3.1 Il progetto
Questo studio è stato realizzato nell’ambito di un progetto finanziato dal Ministero delle Politiche Agricole, Alimentari e Forestali, nel quadro del VI Piano Triennale della Pesca e dell’Acquacoltura (codice progetto (6-B-1)) dal titolo: ”Studio di fattibilità dell’utilizzo di attrezzi selettivi per la cattura di crostacei economicamente importanti nell’ambito di una riconversione dei mestieri e di una riduzione dell’impatto sugli ecosistemi” (De Ranieri, 2005). Tale studio si prefiggeva di valutare, dal punto di vista sperimentale, l’efficacia e la sostenibilità dell’utilizzo di nasse per la cattura di gamberi (Plesionika edwardsii) e scampi (Nephrops norvegicus) nel piano batiale, in alternativa alla pesca a strascico, e di nasse per la cattura delle mazzancolle (Penaeus kerathurus) lungo la fascia costiera toscana, in alternativa alle reti da posta.
Il progetto si è basato sulla collaborazione dell’Ente di Ricerca proponente (C.I.B.M. di Livorno) con altre strutture di Ricerca (A.R.P.A.T. di Livorno, Aplysia di Livorno) e con strutture del settore cooperativo (Ce.S.I.T. di Castiglione della Pescaia) legate al mondo della produzione, coinvolgendo anche gli operatori del settore (associazioni della pesca e pescatori stessi) in tutte le fasi di realizzazione. La raccolta dati si è concentrata sui campionamenti mirati alla cattura di P. edwardsii. In accordo con lo schema del progetto, i dati necessari per questo studio sono stati ottenuti sia da pescate realizzate con nasse sperimentali sia da campionamenti su
crostacei batiali. I dati sono stati raccolti nel periodo 2003 – 2006, in un tratto di mare a Sud dell’Isola di Giannutri, nel Mar Tirreno Settentrionale.
Tali attività rientravano negli obiettivi del progetto: valutare l’efficacia di nasse sperimentali per la cattura di P. edwardsii e monitorare l’attività di pesca usualmente praticata per lo sfruttamento di questa specie.
3.2 Pescate con nasse sperimentali.
Al fine di scegliere la tipologia di nassa più adeguata alla pesca sperimentale di P. edwardsii, sono state effettuate indagini bibliografiche, visite in banchina ed interviste con i pescatori, non solo dei porti toscani, ma anche di altre località marittime italiane. Dato che in Italia l’utilizzo di nasse per la pesca di questa specie è risultato irrilevante, questa indagine ha tenuto conto anche di conoscenze provenienti da altri paesi mediterranei.
Le nasse utilizzate sono state costruite (Fig. 3.2.1) prendendo come riferimento quelle attualmente in uso presso alcune marinerie spagnole (es. Isole Baleari), mirate essenzialmente alla pesca dei gamberi (Gestin e Guennégan, 1989).
Fig. 3.2.1 – Assemblaggio di una nassa per la pesca batiale di P. edwardsii.
1999). Si tratta di nasse che, grazie alla presenza di galleggianti di profondità, vengono mantenute sospese a circa un metro di dal fondo; questa caratteristica le rende particolarmente indicate per la pesca di organismi necto-bentonici, quali il pandalide P. edwardsii.
Le nasse hanno una forma tronco-conica (Fig. 3.2.2) e sono costituite da una intelaiatura in filo di ferro zincato di 4 mm di diametro, circondata da una rete in plastica bianca (con maglie quadrate di 10 mm di lato). La struttura di base è un cilindro di 60 cm di diametro e 30 cm di altezza, al quale sono assemblati due coni di 20 cm di altezza, uno nella parte superiore ed uno in quella inferiore, con aperture al centro. L’apertura nella parte superiore, del diametro di 15 cm, è stata chiusa giustapponendo e legando un coperchio fatto in rete in plastica, da aprire per le operazioni di ispezione e “manutenzione” (per raccogliere gli eventuali individui catturati, per collocare e rimuovere l’esca, ecc.); l’apertura nella parte inferiore, di 8 cm di diametro, costituisce l’entrata per le prede richiamate dalla presenza dell’esca.
