2 Le caratteristiche meccaniche del tessuto nervosoperiferico
La totalità dei lavori reperiti in letteratura, che prendono in esame le caratteristiche meccaniche del tessuto del sistema nervoso periferico, forniscono, accanto ad una descrizione delle procedure e delle attrezzature usate, i grafici delle grandezze misurate. Data la conformazione geometrica dei campioni nervosi usati negli esperimenti, aventi dimensioni della sezione trasversale nettamente inferiori al loro sviluppo in lunghezza, la totalità delle prove svolte ha carattere monoassiale in senso longitudinale.
2.1 La prova di trazione
La più classica delle prove meccaniche a cui sono stati sottoposti campioni di nervi provenienti dal sistema nervoso periferico è la prova monoassiale di trazione. In un primo momento lo scopo prevalente è stato quello di individuare valori che indicassero con una certa esattezza il limite di resistenza a trazione del materiale. Questo perché il limite di funzionalità biologica del tessuto nervoso era considerato coincidente con il suo limite di rottura. Diversi studi, soprattutto in fase iniziale, sono stati compiuti su cadaveri umani. Nel 1866 venne scoperto da Tillaux che nervi ulnari e mediani avevano la capacità di resistere ad un carico variabile fra i 196 N e i 245 N, mostrando un allungamento dal 34% al 45% prima di rompersi. Nel 1880 Trombetta ha quantificato in 373 N il carico di rottura del nervo ulnare, Takimoto, nel 1917, ha indicato il carico di rottura del nervo mediano in un intervallo compreso fra i 196 N e i 491 N. Sunderland e Bradley hanno fissato il limite di resistenza del nervo ulnare e di quello mediano rispettivamente nell’intervallo fra i 65 N e i 155 N e fra i 73 N e i 220 N (Grewal R. et al., 1996). Successivamente agli esperimenti effettuati su cadaveri si sono affiancati sempre più spesso quelli su modello animale e l ’interesse dei
ricercatori si è concentrato sulla determinazione della curva carichi -allungamenti. Si è quindi cercato di determinare il comportamento del tessuto nervoso sotto l’azione di un carico crescente in modo da avere informazioni più ampie, all’interno delle quali integrare quelle acquisite precedentemente, riguardanti il solo carico di rottura a trazione.
2.1.1 L’ottenimento dei campioni
Il primo passo è stato quello di ottenere alcuni campioni del materiale nervoso su cui potesse essere effettuata la prova di trazione. In letteratura i campioni nervosi scelti sono stati prelevati in un momento di poco successivo alla morte e utilizzati nell’arco di poche ore (Arnold G. e Lutge H., 1972), (Rydevik B.L. et al., 1990), (Kwan M.K. et al., 1992), (Millesi H. et al., 1995), (Walbeehm E.T., et al., 2004). In alternativa si è ricorsi all’aggiunta di sostanze chimiche che però alterano la struttura cellulare dei campioni e quindi, anche se non mutano la loro composizione a livello dei materiali costituenti, hanno influenza sulle loro caratteristiche strutturali (Borschel et al., 2003).
2.1.2 I metodi sperimentali
I campioni, prelevati al momento o conservati, devono poter essere montati sulla macchina di misura scelta per caratterizzare le loro proprietà meccaniche. Come già detto più sopra, data la loro geometria, le prove effettuate hanno, nella totalità dei casi, la caratteristica di essere monoassiali nel senso della lunghezza del nervo. Un primo problema che si è posto agli sperimentatori è stato quello del fissaggio delle estremità del campione alla macchina. In passato si è risolto questo problema mediante sutura, ma è risultato in seguito che proprio il materiale del filo usato per la sutura alterava il comportamento del campione (Grewal R. et al., 1996). Il problema è stato affrontato e risolto mediante speciali morse ad alta
al., 1990), (Kwan M.K. et al., 1992), ultimamente con l’ausilio di semplici morse (Fig. 2.1) nelle quali il campione è trattenuto presumibilmente per attrito (Walbeehm E.T., et al., 2004).
