Capitolo 1
Introduzione
Le piante, in quanto organismi sessili, sono frequentemente sottoposte a stress, cioè a condizioni esterne che influiscono sfavorevolmente sulla crescita, lo sviluppo o la produttività. Gli stress possono essere biotici, dovuti cioè ad altri organismi, oppure abiotici, risultanti da un eccesso o da una carenza dei fattori che determinano le caratteristiche chimiche o fisiche dell’ambiente. Nelle piante gli stress inducono un’ampia gamma di risposte, a partire da modificazioni dell’espressione genica e del metabolismo cellulare a cambiamenti delle velocità di crescita e delle rese dei raccolti. La durata, l’intensità e la velocità con cui uno stress colpisce una pianta sono tutti fattori che influenzano la risposta della pianta stessa; la combinazione di diverse condizioni sfavorevoli può provocare una risposta diversa da quella che si ha in seguito ad un singolo tipo di stress. Le caratteristiche della pianta, comprese l’identità degli organi e dei tessuti, lo stadio di sviluppo ed il genotipo, influenzano anch’essi le risposte della pianta allo stress.
I meccanismi di resistenza allo stress possono essere raggruppati in due categorie generali: meccanismi di allontanamento, che prevengono l’esposizione allo stress, e meccanismi di tolleranza, che permettono alla pianta di sopportare le condizioni sfavorevoli. Stomi infossati, spine che
riflettono la luce, radici profonde sono degli esempi di tratti costitutivi geneticamente determinati per la resistenza allo stress, tratti che sono espressi indipendentemente dal fatto che le piante siano sottoposte a stress oppure no: essi rappresentano adattamenti, perfezionamenti evolutivi che aumentano il successo adattativo di una popolazione. Altri meccanismi di resistenza sono ottenuti attraverso l’acclimatazione, cioè mediante una risposta di adattamento ai cambiamenti ambientali da parte dei singoli organismi, che modificano la propria omeostasi, cioè il proprio equilibrio fisiologico; l’esposizione a bassi livelli di stress può quindi permettere la sopravvivenza in condizioni che altrimenti sarebbero letali o il mantenimento della produttività in situazioni che possono essere dannose per i raccolti.
I cambiamenti del metabolismo e dello sviluppo indotti dallo stress possono essere attribuiti alla modificazione dei profili dell’espressione genica; una risposta ad una condizione sfavorevole inizia quando la pianta riconosce lo stress a livello cellulare: tale riconoscimento attiva le vie di trasduzione del segnale che trasmettono l’informazione all’interno delle singole cellule e in tutta la pianta. I cambiamenti dell’espressione genica sono integrati in una risposta da parte dell’intera pianta che può modificare la crescita e lo sviluppo ed influenzare anche le capacità riproduttive: la regolazione di tale risposta coinvolge ormoni (specialmente l’acido abscissico, l’acido jasmonico e l’etilene) e secondi messaggeri come il Ca2+. Alcuni geni vengono espressi maggiormente mentre altri sono repressi: i prodotti proteici dei geni indotti spesso si accumulano ma tale accumulo è influenzato anche da meccanismi di regolazione post-trascrizionale che aumentano le quantità di specifici mRNA, aumentano la traduzione, stabilizzano le proteine, modificano l’attività proteica o determinano un insieme di questi livelli di regolazione. Mediante tecniche di genetica molecolare i ricercatori hanno iniziato ad analizzare le risposte delle piante associate all’esposizione a determinati stress, uno dei quali è quello relativo alle basse temperature.
