioni sono invece state dimostrate in pazienti affetti da diabete, fibrosi polmonare idiomatica, artrite reumatoide ed in emodializzati con bassi livelli di emoglobina e scarsa risposta al trattamento con eritropoietina (Gallian et al., 1999; Bando et al.,
2001; Boysen et al., 2003; Hirata et al., 1998). Inoltre, sono stati evidenziati elevati titoli di TTV in bambini con broncopolmonite (BP) piuttosto che in quelli affetti da forme meno gravi di MRA, suggerendo un possibile ruolo di TTV come cofattore nella patologia di alcune affezioni delle vie aeree (Maggi et al., 2003a).
Recentemente, tenendo conto dell’omologia che TTV mostra con alcuni circovirus animali, è stata presa in esame l’ipotesi che il virus interagisca con il sistema immunitario dell’organismo ospite (Bendinelli et al., 2001; Simmonds, 2002). A conferma di ciò, sono emerse interessanti correlazioni tra TTV e patologie del sistema linfoide: studi condotti da Shibayama e collaboratori hanno evidenziato livelli particolarmente elevati del virus in pazienti immunocompromessi (affetti da AIDS o da altre sindromi intercorrenti), suggerendo un possibile ruolo del sistema immunitario nel controllo della replicazione di TTV; i titoli del virus, quindi, aumenterebbero in risposta ad un progressivo indebolimento della risposta immunitaria (Shibayama et al., 2002; Simmonds, 2002). Tutto ciò è stato anche confermato dall’incremento della viremia di TTV registrato in pazienti sottoposti a trapianto epatico o a terapia con farmaci immunosoppressivi (Shang et al., 2000), oltre che da una correlazione inversa fra i livelli quantitativi del virus ed il numero dei linfociti CD4+ in pazienti HIV positivi (Christensen et al., 2000; Shibayama et al., 2001; Touinssi et al., 2001). Tuttavia, non è stato
osservato un aumento dei livelli di TTV in pazienti HIV positivi; ciò induce ad ipotizzare che in seguito alla deplezione HIV-indotta dei linfociti CD4+, vengano prodotti, solo occasionalmente, fattori necessari per l’incremento del titolo di TTV (Moen et al., 2002a); comunque il reale contributo di TTV nella progressione dell’HIV resta ancora da chiarire.
Le somiglianze con i circovirus animali hanno inoltre suggerito l’eventualità che alcune delle complicanze ematologiche osservate in pazienti immunocompromessi, come la neutropenia, la trombocitopenia, e l’anemia possano dipendere in qualche misura dall’incremento dei livelli di TTV nel midollo osseo (Kikuchi et al., 2000; Tokita et al., 2001c; Simmonds, 2002).
Una relazione tra TTV ed alterazioni del sistema immunitario è emersa da uno studio condotto su bambini affetti da MRA, in cui è stata osservata una correlazione tra i titoli del virus ed i livelli di alcune sottopopolazioni linfocitarie circolanti (in particolare con i linfociti T CD3+ e CD4+ e con i linfociti B CD19+; Maggi et al., 2003b). L’elevata variabilità genetica che caratterizza i virus del gruppo TTV e TTMV ha suggerito anche la possibilità che i diversi genotipi abbiano un differente ruolo in relazione a determinate patologie, come del resto si verifica per alcuni membri della famiglia
Circoviridae: il circovirus porcino (PCV), ad esempio, provoca una
ma tale potenzialità patogenetica è attribuibile soltanto al PCV di tipo 2.
Gli studi condotti al fine di chiarire questo importante aspetto hanno fornito interessanti risultati: è emersa, infatti, una correlazione fra il genotipo 1 di TTV e i livelli di transaminasi in soggetti con patologie epatiche e in più, tale genotipo, risultava particolarmente frequente in pazienti con epatite fulminante o con aggravamento delle condizioni cliniche (Okamura et al., 2000; Shibata et al., 2000; Moriyama et al., 2001; Tokita et al., 2001a; 2001b; 2001c; Tokita et al., 2002). Inoltre è stato dimostrato, in uno studio condotto da Umemura e collaboratori, come due genotipi di SENV (SENV-D e SENV-H) fossero maggiormente presenti nel siero di pazienti con epatite post-transfusionale non-A non-G piuttosto che in soggetti sani (Umemura et al., 2001a). Tuttavia, studi successivi hanno riportato risultati contrastanti (Kao et al., 2002; Mikuni et al., 2002; Yoshida et al., 2002).
1.11 SENSIBILITÀ AI FARMACI ANTIVIRALI
Le attuali conoscenze sulla sensibilità di TTV ai farmaci antivirali deriva da studi effettuati in pazienti coinfettati da altri virus, in particolare da HCV; di conseguenza quasi tutte le osservazioni riguardano il comportamento della viremia di TTV in corso di terapia con interferone (IFN). Le risposte ottenute per TTV sono risultate paragonabili a quelle per HCV. La terapia con IFN induce, in circa il 50% dei pazienti, una scomparsa della viremia plasmatica di TTV, già evidente dopo una o due settimane di somministrazione e persistente per tutta la durata del trattamento (Kao et al., 2000; Dai et al., 2002). Nel rimanente 50% si registrano alcuni casi in cui la terapia con IFN non esercita effetti significativi sui livelli della viremia, mentre in altri casi il titolo virale varia considerevolmente durante il periodo di osservazione, per poi tornare, al termine del trattamento, a livelli basali (Akahane et al., 1999; Berg et al., 1999; Chayama et al., 1999; Maggi et al., 1999). Non è tuttavia escluso che in molti soggetti la ricomparsa di TTV alla fine del trattamento possa essere dovuta ad una nuova infezione (Ali et al., 2002). Vi sono scarsi elementi per definire quali parametri siano responsabili dei diversi tipi di risposta osservati. Tuttavia la presenza di bassi livelli di viremia pre-trattamento di isolati di TTV filogeneticamente distanti dal prototipo TA278 si sono dimostrati parametri predittivi di una migliore risposta all’IFN (Akhane et al., 1999; Chayama et al., 1999).
