20
Capitolo 2
Modello biologico di riferimento: anatomia e biomeccanica del
braccio dell’Octopus vulgaris
Lo studio dell’ Octopus vulgaris è molto stimolante in quanto questo mollusco, nonostante la sua semplicità, presenta soluzioni a problemi ingegneristicamente complessi e funzionalità uniche, che lo rendono un ottimo candidato per uno studio biomimetico.
La morfologia del polpo rappresenta una delle caratteristiche che maggiormente ne rendono interessante lo studio e l’analisi. Infatti, gli infiniti gradi di libertà del braccio del polpo, che tanto appassionano i ricercatori, sono la conseguenza della totale assenza di strutture rigide. La conseguente adattabilità del braccio dell’Octopus vulgaris (mostrato in Fig.2.1), inoltre, rende più facile muoversi in ambienti strutturati di quanto non lo sia per gli animali vertebrati [1] e consente di usarne l'intera lunghezza per la manipolazione degli oggetti, non
Fig. 2.1 Octopus vulgaris, il lato del braccio caratterizzato dalla presenza delle ventose che si osserva in primo piano è denominato “orale”
21
restringendo tale possibilità al solo end-effector. L’altrettanto interessante capacità del polpo di allungare le proprie braccia fino al 60-70% [14] discende anch’essa dalla particolarissima morfologia delle strutture muscolari dell’animale.
2.1 Anatomia del braccio
La muscolatura intrinseca del braccio del polpo consiste di un denso array tridimensionale di fibre muscolari che circondano le fibre del sistema nervoso periferico posizionate al centro del braccio. Lungo le fibre del sistema nervoso si estende un’arteria dalle pareti spesse. Ci sono quattro gruppi di fibre nervose intramuscolari che si estendono lungo il braccio verso la periferia della parte più densa della muscolatura. Il braccio è ricoperto da un semplice epitelio che sovrasta lo spesso e fibroso tessuto connettivo. Tale tessuto contiene al suo interno dei foglietti che sono i siti d’origine e inserzione per gran parte della muscolatura intrinseca del braccio [14].
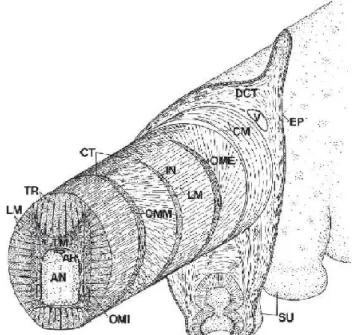
La muscolatura è disposta secondo tre orientamenti primari: le fibre muscolari trasversali sono orientate in piani perpendicolari all’asse maggiore del braccio, le fibre muscolari longitudinali sono orientate parallelamente all’asse maggiore del braccio e le fibre muscolari oblique sono disposte ad elica intorno al braccio come illustrato in Fig.2.2.
Fig. 2.2 schema della disposizione della muscolature del braccio del polpo. AN corrisponde al midollo spinale assiale, AR all’arteria, CM allo strato muscolare circonferenziale, CT al tessuto connettivo, DCT al tessuto dermale connettivo, EP all’epidermide, IN al nervo intramuscolare, LM alla fibre muscolari longitudinali,
22
OME allo strato muscolare obliquo esterno, OMI allo strato muscolare obliquo interno, OMM allo strato muscolare obliquo medio, SU alla ventosa, TM alle fibre muscolari trasversali, TR alle trabecule, V alla vena[1]. Citare fonti
La proporzione della sezione del braccio occupata da ognuno di questi gruppi di fibre muscolari (relativa all’area totale della sezione della muscolatura) rimane costante lungo il braccio anche se il braccio si restringe gradualmente fino ad arrivare alla punta distale [14].