1.1 Raffreddamento e congelamento
Le lesioni da raffreddamento (chilling) avvengono in specie sensibili a temperature che sono troppo basse per una crescita normale ma non abbastanza da formare il ghiaccio; tipicamente tali lesioni si hanno in specie di origine tropicale o sub-tropicale, quali il mais, il pomodoro, il riso, il cotone. Quando piante che crescono a temperature relativamente calde (25-35°C) sono raffreddate a 10-15°C l’accrescimento rallenta, appaiono sulle foglie zone clorotiche o lese e il fogliame risulta come bagnato; la sensibilità al freddo nelle varie specie vegetali è funzione anche dello stadio biologico della pianta e degli organi vegetali interessati: nelle piante superiori gli stadi giovanili sono di solito più sensibili alle basse temperature di quanto lo siano le piante adulte, come dimostrato dalla forte termodipendenza del processo di crescita dei semenzali in genere. Organi particolarmente suscettibili al danno da freddo sono radici, rizomi e bulbi che, se raffreddati, possono portare la pianta all’appassimento. Specie che sono considerate generalmente sensibili al raffreddamento mostrano variazioni apprezzabili nella risposta alle temperature fredde; tuttavia la resistenza spesso aumenta se le piante vengono temprate da esposizione lenta e graduale a temperature fredde ma non lesive; esposizioni repentine a temperature inferiori a 0°C (freezing) producono certamente delle lesioni nelle foglie, che mostrano inibizione della fotosintesi e della traslocazione dei carboidrati, minor respirazione, inibizione della sintesi proteica e aumento della degradazione delle proteine presenti. Tutte queste risposte dipendono probabilmente da un meccanismo primario comune che implica la perdita delle proprietà delle membrane durante il raffreddamento: la perdita di soluti è sintomo di danno alla membrana plasmatica e probabilmente anche al tonoplasto e l’inibizione della fotosintesi e della respirazione riflettono lesioni alle membrane dei cloroplasti e dei mitocondri.
La capacità di tollerare temperature di congelamento in condizioni naturali varia enormemente tra le piante resistenti al raffreddamento. I semi ed altri
tessuti parzialmente disidratati possono essere mantenuti indefinitamente a temperature vicine allo zero assoluto (0 K), quindi le temperature basse non sono intrinsecamente dannose; cellule vegetative idratate possono anche conservare la loro vitalità se vengono raffreddate molto velocemente, in modo da evitare la formazione di grandi cristalli di ghiaccio che, formandosi lentamente, perforerebbero e distruggerebbero la fine struttura cellulare che vi è all’interno del protoplasto vivente; i cristalli di ghiaccio che si formano durante i congelamenti rapidi sono troppo piccoli per causare danni meccanici alle strutture sub-cellulari. Anche il ritorno a temperature normali deve essere estremamente rapido in modo da prevenire la conversione dei cristalli di ghiaccio in vapore acqueo attraverso la sublimazione, o la crescita dei cristalli ad una dimensione dannosa che avviene a temperature intermedie. In condizioni naturali comunque il raffreddamento di organi vegetali multicellulari intatti non è mai abbastanza rapido da prevenire in cellule completamente idratate la formazione di grandi e dannosi cristalli di ghiaccio. Quando un tessuto è raffreddato in condizioni naturali il ghiaccio si forma di solito prima fra gli spazi intercellulari e nei vasi xilematici delle foglie e degli steli, dentro i quali si propaga velocemente a causa delle grandi dimensioni dei vasi e del fatto che la linfa grezza ha un alto punto di congelamento: questa formazione di ghiaccio non è letale e nelle piante resistenti il tessuto si riprende completamente se riscaldato. Se le piante sono esposte al gelo per un periodo esteso la crescita dei cristalli extracellulari è il risultato dello spostamento di acqua allo stato liquido dal protoplasto verso il ghiaccio apoplastico, causando un eccesso di disidratazione, conseguenza della formazione di un gradiente del potenziale idrico: a una certa temperatura, infatti, il potenziale chimico del ghiaccio è minore di quello dell’acqua allo stato liquido, inoltre la pressione di vapore del ghiaccio extracellulare è minore di quella dell’acqua presente nel citoplasma o nel vacuolo e questo comporta la fuoriuscita dell’acqua dalla cellula. Per comprendere meglio questo fenomeno bisogna introdurre il concetto di osmolalità che dipende dalla somma di tutti gli elettroliti ed i soluti osmotici disciolti in una soluzione e si misura come
moli di soluto presenti in 1 kg di acqua. L’osmolalità, per l’azione della legge di massa, deve essere mantenuta uguale e costante all’interno dei diversi compartimenti liquidi separati da una membrana semipermeabile: un suo innalzamento è responsabile di un passaggio di acqua dal compartimento meno concentrato verso quello più concentrato, dove provoca un aumento della pressione osmotica. Nel caso di un’ideale soluzione parzialmente congelata l’osmolalità è in funzione della temperatura (espressa in Kelvin) secondo la relazione:
La quantità della soluzione iniziale che non congela a nessuna temperatura è data invece dalla relazione:
dove MO è l’osmolalità iniziale.
