Risultati e discussione
Dati climatici
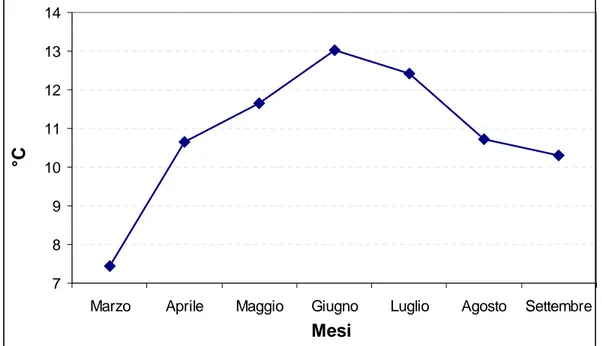
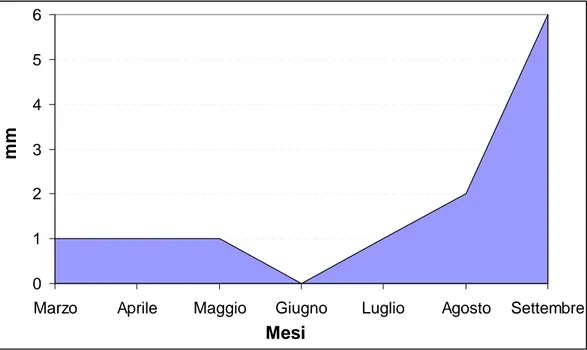
Il comportamento del vigneto è fortemente condizionato dall’andamento meteorologico dell’anno in corso, per cui è sicuramente utile poter disporre dei dati relativi a temperature (massima, minima, media) mensili (Figura 52), escursione termica media mensile (Figura 53), piovosità media mensile (Figura 54) e radiazione solare media mensile (Figura 55).
0 5 10 15 20 25 30 35 Ma rzo April e Ma ggio Giug no Lugl io Agos to Sett embr e Mesi °C
TMax TMed TMin
Figura 52. Andamento mensile delle temperature (massima, minima, media) dal germogliamento alla vendemmia nell’annata 2006.
7 8 9 10 11 12 13 14
Marzo Aprile Maggio Giugno Luglio Agosto Settembre
Mesi
°C
Figura 53. Andamento mensile dell’escursione termica dal germogliamento alla vendemmia nell’anno 2006.
0 1 2 3 4 5 6
Marzo Aprile Maggio Giugno Luglio Agosto Settembre
Mesi
mm
Figura 54. Andamento pluviometrico mensile dal germogliamento alla vendemmia nell’anno 2006.
0 50 100 150 200 250 300
Marzo Aprile Maggio Giugno Luglio Agosto Settembre
Mesi
w/m
q
/o
ra
Figura 55. Andamento della radiazione solare media dal germogliamento al pianto nell’anno 2006
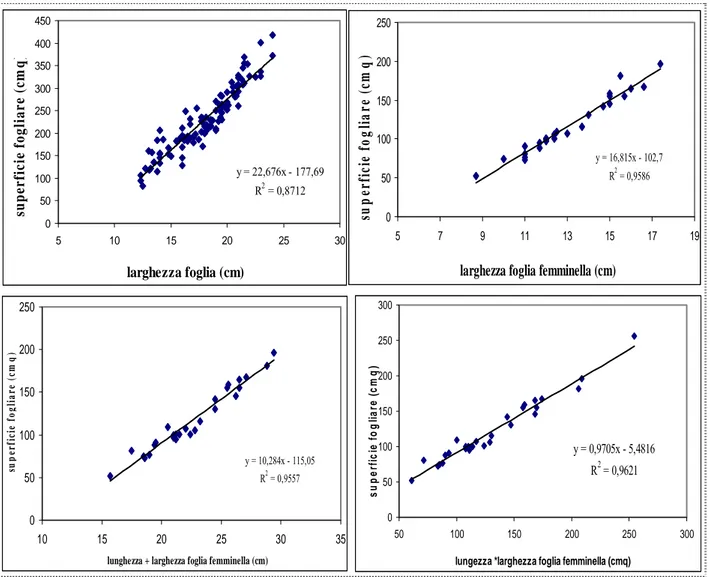
La tabella 1 ci indica alcuni parametri caratteristici delle foglie del tralcio e delle foglie delle femminelle di Ciliegiolo, mentre la fig ci indica come sia la larghezza il parametro più indicativo per la misurazione dell’area fogliare delle foglie del tralcio e delle foglie delle femminelle, tramite l’equazione riportata sulla figura 56; infatti esiste una correlazione molto alta tra larghezza della foglia ed area fogliare, come testimoniato dal valore R² riportato in figura . Per le foglie delle femminelle ci sono altri due parametri, larghezza + lunghezza e larghezza*lunghezza, che hanno un alto coefficiente di correlazione con l’area fogliare (Tabella 1).
Tabella 1. Parametri medi relativi a 150 foglie del tralcio e a 30 foglie delle femminelle.
132,37 23,00 1,38 13,28 9,69 126,72 Foglie femminelle 249,70 32 1,34 18,00 13,56 229,97 Foglie Lunghezza * Larghezza (cm²) Lunghezza + Larghezza (cm) Largezza / Lunghezza Larghezza (cm) Lunghezza (cm) Superficie (cm²) Parametri
y = 22,676x - 177,69 R2 = 0,8712 0 50 100 150 200 250 300 350 400 450 5 10 15 20 25 30 larghezza foglia (cm) sup erfi c ie fog lia re (c m q ) y = 16,815x - 102,7 R2 = 0,9586 0 50 100 150 200 250 5 7 9 11 13 15 17 19
larghezza foglia femminella (cm)
su p er fi ci e f o g lia re ( cm q) y = 10,284x - 115,05 R2 = 0,9557 0 50 100 150 200 250 10 15 20 25 30 35
lunghezza + larghezza foglia femminella (cm)
su p er fi ci e fo g li a re ( cm q) y = 0,9705x - 5,4816 R2 = 0,9621 0 50 100 150 200 250 300 50 100 150 200 250 300
lungezza *larghezza foglia femminella (cmq)
s u p e rf ic ie f o g lia re ( cm q )
Figura 56. Correlazione tra parametri della foglia ed area fogliare
La tabella 2 ci dà indicazioni sull’intero apparato fogliare della pianta e ci permette di valutare se la sfogliatura ha prodotto differenze su di esso. Il gruppo di piante non sfogliate ha avuto una superficie fogliare totale decisamente superiore agli altri due gruppi sotto osservazione; questa differenza è provocata dal numero di foglie ma anche dal numero di femminelle.
Tabella 2. Parametri relativi alla superficie fogliare della singola pianta (valori medi): F/p = foglie del tralcio/pianta; SF/p = superficie delle foglie del tralcio/pianta; f/p = femminelle/pianta; Ff/p = foglie delle femminelle/pianta; Sf/p = superficie delle foglie delle femminelle/pianta; SFT/p = superficie fogliare totale/pianta; Sf/SF = superficie foglie femminelle/superficie foglie del tralcio. Lettere differenti indicano differenze statistiche nell’ambito della colonna per p<0.05.
