Capitolo
1
Stress anossico
1.1 Metabolismo anaerobico
Le piante, in quanto organismi sessili, sono frequentemente sottoposte a stress, cioè a condizioni esterne che influiscono sfavorevolmente sulla crescita, lo sviluppo o la produttività. Gli stress possono essere di natura biotica, dovuti cioè ad altri organismi, oppure di natura abiotica, risultanti da un eccesso o da una carenza dei fattori che determinano le caratteristiche chimico/fisiche dell’ambiente. Nelle piante gli stress inducono un’ampia gamma di risposte, da modificazioni dell’espressione genica e del metabolismo cellulare a cambiamenti delle velocità di crescita e delle relative biomasse prodotte. La durata, l’intensità con cui uno stress colpisce una pianta sono fattori che influenzano la risposta della pianta stessa; la combinazione di diverse condizioni sfavorevoli può provocare una risposta diversa da quella che si ha in seguito ad un singolo tipo di stress. Le caratteristiche della pianta, comprese l’identità degli organi e dei tessuti, lo stadio di sviluppo ed il genotipo, influenzano anch’essi le risposte della pianta allo stress.
I meccanismi di resistenza allo stress possono essere raggruppati in due categorie generali: meccanismi di allontanamento, che prevengono l’esposizione allo stress, e meccanismi di tolleranza, che permettono alla pianta di sopportare le condizioni sfavorevoli. La resistenza può essere ottenuta attraverso degli adattamenti, perfezionamenti evolutivi che aumentano il successo adattativo di una popolazione,
oppure attraverso l’acclimatazione, cioè mediante una risposta ai cambiamenti ambientali da parte dei singoli organismi, che modificano la propria omeostasi, cioè il proprio equilibrio fisiologico (Buchanan et al., 2003).
I cambiamenti del metabolismo e dello sviluppo indotti dallo stress possono essere attribuiti alla modificazione dei profili dell’espressione genica; una risposta ad una condizione sfavorevole inizia quando la pianta riconosce lo stress a livello cellulare: tale riconoscimento attiva le vie di trasduzione del segnale che trasmettono l’informazione all’interno delle singole cellule e in tutta la pianta. I cambiamenti dell’espressione genica sono integrati in una risposta da parte dell’intera pianta che può modificare la crescita e lo sviluppo ed influenzare anche le capacità riproduttive: la regolazione di tale risposta coinvolge ormoni (specialmente l’acido abscissico, l’acido jasmonico e l’etilene) e secondi messaggeri come il Ca2+. Alcuni geni sono espressi maggiormente mentre altri sono repressi: i prodotti proteici dei geni indotti spesso si accumulano, ma tale accumulo è influenzato anche da meccanismi di regolazione post-trascrizionale che aumentano le quantità di specifici mRNA, aumentano la traduzione, stabilizzano le proteine, modificano l’attività proteica o determinano un insieme di questi livelli di regolazione.
Tra gli stress abiotici, sono soprattutto le condizioni anaerobiche che comportano un rapido cambiamento nell’espressione genica, portando alla produzione di una serie di proteine coinvolte nella risposta adattativa alle basse concentrazioni di ossigeno (ANPs, “anaerobic polypeptides” o ASP, “anaerobic stress proteins”) (Geigenberger, 2003). Questo tipo di risposta è stato caratterizzato in diverse specie vegetali, tra cui alcune graminacee, come Echinocloa phyllopogon, mais, riso, piante anfibie, come Iris pseudacorus, e piante semi-acquatiche come la specie Rumex (Visser et al., 2003). A questo elenco di specie vegetali va aggiunto l’utilizzo negli ultimi anni di Arabidopsis thaliana come pianta modello per studiare l’adattamento alle basse concentrazioni di ossigeno (Dolferus et al., 2003).
Come la maggior parte degli organismi eucariotici, le piante sono aerobi obbligati, per cui l’ossigeno rappresenta l’accettore finale degli elettroni nella catena di trasporto mitocondriale.
Una diminuzione dell’ossigeno comporta rapide e profonde modificazioni sulla fisiologia della cellula. Questa condizione altera l’espressione genica, il consumo energetico, il metabolismo cellulare, la crescita e lo sviluppo. Si tratta generalmente di situazioni estreme dovute ai cambiamenti delle condizioni ambientali (ad esempio
la sommersione durante stagioni particolarmente piovose) o ad altre situazioni esterne come la compattazione dei suoli o l’attività di microrganismi presenti nel terreno (Drew, 1997; Geingeberger, 2003; Gibbs e Greenway, 2003; Greenway e Gibbs, 2003). In altri casi la bassa disponibilità di ossigeno può essere dovuta ad un rapido aumento del metabolismo cellulare, ad esempio durante l’imbibizione dei semi e la germinazione oppure durante la maturazione dei frutti (Bailey-Serres e Chang, 2005).