2.1.3 La curva forza – allungamento
Le macchine usate per la quantificazione delle proprietà meccaniche sono le stesse che vengono correntemente utilizzate nel campo dei materiali con funzione strutturale. In esperimenti con nervi tibiali di coniglio (Rydevik B.L. et al., 1990), (Kwan M.K. et al., 1992) è stato utilizzato un Instron (Los Alamitos, CA, U.S.A). In esperimenti con nervi sciatici di ratto (Walbeehm E.T., et al., 2004) la scelta è caduta su una macchina di misura Testometric 220 M (Testometric Co., Lanks., U.K.). Ciò che, evidentemente, risulta modificato, data la tipologia del materiale in esame, sono le condizioni di applicazione del carico e la risoluzione della cella.
Figura 2.1: Campione di tessuto nervoso preso fra le morse della macchina di misura Testometric 220 M
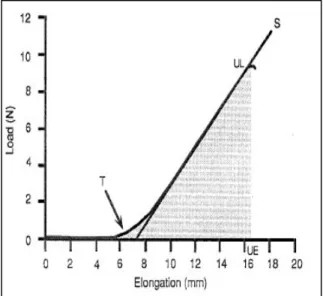
nasce l’esigenza di determinare l’allungamento a cui questo viene sottoposto dal sistema di forze esterne applicate alle sue estremità. In alcune esperienze (Rydevik B.L. et al., 1990), (Kwan M.K. et al., 1992), è stato scelto un video analizzatore in grado di fornire la distanza relativa fra due riferimenti ottenuti sul tessuto nervoso grazie a colorazione chimica. In altre (Walbeehm E.T., et al., 2004) si è preferito misurare la distanza relativa fra due punti situati sulla macchina e implicati nel movimento, mediante un sensore LVDT (Linear Voltage Diffential Trasformer). Correlando fra loro queste informazioni si è riusciti a tracciare la curva forza - allungamento che ha il caratteristico andamento mostrato in Fig. 2.2.
E’ evidente la non linearità del comportamento globale dell’andamento della curva. Si attraversa, infatti, una prima regione chiamata “toe region”, all’interno della quale la pendenza della curva è molto bassa, tanto che nei grafici sembra
Figura 2.2: Curva carico – allungamento di un nervo tibiale di coniglio. Indicata con T la fine della “toe region” in cui la pendenza è praticamente nulla, con UL il carico di rottura, con UE l’allungamento massimo tollerabile prima del cedimento. L’area obreggiata indica l’energia assorbita durante l’allungamento (Grenwal R. et al., 1996).
quasi inesistente se confrontata con il resto dell’andamento. In questa zona a fronte di una piccola sollecitazione in forza si ottengono rilevanti allungamenti. Nella regione successiva si assiste ad un veloce mutamento del coefficiente angolare della curva, che da valori molto bassi assume, con un incremento in allungamento relativamente modesto, valori non trascurabili. Nella terza, e ultima, regione la curva mantiene una pendenza pressochè costante finchè non si arriva a valori di allungamento e quindi di forza che provocano il cedimento strutturale del tessuto nervoso (Grewal R. et al., 1996).
2.1.4 La curva tensione – deformazione
Ottenuta la curva forza allungamento, per passare alla curva tensione -deformazione c’è la necessità di misurare l’area della sezione trasversale del campione, sottoposto a trazione, per ricavarne una tensione media. In tutti gli esperimenti la misura è stata effettuata sull’area della sezione del campione nello stato di riposo. Tutte le esperienze sono state effettuate in vitro, quindi per definire lo stato di riposo, nel quale misurare la sezione laterale, si è assunto uno stato fittizio di riferimento. Questo è stato fatto coincidere di volta in volta, a seconda degli autori:
- con le condizioni nelle quali si veniva a trovare il nervo, vincolato, prima dell’estrazione dal corpo dell’animale. Il nervo, appena estratto, veniva congelato e la sua sezione trasversale misurata con un polarimetro planare (Bora F.W. Jr., et al., 1980).
- con le condizioni del nervo, reciso ed estratto, lasciato penzolare liberamente sulla macchina per alcuni minuti, sottoposto soltanto al proprio peso (Rydevik B.L. et al., 1990), (Kwan M.K. et al., 1992).
- con la sezione misurata al tempo t=0 del nervo montato fra le morse della macchina di misura (Walbeehm E.T., et al., 2004).