La massa muscolare trasversale è principalmente concentrata nella parte centrale del braccio, intorno al midollo spinale centrale. Le fibre dei muscoli trasversali sono orientate perpendicolarmente all’asse longitudinale del braccio. Le trabeculae sono strutture della muscolatura trasversale che si originano nel tessuto connettivo nel lato orale (interno) ed aborale (esterno) del braccio e si estendono verso il centro del braccio in foglietti tra i muscoli longitudinali come è illustrato in Fig. 2.3.
23
Le fibre dei muscoli longitudinali circondano la parte centrale della muscolatura trasversale e sono orientate parallelamente all’asse longitudinale del braccio. Tali fibre si estendono in fasci tra le trabecole della massa muscolare trasversale. Le fibre muscolari longitudinali sono distribuite oralmente, aboralmente e lateralmente in modo che l’intero perimetro esterno del braccio include le fibre muscolari longitudinali. L’area della sezione delle fibre muscolari longitudinali è maggiore nel quadrante aborale del braccio che nei quadranti orale e laterali.
Oltre ai muscoli trasversali e longitudinali nel braccio del polpo sono presenti tre coppie di strati muscolari orientati obliquamente, uno esterno, uno intermedio ed uno interno. Per ciascuna coppia di muscolatura obliqua le fibre sono disposte con orientamento opposto: se da un lato le fibre seguono un orientamento levogiro, dal lato opposto (rispetto al piano di simmetria sagittale) seguono un avvolgimento destrogiro. Inoltre, l’orientamento alternato si nota anche tra i muscoli di un lato: per esempio, se da un lato i muscoli obliqui esterni sono
orientati in modo destrogiro, nello stesso lato i mediali sono disposti in modo levogiro e gli interni sono destrogiri come gli esterni[14].
È altresì presente anche un sottile strato di fibre muscolari disposte circolarmente; tale strato è più spesso nel lato aborale. Esso si estende lungo ogni lato e circonda gli strati muscolari obliqui esterni e si inserisce nello strato orale del tessuto connettivo.
Fig. 2.3 sezione trasversale del braccio del polpo nella quale si possono ritrovare tutti i componenti principali della muscolatura intrinseca del braccio. Il tessuto connettivo ha una colorazione rossastra ed il tessuto muscolare una colorazione marrone. Le lettere sulla foto hanno la corrispondenza: AN: midollo spinale assiale, AR: arteria, ACT: foglio di tessuto connettivo aborale, IN: nervo intramuscolare, CT: tessuto connettivo, OCT: foglio di tessuto connettivo orale, LM: fibre muscolari longitudinali, OME: strato muscolare obliquo esterno, OMI: strato muscolare obliquo interno, OMM: strato muscolare obliquo medio, TM: fibre muscolari trasversali, TR: trabecule. [1] Citare fonti
Fig. 2.4 disposizione delle fibre muscolari oblique e meccanica della torsione. La contrazione di una singola porzione di muscolo avvolto ad elica causa una torsione
24
L’area della sezione della muscolatura intrinseca del braccio e del midollo spinale della base è 144 volte maggiore dell’area della punta. La muscolatura longitudinale occupa il 56% dell’area totale della muscolatura intrinseca e le muscolature obliqua e trasversale occupano il 21% e 17% rispettivamente. Tali misure si riferiscono all’area occupata solo dalla muscolatura, quindi è esclusa l’area occupata dal midollo spinale [14].
La muscolatura sopra descritta si dimostra importantissima nel braccio del polpo dato che in esso non vi sono strutture ossee rigide e la loro funzione è svolta proprio dalla muscolatura. Le braccia e i tentacoli dei cefalopodi, infatti, fanno parte di una classe di strutture muscolari chiamate “muscolar hydrostat”. Queste strutture consistono primariamente di una disposizione tridimensionale delle fibre muscolari e quindi mancano le altre caratteristiche forme di supporto scheletrico osservabile in altri animali. Essi, infatti, non presentano la struttura scheletrica interna o esterna di artropodi, echinodermi e vertebrati, né presentano cavità ripiene di liquido circondate da un contenitore rinforzato da tessuto connettivo che caratterizza lo scheletro idrostatico di animali vermiformi.