L’esca, costituita da sacchetti di rete biodegradabile contenenti sardine, è stata posizionata all’interno di ogni nassa, legata al bordo superiore del cilindro, in modo da rimanere, quando in pesca, al centro della nassa stessa (Fig. 3.2.3.).
Fig. 3.2.2 – Caratteristiche costruttive e dimensioni delle nasse utilizzate per la pesca di P. edwardsii nel piano batiale.
Galleggianti di profondità Moschettone Apertura con coperchio Inganno 30 cm 50 cm 20 cm 15 cm 30 cm 50 cm 20 cm 15 cm 8 cm 60 cm cm 8 cm 60 cm cm
Fig. 3.2.3 – Sacchetto con sardine salate utilizzato per innescare le nasse per la pesca di P. edwardsii.
Ad ogni nassa, in prossimità del suo bordo superiore, sono stati fissati due galleggianti di profondità in plastica da 0,5 litri; dalla parte opposta è stato collegato un “bracciolo” in cordino, di 3 mm di diametro e lungo circa 80 cm, munito di un moschettone per lo sgancio rapido, con la funzione di collegare la nassa al trave o “madre” del calo di nasse. Grazie ai moschettoni, le nasse sono in grado di ruotare su loro stesse e di disporsi nel senso della corrente.
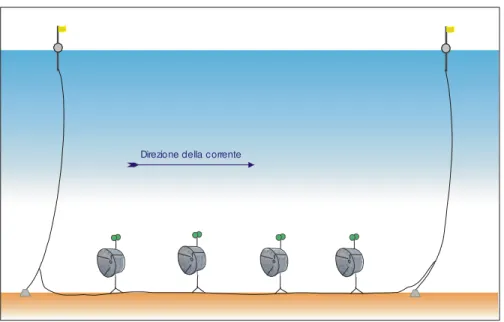
Come trave è stata utilizzata una corda piombata del peso di 40 kg per ogni 100 m con la funzione di mantenere in posizione tutto il calo, limitandone gli spostamenti dovuti alla forza delle correnti. In questo modo le nasse, una volta calate, si posizionano sul fondale (grazie al peso del trave piombato), ma si dispongono sollevate dal fondo ad un’altezza pari a quella del bracciolo, grazie alla spinta dei galleggianti (Fig. 3.2.4).
Fig. 3.2.4 - Schema del posizionamento in pesca di un calo di nasse utilizzato nel piano batiale per la cattura dei gobbetti.
Le nasse sono state posizionate sul trave ad una distanza di 10 m l’una dall’altra; in ogni uscita sperimentale sono stati utilizzati cali di 30 nasse. Alle due estremità del trave, sono stati lasciati due segmenti di 50 cm, privi di nasse, e collegate a due strutture in cemento di 20 kg, collegati a loro volta, tramite una cima, o “caloma”, alla superficie con una boa di segnalazione, un pallone in plastica. La boa è stata munita di un segnale radarabile metallico, in modo da facilitarne le operazioni di avvistamento e recupero.
Le pescate con nasse sperimentali sono state effettuate in un’area con fondali fangosi, tra 294 e 447 metri di profondità media, a più di 20 miglia dal porto di Porto Ercole, caratterizzata da fondali fangosi con ripide pendenze, le cosiddette “bucate” (Fig. 3.2.5).
Fig. 3.2.5 - Localizzazione dei siti in cui sono state calate le nasse sperimentali a gobbetti (P. edwardsii) nell’area di Porto Ercole, nei tre anni di studio.
La sperimentazione delle nasse è stata effettuata utilizzando un peschereccio a strascico appartenente alla marineria di Porto Ercole (Tab. 3.2.1), abitualmente impiegato per la pesca con reti a strascico.
Tab. 3.2.1 – Caratteristiche tecniche della imbarcazione utilizzata per le pescate sperimentali con nasse batiali.