40,52 a 59,48 a 1.75 a 5,23 a 3,17 a 3,29 a 3.93 a 259.27 a 67.76 a 1.95 a 84.90 a no sf 32,11 a 67,89 a 2,30 a 4.31 b 4,17 a 2.99 ab 4,57 a 235.66 ab 53.17 b 1.33 b 57.74 b sf inv 36,75 a 63,24 a 1.78 a 4.01 b 3,22 a 2.50 b 3.70 a 197.59 b 54.32 b 1.51 b 65.48 b sf all SF/SFT (%) Sf/SFT (%) Sf/SF SFT/p (m²) Ff/F Sf/p (m²) Ff/f Ff/p f/p SF/p (m²) F/p
Composizione ed evoluzione della bacca
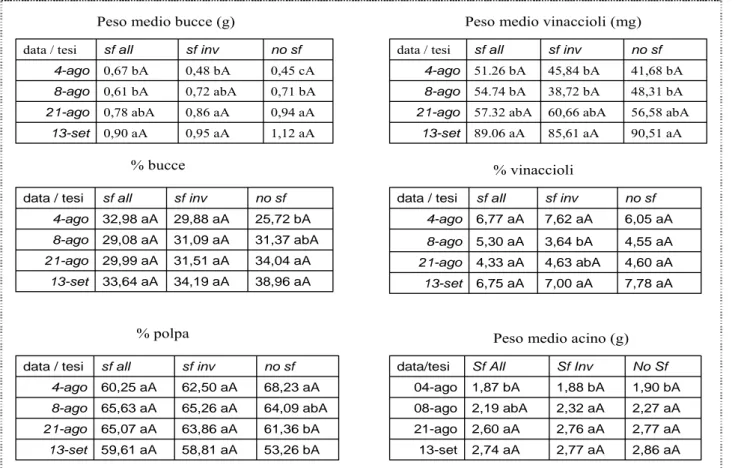
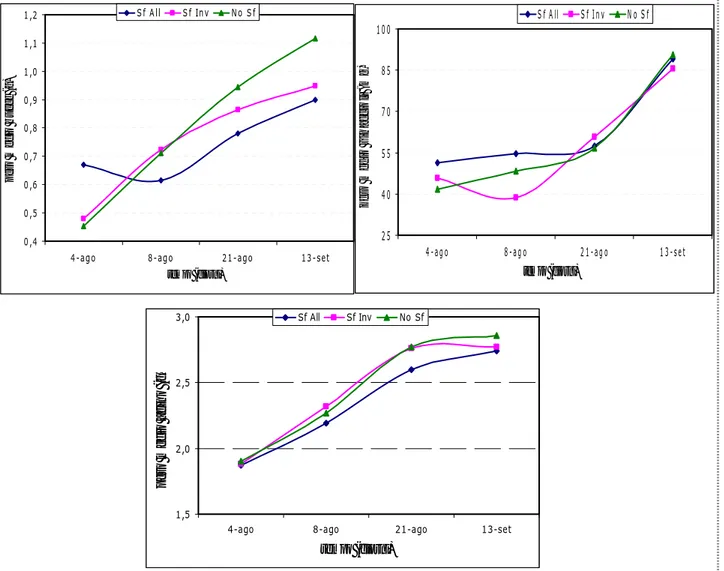
Il peso medio dell’acino è aumentato nel corso della maturazione per le tesi sotto osservazione (Figura 57), esiste infatti una differenza significativa fra l’acino maturo (13-sett) e l’acino in fase di invaiatura (4-ago) (Tabella 3). La sfogliatura non ha prodotto differenze sul peso medio dell’acino ed anche le proporzioni di bucce, vinaccioli e polpa sono simili fra i trattamenti (Tabella 3).
Tabella 3. Composizione della bacca, in relazione alla data di campionamento (4/8/2006 = 4-ago, 8/8/2006 = 8-ago, 21/8/2006 = 21-ago, 13/9/2006 = 13-sett) ed al trattamento di sfogliatura eseguito (Sf All = sfogliatura all’allegagione , Sf Inv = sfogliatura all’invaiatura, No Sf = non sfogliatura). Lettere differenti nella stessa riga, in maiuscolo, indicano differenze statisticamente significative tra le tesi nella stessa data di campionamento (test di Duncan p ≤ 0,05 ). Lettere differenti nella stessa colonna, in minuscolo, indicano differenze statisticamente significative tra le date di campionamento della stessa tesi (test di Duncan p ≤ 0,05).
1,12 aA 0,95 aA 0,90 aA 13-set 0,94 aA 0,86 aA 0,78 abA 21-ago 0,71 bA 0,72 abA 0,61 bA 8-ago 0,45 cA 0,48 bA 0,67 bA 4-ago no sf sf inv sf all data / tesi
Peso medio bucce (g)
90,51 aA 85,61 aA 89.06 aA 13-set 56,58 abA 60,66 abA 57.32 abA 21-ago 48,31 bA 38,72 bA 54.74 bA 8-ago 41,68 bA 45,84 bA 51.26 bA 4-ago no sf sf inv sf all data / tesi
Peso medio vinaccioli (mg)
% bucce % vinaccioli % polpa 38,96 aA 34,19 aA 33,64 aA 13-set 34,04 aA 31,51 aA 29,99 aA 21-ago 31,37 abA 31,09 aA 29,08 aA 8-ago 25,72 bA 29,88 aA 32,98 aA 4-ago no sf sf inv sf all data / tesi 7,78 aA 7,00 aA 6,75 aA 13-set 4,60 aA 4,63 abA 4,33 aA 21-ago 4,55 aA 3,64 bA 5,30 aA 8-ago 6,05 aA 7,62 aA 6,77 aA 4-ago no sf sf inv sf all data / tesi 53,26 bA 58,81 aA 59,61 aA 13-set 61,36 bA 63,86 aA 65,07 aA 21-ago 64,09 abA 65,26 aA 65,63 aA 8-ago 68,23 aA 62,50 aA 60,25 aA 4-ago no sf sf inv sf all data / tesi 2,86 aA 2,77 aA 2,74 aA 13-set 2,77 aA 2,76 aA 2,60 aA 21-ago 2,27 aA 2,32 aA 2,19 abA 08-ago 1,90 bA 1,88 bA 1,87 bA 04-ago No Sf Sf Inv Sf All data/tesi
0,4 0,5 0,6 0,7 0,8 0,9 1,0 1,1 1,2
4-ago 8-ago 21-ago 13-set tempo (giorni) p eso m e d io b u cce ( g ) Sf All Sf Inv No Sf 25 40 55 70 85 100
4-ago 8-ago 21-ago 13-set
tempo (giorni) p es o m ed io vi n a cc iol i (m g) Sf All Sf Inv No Sf 1,5 2,0 2,5 3,0
4-ago 8-ago 21-ago 13-set
tempo (giorni) pe so m e di o ac in o ( g ) Sf All Sf Inv No Sf
Figura 57. Evoluzione del peso medio dell’acino, delle bucce e dei vinaccioli nel corso della maturazione
La figura 58 ci mostra gli andamenti del pH, dell’acidità totale e degli zuccheri (°Brix), sui quali però non è stato possibile fare un’analisi statistica.
2 7 12 17 22 27
04-ago 08-ago 21-ago 13-set
tempo (giorni) °B ri x Sf All Sf Inv No Sf 2,4 2,6 2,8 3,0 3,2 3,4
04-ago 08-ago 21-ago 13-set
tempo (giorni) pH Sf All Sf Inv No Sf 7 8 9 10 11
04-ago 08-ago 21-ago 13-set
tempo (giorni) A cid it à t o ta le g /l Sf All Sf Inv No Sf
Figura 58. Evoluzione del pH, dell’acidità totale (g/l) e degli zuccheri (°Brix) durante la maturazione dell’acino.
E’ sorprendente che il trattamento di sfogliatura non ha influenzato il peso medio dell’acino, ma anche Bledsoe et a.l (1988) e Zoecklein et al. (1992) hanno ottenuto un risultato simile. Varie ipotesi possono essere formulate per spiegare questo risultato, incluso un aumento del tasso di fotosintesi delle foglie, o un cambiamento nel movimento degli assimilati (Ollat et al. 1998), che possono essere confermate indirettamente dalla valutazione della concentrazione degli zuccheri alla vendemmia, pressoché identici nelle tre tesi sotto esame, considerando che il peso medio dell’acino, alla vendemmia, è statisticamente uguale.