Stabilita così l’area della sezione trasversale del nervo, conoscendo la forza che la macchina di misura applica sul campione si è potuta estrarre una tensione media che messa in relazione alla deformazione, calcolata in percentuale rispetto alla lunghezza iniziale fra le morse, produce curve con l’andamento di Fig. 2.3 a e Fig. 2.3 b.
Figura 2.3 b: Curva tensione – deformazione di un nervo sciatico di ratto. Nel tratto a pendenza costante si può parlare di modulo di Young nel senso classico del termine (Borschel et al., 2003).
Figura 2.3 a: Curva tensione – deformazione di un nervo tibiale di coniglio (Kwan M.K. et al., 1992).
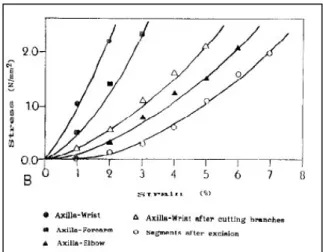
Una ulteriore esperienza ha dimostrato come la rigidezza del nervo dipenda dalla presenza di diramazioni lungo il suo percorso. Sono state misurate le caratteristiche meccaniche di nervi mediani (Fig. 2.4 A) e ulnari (Fig. 2.4 B) mediante una prova di trazione uniassiale. L’andamento delle curve tensione – deformazione si è dimostrato variabile in funzione della presenza o meno delle diramazioni. I nervi ancora muniti delle loro diramazioni secondarie si sono dimostrati più rigidi esibendo una curva nella quale una pendenza maggiore si associa alla mancanza della “toe region”. Quelli con le diramazioni recise invece, pur non presentando “toe region”, hanno rivelato una pendenza minore dei precedenti. Infine, gli stessi segmenti nervosi, una volta recisi, hanno mostrato la presenza di “toe region” e una pendenza ancora minore dei precedenti. Questo fenomeno è stato spiegato con la presenza in situ di tessuto connettivo che rende pìù difficile la distensione del nervo, conferendogli una resistenza maggiore di quella che riesce ad esibire in esperienze in vitro (Millesi H. et al., 1995).
Figura 2.4 A: Curva tensione – deformazione di un nervo mediano di cadavere. Le curve con pendenza minore sono quelle che descrivono il comportamento di nervi recisi o soggetti ad una poco rilevante influenza delle diramazioni nervose. Al contrario le curve più ripide sono relative alla descrizione del comportamento di nervi considerati nella loro interezza e soggetti all’influenza di diramazioni. (Millesi H. et al., 1995).
2.1.5 Le condizioni in situ
Una volta ottenuta la curva tensione – deformazione uno dei problemi che si sono presentati ai ricercatori è stato quello di determinare se all’interno di un organismo vivente i nervi si trovassero in tensione e a quali tipi di allungamenti essi fossero soggetti. Una interessante esperienza svolta su modello animale ha portato alla determinazione dell’allungamento al quale si trovano sottoposti alcuni nervi in situ. A questo scopo, fissati il ginocchio e l’anca di 18 conigli bianchi della Nuova Zelanda a 90° di flessione, sono stati esposti i nervi tibiali di ciascun animale. Si è poi misurata, tramite un video analizzatore, la distanza intercorrente fra due linee tracciate in situ, mediante macchia di Verhoff, sulla superficie del nervo. Successivamente i nervi sono stati estratti, mediante incisione, e lasciati liberi di assestarsi meccanicamente per un tempo di 5 minuti. In queste condizioni si è misurata la lunghezza di riposo e la sezione trasversale iniziale. Un confronto fra le distanze relative dei riferimenti in queste condizioni e quelle misurate in situ ha dimostrato la presenza di un allungamento del nervo tibiale dello 11±1,5% all’interno dell’organismo dei conigli, nelle condizioni sopra descritte e
Figura 2.4 B: Curva tensione – deformazione di un nervo ulnare di cadavere. Il comportamento è simile a quello del nervo mediano (Millesi H. et al., 1995).
considerate come standard in questi tipo di esperimento. Dalla curva tensione – deformazione poi si può ricavare la tensione in situ (Rydevik B.L. et al., 1990), (Kwan M.K. et al., 1992).