Marineria Porto Ercole Imbarcazione Angela Madre Stazza (TSL, t) 50
Potenza (HPA) 280 Lunghezza (LFT, m) 22,7 Verricello salpareti No Verricello strascico Sì
Per quanto riguarda i tempi di pesca, gli attrezzi hanno pescato per un periodo di circa 24 ore; questo si è rilevato il tempo migliore, consentendo il migliore compromesso tra aspetti logistici e quantità della cattura.
Il tempo necessario per calare le nasse è stato di circa mezz’ora mentre il loro recupero, avvenuto un’ora dopo l’alba del giorno successivo, ha richiesto in media un’ora. Non sempre è stato possibile fare le uscite in mare, a causa della lontananza della zona di pesca e delle condizioni metereologiche sfavorevoli.
Al termine di ogni pescata, tutte le eventuali catture (Fig. 3.2.6) di ogni nassa sono state disposte in sacchetti separati e contrassegnati con il numero della nassa. Tutto il materiale è stato congelato e trasferito in laboratorio per le successive analisi.
Fig. 3.2.6 – Esemplari di P. edwardsii catturati da una nassa durante un’uscita sperimentale.
3.3 Osservazioni a bordo
Altro scopo del progetto nel quale è inserito questo lavoro di tesi era quello di monitorare la pesca professionale, al fine di conoscere la composizione in taglie dei crostacei catturati e, per ottenere stime della eventuale frazione scartata. Sono state effettuate, nei tre anni di indagine, giornate di osservazioni a bordo su pescherecci che effettuano normalmente la pesca a strascico a crostacei batiali. Sugli esemplari catturati di P. edwardsii, per ogni cala rilevata, sono stati acquisiti dati su numero e peso, sulla struttura demografica e sulla biologia riproduttiva. I rilevamenti sono stati realizzati a bordo di un peschereccio, appartenente alla marineria di Porto Ercole, la stessa imbarcazione utilizzata per le uscite con nasse sperimentali.
La pesca a strascico è l’attività più diffusamente praticata a livello semi-industriale ed semi-industriale per lo sfruttamento delle risorse ittiche. La rete a strascico è l’attrezzo più efficiente per catturare le specie demersali o nectobentoniche, ovvero quelle specie che contraggono rapporti sistematici con il fondo, trascorrendo gran parte del loro ciclo vitale nello strato della colonna d’acqua prossimo al substrato. Tuttavia questo attrezzo può catturare anche specie tipicamente pelagiche (quando esse si trovano in prossimità del fondo) e specie tipicamente bentoniche (in particolare quando non si trovano infossate nel sedimento). (Fig. 3.3.1)
Fig. 3.3.1 - Raggruppamenti ecologici degli organismi interessati dalla pesca a strascico: A) specie pelagiche gregarie, B) specie demersali, C) specie bentoniche che possono infossarsi nel sedimento, D) specie bentoniche che conducono parte della loro vita in tane scavate nel sedimento. In figura sono mostrate anche specie non catturabili dalla rete a strascico (da Froglia, 1982).
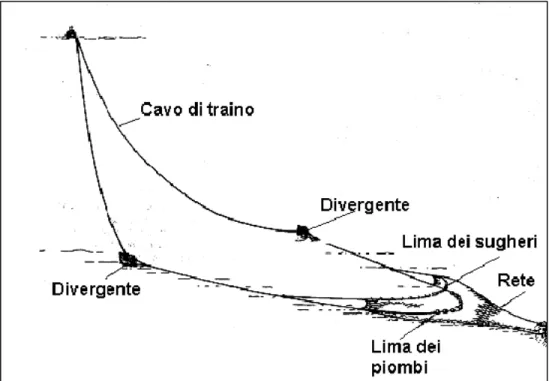
La rete a strascico è un attrezzo da pesca costituito da molte pezze di rete (Fig. 3.3.2), diverse per le dimensioni del filato e della maglia, cucite tra loro in modo da formare durante il traino un tronco di cono; gli organismi vengono catturati dal progressivo avanzamento della rete. Nella parte anteriore della rete si trova un’apertura, detta bocca, alle cui estremità sono fissate le mazzette, atte a collegare la rete all’imbarcazione attraverso due cavi, detti calamenti. La parte posteriore della rete, detta sacco, ha le maglie più piccole, per evitare che gli organismi di piccola taglia possano fuggire, e possiede un sistema di chiusura costituito da una cima annodata, per il recupero della pescata. Superiormente si trova la lima dei sugheri che garantisce l’apertura verticale della rete, mentre inferiormente è presente una lima piombata che consente il posizionamento della rete sul fondo.