Profilo di carotenoidi e clorofille di uve di Ciliegiolo.
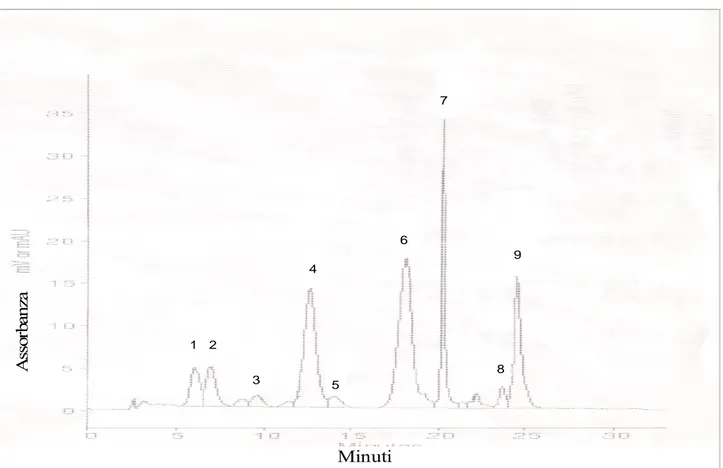
Le analisi relative alla separazione dei carotenoidi di uve di Ciliegiolo tramite HPLC P200 Spectra Series (Spectra Physics) mostra che sono stati identificati vari composti (Figura 59): β carotene, α carotene, luteina, neoxantina, violaxantina, anteraxantina, zeaxantina e le clorofille a e b.
A sso rb an za Minuti 1 2 3 4 5 6 7 8 9
Figura 59. Cromatogramma HPLC di carotenoidi e clorofille estratte da uva Ciliegiolo. Picchi : (1) Neoxantina, (2) Violaxantina, (3) Anteraxantina, (4) Luteina, (5) Zeaxantina, (6) Clorofilla B, (7) Clorofilla A, (8) α-carotene, (9) β-carotene.
.
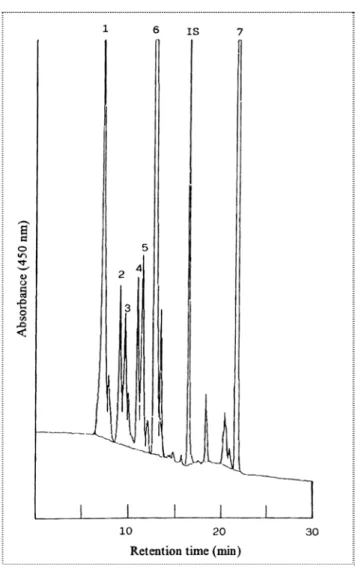
I composti presenti in maggior quantità nelle uve sono la luteina (picco 4), il β-carotene (picco 9) e clorofilla b (picco 6), come riportato da altri autori (figura 60, figura 61) (Guedes de Pinho et al. 2001, Mendes Pinto et al. 2005, Razungles et al. 1996). Gli altri picchi non identificati probabilmente sono isomeri dei composti da noi identificati mediante l’utilizzo di standard specifici
Figura 60. Esempio di separazione HPLC dei carotenoidi da bacche mature di Syrah, monitorate a 450nm: (1) neoxantina; (2) violaxantina; (3) luteoxantina; (4) luteina 5-6 epossido; (5) flavoxantina; (6) luteina; (7) β carotene; (IS) internal standard ( β-apo-8′-carotenal) (Razungles et al. 1996).
Figura 61. Esempio di profilo HPLC di carotenoidi, clorofilla e derivati della clorofilla isolati dall’uva. Picchi: (1) neocromo a; (2) neoxantina; (3) neocromo b; (4) violaxantina; (5) luteina 5,6 – epossido; (6) flavoxantina; (7) non identificato; (8) non identificato; (9) non identificato; (10) ) non identificato; (11) (13z) o (13′z) luteina; (12) clorofilla b; (13) feofitina b; (14) feofitina a; (15) (E) β carotene; (16) (13z) β carotene; (IS) internal standard ( β-apo-8′-carotenal) (Mendes Pinto et al. 2005).
Evoluzione di carotenoidi e clorofille durante la maturazione del grappolo
Le quantità di carotenoidi e clorofille sono state monitorate durante l’ultimo mese di maturazione delle uve di Ciliegiolo, prelevando 4 campioni in date differenti (4/8/2006, 8/8/2006, 21/8/2006, 13/9/2006) per le tre tesi sotto osservazione ( Sf All = sfogliatura all’allegagione , Sf Inv = sfogliatura all’invaiatura, No Sf = non sfogliatura) (Tabella 4).
Tabella 4. Contenuto di carotenoidi (α-carotene, β-carotene, caroteni totali, luteina, neoxantina, violaxantina, anteraxantina, zeaxantina, xantofille totali, carotenoidi totali) e clorofille (clorofilla a, clorofilla b, clorofilla totale) in uve di Ciliegiolo, espressi in μg/g di peso secco, in relazione alla data di campionamento (4/8/2006 = 4-ago, 8/8/2006 = 8-ago, 21/8/2006 = 21-ago, 13/9/2006 = 13-sett) ed al trattamento di sfogliatura eseguito (Sf All = sfogliatura all’allegagione , Sf Inv = sfogliatura all’invaiatura, No Sf = non sfogliatura).
Lettere differenti nella stessa riga, in maiuscolo, indicano differenze statisticamente significative tra le tesi nella stessa data di campionamento (test di Duncan p ≤ 0,05 ). Lettere differenti nella stessa colonna, in minuscolo, indicano differenze statisticamente significative tra le date di campionamento della stessa tesi (test di Duncan p ≤ 0,05).