Attraverso queste due equazioni è possibile calcolare il punto di congelamento e la quantità di acqua congelata ad una data temperatura. Poiché la formazione del ghiaccio aumenta la concentrazione della soluzione extracellulare, si viene a creare un gradiente nel potenziale chimico dell’acqua (potenziale idrico) tra citoplasma e apoplasto; l’acqua tenderà a muoversi al di fuori della cellula per portare all’equilibrio il potenziale e ne causerà la disidratazione.
Figura 1.1: l’acqua si
sposta dal citoplasma verso l’apoplasto creando nella cellula una situazione di stress idrico.
Tale disidratazione del citoplasma nei tessuti non acclimatati porta al collasso della cellula, in quelli acclimatati è reversibile fino ad una data soglia.
Figura 1.2: congelamento e disidratazione in cellule non-acclimatate e in cellule
acclimatate; in queste il processo è reversibile fino ad un certo punto, superato il quale, tuttavia, si va incontro ad un’eccessiva perdita di turgore ed alla morte cellulare.
Il fenomeno del congelamento nelle cellule vegetali può quindi essere riassunto in tre punti fondamentali:
1. grandi flussi di acqua attraversano il plasmalemma durante il congelamento e lo scongelamento;
2. i cristalli di ghiaccio si espandono negli spazi extracellulari andando a gonfiare i tessuti e comportando la rottura delle interconnessioni cellulari;
3. il citoplasma va incontro a essiccazione e le cellule collassano.
La membrana plasmatica assume quindi una funzione critica poiché deve restare fisicamente intatta per preservare l’integrità della cellula durante le contrazioni e i rigonfiamenti tipici del congelamento e dello scongelamento; le sue proprietà inoltre influenzano il flusso di acqua, di ioni e di soluti tra citoplasma e apoplasto. Per tali ragioni non deve sorprendere se lo sviluppo della tolleranza al gelo è associato con i cambiamenti che avvengono nelle proprietà fisiche e nella composizione della membrana.
1.2 Danni da congelamento
Esistono diverse teorie che cercano di definire la natura delle lesioni alla membrana plasmatica:
• ipotesi del solfidrile-disolfuro: nel 1980 Levitt propose che i danni da congelamento nelle cellule fossero dovuti alla denaturazione proteica,
responsabile di modifiche nella struttura terziaria delle proteine: in seguito alla disidratazione cellulare si ha la formazione irreversibile di nuovi ponti disolfuro intramolecolari che legano trasversalmente le proteine; quando inizia lo scongelamento la cellula tende ad espandersi grazie all’ingresso di acqua ma i nuovi legami formatisi tra le proteine non si rompono ed impediscono il ripristino del volume originale della cellula (Levitt, 1980); inoltre i ponti disolfuro sono responsabili dell’inattivazione enzimatica. In questo modello la tolleranza al congelamento comporta la diminuzione della denaturazione proteica prevenendo la formazione dei ponti disolfuro. Sembra quindi che questi legami siano il risultato e non la causa dei danni dovuti al congelamento o, in altre parole, siano un effetto secondario e non primario: quando Levitt propose questa teoria non poteva conoscere il significato dello stress ossidativo e la possibilità che forme attivate dell’ossigeno siano in grado di sottrarre ioni H+ dai gruppi tiolici,
permettendo la formazione di legami tra due atomi di zolfo.