Anche in situazioni di ambienti ben ossigenati (21% v/v di ossigeno esterno), i tessuti vegetali che presentano un’elevata attività metabolica possono divenire ipossici; questo soprattutto quando mancano di ampi spazi intercellulari per il passaggio dell’aria, contengono cellule che sono poco vacuolate, o si trovano localizzati al centro di organi che sono distanti dai siti dove l’ossigeno entra nella pianta. Esempi di tessuti che presentano un elevato e rapido consumo di ossigeno e pochi spazi intercellulari sono le cellule meristematiche radicali (Ober e Sharp, 1996) e i tessuti floematici (van Dongen et al., 2003), al cui interno si riscontrano condizioni ipossiche. Stessa condizione è stata riscontrata all’interno delle silique di
Arabidopsis e nei semi di Vicia e Pisum, così come in organi di accumulo voluminosi
come mele (Magness, 1920), banane (Banks, 1983) e durante la crescita dei tuberi di patata (Geigenberger et al., 2000).
Nelle normali condizioni aerobiche le piante possono ossidare una mole di zuccheri esosi attraverso la glicolisi, il ciclo dell’acido citrico e la fosforilazione ossidativa fino a ottenere dalle 36 alle 38 moli di ATP. In assenza di ossigeno la produzione mitocondriale di ATP mediante la via della fosforilazione ossidativa risulta inibita. Inoltre i nucletotidi pirimidinici ridotti nel corso della via glicolitica e del ciclo di Krebs non possono più essere riossidati attraverso la catena mitocondriale di trasporto degli elettroni, per cui, per far procedere la glicolisi in assenza di ossigeno, deve essere rigenerato il substrato glicolitico NAD+.
Riguardo alle modalità del riciclaggio anaerobico dei nucleotidi pirimidinici sono state proposte diverse vie, ma la via fermentativa sembra giocare un ruolo fondamentale (Perata e Alpi, 1993).
Il lattato e l’etanolo sono i principali prodotti finali della via fermentativa, da cui si possono formare anche altri prodotti secondari come l’alanina, il succinato e il
relazione alle specie vegetali, al genotipo e al tessuto, così come alla durata e all’intensità della mancanza di ossigeno.
Sia la via fermentativa che porta al lattato sia quella che porta alla formazione di etanolo producono NAD+, ma solo il lattato abbassa il pH citosolico. In accordo con l’ipotesi di Davies-Roberts sull’omeostasi del pH basata sul rapporto tra lattato deidrogenasi (LDH) e piruvato decarbossilasi (PDC), il metabolismo anaerobico viene regolato dalle attività di enzimi sensibili al pH. In base a questo modello il piruvato prodotto inizialmente dalla glicolisi viene convertito a lattato tramite l’azione della LDH, un enzima che ha come pH ottimale il pH fisiologico. La produzione di lattato riossida il NADH e al tempo stesso abbassa il pH citoplasmatico. Con l’acidificazione del citosol, l’LDH viene progressivamente inibito e viene attivato un secondo enzima, la PDC, per il quale il pH ottimale è più basso di quello che normalmente si ha nel citoplasma e che stimola la conversione di piruvato ad acetaldeide. L’alcol deidrogenasi (ADH) riduce successivamente l’acetaldeide a etanolo, ossidando nello stesso tempo il NADH a NAD+. Come risultato il pH citoplasmatico viene stabilizzato su un valore leggermente acido. Dunque l'accumulo di acido lattico con la conseguente acidificazione del citosol potrebbe fornire il segnale che innesca la fermentazione alcolica. Questa osservazione è stata supportata dal fatto che in molte piante in anossia la produzione di lattato precede la sintesi di etanolo (Perata e Alpi, 1993).
Fig.1.1:meccanismi biochimici di rigenerazione del NAD+ nei tessuti vegetali.
Sulla regolazione del metabolismo fermentativo sono comunque ancora aperti diversi dibattiti, ma non vi sono dubbi sul fatto che la fermentazione avvenga nei tessuti delle piante esposte all’anossia. Un’ipotesi diversa sostiene che siano le
concentrazioni di piruvato, piuttosto che il pH citoplasmatico, a regolare la fermentazione etanolica, anche se le due ipotesi sostanzialmente non si escludono a vicenda.