Il principio per cui tali strutture sono capaci di muoversi e di rimanere compatte risiede nella morfologia stessa. Il tessuto muscolare, infatti, è un tessuto ad alta densità che quindi riesce a mantenere il proprio volume inalterato durante le deformazioni che subisce. Poiché il volume totale della struttura non varia, ogni diminuzione quantitativa in qualsiasi dimensione deve essere controbilanciata da un aumento quantitativo in un’altra dimensione. Nei “muscolar hydrostat” le fibre muscolari sono tipicamente orientate in più direzioni in modo che tutte le tre dimensioni possano essere controllate attivamente. Quindi, attivando selettivamente specifiche orientazioni delle fibre muscolari si può creare una grande varietà e complessità di movimenti [14].
2.2 Biomeccanica dei movimenti di base
Da un punto di vista biomeccanico tutti i movimenti complessi a cui ci si è riferiti sono prodotti dalla combinazione di cinque deformazioni base del braccio dell’ Octopus vulgaris: l’allungamento, l’accorciamento, il piegamento o bending, l’irrigidimento o stiffening e la torsione. Queste deformazioni possono essere localizzate o interessare l’intero braccio. Il movimento di bending può esser espletato in ogni direzione ed i movimenti torsionali
25
possono essere attuati sia in verso orario che antiorario. Si possono quindi presentare le singole attivazioni muscolari che sottendono alla creazione di questi movimenti di base.
Allungamento: il principio di funzionamento di tale movimento è molto semplice: la
diminuzione dell’area della sezione causa un allungamento visto che il tessuto del braccio resiste al cambiamento di volume. A causa dell’orientamento e dei punti di attacco dei muscoli trasversali si può supporre che la diminuzione dell’area della sezione possa essere creata dalla contrazione delle fibre muscolari trasversali. La diminuzione dell’area della sezione che porta all’allungamento può essere marcatamente localizzata ma può anche riguardare tutto il braccio. Ciò è molto differente dalla meccanica degli scheletri idrostatici nei quali la contrazione dei muscoli aumenta la pressione globalmente, rendendo più difficile ottenere delle deformazioni locali.
Accorciamento: Tale movimento richiede la contrazione di fibre muscolari orientate in
modo da fornire una forza di compressione longitudinale ed i fasci delle fibre muscolari longitudinali sono orientati proprio in tal modo. Quindi si assume che siano i muscoli longitudinali ad attuare l’accorciamento. Anche in questo caso, visto che il volume totale non cambia, l’accorciamento del braccio dovuto alla contrazione delle fibre muscolari longitudinali comporta una compensazione radiale ed in particolare un aumento nell’area della sezione.
La contrazione delle fibre muscolari longitudinali porta ad un allungamento delle fibre muscolari trasversali e vice versa. Si evince, quindi, che i muscoli longitudinali e trasversali funzionano come muscoli antagonisti [14].
Bending: il piegamento o bending attivo del braccio può potenzialmente coinvolgere una
sequenza di attivazione di maggior complessità rispetto a quella richiesta per l’allungamento e l’accorciamento. Tale movimento richiede la contrazione delle fibre muscolari lungo un lato del braccio che crea una forza di compressione asimmetrica che accorcia un lato del braccio e causa quindi il piegamento. Perché tale azione sia efficace il lato opposto del braccio deve resistere a tale forza di compressione, altrimenti questa contrazione unilaterale accorcerà il braccio e genererà il bending in maniera meno efficace. Poiché l’accorciamento
Fig. 2.5 (a): bending generato in maniera non efficace, (b): bending efficiente, grazie alla co-contrazione dei muscoli trasversali [14]
26
del braccio risulta in un aumento nella grandezza della sezione e le fibre dei muscoli trasversali sono orientate in modo da controllare il diametro si può creare un’azione di resistenza alla compressione longitudinale grazie alla co-contrazione delle fibre muscolari trasversali, come si può osservare in Fig. 2.5. risulta quindi che il piegamento possa coinvolgere simultaneamente sia la muscolatura trasversale sia quella longitudinale.