α-carotene β-carotene
Caroteni totali Luteina
Neoxantina Violaxantina 0,75 cA 0,10 cA 0,52 bA 13-set 1,52 bcA 0,43 cB 0,55 bB 21-ago 2,65 bA 1,72 bB 1,36 bB 08-ago 7,56 aA 7,56 aA 4,73 aB 04-ago no sf sf inv sf all data / tesi 4,90 cA 4,44 cA 5,80 cA 13-set 6,65 bcA 4,45 cB 7,46 bA 21-ago 7,510 bA 7,00 bA 6,51 bcA 08-ago 11,74 aA 11,74 aA 10,71 aA 04-ago no sf sf inv sf all data / tesi 5,66 cA 4,53 bA 6,33 bA 13-set 8,20 bcA 4,90 bB 8,18 bA 21-ago 10,16 bA 8,71 bA 7,87 bA 08-ago 19,30 aA 19,30 aA 15,43 aB 04-ago no sf sf inv sf all data / tesi 2,25 cA 1,91 bA 2,58 cA 13-set 3,11 bcAB 2,24 bB 3,74 bA 21-ago 4,73 bA 4,43 bA 4,36 bA 08-ago 10,12 aA 10,12 aA 9,60 aA 04-ago no sf sf inv sf all data / tesi 0,56 cA 0,36 cA 0,58 cA 13-set 0,84 bcA 0,52 cA 0,77 bcA 21-ago 1,17 bA 1,16 bA 1,02 bA 08-ago 2,86 aA 2,86 aA 2,45 aB 04-ago no sf sf inv sf all data / tesi 0,61 aB 0.60 aB 0,89 bA 13-set 0,84 aB 0,74 aB 1,30 aA 21-ago 0,60 aA 0.74 aA 0,58 cA 08-ago 0,62 aB 0.62 aB 0,83 bA 04-ago no sf sf inv sf all data / tesi Carotenoidi totali 9,47 cA 7,61 cA 10,82 cA 13-set 13,48 bcA 9,11 bcB 14,36 bA 21-ago 17,01 bA 15,69 bA 14,40 bA 08-ago 34,32 aA 34,32 aA 29,70 aB 04-ago no sf sf inv sf all data / tesi 3,81 cA 3,10 cA 4,50 cA 13-set 5,30 bcA 4,24 bcB 6,36 bA 21-ago 6,85 bA 6,98 bA 6,54 bA 08-ago 15,01 aA 15,01 aA 14,26 aA 04-ago no sf sf inv sf all data / tesi Xantofille totali Zeaxantina 0,18 bA 0,04 cA 0,16 bA 13-set 0,21 bA 0,33 bA 0,14 bA 21-ago 0,12 bB 0,30 bA 0,23 abA 08-ago 0,89 aA 0,89 aA 0,48 aB 04-ago no sf sf inv sf all data / tesi Anteraxantina 0,20 bA 0,17 cA 0,27 bA 13-set 0,30 bA 0,41 abA 0,41 bA 21-ago 0,23 bA 0,35 bA 0,34 bA 08-ago 0,59 aA 0,59 aA 0,82 aA 04-ago no sf sf inv sf all data / tesi
Clorofilla A 13,60 aA 20,17 aA 21,56 cAB 13-set 17,40 aB 23,04 aB 36,80 bcA 21-ago 26,45 aA 44.32 aA 39,53 bA 08-ago 45,84 aA 45,84 aA 67,35 aA 04-ago no sf sf inv sf all data / tesi Clorofilla B 10,61 cA 8,25 cA 11,68 cA 13-set 17,00 bcA 11,00 cB 17,00 bA 21-ago 27,22 bA 28,07 bA 23,24 bA 08-ago 56,40 aA 56,40 aA 48,33 aB 04-ago no sf sf inv sf all data / tesi 24,21 bA 28,42 bA 33,24 cA 13-set 34,31 bB 34,00 bB 53,90 bcA 21-ago 53,67 bcA 72,39 abA 62,77 bA 08-ago 102,25 aA 102,25 aA 115,68 aA 04-ago no sf sf inv sf all data / tesi Clorofilla totale
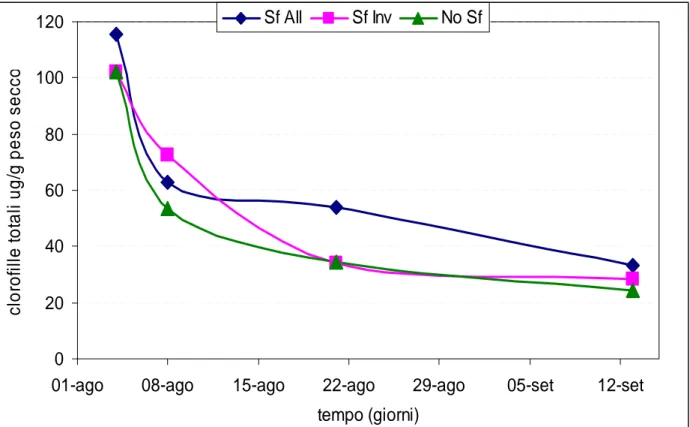
In generale, i risultati mostrano che, per tutte e tre le tesi, il contenuto in carotenoidi totali (Figura 62) e clorofille totali (Figura 63) decresce durante la maturazione, anche se in modo non lineare: decade rapidamente durante l’invaiatura (1-8 agosto), mentre questo fenomeno rallenta con l’avvicinarsi della maturazione e si possono ancora trovare tracce di questi composti nell’uva matura. Quando si prendono in considerazione i singoli carotenoidi, α-carotene, β-carotene, luteina, neoxantina, anteraxantina, zeaxantina, i caroteni totali (α-carotene + β-carotene) e le xantofille totali (luteina + neoxantina + anteraxantina + violaxantina + zeaxantina) (Figura 64), l’andamento è simile a quello dei carotenoidi totali, così come avviene anche per le clorofille a e b (Figura 65) rispetto alla clorofilla totale, come dimostrato nella tabella, dove per tutti i parametri analizzati, con eccezione della violaxantina, sussite una differenza significativa tra la data di vendemmia (13/9/2006) e l’invaiatura (4/8/2006) per le tre tesi sotto osservazione.
E’ durante l’ invaiatura che avviene la variazione nel contenuto di carotenoidi e clorofille; il decremento inizia quando il colore delle bacche comincia a virare verso il rosso e da questo momento il decadimento incrementa sempre più fino al termine dell’invaiatura quando la bacca è completamente rossa. E’ quindi la fine dell’invaiatura (8/8/2006) il momento chiave per il cambiamento nel contenuto di carotenoidi e clorofille, come dimostra la tab, in cui si notano differenze statistiche significative per tutti i componenti, fatta eccezione per la violaxantina, tra la data del 4/8/2006 e quella del 8/8/2006.
0 5 10 15 20 25 30 35 40
01-ago 08-ago 15-ago 22-ago 29-ago 05-set 12-set
tempo (giorni) ca rot en oi di to ta li ug /g pe so s ec co Sf All Sf Inv No Sf
Figura 62. Evoluzione dei carotenoidi totali durante la maturazione
0 20 40 60 80 100 120
01-ago 08-ago 15-ago 22-ago 29-ago 05-set 12-set
tempo (giorni) c lo ro fi ll e to ta li u g /g p e s o s e c c o Sf All Sf Inv No Sf
0 2 4 6 8 10 12 14
01-ago 08-ago 15-ago 22-ago 29-ago 05-set 12-set tempo (giorni) B c a ro te n e u g /g p e s o s e c c o Sf All Sf Inv No Sf 0 1 2 3 4 5 6 7 8
01-ago 08-ago 15-ago 22-ago 29-ago 05-set 12-set tempo (giorni) a -c a ro tene u g /g p e s o se cco Sf All Sf Inv No Sf 0 2 4 6 8 10 12 14 16 18 20
01-ago 08-ago 15-ago 22-ago 29-ago 05-set 12-set tempo (giorni) C a ro te n i to ta li u g /g p e s o s e c c o Sf All Sf Inv No Sf 0 2 4 6 8 10 12
01-ago 08-ago 15-ago 22-ago 29-ago 05-set 12-set tempo (giorni) L u te in a ug /g pe s o s e c c o Sf All Sf Inv No Sf 0,0 0,5 1,0 1,5 2,0 2,5 3,0
01-ago 08-ago 15-ago 22-ago 29-ago 05-set 12-set tempo (giorni) n eox ant ina u g /g p es o s e c c o Sf All Sf Inv No Sf 0,0 0,2 0,4 0,6 0,8 1,0
01-ago 08-ago 15-ago 22-ago 29-ago 05-set 12-set tempo (giorni) ant er ax ant in a u g /g p es o s e c c o Sf All Sf Inv No Sf 0,0 0,2 0,4 0,6 0,8 1,0
01-ago 08-ago 15-ago 22-ago 29-ago 05-set 12-set
tempo (giorni) z e ax a n ti n a u g /g pes o s e c c o Sf All Sf Inv No Sf 2 4 6 8 10 12 14 16
01-ago 08-ago 15-ago 22-ago 29-ago 05-set 12-set
tempo (giorni) X a n to fi ll e to ta li u g /g p e s o s e c c o Sf All Sf Inv No Sf
Figura 64. Evoluzione del β carotene, α carotene, caroteni totali, luteina, neoxantina, anteraxantina, zeaxantina, xantofille totali nel corso della maturazione del grappolo
0 10 20 30 40 50 60 70
01-ago 08-ago 15-ago 22-ago 29-ago 05-set 12-set
tempo (giorni) c lor of il la A u g /g p e s o s e c c o Sf All Sf Inv No Sf 0 10 20 30 40 50 60
01-ago 08-ago 15-ago 22-ago 29-ago 05-set 12-set
tempo (giorni) cl o ro fi ll a B u g /g p e so se cco
Figura 65. Evoluzione della clorofilla a e della clorofilla b durante la maturazione
Mentre per le clorofille la causa del decremento è senza dubbio la perdita dei cloroplasti e la fine dell’attività fotosintetica, la causa del decremento dei carotenoidi, durante la maturazione del grappolo, non è conosciuta con certezza ed è possibile solo fare ipotesi. E’ possibile che dopo la loro distruzione, i cloroplasti non siano trasformati in cromoplasti capaci di sintetizzare i carotenoidi o alternativamente ci può essere una competizione tra la sintesi di carotenoidi e flavonoidi per i loro precursori comuni, l’acetato e il mevalonato. La sintesi dei flavonoidi avverebbe a discapito di quella dei carotenoidi, che durante la maturazione sono progressivamente degradati, per via enzimatica o fotochimica, senza che una nuova sintesi li possa rimpiazziare e successivamente trasformati in C-13 norisoprenidi, liberi o glicosilati. I quantitativi di carotenoidi presenti ancora nell’uva matura vengono trasferiti in seguito nel vino e saranno poi qui trasformati in aromi durante l’invecchiamento. E’ ipotizzabile che anche dalla degradazione delle clorofille si possano originare composti implicati nella qualità organolettica dell’uva e del vino, anche se questo non è stato ancora dimostrato e può rappresentare un’interessante linea di ricerca per il futuro. Mendes Pinto et al. (2005) hanno identificato alcuni composti di degradazione della clorofilla nelle uve mature di varietà portoghesi, come Touriga Nacional o Tinta Roiz, e nei vini Porto.