Fig. 3.3.2 - Schema di rete a strascico e suoi componenti.
I calamenti sono collegati a due strutture, di solito in ferro o in acciaio, dette divergenti, aventi la funzione di stabilizzare la rete sul fondo e di mantenerne l’apertura orizzontale. Dai divergenti partono due cavi in acciaio, che sono collegati al verricello dell’imbarcazione. La rete adottata dall’imbarcazione su cui sono state fatte le osservazioni, era del tipo tradizionale (o “italiana”), ovvero con apertura verticale della bocca è di circa 1 m. la maglie al sacco misuravano 40 mm di lato, la misura consentita dalla legge (Regolamento UE 1626/94).
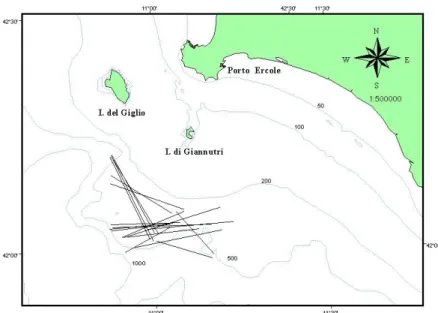
Le osservazioni a bordo sono state effettuate durante normali battute di pesca mirate ai crostacei batiali, effettuate a sud dell’Isola di Giannutri; la Fig. 3.3.3. mostra il tracciato delle cale monitorate. Generalmente, in ciascuna giornata di pesca sono state effettuate tre cale della durata di un’ora ciascuna, ad una velocità di traino è circa 3 nodi. Non c’è stata
nessuna interferenza, da parte del personale scientifico, con il modus operandi dell’equipaggio (scelta della zona di pesca, durata della cala e modalità smistamento del pescato). Tutta la cattura di P. edwardsii è stata congelata direttamente a bordo, per le successive analisi di laboratorio.
Fig. 3.3.3 - Tracciato delle cale commerciali su fondali batiali monitorate durante questo studio.
3.4 - Analisi di laboratorio
I principali aspetti indagati su P. edwarsii in questo studio hanno riguardato la demografia, la definizione del periodo riproduttivo e della taglia di maturità sessuale, la crescita relativa e stime di fecondità (assoluta e relativa). Sono stati prelevati campioni mensili, da dicembre 2003 a aprile 2006 per un totale di 4919 individui di P. edwardsii, di cui 1451 maschi, 2385 femmine e 1083 indeterminati (Tab. 3.4.1).
Tab. 3.4.1 - Numero di individui di P. edwardsii analizzati per mese (con nasse sperimentali o con rete a strascico), suddivisi per maschi, femmine e
indeterminati).
Anno Mese Attrezzo Maschi Femmine Indet. TOT
2003 dicembre nasse 56 70 8 134 2004 Febbraio nasse 23 48 7 78 2004 Aprile nasse 3 4 0 7 2004 Maggio nasse 113 174 103 390 2004 Giugno nasse 24 35 38 97 2004 Luglio nasse 175 193 326 694 2004 Ottobre nasse 153 258 357 768 2005 Marzo strascico 48 108 0 156 2005 Maggio nasse 68 266 218 552 2005 Giugno nasse 29 22 0 51 2005 Luglio nasse 195 292 3 490 2005 Agosto strascico 77 122 0 199 2005 settembre strascico 44 108 20 172 2005 novembre strascico 125 185 3 313 2006 Febbraio strascico 109 197 0 306 2006 Marzo strascico 107 177 0 284 2006 Aprile strascico 102 126 0 228 totale 1451 2385 1083 4919
3.4.1 Rilevamento parametri biologici
- Sesso, stadio maturativi, taglia e peso.