Come si può notare da Fig. 2.3 l’area della sezione delle fibre muscolari longitudinali nel quadrante aborale del braccio risulta maggiore di quella negli altri quadranti, indicando che i movimenti di piegamento aborale potrebbero necessitare di maggior forza che quelli in altre direzioni.
Stiffening: la variazione di rigidezza, stiffening, del braccio può essere causata dalla
co-contrazione della muscolatura trasversale e longitudinale. Tale pattern di attivazione si osserva frequentemente, ed è un componente del movimento di reaching. In alcuni studi [15] viene suggerito, infatti, che durante il movimento di reaching venga trasmessa un’onda di co-contrazione della muscolatura trasversale e longitudinale dalla base alla punta del braccio. Mentre lo stiffening si propaga dalla base alla punta si forma una piega (punto di massima curvatura del braccio) passiva durante la transizione dalla base irrigidita alla porzione terminale del braccio rilassata che viene trascinata in avanti dall’onda di co-contrazione. Un’onda di stiffening simile è osservabile in altri movimenti del polpo e potrebbe essere un’importante componente stereotipata di molti movimenti di questo animale. In alcuni movimenti una porzione significativa della parte prossimale del braccio potrebbe essere contratta ed il braccio potrebbe essere trascinato dalla rotazione della base. Tale dinamica somiglierebbe alla rotazione intorno all’articolazione della spalla di un vertebrato. Un tale comportamento probabilmente richiede la co-contrazione della muscolatura trasversale e longitudinale della massa muscolare intrinseca del braccio per poterlo irrigidire, mentre la contrazione alla base del braccio potrebbe causare o solo aumentare il movimento di trascinamento.
Torsione: poiché la torsione in un “muscular hydrostat” deriva presumibilmente dalla
contrazione di fibre muscolari disposte ad elica si pensa che tale movimento sia dato dalle fibre muscolari oblique che sono disposte proprio in tal maniera. Per ottenere il movimento di torsione sono probabilmente attivate insieme le fibre esterne di un lato del braccio e le
27
fibre mediane dell’altro lato del braccio visto che possiedono lo stesso orientamento. Il momento torsionale dei muscoli obliqui dipende dal prodotto della forza che producono (data dalla tensione delle fibre muscolari e dall’area della sezione) e la loro distanza dall’asse neutro del braccio. Sono quindi gli strati di fibre muscolari esterno e mediano ad avere un momento relativamente grande visto che sono lontani dall’asse neutro.
Il problema nel comprendere ulteriormente i pattern di movimento del polpo è dato dalla potenziale complessità del controllo motorio richiesto. Infatti, le deformazioni sopra descritte potrebbero avvenire in ogni punto o anche in più punti lungo il braccio.
2.2 Biomeccanica del movimento di reaching
Il polpo usa un numero limitato di pattern motori stereotipati per raggiungere e prendere o ispezionare un oggetto. Viene creata una piega, di solito vicino alla base del braccio (ma può essere creata ovunque lungo il braccio) ed è propagata lungo il braccio verso la punta, come illustrato in Fig. 2.6. Il segmento del braccio in posizione prossimale rispetto alla piega, cioè il punto di massima curvatura del braccio, è mantenuto relativamente dritto e la piega è sempre curvata dorsalmente. Il percorso di tale piega rimane su un piano ed essa si propaga secondo un profilo di velocità costante. Alcuni studi [16] hanno proposto l’idea che questa modalità di reaching risolva il problema dei multipli gradi di libertà riducendone il numero a tre. Un grado di libertà, infatti, servirebbe per stabilire la velocità di propagazione della piega lungo il braccio e due per l’orientamento della base del braccio nello spazio.
È stato anche suggerito che il sistema nervoso centrale mandi l’ordine di orientare la direzione del braccio ed identificare i parametri dell’estensione mentre i dettagli per l’attivazione muscolare sono deputati al sistema neuromuscolare del braccio stesso [17].