Per quanto riguarda i carotenoidi i maggiori componenti sono il β-carotene e la luteina, che sono anche rispettivamente i principali composti del gruppo dei caroteni e delle xantofille in tutte le tesi. Anche l’α-carotene è presente in notevole quantità e questo fa sì che il gruppo dei caroteni sia quello che incide di più sul peso totale dei carotenoidi. Le clorofilla a e b incidono im modo simile sulla quantità di clorofilla totale.
La violaxantina ha un comportamento particolare durante il periodo di maturazione, la sua evoluzione non segue quella degli altri carotenoidi (Figura 66). E’ presente in minima quantità all’inizio dell’invaiatura ed il suo contenuto cresce, probabilmente a causa di una bioconversione da β-carotene a violaxantina, per poi tornare sui livelli iniziali, come si può notare dalla tabella , dove alla data 21/8/2006 si registra il valore più alto di violaxantina in tutte le tesi. A questi risultati sono giunti anche altri autori (Baumes et al. 2002, Giovannelli G. et al. 2006, Guedes de Pinho et al. 2003, Mendes Pinto et al. 2005, Oliveira et al. 2004, Razungles et al. 1988, Razungles et al. 1996, Razungles et al. 1998) 0,5 0,8 1,0 1,3 1,5
01-ago 08-ago 15-ago 22-ago 29-ago 05-set 12-set
tempo (giorni) V iol axan ti na ug /g peso secc o Sf All Sf Inv No Sf
Effetto della sfogliatura sul contenuto in carotenoidi e clorofille di uve Ciliegiolo
Per valutare l’effetto dei trattamenti di sfogliatura sul contenuto in carotenoidi e clorofille di uve Ciliegiolo è necessario focalizzare l’attenzione su due date, il 4/8/2006 ed il 13/9/2006.La valutazione dei parametri al 4/8/2006 (Tabella 4) ci fornisce indicazioni sull’effetto della sfogliatura precoce durante la fase erbacea dell’acino, rispetto alle altre due tesi (controllo e sfogliatura all’invaiatura), che fino a questo momento possono considerarsi identiche.
Le variazioni statisticamente significative riguardano i singoli metaboliti α carotene, neoxantina, zeaxantina, violaxantina, e per quanto riguarda i totali i caroteni totali e i carotenoidi totali (Figura 67). 0 5 10 15 20 25 Sf All Sf Inv No Sf tesi ca ro te n i t o ta li u g /g p e so se cco 2,2 2,3 2,4 2,5 2,6 2,7 2,8 2,9 3,0 Sf All Sf Inv No Sf tesi ne o x a n ti na ug/ g pe s o s e c c o 0,0 0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9 1,0 Sf All Sf Inv No Sf tesi z eaxa n ti n a u g /g p eso secco B A A B B B A A A A A 0 2 4 6 8 10 Sf All Sf Inv No Sf tesi a -c a rot e n e ug/ g pe s o s e c c o A A B
0,0 0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9 Sf All Sf Inv No Sf tesi vi o laxan ti n a u g /g p eso secco A B B B A 20 24 28 32 36 Sf All Sf Inv No Sf tesi c a ro te noi di t o ta li ug/ g pe s o s e c c o A A B
Figura 67. Effetto della sfogliatura sul contenuto di carotenoidi (μg/g peso secco) di uve di Ciliegiolo in relazione ai trattamenti di sfogliatura. Parametri statisticamente differenti al 4/8/2006: α-carotene, caroteni totali, neoxantina, zeaxantina, violaxantina, carotenoidi totali. Lettere differenti, in maiuscolo, indicano differenze significative, secondo il test di Duncan (α ≤ 0,05 ), tra le tesi della stessa data di campionamento.
Si nota che la sfogliatura all’allegagione ha prodotto uve con un minor quantitativo di carotenoidi totali e quindi con minor potenziale aromatico, in quanto questi composti durante la maturazione saranno convertiti in C-13 norisoprenoidi. E’ il gruppo dei caroteni totali, ed in particolar modo l’ α-carotene, che ha generato questa differenza, con le xantofille interessate in modo minore. Le ragioni di questa differenza si possono ascrivere all’eccessivo stress luminoso subìto da i grappoli delle piante sfogliate all’allegagione, che se da una potrebbe indurre la sintesi ex novo dei carotenoidi, dall’altra ne può determinare la degradazione a causa dalla presenza di specie attive dell’ O2 (ROS), generate in conseguenza di un eccesso (stress fotoossidativo) di energia, che può
colpire i centri di reazione dei fotosistemi e cioè quando si verifica la situazione in cui la quantità di fotoni disponibili risulta troppo elevata rispetto a quella richiesta per l’assimilazione della CO2.
Questa situazione si verifica quando ancora i cloroplasti sono integri e svolgono un attiva fotosintesi. Nel nostro caso la diminuzione del contenuto di caroteni totali a seguito di intensa illuminazione nella tesi sfogliatura all’allegagione, trova giustificazione nel ruolo fotoprotettivo svolto da questi pigmenti, grazie alla loro capacità di deeccitare sia la clorofilla tripletto (3Chl*), quando i cloroplasti sono attivamente funzionanti, che di eliminare specie attive dell’O2 , che si
originano in seguito a processi fotoossidativi che possono indurre una distruzione di questi metabolici (Senger et al. 1993). La degradazione dei pigmenti è un sintomo comune del danno ossidativo che i vari organi delle piante possono subire e spesso e spesso si verifica in seguito a condizioni di stress ambientali, quali carenze nutrizionali, elevata irradiazione, alte o basse temperature, deficit idrico, inquinamento atmosferico, che hanno la conseguenza di aumentare la probabilità di produzione di specie attive dell’ossigeno, come lo ione superossido (O2ֿ) e l’ossigeno
singoletto (1O
2 ). E’ generalmente provato che i radicali dell’ossigeno e l’ossigeno singoletto (1O2 )
sono coinvolti nella degradazione sia dei pigmenti fotosintetici che accessori. La fotoossidazione dei pigmenti avviene in conseguenza ad una perdita o ad un danno dei fattori protettivi per la fotoinibizione, il deeccitamento degli stati eccitati e l’inattivazione delle specie reattive
dell’ossigeno (Asada 2006, Merzlyack et al. 1998, Adams et al 1996 A, Demmings-Adams et al. 1996 B). La violaxantina mostra un comportamento opposto e si trova in maggiori quantità nella tesi sfogliatura precoce; essa infatti viene prodotta all’inizio dell’invaiatura e poichè i grappoli di questa tesi sono già esposti al sole, il suo accumulo è stato favorito.