Gli esemplari catturati sono stati distinti per sesso, sulla base della presenza dei caratteri sessuali secondari: nel maschio sono presenti due appendici sessuali, una delle quali è definita “appendice mascolina” (King e Moffitt, 1984); questa si trova a livello dell’endopodite del secondo paio di pleopodi (Fig. 3.4.1.1 e Fig. 3.4.1.2); inoltre il maschio, quando è maturo, evidenzia sotto il carapace, un canale di colore bianco - trasparente con funzione di trasporto degli spermi. La femmina si distingue per la presenza dell’ovario a livello del cefalotorace che assume colorazioni diverse in base allo stadio maturativo (Fig. 3.4.1.3) e per la presenza di uova sotto l’addome. Il metodo migliore per la determinazione resta quello che si basa sulla presenza dell’appendice mascolina. Il riconoscimento non è molto semplice, poiché l’espansione è molto piccola e, nel caso di maschi non adulti, spesso poco visibile (poco sviluppata) o non ancora presente (King e Moffitt, 1984); l’identificazione è stata effettuata stringendo il pleopode alla base dell’endopodite con l’ausilio di pinzette da laboratorio, in modo da osservare la presenza di una o due espansioni.
Poiché il lavoro era focalizzato sullo studio della biologia riproduttiva e fecondità di P. edwardsii, sulle femmine sono stati assegnati gli stadi maturativi della gonade e sono stati identificati gli esemplari con uova trattenute dai pleopodi addominali (femmine ovigere); infine è stato assegnato lo stadio di sviluppo delle uova.
Fig. 3.4.1.1 – Immagine delle due appendici sessuali presenti sul secondo pleopode di un maschio di P. edwardsii.
Fig. 3.4.1.2 – Dettagli del secondo pleopode di una femmina e di un maschio di P. edwardsii a confronto. La freccia indica l’appendice mascolina.
Fig. 3.4.1.3 - Immagine di una femmina con ovario maturo (colore blu) e di un maschio di P. edwardsii.
Le femmine sono state classificate secondo tre stadi maturativi della gonade, a partire dall’esame macroscopico degli ovari (dimensioni e colorazione):
1. Femmina con ovario ridotto, di colore chiaro, tra il giallo e il rosa: individui immaturi (Fig. 3.4.1.4).
2. Femmina con ovario sviluppato, di colore celeste - violaceo. E’ lo stadio più difficile da individuare, perché si trova in una fase intermedia: individui in maturazione (Fig. 3.4.1.5).
3. Femmina con ovario ben sviluppato e di colore blu intenso: individui maturi (Fig. 3.4 1.6).
♀
Fig. 3.4.1.4 - Femmina di P. edwardsii con ovario in stadio 1.
Fig. 3.4.1.5 - Femmina di P. edwardsii con ovario in stadio 2.
Le femmine, sulla base della presenza o assenza di uova sui pleopodi, sono state distinte in ovigere e non ovigere. Le femmine ovigere, a loro volta, sono state distinte in:
- Femmine ovigere stadio 1: uova con assenza di embrioni e di colore azzurro intenso, definite uova “senza occhi” poiché non sono ancora presenti gli abbozzi oculari delle future larve (Fig. 3.4.1.7).
- Femmine ovigere stadio 2: uova con presenza di embrioni e di colore marrone chiaro, definite uova “con occhi” poiché sono presenti gli abbozzi oculari delle future larve (Fig. 3.4.1.8).
Le prime osservazioni sugli stadi di uova e ovari sono state effettuate a livello macroscopico, mentre lo studio approfondito dell’intero sviluppo di uova e gonadi è stato fatto con l’ausilio di uno stereomicroscopio.
Fig. 3.4.1.7 - Femmine di P. edwardsii con uova senza embrioni, di colore azzurro intenso (“senza occhi”); si tratta di uova stadio 1.
In Tab .3.4.1.1 è riportato il modulo utilizzato per il rilevamento dei parametri biologici.
Una volta suddiviso il campione in maschi e femmine e suddivise queste per stadio maturativo, su ogni esemplare è stata rilevata la taglia espressa come lunghezza del carapace (LC), cioè la lunghezza che va dall’orbita dell’occhio fino all’estremità posteriore del cefalotorace (Fig. 3.4.1.9). Questa misurazione è stata rilevata al millimetro inferiore, impiegando un calibro di precisione.