La combinazione di IFN con ribavirina mostra, allo stesso modo di quanto osservato per HCV, una maggiore efficacia nell’indurre una risposta a lungo termine verso TTV (Berg et al., 1999). Non è attualmente noto se esistano differenze nella suscettibilità all’IFN fra i differenti genotipi di TTV. Tuttavia, isolati di SENV sono risultati particolarmente sensibili all’azione dell’IFN: il 60-80% dei soggetti infettati mostrava infatti la scomparsa del virus circolante dopo trattamento farmacologico (Umemura et al., 2002; Wong et al., 2002). Contrariamente a quanto osservato con l’IFN, la terapia antiretrovirale in pazienti HIV positivi non ha alcun effetto sulla viremia da TTV, né la presenza del virus sembra alterare l’efficacia dei farmaci antiretrovirali (Watanabe et al., 2000; Kawanaka et al., 2002).
2. ASMA
L’asma è una delle malattie respiratorie croniche più diffuse nel mondo, presente in tutti i paesi anche se con livelli molto variabili. Rappresenta quindi un consistente problema di sanità pubblica, anche perché la sua prevalenza è in aumento soprattutto nei paesi industrializzati e ciò non può essere imputabile a fattori genetici. E’ stato ritenuto che, probabilmente, il fattore centrale dell’aumento della prevalenza dell’asma sia rappresentato dalle modificazioni ambientali spesso molto rapide in rapporto alle capacità di adattamento degli organismi biologici. Tra i fattori ambientali sospettati di questo ruolo causale o comunque scatenante l’asma sono stati considerati l’inquinamento atmosferico insieme all’esposizione passiva al fumo di tabacco, ma anche l’esposizione agli allergeni e le infezioni respiratorie, specialmente di origine virale. Nella maggior parte dei casi i sintomi compaiono prima dei 6 anni, ma in questa fase difficilmente possono essere ricondotti ad una diagnosi precisa. Infatti, il wheezing, soprattutto se indotto da infezioni virali è una manifestazione clinica assai frequente nei primi anni di vita ed è molto difficile identificare, nei primi anni di vita, quali tra i bambini con tale sintomatologia continueranno a presentare manifestazioni di tipo asmatico nelle età successive.
La caratterizzazione precisa della patologia è stata sancita nell’ambito del progetto “G.I.N.A” (Global Iniziative for Asthma), promosso nel 1992 dal NHBLI (National Heart Lung and Blood Institute USA) e dall’OMS (Organizzazione Mondiale della Sanità) con lo scopo si sviluppare una strategia mondiale per ridurre la mortalità e la morbilità di tale patologia. L’asma viene definita come “ una malattia infiammatoria cronica a carico delle vie aeree nella quale giocano un ruolo numerose cellule, in particolare i mastociti, gli eosinofili ed i linfociti T. Nei soggetti predisposti questa infiammazione provoca episodi ricorrenti di respiro sibilante, dispnea, senso di costrizione toracica e tosse, in particolare durante la notte e/o al mattino presto. Questi sintomi sono di solito associati con diffusa, ma variabile ostruzione del flusso aereo che è almeno parzialmente reversibile spontaneamente o dopo terapia. L’infiammazione provoca, inoltre, un aumento della responsività delle vie aeree a numerosi stimoli irritativi”.
L’infiammazione delle vie aeree è quindi considerata la più importante caratteristica dell’asma, sia in quanto causa primaria della malattia, sia come fattore in grado di condizionare la severità del decorso. È responsabile, infatti, delle riacutizzazioni e di alterazioni della struttura delle vie aeree (rimodellamento) che sono alla base della persistenza dei sintomi nel tempo, della limitazione variabile del flusso aereo e della ipereattività bronchiale, caratteristiche della
malattia. Come indicato nelle raccomandazioni delle linee guida G.I.N.A., è quindi necessario individuare appropriati test non invasivi da applicare nella ricerca e nella pratica clinica, per la diagnosi, il monitoraggio e il trattamento della flogosi delle vie aeree.
Fino a pochi anni fa non è stato comunque possibile misurare in modo semplice l’infiammazione delle vie aeree che veniva valutata raccogliendo campioni biologici (cellule endoluminale delle vie aeree e componenti molecolari) mediante lavaggio bronco-alveolare o campioni bioptici prelevati dalla biforcazione dei bronchi prossimali in corso di broncoscopia. Tali metodi, però, sono invasivi mal tollerati dal paziente e non privi di rischi nei soggetti ostruiti; pertanto non sono considerati adeguati per un monitoraggio ripetuto, soprattutto nei bambini.
Negli ultimi anni la ricerca si è focalizzata su tecniche non invasive, come l’analisi dei marker dell’infiammazione nell’aria espirata. Tra questi, la misura dell’ossido nitrico (NO) ha rappresentato una risposta alle necessità sopra discusse ed ha aperto nuove prospettive per lo studio dell’infiammazione nelle malattie respiratorie. Una delle più importanti caratteristiche della misura dell’NO nella pratica clinica consiste nel fatto che i risultati si ottengono in pochi minuti e possono, quindi, essere utilizzati nel momento stesso in cui si valuta il paziente.