2.1.6 Gli allungamenti fisiologici
Per quanto riguarda la valutazione quantitativa degli spostamenti, a cui sono soggetti alcuni nervi periferici durante i normali movimenti corporei, studi effettuati su cadaveri hanno dimostrato che il nervo mediano, alla massima estensione della giunzione del gomito, è sottoposto, in media, ad un allungamento del 4,5% circa, destinato a subire un ulteriore incremento in caso di estensione del polso e delle dita. Il nervo mediano subisce quindi allungamenti e accorciamenti compresi in un intervallo che va dal 4% circa in trazione al 15% circa in compressione (Millesi H. et al., 1995). In un recente studio un gruppo di ricercatori ha correlato alcuni movimenti corporei con lo spostamento e la deformazione subita dal nervo mediano. I movimenti esaminati sono stati l’estensione del polso, l’adduzione della spalla, l’estensione dall’articolazione del gomito e la flessione controlaterale del collo. E’ stato esaminato un campione costituito da 23 donne e 11 uomini di età compresa fra i 20 e i 59 anni, sottoposti preventivamente a visita medica per accertare l’assenza di patologie spinali a loro carico. Per ognuno di essi sono state misurate la lunghezza del braccio e la distanza intercorrente fra il sesto processo cervicale e la punta del dito del medesimo braccio steso sul fianco, così da avere una buona approssimazione del percorso del nervo mediano. Il movimento del nervo è stato stimato mediante l’analisi e la comparazione, fornita da algoritmi numerici, di immagini provenienti da trasduttori ad ultrasuoni. In fig. 2.5 a e fig. 2-5 b sono visibili i movimenti del nervo mediano in relazione all’angolo assunto dall’articolazione del gomito per due diversi valori dell’angolo di adduzione della spalla. In fig. 2-6 a e fig. 2-6 b invece si possono apprezzare i movimenti del nervo mediano in funzione degli
differenza fra i movimenti del nervo in corrispondenza dei trasduttori e la distanza fra i trasduttori stessi. La deformazione complessiva del nervo mediano è influenzata dal comportamento di tutte le articolazioni che interessano il suo percorso. Finchè il nervo mediano appare scarico, in condizione di adduzione della spalla o di flessione dell’articolazione del gomito, la deformazione totale può essere determinata come la somma di quelle di tutte le articolazioni. Quindi in condizioni di adduzione della spalla di 90° e di estensione dell’articolazione del gomito di 40°, la deformazione complessiva a livello dell’avambraccio prossimale è del 2,1%. Una estensione dell’articolazione del gomito fino a 60° porta la questa deformazione a valori del 2,5-3%. In conclusione questo lavoro dimostra che l’uso funzionale del braccio nelle posizioni di adduzione e di estensione sono caratterizzate da avere l’articolazione del gomito piegata e la spalla in adduzione. In questa condizione il nervo mediano è praticamente scarico ed in corrispondenza delle articolazioni presenta una piccola deformazione. La situazione è differente se tutte le articolazioni sono distese, in questo caso si arriva ad una deformazione del 3-4%. (Dilley A. et al., 2003).
Figura 2.5 a: Movimenti del nervo mediano all’interno dell’avambraccio distale, medio e prossimale in funzione dell’angolo del polso misurato da un elettrogoniometro. L’escursione angolare va dai 60° di flessione ai 40° di estensione con 45° di abduzione della spalla (Dilley A. et al., 2003).
Figura 2.6 a-b: Movimenti del nervo mediano all’interno dell’avambraccio medio e prossimale in funzione dell’angolo del collo misurato da un elettrogoniometro. I movimenti del nervo seguono la
Figura 2.5 b: Movimenti del nervo mediano all’interno dell’avambraccio distale, medio e prossimale in funzione dell’angolo del polso misurato da un elettrogoniometro. L’escursione angolare va dai 60° di flessione ai 40° di estensione con 90° di abduzione della spalla (Dilley A. et al., 2003).