Fig. 3.4.1.9 – Rappresentazione della rilevazione della lunghezza del carapace (LC) in un crostaceo (da Sparre et al., 1989, modificato).
Successivamente, su un campione, è stato rilavato il peso individuale, espresso al centesimo di grammo.
Si è anche osservata la presenza/assenza di parassiti, di solito situati nelle branchie. Questi si vedono bene, perché causano una ciste/rigonfiamento sulla parte laterale del carapace. Altre cisti possono formarsi sulle uova sotto forma di filamenti biancastri.
Durante le analisi di laboratorio, è stata anche presa nota della presenza o assenza di individui in fase di muta.
Tab. 3.4.1.1 - Foglio di lavorazione per l’acquisizione dei dati biologici.
PROGETTO NASSE (C.I.B.M. – A.R.P.A.T. – Ce.S.I.T.) BIOLOGIA CROSTACEI DATA____________ OPERATORI______________ SPECIE__________________________ CALA_______________ N.TOT_____________PTOT____________SUBCAMPIONE____________ N° nassa LC (mm) P.TOT (0,1 g) Sesso Stadio maturativo femmine
Muta Ovigera Parassiti 1 2 3 4 5 6 7 8 9 10 11 12 13 14 15 16 17 18 19 20 21 22 23 24 25 26 27 28 29 30 31 32 33
- Studio della fecondità
La fecondità è un parametro biologico molto importante che può essere usato per distinguere le varie popolazioni di una specie. Può cambiare a seconda dell’area presa in esame (Ihssen et al., 1981). La fecondità può essere anche definita come un volano biologico che agisce da regolatore assicurando la sopravvivenza delle popolazioni naturali nel loro territorio, mantenendo le dimensioni della stessa intorno ad un valore medio che può essere considerato la capacità portante in quel particolare ambiente. Questo permette alle specie e alle popolazioni di ottimizzare le loro opportunità di sopravvivenza e definire il loro livello di dominanza nell’ecosistema (Ehrhardt, 1986).
Lo studio della fecondità è importante per approfondire i meccanismi di riproduzione della specie, dando informazioni sulla prolificità e sul tipo di strategia riproduttiva e offre la possibilità di individuare la porzione di femmine riproduttrici che contribuisce in modo maggiore al rinnovamento della popolazione (stock riproduttivo). La conoscenza della fecondità di una specie o di una popolazione è fondamentale nella corretta gestione degli stock.
Negli studi di biologia riproduttiva di solito viene stimata la fecondità assoluta e relativa. Quella assoluta è rappresentata dal numero di ovociti presenti nell’ovario appena prima della deposizione o rilascio. La fecondità relativa è definita come il numero di uova in funzione della taglia o del peso individuale.
Lo stima della fecondità può avvenire secondo vari metodi, come il metodo gravimetrico, il metodo volumetrico o tramite il conteggio diretto di tutte le uova contenute nell’ovario.
Nel metodo gravimetrico (Caillet et al., 1986; Holden e Raitt, 1974), le uova liberate dal tessuto ovarico, dopo il soggiorno in un adeguato conservante, vengono lavate ed essiccate. Dopo essiccatura, il quantitativo totale di uova viene pesato e da questo vengono estratti uno o più subcampioni di peso prederminato. Ciascun subcampione viene poi sottoposto alle procedure di conteggio.
Nel metodo volumetrico la soluzione contenente gli ovociti ed i residui di tessuto ovarico viene filtrata al fine di pulire il più possibile gli ovociti. In seguito le uova vengono messe in un contenitore graduato riempito con acqua o alcool fino ad un volume noto ed agitate manualmente o con un agitatore magnetico. Immediatamente dopo o durante l’agitazione si preleva con una pipetta un subcampione di volume noto di cui si conteranno gli ovociti. Questi primi due metodi sono utilizzati soprattutto per specie che possiedono ovari molto grandi con molte uova, come nel caso dei pesci.