Pur essendo provata la tossicità dell’etanolo sulle piante, tale prodotto finale non può essere considerato una causa primaria per i danni da anossia. Piante tolleranti, secondo Crawford (1978), limitano la quantità di etanolo prodotto in due modi: non producendolo, o rimuovendolo per diffusione dalle radici o con trasporto in altre parti della pianta. Specie intolleranti la sommersione mostrano invece un’accelerazione della via glicolitica con conseguente accumulo nei tessuti di etanolo. Questo tuttavia non è valido nel caso del riso, la cui capacità di germinare e di allungare il coleoptile in condizioni di anossia è dovuta ad una produzione piuttosto elevata di etanolo e non ad un abbassamento della sua concentrazione accumulata. Jackson e collaboratori (1982), sulla base di risultati sperimentali, conclude che l’etanolo non svolge il ruolo fondamentale nell’induzione dei danni relativi all’anossia. Solo elevate concentrazioni di tale prodotto, non fisiologiche e fornite quindi esogenamente, risultano essere estremamente tossiche per la pianta.
Studi condotti da Roberts e collaboratori (1984b) hanno dimostrato che mutanti di mais ADH1-null, mancanti cioè di un enzima che non è necessario per la crescita in condizioni aerobiche, sono più sensibili in ipossia di quanto non lo siano i genotipi
wild-type isogenici. Il mutante mostra accumulo di acetaldeide ed un prolungato
abbassamento del pH citoplasmatico rispetto al wild-type attribuiti ad una continua produzione di lattato. Il livello di ATP in questi mutanti, in condizioni di ipossia, è molto basso in quanto la fermentazione lattica non è in grado di fornire le condizioni necessarie per una efficiente produzione di ATP attraverso la fermentazione. Roberts conclude affermando che la sensibilità del mutante in ipossia sia dovuta alla sua inabilità nella regolazione del pH citoplasmatico. Sempre in questi studi (Roberts et
al., 1984a) è stato dimostrato che l’acidificazione citoplasmatica sia dovuta alla
perdita di acidi dal vacuolo a causa della minore disponibilità di molecole di ATP, non sufficiente a mantenere il gradiente protonico del tonoplasto attraverso la pompa H+-ATPasi.
La riduzione della concentrazione interna di ossigeno comporta una generale diminuzione dell’attività metabolica. Attraverso studi condotti su tuberi di patata (Geigenberger et al., 2000), semi di Arabidopsis (Rolletscheck et al., 2002) e nei
tessuti floematici di piante di Ricinus (van Dongen et al., 2003) è stato riscontrato un abbassamento del rapporto ATP/ADP, indicando l’inibizione della respirazione.
Geigenberger e collaboratori hanno dimostrato in patata e Ricinus che la riduzione del rapporto ATP/ADP che accompagna l’abbassamento della concentrazione interna di O2, non è associata ad un incremento dello stato redox
cellulare (ad esempio rapporto NADH/NAD+), almeno fin quando i valori di O2 non
scendono sotto l’1%. Per cui se il sistema NAD+ citosolico rimane altamente ossidato mentre [O2]int diminuisce, allora significa che il metabolismo aerobico è mantenuto
anche in un ampio range di [O2] nei diversi tessuti. Inoltre la riduzione dello stato adenilato non è associata con l’accumulo di lattato o etanolo (Geigenberger et al., 2000; Rolletscheck et al., 2002; van Dongen et al., 2003) e il suo valore è già significativo a [O2]int che sono superiori alla Km della citocromo ossidasi (Km [O2]cyt ox
= 14 µM, equivalente a 0,013% di ossigeno) (Drew, 1997).
Dunque si può riassumere affermando che la riduzione della [O2] percepita dai sistemi vegetali comporta una rapida inibizione della respirazione, una diminuzione dello stato adenilato e una coordinata diminuzione dell’attività glicolitica e del ciclo di Krebs. Il primo sito di regolazione per queste risposte è la catena di trasporto elettronico del mitocondrio. L’inibizione avviene comunque quando la concentrazione di ossigeno si trova al di sopra della Km della citocromo ossidasi, per cui l’ossigeno non limita direttamente la catena di trasporto elettronico. La conseguente riduzione del consumo di ossigeno è adattativa, in quanto consente alle piante di evitare che i tessuti interni diventino anossici (Geigenberger et al., 2003).