2.2 I fenomeni connessi alla viscoelasticità
Il tessuto nervoso possiede caratteristiche meccaniche comuni a molti tessuti biologici che, oltre a manifestare una certa elasticità nel breve periodo, sono soggetti a fenomeni di scorrimento viscoso con il passare del tempo. Questa tipologia di fenomeni influenza globalmente tutte le caratteristiche meccaniche del tessuto nervoso, rendendole dipendenti dal tempo, dalla velocità di applicazione del carico e dalla successione di carichi applicati. Appare quindi logico come anche le curve riportate nei paragrafi precedenti, relative ai valori di tensione – deformazione e di forza – allungamento siano da intendersi come relative ad un lasso di tempo abbastanza breve ed a una velocità di applicazione del carico ben definita. Generalmente i fenomeni viscosi che influenzano il comportamento dei materiali vengono studiati mediante prove di creep e di rilassamento.
2.2.1 Il fenomeno del creep
Un materiale sottoposto ad un carico applicato impulsivamente, e mantenuto di entità costante, risulta soggetto ad allungamento progressivo con il passsare del tempo. Questo fenomeno, denominato creep, è riscontrabile nei tessuti biologici, nei polimeri, nelle gomme ed in tutti quei materiali che risentono in qualche modo di effetti viscosi. Il creep risulta dipendente dal tempo, dal carico e dalle modalità della sua applicazione, dalla temperatura sia del campione che dell’ambiente in cui esso si trova. In caso di presenza di rilevanti gradienti termici, la temperatura ha un effetto di accelerazione del fenomeno, in condizioni fisiologiche però il tessuto nervoso si trova soggetto ad una escursione termica molto esigua, quindi in una condizione ben approssimabile come isoterma.
2.2.2 Il fenomeno del rilassamento
Le proprietà viscoelastiche dei nervi sono state diffusamente indagate mediante esperimenti di rilassamento, probabilmente perché questo fenomeno ha, contrariamente al creep, ripercussioni dirette su problematiche di carattere medico. Il fenomeno del rilassamento consiste in una progressiva diminuzione, con il passare del tempo, della tensione interna al campione sottoposto ad un allungamento applicato impulsivamente e mantenuto costante. Un vasto studio sul comportamento meccanico dei materiali biologici è stato svolto da Fung1. Egli ha rilevato che materiali costituiti prevalentemente da componenti elastici mostravano un esiguo rilassamento. Tessuti contenenti collagene, dimostravano un rilassamento moderato. Un marcato rilassamento, infine, era caratteristico di tessuti del tipo di quello muscolare. Uno fra i primi studi a dimostrare l’esistenza di rilassamento nei nervi, mediante l’applicazione di carichi ciclici, è attribuibile ad Hartung2. Altri ricercatori, fra cui Yamada3, hanno osservato un decremento nel tempo della tensione in nervi sciatici di coniglio mantenuti ad un allungamento pari al 6% della loro lunghezza iniziale. L’esistenza del fenomeno del rilassamento unitamente a quello di creep in nervi sciatici di ratto, è attribuibile alle ricerche di Kendall et al.4. I risultati di questo lavoro, dimostrano che la tensione, in nervi sottoposti ad allungamento impulsivo mantenuto costante nel tempo, decresce del 30% nei primi 10 minuti, mentre nei successivi 20 ha un decremento più lento, per poi stabilizzarsi nei minuti successivi (Grewal R. et al., 1996). Un lavoro più recente, svolto su modello animale, si è preposto la studio del fenomeno del rilassamento in vista di applicazioni alle operazioni di sutura di
1
Fung Y., ”Mechanical properties of living tissues” in Biomechanics. New York, Springer Verlag, 1985, p 187.
2
Hartung C., Arnold G.,”Histomechanische Eigenshaften Peripherer Nerven” Nervenarts 44:80, 1973.
3
Yamada Y., “Studies of electrophysiological and morphological changes in sciatic nerve of rabbit under various types if stretch and relaxation”. In Journal of Japanese Orthopaedic Association, 61:217, 1987.