La valutazione dei parametri alla data del 13/9/2006 (Tabella 4) è importante per stabilire se tra la pratica della sfogliatura all’invaiatura rispetto alla sfogliatura precoce e rispetto al controllo, si siano verificate differenze nella degradazione dei pigmenti. La sfogliatura non ha prodotto differenze significative tra le tesi sotto osservazione sul contenuto di carotenoidi e di clorofille al momento della vendemmia (13/9/2006), ad eccezione della violaxantina, come riportato in tabella 4. Questo è un risultato inaspettato, poiché se la degradazione dei carotenoidi viene promossa dalla luce che colpisce il grappolo, il gruppo di piante non sfogliate avrebbe dovuto mostrare un contenuto maggiore di carotenoidi alla vendemmia rispetto alle tesi sfogliate; evidentemente durante la maturazione del grappolo si sono verificate delle condizioni ambientali per cui la luce non è risultata un fattore dicriminante. Quindi la sfogliatura all’invaiatura ed all’allegagione, in questo caso, non hanno avuto alcun effetto sull’evoluzione di carotenoidi e clorofille durante la maturazione del grappolo, eccetto che per la violaxantina che peraltro rappresenta un carotenoide minore per la sua bassa concentrazione.
Polifenoli della buccia
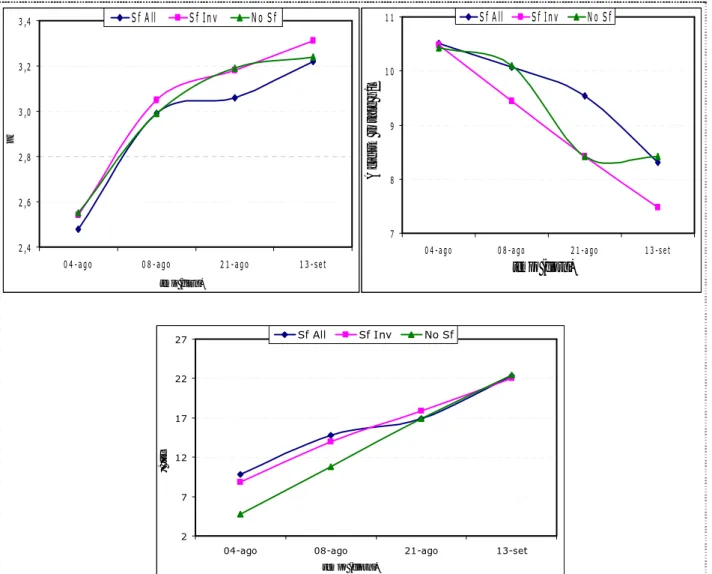
Esprimendo i polifenoli delle bucce in mg/Kg acino non si trovano differenze né sull’evoluzione dei polifenoli nel corso della maturazione né fra i trattamenti di sfogliatura (Tabella 5). Quando si esprimono in mg/kg bucce acino si nota sia un’incremento nella concentrazione di polifenoli delle bucce durante la maturazione, sia una differenza di concentrazione fra le tesi, già a partire dall’acino completamente invaiato (8-ago); in particolare alla vendemmia (13-sett) si registra un valore più alto di polifenoli delle bucce negli acini provenienti da piante sfogliate all’allegagione rispetto al controllo ed alla sfogliatura all’allegagione (Tabella 5). Quando l’unità di misura è mg/peso medio dell’acino si può notare solo l’incremento dei polifenoli delle bucce nel corso della maturazione (Figura 68), mentre l’effetto della sfogliatura risulta nullo (Tabella 5).
Tabella 5. Contenuto dei polifenoli delle bucce in uve di Ciliegiolo, in relazione alla data di campionamento (4/8/2006 = 4-ago, 8/8/2006 = 8-ago, 21/8/2006 = 21-ago, 13/9/2006 = 13-sett) ed al trattamento di sfogliatura eseguito (Sf All = sfogliatura all’allegagione , Sf Inv = sfogliatura all’invaiatura, No Sf = non sfogliatura).
Lettere differenti nella stessa riga, in maiuscolo, indicano differenze statisticamente significative tra le tesi nella stessa data di campionamento (test di Duncan p ≤ 0,05 ). Lettere differenti nella stessa colonna, in minuscolo, indicano differenze statisticamente significative tra le date di campionamento della stessa tesi (test di Duncan p ≤ 0,05).
1209 aA 1224 aA 1674 aA 13-set 920 aB 1225 aAB 1444 aA 21-ago 870 aA 847 aA 1217 aA 8-ago 1243 aA 1150 aA 1002 aA 4-ago no sf sf inv sf all data/tesi
Polifenoli bucce mg/Kg acino Polifenoli mg/Kg bucce
3,43 aA 3,33 aA 4,38 aA 13-set 2,54 bB 3,37 aA 3,75 abA 21-ago 1,95 bB 1,97 bB 2,58 bcA 8-ago 2,19 bA 1,78 bA 2,04 cA 4-ago no sf sf inv sf all data/tesi
Polifenoli bucce mg/acino
3133 bB 3663 aAB 4938 aA 13-set 2700 bB 4141 aAB 4803 aA 21-ago 2755 bB 2777 aB 4300 abA 8-ago 4771 aA 3935 aA 3045 bA 4-ago no sf sf inv sf all data/tesi 2500 3000 3500 4000 4500 5000
4-ago 8-ago 21-ago 13-set tempo (giorni) p o li fen o li b u cce m g /K g b u cci a Sf All Sf Inv No Sf 750 1000 1250 1500 1750
4-ago 8-ago 21-ago 13-set tempo (giorni) p o lif en o li b u cc e m g /K g ac in o Sf All Sf Inv No Sf 1 2 3 4 5
4-ago 8-ago 21-ago 13-set
tempo (giorni) po lif enoli bucce mg/ a cino Sf All Sf Inv No Sf
L’icremento dei polifenoli delle bucce durante la maturazione dell’acino è stato verificato anche da altri autori (Fournand et al. 2006, Mazza et al. 1999). E’ importante sottolineare come i polifenoli delle bucce di uve provenietenti dai trattamenti di sfogliatura, quando sono espressi per unità di buccia (mg/Kg bucce), sono risultati superiori ai valori riscontrati nel controllo; ciò significa che c’è stata un’effettiva maggiore biosintesi di questi composti, considerando che non ci sono differenze in peso tra le bucce delle uve sfogliate e non sfogliate (Tabella 5). Infatti la biosintesi dei flavonoidi è influenzata positivamente dall’esposizione alla luce e vari autori hanno riscontrato un contenuto di flavonoli e antociani maggiori in grappoli esposti alla luce piuttosto che in grappoli ombreggiati (Crippen et al. 1986, Haselgrove et al. 2000, Berqvist et al 2001). Inoltre la sfogliatura all’allegagione ha generato uve con un contenuto di polifenoli delle bucce superiore a quello di uve della tesi sfogliatura all’invaiatura, questo perché sfogliare dopo l’invaiatura vuol dire sottoporre il tessuto della buccia improvvisamente ad un regime radiativo e termico che non è abitutato a sostenere, a differenza dei grappoli di piante sfogliate all’allegagione. Le cellule, infatti, sono nel frattempo passate alla fase distensione cellulare, quindi subiscono un grande stress, che ne determina il collasso (Palese 2003). Quindi possiamo sostenere che la sfogliatura all’allegagione ha avuto un effetto positivo sulla biosintesi dei polifenoli delle bucce.