Conducendo sempre studi su tuberi di patata è stato dimostrato (Geigenberger et
al., 2003) che la riduzione della [O2] (in un range da 21% fino a 0%) comporta una progressiva inibizione dei processi biosintetici che portano alla sintesi di saccarosio, amminoacidi, proteine e lipidi, durante le prime due ore di trattamento. Ciò è accompagnato da una concomitante riduzione del consumo di ATP quando appunto la respirazione viene inibita. Risulta invece interessante il fatto che la sintesi di amido (Geigenberger et al., 2000) rimanga relativamente elevata se confrontata con gli andamenti della altre vie biosintetiche. Per quanto riguarda invece la degradazione degli zuccheri, questa subisce una sensibile riduzione quando la concentrazione di ossigeno scende al di sotto del 12%, mentre subisce un’ulteriore incremento quando si trova allo 0%, semplicemente perché questo segna l’inizio del processo fermentativo.
Da notare inoltre che le condizioni ipossiche inibiscono le risposte allo stress da
wounding, una tipica risposta cellulare che richiede appunto ampi incrementi delle
attività metaboliche (Butler et al., 1990; Geigenberger et al., 2000).
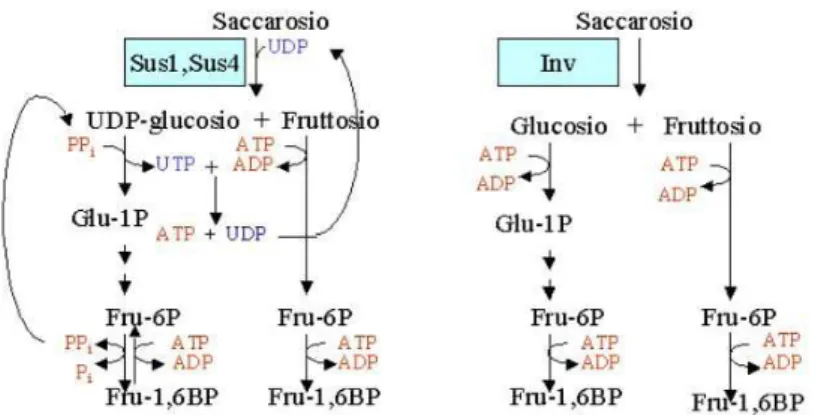
Una strategia complementare alla riduzione del metabolismo potrebbe essere quella che favorisce vie metaboliche e strategie in grado di conservare energia, e che quindi ridurranno anche il consumo di ossigeno (Geigenberger et al., 2000). Un esempio è la degradazione del saccarosio. Le piante hanno due vie biochimiche alternative per la degradazione del saccarosio, che differiscono soltanto per il loro costo energetico per ottenere esosi fosfati. La degradazione del saccarosio tramite invertasi e esochinasi richiede due molecole di ATP, mentre tramite la saccarosio sintasi e la UDP glucosio fosforilasi richiede una sola molecola di pirofosfato inorganico (PPi) e non ATP. Dunque il costo energetico attraverso la saccarosio sintasi è minore, in considerazione anche del fatto che questa via ricicla PPi, prodotto da diverse reazioni biosintetiche.
Fig. 1.2: Confronto dei primi passaggi della via glicolitica mediati da saccarosio sintasi e
invertasi
I geni codificanti per l’enzima saccarosio sintasi (SuSy genes) risultano essere particolarmente indotti dalle condizioni di ipossia mentre i geni codificanti per l’enzima invertasi sono fortemente repressi (Geigenberger et al., 2003).
Essendoci una forte induzione per i geni della saccarosio sintasi, in particolare dei geni SUS1 e SUS4 (Loreti et al., 2005) anche in plantule di Arabidopsis, questo suggerisce che la via della saccarosio sintasi sia attiva per l’utilizzo di saccarosio in condizioni di anossia. Questo era già stato proposto per il riso (Guglielminetti et al., 1995).
Per quanto riguarda invece la via di degradazione dell’amido, sempre in plantule di Arabidopsis, questa viene moderatamente indotta in anossia, indicando quindi che, in termini di conservazione di energia, viene seguita la via degradativa del saccarosio e non dell’amido.
Risulta molto interessante notare che, mentre l’espressione dei geni codificanti per proteine coinvolte nel ciclo di Krebs non subisce variazioni durante l’anossia, un ampio numero di geni codificanti per proteine appartenenti alla catena di trasporto mitocondriale, soprattutto NADH deidrogenasi, risulti indotta in anossia. Questo potrebbe essere dovuto al fatto che bassi livelli di ATP potrebbero portare ad un
up-regulation dei geni chiave per la produzione di energia, includendo la catena di
trasporto mitocondriale. Questa risposta potrebbe essere poco utile durante l’anossia, sebbene i mitocondri necessitino di maggiore tempo per diventare anossici, mentre potrebbe giocare un ruolo importante nell’adattamento all’ipossia e alla sommersione (Loreti et al., 2005).