4
nervi danneggiati. Sono stati studiati nervi tibiali di 24 conigli bianchi della Nuova Zelanda. Sulla superficie dei nervi, portati allo scoperto dopo l’uccisione delle cavie, sono stati creati, in situ, riferimenti ottici mediante colorazione chimica dovuta a macchia di Verhoeff. Questa procedura è stata attuata per creare uno stato iniziale catalogato in base alla distanza fra i riferimenti ottici, misurata da un video analizzatore. La porzione di tessuto nervoso da sottoporre allo studio è stata estratta e successivamente montata, racchiusa fra due pinze, su una macchina per l’analisi strutturale di materiali del tipo Instron (Instron Corp., Canton Mass.). Il campione è stato mantenuto in un bagno di soluzione salina termostata a 37° C. Ricondotti alla loro lunghezza iniziale, computata in base alla distanza fra i riferimenti ottici, i campioni, divisi casualmente in tre gruppi distinti, sono stati precondizionati mediante un ciclo di allungamento, al tasso di deformazione dello 0,03% al secondo, al fine di giungere ad un livello di deformazione complessiva del 6%, 9%, 12%, rispettivamente, al variare del gruppo di appartenenza. In una fase successiva, è stato permesso ai nervi di equilibrare il loro stato tensionale interno per il tempo di un’ora. Infine essi sono stati riallungati, al tasso di deformazione del 3% al secondo fino al livello raggiunto nel precondizionamento e lasciati in questa condizione per il tempo di un’ora. Il livello di tensione al variare del tempo è stato computato in base alla forza, data in uscita dalla macchina di misura, e alla sezione laterale, misurata nelle condizioni iniziali con l’ausilio di un calibro digitale. Il rilassamento è stato espresso come percentuale del valore iniziale di picco, utilizzando la cosiddetta funzione di rilassamento ridotta. Per la valutazione e la comparazione dei risultati provenienti dai tre gruppi è stato usato il testi t di Student. Il risultato di questa esperienza è mostrato in Fig. 2.7 (Wall E. et al., 1991).
2.2.3 Il fenomeno del “mushrooming”
Il fenomeno del “mushrooming” consiste nella lenta e progressiva fuoriuscita dell’interno della fibra nervosa dalla sezione di recisione. In passato si è cercato di correlare questo comportamento con la presenza della pressione endoneurale5. Ma ciò non sarebbe consistente con la dinamica del fenomeno che si evolve lentamente, per periodi variabili da alcuni minuti a ore, dal momento della recisione. La presenza della pressione endoneurale infatti provocherebbe una veloce fuoriuscita dell’interno della fibra. Una più moderna teoria tende a spiegare il fenomeno come una combinazione del ritorno elastico del tessuto perineurale, che perde la propira integrità dopo la recisione del nervo, e della fuoriuscita del tessuto endoneurale. Secondo questa teoria la fibra nervosa incrementa il proprio diametro a causa della pressione endoneurale rimasta e, di
5
Lundborg G., “Intraneural microcircolation” in Orthop. Clin. North Am 19:1-12, 1988 Figura 2.7: Curva di rilassamento di tre gruppi di nervi periferici di coniglio mantenuti ripettivamente al 6%, 9%, 12% di allungamento per il tempo di un ora . E’ mostrata la tensione media come percentuale del picco iniziale (funzione di rilassamento ridotta). Le barre rappresentano l’errore (Wall E. et al., 1991).
conseguenza, diminuisce la propria lunghezza6. Per verificare l’ipotesi che il “mushrooming” fosse provocato in primo luogo dal comportamento del tessuto endoneurale sono stati fatti esperimenti tesi alla determinazione della dinamica dell’assorbimento di liquidi da parte della fibra nervosa e alla quantificazione del ritiro del tessuto nervoso dopo la recisione. Questo per poter distinguere i due diversi meccanismi, quello di accorciamento del perineuro e quello del rigonfiamento e trabocco dell’endoneuro, supposto quindi dovuto ad assorbimento di liquidi. Il risultato di questo lavoro, mostrato in fig. 2.8, è una curva che rappresenta rispettivamente l’aumento di peso di un nervo intero e del solo interno in funzione del tempo. Secondo gli stessi autori questo andamento non rispecchia l’andamento del fenomeno del “mushrooming” nel tempo, ma rende conto dell’esistenza all’interno del nervo integro di tessuti non completamente idratati e meccanicamente vincolati dalla presenza dello strato esterno.