Figura 1.3: modello di Levitt relativo alla formazione dei ponti disolfuro nelle cellule
• lisi indotta da espansione: Steponkus (1984) ha basato molte delle sue ricerche sull’osservazione di protoplasti (cioè cellule senza parete, digerita enzimaticamente), durante un ciclo di congelamento-scongelamento; questo sistema fornisce un ottimo metodo per osservare il comportamento della membrana ed il flusso dell’acqua che la attraversa. I protoplasti si contraggono e si espandono durante un ciclo di congelamento-scongelamento. In un mezzo isotonico sul plasmalemma viene esercitata una pressione minima, in quanto si è in una condizione di equilibrio osmotico; a causa dell’abbassamento termico il mezzo si congela e diventa ipertonico: l’acqua tende quindi a diffondere fuori dal cloroplasto per ristabilire l’equilibrio osmotico. Nei protoplasti delle foglie non acclimatate questo fenomeno è accompagnato da una vescicolazione endocitotica: piccole vescicole di plasmalemma si separano dalla membrana per finire nel citoplasma. Questo non è dannoso di per sé ma causa la riduzione della superficie della membrana che comporta la lisi cellulare al momento dello scongelamento: infatti il mezzo diventa ipotonico e l’acqua comincia a diffondere nuovamente nella cellula rimpicciolita che non è più in grado di espandersi completamente e di conseguenza avviene la lisi. Nei protoplasti delle foglie acclimatate la membrana si comporta in una maniera molto differente durante la contrazione osmotica: invece della vescicolazione endocitotica, i protoplasti acclimatati formano estrusione esocitotiche quando vengono congelati o posti in una soluzione ipertonica (Dowgert and Steponkus, 1984) quindi, al momento dello scongelamento, il processo di espansione dovuto alla reidratazione è completamente reversibile e i protoplasti recuperano il loro volume originario. Il diverso comportamento dei protoplasti acclimatati e quelli che non lo sono si ipotizza sia dovuto a differenze nelle interazioni dei lipidi formanti le membrane.
Figura 1.4: piante non-acclimatate e lisi. Figura 1.5: piante acclimatate.
• fase esagonale II: l’essiccazione citoplasmatica durante il congelamento causa dei cambiamenti non reversibili a livello della composizione lipidica del plasmalemma. Analisi effettuate con il microscopio elettronico su tessuti non acclimatati hanno permesso l’individuazione di regioni di membrana che mancano di particelle intramembranose (proteine): la loro assenza comporta una modifica del plasmalemma che assume una conformazione definita “fase esagonale II” e perde la sua tipica struttura caratterizzata da un doppio strato fosfolipidico (Gordon-Kamm e Steponkus, 1984). La perdita di fosfolipidi e l’accumulo di acidi grassi come conseguenza di reazioni di de-esterificazione sono il risultato del congelamento dei tessuti; nelle piante acclimatate non si formano fasi esagonali: probabilmente questo è dovuto a cambiamenti nella composizione lipidica della membrana che avvengono in seguito all’esposizione a basse temperature e che riducono la tendenza dei lipidi di legarsi in maniere non idonee.
Figura 1.6: struttura tipica della
membrana plasmatica: doppio strato fosfolipidico.
Figure 1.7 e 1.8 : formazione della fase esagonale II.