1.2 Ciclo Hb/NO come meccanismo diretto di
percezione della carenza di ossigeno
L’assenza di ossigeno altera lo stato redox della cellula, portando ad esempio all’aumento del rapporto NADH/NAD+ dovuto proprio all’incapacità dei mitocondri ad usare NADH in anossia. Sebbene l’attivazione della fermentazione alcolica potrebbe in qualche modo ridurre l’effetto di un eccessivo accumulo di NADH, anche l’attivazione di altri geni in grado di compensare questo squilibrio dello stato redox potrebbe rivelarsi molto importante. Fra questi l’emoglobina non-simbiotica (Hb1) è quella maggiormente indotta e un suo ipotetico ruolo fisiologico potrebbe essere quello di funzionare come un fattore, NADH-dipendente, ossidante l’ossido nitrico (NO), con produzione di nitrato che viene poi convertito a nitrito dalla nitrato riduttasi (NR1) (vedi figura 1.3). Studi recenti (Dordas et al., 2004) hanno proprio dimostrato come NO sia prodotto nei tessuti vegetali in condizioni ipossiche e che l’emoglobina di classe 1 svolgano un ruolo essenziale nella regolazione dei livelli di NO. Dunque anche il sistema Hb/NR1 svolge l’importante funzione di rigenerazione di NAD+ in condizioni anossiche come supporto per la glicolisi. L’incremento dell’espressione del gene Hb1 in seguito alla interruzione della sintesi di ATP a
livello mitocondriale, supporta l’ipotesi che l’emoglobina possa svolgere un ruolo nella mitigazione degli effetti dello stress anossico (Nie e Hill, 1997). Questa considerazione è stata inoltre confermata dal fatto che piante overesprimenti Hb mostrano una bassa attività dell’ADH, suggerendo appunto che l’emoglobina possa essere un’alternativa alla fermentazione (Dat et al., 2004).
Uno dei problemi fondamentali è come gli organismi vegetali siano in grado di percepire e di trasmettere il segnale in caso di abbassamento dei livelli di O2. In una ipotetica via di percezione dei livelli di O2 vi deve essere sicuramente a monte un sensore in grado di percepire le variazioni della concentrazione di ossigeno nell’ambiente circostante, instaurando un meccanismo di trasduzione del segnale a cascata verso specifici fattori di trascrizione i quali medieranno a loro volta la risposta cellulare. L’identificazione di tali ipotetici sensori e l’esatto meccanismo di percezione restano comunque ancora indefiniti. Nelle piante sono stati identificati 2 tipi di emoglobine: una simbiotica (leghemoglobina), l’altra non simbiotica. La prima è stata isolata inizialmente in specie leguminose ed è presente soprattutto a livello dei noduli dove contribuisce appunto al trasporto di ossigeno. Al contrario la funzione di quella non simbiotica (Hb) non è ancora stata definita (Chaparro-Giraldo
et al., 2000; Duff et al., 1997) sebbene sia noto che venga indotta in tessuti sottoposti
a stress ipossici (Taylor et al., 1994).
Sono stati proposti diversi ruoli per Hb in condizioni ipossiche:
• può svolgere la funzione di trasportatore dell’O2 come nelle cellule animali per aiutare a preservare la respirazione mitocondriale in condizioni anossiche;
• può agire come proteina in grado di trasferire elettroni;
• può agire come sensore dell’ossigeno in grado di regolare l’espressione
genica in condizioni anossiche o di sostenere il metabolismo glicolitico nei tessuti stressati.
L’induzione di emoglobina (Hb) è stata studiata in condizioni ipossiche per il suo coinvolgimento nell’acclimatazione delle piante a bassi livelli di ossigeno. Come abbiamo visto sopra, uno dei suoi principali ruoli consiste nel modulare l’ossido nitrico (NO) prodotto in risposta a stress ipossici. La molecola NO è coinvolta in molte vie di trasduzione del segnale in connessione con risposte da stress, e inoltre la modulazione dei livelli di NO può portare a significativi cambiamenti nelle risposte ormonali delle piante.
Esiste un efficiente sistema NADPH e Hb-dipendente per lo scavenging di NO in condizioni ipossiche.