Il pattern descritto per creare il movimento di bending può essere propagato lungo il braccio creando un movimento di estensione. In alcuni studi è stata proposta l’idea che il meccanismo per la propagazione della piega fosse differente, che fosse un’onda di stiffening causata da un pattern di attivazione muscolare simmetrica che si propaga lungo il braccio. È stato creato anche un modello per testare la plausibilità dell’ipotesi e per analizzare come una tale onda possa essere controllata per produrre movimenti di estensione [18].
28
Quando gli viene presentato un target, il polpo estende uno o più braccia verso di esso usando l’onda di propagazione della piega verso la punta del braccio mentre la parte prossimale del braccio viene mantenuta quasi dritta. Tale parte non si muove quindi non risente delle forze di resistenza fluidodinamica. L’area vicino alla piega, invece, è relativamente piccola ma esposta a grandi forze di resistenza. La parte distale, cioè quella che va dalla piega alla punta del braccio ha una superficie molto maggiore ma la forza di
drag è comunque inferiore. La piega si propaga lungo una traiettoria planare che unisce il corpo dell’animale al target.
2.4 Caratteristiche cinematiche del movimento di reaching
Alcuni studi [5] sono stati compiuti sul movimento di reaching per analizzare i parametri cinematici del movimento. Visto che tali studi venivano compiuti direttamente sull’animale, il movimento di reaching doveva esser separato dal movimento dell’intero animale, in quanto che anche quest’ultimo si muoveva verso il target. Per ottenere ciò si è sottratto vettorialmente il movimento dell’occhio del polpo (indicativo del movimento dell’animale) dal movimento del punto di bending, vedi Fig.2.6.
29
Per ottenere una descrizione dell’onda di stiffening è stato identificato, nei video del movimento, il punto di massima curvatura del braccio del polpo, cioè il punto di bending, e ne è stato fatto il tracking. Attraverso un’analisi di questo tipo è stato possibile notare che il punto di bending si muove in un piano (vedi Fig. 2.7) secondo una traiettoria leggermente curva come si nota in Fig. 2.8. La planarità del movimento permette di ridurre lo spazio d’interesse da tre a due dimensioni.
Fig.2.7 A: movimento della posizione della piega nello spazio tridimensionale. La posizione dell’occhio è dato dai rettangoli, la posizione del punto della posizione della piega è segnalata con dei cerchi. La posizione del target è identificata da una croce. B: il movimento della posizione della piega relativo all’occhio. La freccia indica la direzione del movimento.
30
In Fig. 2.9 sono mostrati quattro movimenti di reaching per 3 distinti animali. I punti si riferiscono ai dati estratti, le linee al fitting con un polinomio di quinto grado. La direzione del movimento è indicata dalle frecce. La forma del grafico è leggermente curvata o diritta, indicando che il punto di bending si muove lungo una traiettoria semplice rispetto al corpo dell’animale.
Fig.2.8 il movimento della posizione della piega è planare, infatti il valore delle coordinate y durante il movimento varia molto poco. Il simbolo quadrato rappresenta la posizione della piega.
31
Fig.2.9 esempi di traiettoria del punto della piega è planare, in differenti momenti ed in differenti animali. La freccia indica la direzione del movimento.