6
Ushiki T., Ide C., “Three-dimensional organization of the collagen fibrils in the rat sciatic nerve as revelated by transmission and scanning electron microscopy” in Cell Tissue Res 260:175-184
Figura 2.8: Rigonfiamento di un nervo nella sua interezza e del solo suo interno in acqua distillata. La curva, che rappresenta l’aumento di peso in funzione del tempo, dimostra la tendenza all’assorbimento del liquido da parte di tessuti interni al nervo che non sono ben idratati (Walbeehm E. T. et al., 2004).
Le esperienze sulla ritrazione degli strati esterni del nervo dopo la sua recisione hanno avuto lo scopo di quantificare il fenomeno e, dal punto di vista concettuale, di comprendere se questo fosse dovuta alle sole caratteristiche del materiale o fosse in qualche modo influenzato anche da fattori cellulari. L’esperienza è stata condotta su due gruppi di tessuti nervosi, il primo mantenuto in un incubatore per 24 ore, il secondo composto da tessuti mantenuti congelati per 24 ore e poi scongelati. Il fenomeno della ritrazione non ha mostrato sostanziali differenze nei due casi, e quindi è stata esclusa l’influenza a livello cellulare, sulla dinamica del fenomeno. I risultati sono mostrati in tab. 2.1 e tab. 2.2 (Walbeehm E.T., et al., 2004).
Tabella 2.1: Ritrazione immediata dopo il taglio misurata in tutti i 18 nervi usati nell’esperimento. In 9 è stato misurato anche un aumento della sezione (Walbeehm E. T. et al., 2004).
Tabella 2.2: Ritrazione immediata dopo il taglio misurata in 8 nervi dei quali 4 sono stati mantenuti in un incubatore a 37°C, gli altri congelati e scongelati successivamente. Non ci sono differenze apprezzabili nei due casi (Walbeehm E. T. et al., 2004).
2.2.4 Il fenomeno dell’isteresi
Le caratteristiche viscoelastiche del tessuto nervoso influenzano anche le curve forza – allungamento, tensione – deformazione, e quelle da esse derivate come la curva carico- deformazione. Infatti un materiale, che non presenti al suo interno fenomeni viscosi, produce una curva che, nel piano delle forze e degli allungamenti, rappresenta una funzione monodroma. Inoltre questa curva, funzione soltanto dalla combinazione dei carichi e degli allungamenti, viene percorsa in un verso ascendente nella fase di carico e discendente durante quella di scarico. Questo si ripete per tutti i cicli di carico – scarico che non portino a rottura il materiale. Nel caso di un materiale viscoelastico invece il punto che rappresenta lo stato nel piano tensione -deformazione, dati gli scorrimenti interni che avvengono durante il processo di allungamento percorrerà la traiettoria della fase di scarico lungo una curva differente da quella della fase di carico precedente. Questa è solitamente più bassa in riferimento alle tensioni e spostata a destra sull’asse delle deformazioni. Ciò rende evidentemente conto dei fenomeni di rilassamento e di creep che si sviluppano all’interno della fibra.
2.3 Effetti delle sollecitazioni meccaniche sulla fisiologia dei nervi periferici
Le sollecitazioni meccaniche esterne possono avere delle ripercussioni anche gravi sulla fisiologia del tessuto nervoso periferico. I danni possono essere di carattere strutturale, connessi alla circolazione interna alla fibra nervosa oppure relativi alla capacità di quest’ultima di condurre gli stimoli elettrici in modo corretto. Il grado di deterioramento funzionale dipende sia dalla grandezza, dal carattere e dalla durata di applicazione della sollecitazione deformante, che dall’anatomia della fibra nervosa ad essa sottoposta. Le proprietà meccaniche del tessuto nervoso esposte nelle sezioni precedenti tendono a salvaguardare
l’integrità strutturale delle fibre ed il loro corretto funzionamento (Grewal R. et al., 1996).
2.3.1 Gli effetti dello stiramento sulla permeabilità epineurale
Il perineuro fornisce una barriera alla diffusione di sostanze chimiche, fra le quali per importanza spiccano le proteine. Contribuisce così, in maniera specifica, alla conservazione della composizione dell’ambiente intrafascicolare. Danni al perineuro comportano un cambiamento della sua permeabilità, che si ripercuote in un impoverimento della concentrazione di proteine dell’ambiente intrafascicolare. A questo impoverimento segue edema intrafascicolare, che sfocia in processi fibrotici, che compromettono la possibilità di recuperare una normale funzionalità da parte del nervo. Nonostante risulti evidente l’importanza di poter stimare con sufficiente precisione il limite oltre il quale lo stiramento produce danni irreversibili alla fibra nervosa, questo limite non è stato ben quantificato (Grewal R. et al., 1996).