In ipossia, il nitrato è utilizzato come accettore di elettroni intermedio. L’emoglobina potrebbe essere un componente di una ossigenasi solubile per NO (probabilmente citosolica), che contribuisce all’ossidazione del NADH ed è associata al miglioramento dello stato energetico cellulare, indipendentemente dalla fosforilazione ossidativa mitocondriale.
Fig.1.3 (Dordas et al., 2003): meccanismi di interazione tra Hb e NO (Hb/NO cycle) che
portano alla rigenerazione del NADH e al mantenimento dello status energetico. NR, nitrato riduttasi; E, ipotetico complesso che porta alla formazione di NO costituito da nitrito:NO riduttasi, NO sintasi e mitocondrio. NO viene ossidato a nitrato per mezzo della oxy-emoglobina [Hb(Fe2+)O
2]. L’alta affinità tra Hb(Fe2+) e O2 determina una immediata
ossigenazione anche a concentrazioni (nanomolari) molto basse di O2. La riduzione del
nitrito a NO nel mitocondrio è invece associata all’ossidazione del NADH tramite il complesso I, essendo un ipotetico sito per la sintesi di ATP.
L’ossido nitrico è un segnale ubiquitario negli organismi vegetali dove le forme inorganiche di azoto ossidato rappresentano una risorsa per la formazione di NO. In determinate condizioni NO può essere prodotto attraverso riduzioni enzimatiche e non enzimatiche di nitrato e nitrito. Tuttavia la formazione non-enzimatica è favorita a valori di pH acidi, per cui la sua formazione nell’apoplasto dipenderà essenzialmente dal pH acido e dalla presenza di nitrito extracellulare, condizioni che
sono appunto presenti durante l’anaerobiosi. L’enzima chiave che sfrutta il nitrato come substrato per la formazione di NO è una nitrato riduttasi citosolica (vedi fig.1.3) (Rockel et al., 2002) ed è stato interessante osservare che il nitrito può essere accumulato a livello intracellulare solo quando le cellule sono in condizioni anaerobiche, condizione tipica delle cellule radicali in condizioni di sommersione (Botrel et al., 1996). Stohr e collaboratori hanno identificato la nitrito:NO riduttasi localizzata sulla membrana citoplasmatica responsabile della generazione di NO nelle radici (Stohr et al., 2001).
L’ossido nitrico risulta essere coinvolto in reazioni con radicali liberi, in legami reversibili con metallo proteine e la sua formazione nelle radici è stimolata soprattutto da condizioni anaerobiche. Il fatto che questa molecola sia associata all’emoglobina e all’omeostasi dello stato redox, potrebbe far supporre che NO possa essere uno dei primi segnali nella percezione della diminuzione di ossigeno, come ad esempio durante periodi di sommersione (Dordas et al., 2003).
1.3 Il flusso del Ca
2+è necessario per l’induzione dell’ADH in
assenza di O
2Nelle specie vegetali come mais, riso e Arabidopsis, il flusso di Ca2+ è un prerequisito per l’alterazione dell’espressione di alcuni geni in risposta all’anossia e all’ipossia (Subbaiah e Sachs, 2003). Per comprendere l’importanza della modulazione di Ca2+, Sedbrook e collaboratori (Sedbrook et al., 1996) hanno osservato plantule di Arabidopsis in grado di esprimere una proteina luminescente sensibile al calcio, una AQUORINA di medusa, ed hanno osservato tre oscillazioni della concentrazione di Ca2+ citosolico in risposta all’anossia. In particolare il trasferimento delle plantule in condizioni anossiche causa, dopo appena 10 minuti, un rapido picco transiente del livello di Ca2+cit seguito da un prolungato incremento tra 1.5h e 4h di esposizione. Anche la riossigenazione è in grado di indurre un picco transiente nella [Ca2+]cit.
Il picco iniziale individuato risulta molto meno accentuato se le plantule vengono pre-trattate o trattate ripetutamente durante gli iniziali 10 minuti di anossia con rutenio rosso, con un chelante per il calcio (EGTA), o con lantanio o cloruro di gadolinio entrambi in grado di bloccare i canali per il calcio situati sulla membrana
citoplasmatica. Durante invece il prolungato incremento di [Ca2+] cit sia il lantanio che il cloruro di gadolinio hanno in questo caso un effetto positivo sull’incremento.
Queste osservazioni consentono di ipotizzare che l’incremento iniziale di calcio sia dovuto al rilascio di calcio da riserve interne al citoplasma, mentre l’incremento successivo sembra possa essere dovuto alle attività dei canali situati lungo la membrana citoplasmatica (Sedbrook et al., 1996).