32
Un altro parametro preso in considerazione nello studio del movimento di reaching nel polpo è la velocità del punto di bending. Il parametro preso in analisi è la velocità tangenziale, cioè:
Vtan =
Tale velocità è stata calcolata in funzione del tempo in un gruppo di prove sperimentali. Dato che la piega è una struttura dinamica che può evolvere o scomparire durante il movimento del braccio la velocità potrebbe partire da valori diversi da zero. Sono mostrati in Fig. 2.10 tre profili di velocità presi da movimenti dello stesso animale. Sebbene il range di velocità di questi movimenti vari, tutti dimostrano un profilo di velocità caratteristico. Si possono distinguere tre fasi in tale profilo. La prima fase è la parte iniziale del movimento e termina con un minimo. Essa è caratterizzata da una decrescita monotona in velocità (profili a e c) o da un aumento iniziale seguito da un picco locale (profilo b). La fase due corrisponde alla propagazione della posizione della piega lungo il braccio, questa è la porzione di movimento dove avviene la maggior parte dell’estensione del braccio. La velocità, di solito, raggiunge il massimo vicino al target ed in seguito sembra che la piega si muova verso la punta con un’onda di propagazione passiva. Tale parte corrisponde alla diminuzione di velocità nella fase tre. In molti casi, però, la piega scompare prima di aver raggiunto la punta del braccio.
33
L’invarianza del profilo di velocità tangenziale è testato normalizzando la velocità di movimento rispetto all’ampiezza ed alla durata. Usando questo metodo d’indagine sorge il problema di individuare l’inizio e la fine del movimento, visto che esso può iniziare e terminare con varie velocità. Si risolve tale problema facendo riferimento al fatto che gli andamenti della velocità tangenziale seguono un profilo con dei punti di massimo e minimo ben definiti. Si normalizzano quindi la velocità ed il tempo secondo la velocità massima e la distanza percorsa dalla posizione della piega tra questi punti di minimo e massimo:
Vnormalizzata=
Tnormalizzata= * t/D D viene calcolata come:
D =
Dove X e Y sono le coordinate dei dati dopo il fitting e l’indice t indica il numero dell’immagine (tempo).
Nella Fig. 2.11 i tre profili sono stati normalizzati e sovrapposti allineando i picchi. Queste curve normalizzate dimostrano una sovrapposizione nella fase due, suggerendo che ci sia una sorta di vincolo che causa l’aumento di velocità in questa fase. I profili di velocità,
Fig.2.10 La velocità della posizione della piega, calcolata grazie ai dati cinematici sottoposti a fitting sono graficati sul tempo. I profili hanno un pattern simile e si possono dividere in tre fasi del movimento (marcate come PhaseI, PhaseII, PhaseIII)
Fig.2.11 i profili di velocità tangenziale sono invarianti. I profili presentati sono gli stessi di Fig.2.8 ma con velocità e distanza normalizzate ed i picchi allineati. Si dimostra una sovrapposizione della fase due.
34
inoltre, risultano essere simili anche tra animali diversi.
2.5 Estrazione delle specifiche per l’implementazione robotica
Partendo dalla conoscenza dell’anatomia del braccio del polpo, dai movimenti di base e dalle analisi del movimento di reaching sopra esposti si è giunti ad una conoscenza sufficiente per poter tentare di riprodurre il movimento di reaching utilizzando una piattaforma robotica (descritta nel capitolo seguente).
Le caratteristiche salienti per la riproduzione del movimento sono:
• Viene creata una piega, di solito vicino alla base del braccio, ma in realtà quasi ovunque nel braccio, e viene propagata lungo il braccio verso la punta
• Il segmento del braccio prossimale alla piega è mantenuto relativamente dritto e la piega è ottenuta contraendo il lato aborale del braccio
• L’onda di attivazione muscolare di stiffening si muove a velocità costante
• Il tragitto della piega si muove su un piano
• Viene ipotizzata una riduzione dei parametri che determinano il movimento: un grado di libertà per il movimento della piega lungo il braccio e due gradi di libertà per l’orientamento della base del braccio
• Durante il movimento di reaching il movimento del segmento prossimale del braccio non si muove, quindi non risente delle forze di resistenza fluidodinamica
I movimenti ottenuti vengono poi filmati per poter estrarre le caratteristiche cinematiche ravvisate nei movimenti dell’animale come negli studi sopra citati. In tale modo sarà possibile ottenere un confronto diretto quantitativo oltre che qualitativo della bontà della riproduzione del movimento di reaching con i pattern di attivazione muscolare scelti ed il setup utilizzato.