La stima della fecondità di P. edwardsii è avvenuta tramite il conteggio delle uova, poiché il numero di uova per individuo non è molto alto. L’analisi della fecondità è iniziata con il prelievo delle uova, effettuato in modo molto accurato, con l’ausilio di pinzette da laboratorio. Si è cercato di prelevare tutte le uova, eliminando anche quelle trattenute dai pleopodi (Fig. 3.4.1.10).
Fig. 3.4.1.10 – Foto effettuata al microscopio ottico di parte di uova di una femmina di P. edwardsii appena prelevate dall’addome. La doppia feccia indica la
lunghezza del diametro maggiore di un uovo.
Le uova sono state poi trasferite in provette contenente formalina al 5% per il fissaggio. Dopo circa una settimana, i campioni di P. edwardsii sono stati agitati per circa 10 minuti con un agitatore meccanico, lavati con acqua in un setaccio da 100 µm e trasferiti in alcool al 70 % per la conservazione.
Il conteggio delle uova è avvenuto disponendole su una cella Dollfus (Fig. 3.4.1.11), una vaschetta di vetro rettangolare, di dimensioni interne 5X10 cm per 1 cm di altezza circa. La cella è divisa in 200 cellette, disposte in 20 colonne per 10 righe, ognuna di 0,5 cm di lato, separate le una dalle altre da piccole pareti alte 1 mm.
Fig. 3.4.1.12 - Immagine di una cella Dollfus vuota.
Una volta riversato il contenuto di una provetta nella cella, questo si sedimenta e può essere contato. Si è cercato di distribuire le uova più omogeneamente possibile con le pinzette o con uno palettina adatta all’interno della cella Dollfus. Durante il conteggio, le pareti fra una celletta e l’altra impediscono lo spostamento del materiale fra queste, eliminando così gli errori del conteggio. Il grado di precisione di questo metodo dipende prima di tutto da come viene prelevato il campione, dalla cura con cui sono mescolate le uova, da come viene effettuato il conteggio e dalle modalità di distribuzione degli ovociti nelle cellette. Per stabilire il numero di cellette minimo da contare, per avere una stima rappresentativa della fecondità reale, sono state fatte molte prove e si è visto che le stime ottenute contando 25 cellette (1/8 del totale) differivano dai conteggi della totalità delle cellette al massimo di ± il 10%. Si è ritenuto questo errore accettabile; le 25 cellette da contare sono state scelte sorteggiandole di volta in volta con la tavola dei numeri
casuali. Le cellette da contare sono state individuate sulla Dollfus con l’ausilio di pallini di piombo dal diametro molto piccolo (Fig. 3.4.1.12).
Fig. 3.4.1.12 - Immagine di una cella Dollfuss contenente le uova in stadio 1 di un esemplare di P.edwardsii. I pallini indicano le cellette prescelte per il
conteggio delle uova.
I conteggi delle uova sono stati effettuati su campioni di ciascun mese: sono state scelte 24 femmine ovigere di diversa taglia con uova nei due stadi di maturazione (con occhi e senza occhi) per mese. Quando possibile, i 24 individui sono stati scelti fra 4 classi di taglia (15-18 mm LC, 19-22 mm LC, 23-26 mm LC, 27-30 mm LC).
Successivamente si è proceduto con la misurazione delle uova: è stato misurato il diametro maggiore delle uova con lo scopo di osservare lo sviluppo dell’uovo e dell’embrione, fino alla schiusa della larva. Le uova di P. edwardsii, nelle prime fasi dello sviluppo sono sferiche, poi tendono a svilupparsi maggiormente secondo un asse, diventando ellissoidali. Si è scelto di misurare 10 campioni di uova di P. edwardsii per mese, utilizzando un microscopio ottico dotato di oculare micrometrico. Non è sempre stato facile effettuare le misurazioni, soprattutto quando si è
dovuto misurare le larve, poiché queste mantengono la forma ad “U”, che possedevano all’interno dell’uovo con il telson incastrato nella fossetta che si forma tra gli occhi; in questo caso non si è misurato il diametro, ma la lunghezza totale e per fare ciò è stato necessario distendere le larve, grazie all’ausilio di un ago particolare e di pinzette da laboratorio. Alcuni campioni di uova sono stati misurati anche a fresco, al fine di stimare il restringimento dimensionale dovuto alla fissazione in formalina. Durante i conteggi e le misurazioni è stata raccolta un’ampia documentazione fotografica delle uova, al fine di individuare gli stadi di sviluppo delle uova e delle future larve.