2.3.2 Effetti dell’allungamento sulla microcircolazione interna
Il tessuto nervoso presenta un elevata complessità dovuta alla contemporanea presenza di diversi tipi di strutture cellulari e tessuti che conferiscono, nella loro globalità, alla fibra nervosa le caratteristiche proprietà funzionali. Un aspetto fondamentale per la conservazione delle suddette funzionalità, è la possibilità da parte del sistema microcircolatorio interno di portare il nutrimento alla totalità dei tessuti presenti nel nervo. In caso di allungamento si verifica il fenomeno della contrazione laterale, che diminuendo la sezione provoca lo schiacciamento del tessuto. A questo segue la compressione dei vasi sanguigni che lo irrorano e lo nutrono. Ben si comprende, quindi, come schiacciamneti troppo elevati possano avere effetti pesanti sulla microcircolazione interna. Studi effetuati e riportati in (Rydevik et al, 1989) e (Millesi et al., 1995), riportano per un nervo tibiale di
si ha il totale blocco del flusso sanguigno dei capillari. Altre esperienze su modello animale dimostrano che all’8% di allungamento si susseguono, riduzione del flusso sanguigno del 50%, lesioni vascolari e in alcuni casi anche inizio di necrosi del tessuto. Recenti ricerche svolte sul plesso brachiale di coniglio (Takai et al. 2002), confermano che in corrispondenza dell’8% di elongazione si hanno danni al tessuto ed alla circolazione interna. Per quanto riguarda l’essere umano questo limite è già superiore a quello del 6% in cui si va incontro a danneggiamento strutturale (Millesi et al., 1995). Per quel che riguarda i livelli di compressione a cui si verificano fenomeni degenerativi si è dimostrato (Dahlin et al. ,1986) che una pressione interna al nervo di 80 mmHg, mantenuta per il tempo di 2 ore provoca la decrescita graduale della velocità di conduzione delle fibre e la diminuzione del potenziale d’azione, unita ad una completa ishemia del tessuto. Ad una pressione di 400 mmHg, mantenuta per 15 minuti, tutti i suddetti fenomeni degenerativi si velocizzano e comprendono la maggioranza dei tessuti interni.
2.4 Conclusioni e considerazioni
In base alla conoscenza acquisita negli anni sulle proprietà biomeccaniche del tessuto nervoso periferico si è cercato di stabilire, integrando i molteplici contributi, i limiti di sollecitazione che ne mantengano l’integrità ed il trofismo fisiologico. Si è quindi giunti alla conclusione che il limite di allungamento a cui è possibile sottoporre il tessuto nervoso vanno dal 4% al 6% e che la pressione interno dovuta a schiacciamento non deve in alcun modo superare gli 80 mmHg. Per quanto riguarda lo studio delle proprietà meccaniche del tessuto, sottoposto ad allungamento su macchine di prova per materiali, si sono riscontrate lacune nella trattazione del problema. In effetti non è corretto misurare gli estremi del nervo collegato alla macchina, per avere una misura affidabile dell’allungamento. Difatti le estremità schiacciate influenzano negativamente la misura in un modo non
condizioni riportate in letteratura si misura un comportamento “medio”, di un tessuto in parte schiacciato ed in parte no. Nell’indicazione dei risultati delle prove di trazione, eccettuato (Arnold G. e Lutge H., 1973), manca completamente la curva di scarico del materiale. Nessun accenno preciso viene inoltre fatto, tranne in alcuni casi, a procedure di precondizionamento del tessuto che tendano a diminuire l’ampia isteresi che si incontra nelle fasi iniziali della prova. Quando poi questo importante aspetto viene citato non si adotta un protocollo standard. Nessuno ha infine proposto un qualche modello quantitativo, che possa decrivere con una certa precisione i comportamenti del tessuto nervoso periferico sottoposto a sollecitazioni esterne.