Un altro dato interessante osservato dal gruppo di lavoro di Sedbrook è che le radici non mostrano alcun incremento di [Ca2+] cit , per cui è assai probabile che radici e tessuti aerei della pianta possano avere una risposta distinta oppure l’AQUORINA di questi sistemi non possiede un’adeguata sensibilità per registrare eventuali modulazioni di [Ca2+] cit in questi tessuti (Sedbrook et al., 1996). Questa seconda ipotesi sembra essere più corretta e confermata dal fatto che l’incremento di [Ca2+] cit sia necessario per guidare l’espressione del promotore di ADH1 sia nelle radici che nei tessuti aerei di A. thaliana (Chung e Ferle, 1999).
Alla luce di questi risultati sarà importante determinare lo specifico ruolo di Ca2+ nella trasduzione del segnale in condizioni sia ipossiche che anossiche.
1.4 La percezione dell’etilene influenza positivamente la
risposta alla carenza di O
2Fra gli ormoni studiati in risposta a stress da carenza di ossigeno, l’etilene è stato indubbiamente quello che ha suscitato maggiore interesse. In diverse specie vegetali è stato infatti osservato che la biosintesi dell’etilene incrementa nelle prime fasi (4 ore) di esposizione a trattamenti ipossici (Drew et al., 1979; Lorbiecke e Sauter, 1999). In Arabidopsis la fosforilazione dell’enzima acido 1-aminociclopropano-1-carbossilico sintasi (ACS) dovuta alla proteina kinasi MAPK6 comporta un accumulo della proteina ACS (Liu e Zhang, 2004) per cui, essendo elevata l’attività dell’enzima ACC sintasi, risulterà elevata anche la produzione di etilene. Ovviamente la biosintesi di etilene non potrà avvenire se la cellula si trova in condizioni strette di anossia, in quanto la conversione di ACC a etilene da parte dell’enzima ACC ossidasi (ACO) richiede consumo di ossigeno (Yang e Hoffman, 1984; Kende, 1993). Non è comunque ancora noto se la via di trasduzione del segnale da parte di MAPK venga attivata dalla diminuzione dei livelli di ossigeno.
Nella specie Rumex palustris è stato visto che, in condizioni di ipossia, il livello di concentrazione dell’etilene subisce una immediata variazione, tanto che dopo 16 ore di sommersione è possibile osservare un incremento di questi valori, da 0,5µl l-1 iniziali fino a 8 µl l-1 (Banga et al., 1996).
L’etilene è coinvolto in numerose risposte di acclimatamento a condizioni di sommersione e quindi di ipossia, portando ad esempio all’allungamento degli internodi e alla formazione di radici avventizie in riso durante la sommersione Lorbiecke e Sauter, 1999), allungamento del picciolo e movimento iponastico delle foglie in R. palustris (Voesenek et al., 2003).
In Arabidopsis l’etilene è in grado di incrementare l’induzione ipossica del gene
ADH1 (Peng et al., 2001).
1.5 Variazioni nella omeostasi delle “reactive oxygen
species” (ROS) nella percezione della carenza di O
2I mitocondri sono per loro natura estremamente sensibili alle variazioni dei livelli di O2 nell’ambiente, in quanto a livello cellulare ne sono i principali consumatori e
rivestono un ruolo cruciale nel percepire la sua variazione (Dat et al., 2004). Infatti, l’O2 è l’accettore finale del trasporto elettronico nella catena respiratoria, un processo
da cui si originano anche le cosiddette specie reattive dell’ossigeno (ROS) come co-prodotti di questo metabolismo.
L’importanza del ruolo delle ROS nella percezione della variazione dei livelli di O2 non è stata ipotizzata esclusivamente per i mitocondri, ma altri componenti
cellulari in grado di formare ROS sono stati identificati come ipotetici sensori di stati ipossici. L’incremento transiente nel livello del trascritto di una respiratory burst oxidase in tessuti radicali di Arabidopsis durante trattamenti ipossici (Klok et al., 2002) e l’induzione di ADH dipendente dall’attività di una NADPH ossidasi, generatrice di ROS, sono altri esempi dell’attualità del dibattito sull’importanza delle ROS nella percezione di bassi livelli di concentrazione di O2.