4.5 Analisi dei dati
Per la registrazione dei dati è stato creato un Data base ad hoc che ha permesso, una volta archiviate le informazioni raccolte, di ottenere vari tipi di elaborazioni secondo “queries” prestabilite. È stato così possibile stimare la demografia di P. edwardsii.
Demografia
È stato prodotto un istogramma taglia-frequenza. Relativamente a ciascun campione mensile. Sempre per ciascun mese sono state calcolate taglia media ed errore standard. La struttura in taglia è stata calcolata sia per tutta la popolazione che separatamente per maschi e femmine.
Rapporto sessi
Il rapporto sessi è stato calcolato come percentuale di femmine su tutta la popolazione campionata. Per verificare differenze significative dal rapporto atteso di 1:1 tra maschi e femmine è stato utilizzato il test del chi quadro:
χ
2 = ∑ (numero atteso – numero osservato)2/ numero attesoI valori ottenuti sono stati poi confrontati con la tabella del chi quadro (con 2 gradi di libertà). I confronti sono stati effettuati per ciascun campione mensile, nei casi con catture superiori a 50 individui.
Relazione taglia – peso
La relazione taglia peso è stata calcolata separatamente per i due sessi. La significatività delle differenze tra le curve di maschi e femmine sono state analizzate attraverso l’analisi della Covarianza, utilizzando il programma Statistica.
E’ stato utilizzato il test t di Student con n-2 gradi di libertà (Pauly, 1984) per testare se esiste una differenza statisticamente significativa da 3 del parametro b dell’equazione della curva di crescita.
Riproduzione e fecondità
Per l’analisi del periodo riproduttivo della specie è stata considerata la percentuale di femmine ai diversi stadi di maturità sia delle gonadi che delle uova e la percentuale di femmine ovigere, nei diversi mesi dell’anno.
L’analisi della taglia a cui avviene la riproduzione è stata effettuata considerando sia la percentuale di femmine con gonadi mature che quella con uova sui pleopodi (ovigere) per classi di taglia. La taglia di prima maturità fisiologica è stata definita come la taglia (LC) alla quale il 50% delle femmine presenta gonadi mature o in stato di maturazione avanzata. La taglia di prima maturità funzionale è stata definita come la taglia (LC) a cui il 50% delle femmine è ovigere (Guénnegan et al., 1992).
Le stime sono state effettuate adattando i dati sperimentali ad una curva logistica.
Le stime di fecondità assoluta sono state effettuate espandendo i conteggi del campione (25 cellette) al totale delle cellette (200) presenti nella Dollfuss. In pratica è stato moltiplicato il campione x 8 celle contate)
La fecondità relativa (numero di uova in relazione alla lunghezza o al peso individuale) è stata stimata attraverso regressioni lineari o esponenziali di queste misure. È stato utilizzato il coefficiente di
regressione (R2) per valutare l’adattamento dei dati sperimentali al modello utilizzato.
Sono stati prodotti istogrammi di frequenza di taglia delle uova, per ogni stadio di sviluppo delle uova stesse.
Relazione taglia – peso
La relazione taglia peso è stata calcolata separatamente per i due sessi, mediante lo studio delle regressioni tra i dati di lunghezza e di peso. E’ stato utilizzato il test t di Student con n-2 gradi di libertà (Pauly, 1984) per testare se esisteva una differenza statisticamente significativa da 3 del parametro b dell’equazione della curva di crescita. Questo test è stato utilizzato anche per verificare differenze significative tra il b dei maschi e quello delle femmine.
Tutti i risultati ottenuti da questo lavoro di tesi sono stati confrontati con quelli noti in bibliografia.