Polifenoli dei vinaccioli
I polifenoli dei vinaccioli decrescono durante la maturazione dell’acino (Tabella 6, Figura 69), ma la sfogliatura non ha prodotto alcune differenze rilevanti tra le tesi sotto esame (Tabella 6).
Tabella 6. Contenuto dei polifenoli dei vinaccioli uve di Ciliegiolo, in relazione alla data di campionamento (4/8/2006 = 4-ago, 8/8/2006 = 8-ago, 21/8/2006 = 21-ago, 13/9/2006 = 13-sett) ed al trattamento di sfogliatura eseguito (Sf All = sfogliatura all’allegagione , Sf Inv = sfogliatura all’invaiatura, No Sf = non sfogliatura).
Lettere differenti nella stessa riga, in maiuscolo, indicano differenze statisticamente significative tra le tesi nella stessa data di campionamento (test di Duncan p ≤ 0,05 ). Lettere differenti nella stessa colonna, in minuscolo, indicano differenze statisticamente significative tra le date di campionamento della stessa tesi (test di Duncan p ≤ 0,05).
20330 bA 22243 bA 23868 cA 13-set 36270 abA 37017 aA 35568 bcA 21-ago 46766 aA 50989 aA 43911 abA 8-ago 50352 aA 47897 aA 57470 aA 4-ago no sf sf inv sf all data/tesi Polifenoli mg/Kg vinaccioli 1357 bA 1537 bA 1463 bA 13-set 1631bA 1743bA 1700 bA 21-ago 2073 bA 1816 bA 2328 bA 8-ago 3056 aA 3589 aA 3870 aA 4-ago no sf sf inv sf all data /tesi
20000 30000 40000 50000 60000
4-ago 8-ago 21-ago 13-set
tempo (giorni) p o lif en o li m g /K g v in a cc io li Sf All Sf Inv No Sf 1000 1500 2000 2500 3000 3500 4000
4-ago 8-ago 21-ago 13-set
tempo (giorni) p o lif en o li v in a cc io li m g /K g a cin o Sf All Sf Inv No Sf
Figura 69. Andamento dei polifenoli dei vinacciolo nel corso della maturazione
I geni coinvolti nella sintesi dei polifenoli hanno differenti modi di espressione nei vinaccioli e nelle bucce, che ne determinano la concentrazione (Cortell et al. 2006). Infatti non si è verificata, come nel caso dei polifenoli delle bucce, un’influenza della luce sul contenuto dei polifenoli dei vinaccioli. Anche Downey et al. (2004) non hanno trovato nessun effetto della luce sul contenuto dei polifenoli dei vinaccioli, dovuto anche al fatto che diminuisce l’estraibilità dei polifenoli dei vinaccioli con il progredire della maturazione.
Polifenoli totali
I polifenoli totali si riducono notevolmente durante la maturazione dell’acino in tutte le tesi sotto osservazione (Figura 70), a causa del decremento dei polifenoli dei vinaccioli, che fino alla data del terzo campionamento (21-ago) rappresentano più del 50% in peso dei polifenoli totali (Tabella 7); da questo momento si assiste ad un recupero in percentuale dei polifenoli della buccia, che alla data di vendemmia (13-sett) si assestano su valori simili a queli dei polifenoli dei vinaccioli (Tabella 7). La sfogliatura non ha prodotto differenze rispetto al controllo (Tabella 7).
Tabella 7. Contenuto dei polifenoli totali in uve di Ciliegiolo, in relazione alla data di campionamento (4/8/2006 = 4-ago, 8/8/2006 = 8-ago, 21/8/2006 = 21-ago, 13/9/2006 = 13-sett) ed al trattamento di sfogliatura eseguito (Sf All = sfogliatura all’allegagione , Sf Inv = sfogliatura all’invaiatura, No Sf = non sfogliatura).
Lettere differenti nella stessa riga, in maiuscolo, indicano differenze statisticamente significative tra le tesi nella stessa data di campionamento (test di Duncan p ≤ 0,05 ). Lettere differenti nella stessa colonna, in minuscolo, indicano differenze statisticamente significative tra le date di campionamento della stessa tesi (test di Duncan p ≤ 0,05).
2566 bA 2761bA 3137 bA 13-set 2551 bA 2968 aA 3145 bA 21-ago 2943 bA 2664 aA 3546 abA 8-ago 4300 aA 4739 aA 4872 aA 4-ago no sf sf inv sf all data/tesi
Polifenoli totali mg/Kg acino
47,40 aA 44,50 aA 52,20 aA 13-set 37,60 bA 42,30 aA 45,90 aA 21-ago 33,10 bA 32,60 abA 34,10 bA 8-ago 25,60 bA 19,06 bA 21,01 cA 4-ago no sf sf inv sf all data / tesi % Polifenoli bucce 52,60 bA 55,50 bA 47,80 cA 13-set 62,40 aA 57,70 bA 54,10 cA 21-ago 66,90 aA 67,40 abA 65,90 bA 8-ago 74,40 aA 80,40 aA 79,00 aA 4-ago no sf sf inv sf all data / tesi % Polifenoli vinaccioli 2000 2500 3000 3500 4000 4500 5000
4-ago 8-ago 21-ago 13-set
tempo (giorni) p o li fe n o li t o ta li m g /K g a cin o Sf All Sf Inv No Sf
La sintesi dei polifenoli dei vinaccioli non è influenzata dall’esposizione luminosa e poiché il loro contenuto rappresenta all’incirca il 50% dei polifenoli totali alla vendemmia, si capisce come la sfogliatura non abbia avuto effetti sul quantitativo di polifenoli totali.
Antociani
La concentrazioni in antociani, nelle tre tesi, aumenta durante la maturazione dell’acino (Tabella 8, Figura 71). La sfogliatura ha inciso sulla concentrazione di antociani, quando questi sono espressi in mg/peso medio dell’acino si nota come la sfogliatura all’allegagione ha prodotto acini con un contenuto superiore di antociani rispetto al controllo e alla sfogliatura all’invaiatura (Tabella 8). Tabella 8. Contenuto degli antociani in uve di Ciliegiolo, in relazione alla data di campionamento (4/8/2006 = 4-ago, 8/8/2006 = 8-ago, 21/8/2006 = 21-ago, 13/9/2006 = 13-sett) ed al trattamento di sfogliatura eseguito (Sf All = sfogliatura all’allegagione , Sf Inv = sfogliatura all’invaiatura, No Sf = non sfogliatura).Lettere differenti nella stessa riga, in maiuscolo, indicano differenze statisticamente significative tra le tesi nella stessa data di campionamento (test di Duncan p ≤ 0,05 ). Lettere differenti nella stessa colonna, in minuscolo, indicano differenze statisticamente significative tra le date di campionamento della stessa tesi (test di Duncan p ≤ 0,05).