Baxter-Burrel e collaboratori (2002) hanno osservato che la trasduzione del segnale anossico è, almeno in parte, mediata da una proteina in grado di legare il nucleotide guanidinico trifosfato (GTP), la RopGAP4 (Rop GT-Pase activating protein 4), che regola l’attività di una NADPH ossidasi. Nelle piante esistono due
classi di tali peptidi: le proteine G eterotrimeriche, e le Rop monometriche. Entrambe le classi sembrano svolgere ruoli chiave nell’adattamento alla carenza di O2: ad esempio é stato dimostrato che un attivatore di sintesi della segnalazione mediata da proteine, il guanosin-5’-O-(3-tiotrifosfato) (GTPγS), promuove la formazione di
aerenchima mediante meccanismi apoptotici in radici di mais (Drew et al., 2000). Le Rop fungono da interruttori molecolari, attivi nello stato legato al GTP e inattivi se legate al GDP, in grado di scatenare una catena di segnalazione o di attivare, in maniera più o meno diretta, alcuni enzimi (Yang e Poovaiah, 2002).
Il lavoro condotto da Buxter-Burrel e collaboratori risulta particolarmente interessante in quanto riporta in Arabidopsis l’aumento dell’espressione di una
RopGAP4 in condizioni di anossia. Il mutante inserzionale per RopGAP4 mostra
inoltre livelli di messaggero per l’ADH significativamente superiori rispetto al wild
type. Valutando poi i livelli di H2O2, le quantità di messaggero e l’attività dell’enzima ADH in mutanti che esprimono forme costitutivamente attive o inattive di proteine Rop, gli autori hanno ipotizzato che l’espressione dell’ADH sia mediata dalla produzione di H2O2, riferibile all’attività di una NADPH ossidasi. Questa NADPH ossidasi risulterebbe a sua volta regolata da Rop e inibita da cloruro di iodio difenile (DPI), considerato un inibitore di proteine legate a gruppi flavinici quali le NADPH ossidasi.
Figura 1.4: Rappresentazione schematica del ruolo delle Rop nella trasduzione del segnale anossico.
Alla luce di questi studi è possibile speculare su una connessione tra la produzione di H2O2 dovuta all’attività delle proteine ROP e l’incremento di [Ca2+]cit dovuto, invece, ad uno stato ipossico cellulare (Bailey-Serres e Chang, 2005). Interessante è infatti notare che l’incremento di [Ca2+]cit sembra essere un prerequisito per l’attivazione di H2O2 a livello apoplastico, dovuta all’attività di una NADPH ossidasi Ca2+ dipendente (Keller et al., 1998; Sagi e Fluhr, 2001).
In radici di Arabidopsis, analisi di mutanti per una subunità della NADPH ossidasi di membrana mostrano che la produzione di ROS da parte di questa ossidasi attiva canali specifici per Ca2+ e ne facilita il flusso necessario per la crescita radicale (Foreman et al., 2003). Diverse analisi microarray (Klok et al., 2002; Branco-Price
et al., 2005) hanno individuato l’induzione di tale subunità in condizioni ipossiche.
Inoltre, sempre in Arabidopsis, è stato osservato che la proteina Rop1 è responsabile del gradiente intracellulare di Ca2+ a livello dei peli radicali (Jones et al., 2002). Questi dati possono far ipotizzare che le proteine Rop, mediate dall’attività di una NADPH ossidasi, presumibilmente localizzata sulla membrana plasmatica, siano in grado di promuovere un incremento del Ca2+ citosolico in condizioni ipossiche. Di conseguenza, il flusso di Ca2+ sembra provvedere a un bilanciamento tra la
produzione di H2O2 come molecola segnale e il danno che essa può causare, trattandosi di una specie reattiva dell’ossigeno (Bailey-Serres e Chang, 2005).
Fig 1.5 (Bailey_Serres e Chang, 2005): Modello di percezione e trasduzione del segnale a seguito di
carenza di O2: una diretta percezione del segnale non è ancora nota, mentre variazioni del pH
citosolico, di [Ca2+]
cit, produzione di ROS e riduzione dello stato adenilato possono rappresentare
meccanismi indiretti per la trasmissione del segnale. L’accumulo della forma attiva delle ROP GTPasi stimola in Arabidopsis l’induzione dell’espressione del gene dell’ADH e la fermentazione alcolica. L’attivazione delle ROP come molecole segnale comporta un incremento dei livelli di H2O2, per cui i
genotipi che non sono in grado di regolare negativamente questo segnale saranno maggiormente sensibili alle condizioni ipossiche, nonostante la forte induzione dell’espressione del gene ADH. RopGAP consente l’idrolisi di GTP legata alla proteina ROP e questa regolazione negativa limita la produzione di ROS e consente la conservazione dei carboidrati.