615 aA 694 aA 864 aA 13-set 532 abA 655 abA 559 aA 21-ago 277 bcB 439 bAB 472 abA 8-ago 12 cA 77 cA 123 bA 4-ago no sf sf inv sf all data/tesi Antociani mg/Kg acino 1608 aA 2143 aA 2610 aA 13-set 1569 aA 2142 aA 1831 aA 21-ago 887 abB 1443 aAB 1677 aA 8-ago 49 bA 262 bA 366 bA 4-ago no sf sf inv sf all data/ tesi Antociani mg/Kg buccia 1,73 bB 1,87 aAB 2,30 aA 13-set 1,47 bA 1,81 aA 1,47 bA 21-ago 0,63 aB 1,02 bA 1,00 bcA 8-ago 0,02 aA 0,12 cA 0,25 cA 4-ago no sf sf inv sf all data/tesi Antociani mg/acino
0 250 500 750 1.000
4-ago 8-ago 21-ago 13-set tempo (giorni) an to c ian i m g /k g ac in o Sf All Sf Inv No Sf 0 400 800 1.200 1.600 2.000 2.400 2.800
4-ago 8-ago 21-ago 13-set
tempo (giorni) a n to ci an i m g /k g b u cc ia Sf All Sf Inv No Sf 0,0 0,5 1,0 1,5 2,0 2,5
4-ago 8-ago 21-ago 13-set
tempo (giorni) an to c ia n i m g /a c in o Sf All Sf Inv No Sf
Figura 71. Andamento degli antociani durante la maturazione
Il metabolismo degli antociani risente dei cambiamenti delle condizioni di luce e temperatura. Se le condizioni della luce dentro la chioma sono tali che i grappoli ricevono sufficiente luce di moderata intensità, si assite ad maggiore biosintesi di questi composti (Haselgrove et al. 2000). Berqvist et al (2001) nei loro studi hanno trovato un contenuto maggiore di antociani in grappoli esposti alla luce. Anche in questo caso i grappoli esposti dal trattamento di sfogliatura hanno incrementato il livello di antociani rispetto al controllo, ed in particolare il valore più alto si è registrato nelle uve maturate su piante sfogliate all’allegagione, questo perché sfogliare dopo l’invaiatura vuol dire sottoporre il tessuto della buccia improvvisamente ad un regime radiativo e termico che non è abitutato a sostenere, a differenza dei grappoli di piante sfogliate all’allegagione (Palese 2003).
Conclusioni
Nella attuale tecnica viticola si presta molta attenzione agli effetti del microclima del grappolo sulla composizione dell’uva e sulla qualità del vino, e in particolare agli interventi che consentono di modulare l’esposizione dei grappoli. Il quantitativo di radiazione solare che raggiunge le foglie ed i grappoli interni alla chioma decresce in proporzione con l’aumento degli strati fogliari. Chiome dense, ombreggiate, costituiscono un ambiente avverso per la produzione di uva di qualità, poiché caratterizzate da: una riduzione della quantità della luce, una qualità dello spettro luminoso meno adatto alla maturazione dei grappoli, ridotta ventilazione ed elevata umidità e quindi maggiore incidenza di malattie fungine favorite anche dalla scarsa penetrabilità dei pesticidi. La sfogliatura è una pratica che permette di ridurre la densità della chioma e l’ombreggiamento, migliorando l’esposizione dei grappoli alla luce e favorendo l’efficacia d’azione dei pesticidi. La luce è un fattore determinante per la produzione di uve di qualità, perché influenza la biosintesi degli zuccheri, dei polifenoli, degli aromi, l’acidità totale el pH.
Nel presente lavoro di tesi uno degli obiettivi è stato quello di determinare se l’asportazione di 6 foglie alla base del germoglio uvifero, effettuata a stadi fenologici differenti, cioè all’allegagione e all’invaiatura, su due gruppi distinti di piante di Ciliegiolo, ha avuto eventuali effetti benefici sulla composizione delle uve, nell’ottica di valorizzare vitigni autoctoni come il Ciliegiolo. Infatti, un mercato sempre più omologato e competitivo, la ricerca della tipicità e la valorizzazione di vitigni autoctoni come il Cilegiolo, può sicuramente costituire una carta vincente da spendere nei confronti del consumatore.
Le indagini sono state concentrate su componenti della qualità, come i carotenoidi, precursori degli aromi C-13 norisoprenoidi che hanno un ruolo importante nella definizione dell’aroma del Ciliegiolo, ed i polifenoli. Tramite HPLC nelle uve sono stati determinati i carotenoidi (β-carotene, α-carotene, luteina, neoxantina, anteraxantina, violaxantina, zeaxantina) e le clorofille A e B. Oltre ai carotenoidi come precursori aromatici, è possibile che anche dalla degradazione delle clorofilla si originino composti volatili, ipotesi che non è stata ancora verificata e che rappresenta un’interessante linea di ricerca per il futuro. Il cambiamento del contenuto di carotenoidi e clorofille si verifica principlamnete durante invaiatura; l’effetto della luce sulla sintesi e degradazione dei precursori aromatici è di grande importanza: la sfogliatura precoce, che espone il grappolo durante il periodo di sintesi di questi composti, è risultata essere dannosa per la formazione del potenziale aromatico, in quanto i grappoli sono stati esposti ad un stress luminoso eccessivo che ha determinato una riduzione nel contenuto in carotenoidi totali; mentre la sfogliatura all’invaiatura non ha generato differenze rispetto al controllo. Questi dati sono importanti per capire il metabolismo dei carotenoidi e per promuovere la sintesi di questi composti, che sono degli interessanti precursori d’aroma; inoltre ci confermano l’importanza del fattore luce e della gestione della chioma per la qualità dell’uva, considerando che la sfogliatura, sia effettuata all’allegagione che all’invaiatura, ha anche generato uve con un contenuto maggiore di polifenoli e di antociani delle bucce rispetto al controllo.
Un altro obiettivo del presente lavoro di tesi è stato monitorare gli effetti della sfogliatura sulla crescita dell’acino. La gestione del rapporto foglie/frutti è alla base di molte pratiche colturali che aspirano ad ottenere una produzione di uva con determinate caratteristiche composizionali. La sfogliatura modifica la relazione source:sink, importante per raggiungere un equilibrio tra la crescita vegetativa e la crescita riproduttiva (equilibrio vegeto-produttivo), in modo da ottenere una adeguata composizione dell’uva. Per le varietà da vino, specialmente le rosse, le dimensioni dell’acino e la qualità del mosto sono strettamente correlate dal contenuto in acqua e dal rapporto buccia/polpa durante la vinificazione. Nel presente lavoro la sfogliatura non ha provocato nessuna differenza statistica tra le tesi considerate riguardo al peso medio dell’acino, nemmeno, come ci si aspettava, nel caso della sfogliatura precoce. Si possono formulare alcune ipotesi al riguardo, tra cui un aumento del tasso di fotosintesi delle foglie restanti o una intensificazione nella traslocazione
degli assimilati dagli organi di riserva che ha sopperito alla mancaza si assimilati dovuta all’asportazione delle foglie.
E’ in ogni caso da rilevare come il peso medio dell’acino proveniente da piante sfogliate all’allegagione sia sempre inferiore al controllo, anche se questo non è risultato statisticamente significativo, tendenza che, però, congiuntamente all’aumento della concentrazione dei polifenoli delle bucce, lascia intendere come la pratica della sfogliatura precoce possa essere idonea a valorizzare le potenzialità qualitative del vitigno Ciliegiolo, ma che essa debba essere eseguita in sinergia con altre pratiche viticole che permettano di salvaguardare il potenziale aromatico, altrimenti compromesso da una esposizione alla luce troppo intensa. Ulteriori ricerche sono necessarie per confermare questa ipotesi e per verificare le possibilità di migliorare anche la qualità del vino prodotto.