• permeabilità della membrana: gli effetti del congelamento non necessariamente sono legati alla drastica lisi cellulare, come risulta dalle ricerche di Steponkus e colleghi: spesso infatti i danni dovuti alle basse temperature riguardano la semplice perdita della semi-permeabilità della membrana. Studi effettuati sulla cipolla dimostrano che il congelamento implica una modifica a livello delle proteine integrali di membrana responsabili del trasporto di ioni K+ (Palta e Li, 1980). Ulteriori studi confermano questi risultati; Palta ha proposto che stress termici sub-letali causano effettivamente delle alterazioni alla membrana associate con i trasportatori H+-ATPasi: l’attività delle ATPasi, infatti, decresce dopo un ciclo di congelamento-scongelamento ed il loro recupero è impedito dalla presenza di vanadato, un inibitore di queste pompe protoniche.
• campi elettrici transienti: quando una soluzione acquosa congela si viene a formare una differenza di potenziale tra ghiaccio e soluzione non ancora ghiacciata, che dipende dal tipo di ioni presenti, dalla loro concentrazione e dalla velocità di formazione del ghiaccio. A volte questa differenza di potenziale è talmente alta da comportare una destabilizzazione della membrana plasmatica con conseguente lisi cellulare (Steponkus et al., 1985). L’acclimatazione consente, attraverso modifiche a livello della membrana, un aumento della soglia critica di potenziale (sopra la quale viene indotta la lisi cellulare).
• stress ossidativo: come analizzato fino ad ora, il congelamento causa numerosi danni alla membrana, tra i quali modifiche strutturali, incremento della microviscosità, degradazione dei fosfolipidi e accumulo di acidi grassi; tutti questi sintomi possono essere simulati in
vitro se le cellule vengono trattate con il superossido: ciò fa supporre
che questa molecola partecipi nel danneggiamento cellulare indotto dagli stress termici e che la tolleranza allo stress ossidativo sia un’importante componente della tolleranza al congelamento. Esistono diverse ipotesi sul legame tra congelamento e stress ossidativo: questo può essere considerato come un effetto secondario dell’esposizione alle basse temperature, le quali causano disfunzioni nel trasporto elettronico di membrana con conseguente sintesi del superossido. Tuttavia il congelamento implica anche la produzione di un potenziale tra le particelle di ghiaccio e la soluzione non ghiacciata dovuto all’esclusione dei gas dal ghiaccio in formazione, come discusso precedentemente: si creano, così, alte pressioni di ossigeno che, in presenza di ioni metallici ridotti, favoriscono la sintesi del superossido. Kendall e McKersie (1989) hanno suggerito che le membrane delle cellule acclimatate accumulino antiossidanti solubili in grado di disattivare le specie reattive dell’ossigeno e minimizzare così le pericolose reazioni di perossidazione.
1.3 Formazione del ghiaccio nelle cellule
Durante il congelamento rapido il protoplasto, compreso il vacuolo, si sovraraffredda, cioè l’acqua della cellula rimane liquida anche a temperature di svariati gradi inferiori al suo punto di congelamento teorico. Il processo che porta alla formazione e alla propagazione di un cristallo di ghiaccio a partire da punti di potenziale osmotico meno negativo è definito nucleazione del
ghiaccio (Burke e Lindow, 1990) e vi partecipano svariate centinaia di
molecole, cioè grandi polisaccaridi e proteine (nucleatori del ghiaccio): alcune di queste proteine facilitano il processo allineando le molecole d’acqua lungo domini di amminoacidi ripetuti all’interno della proteina stessa.
Il sovraraffreddamento (supercooling) è quel fenomeno sfruttato principalmente dalle piante arboree per evitare l’essiccazione del citoplasma durante il congelamento, per il quale durante la fase iniziale della perdita del calore dal tessuto la temperatura scende sotto il reale punto di congelamento del citosol e del vacuolo senza un cambiamento di fase da quella liquida a quella solida. A mano a mano che le temperature diminuiscono ulteriormente si forma ghiaccio negli spazi intercellulari e vi è una perdita di energia di calore dovuta al calore latente di fusione dell’acqua; a questo stadio la temperatura del tessuto rispecchia il bilancio tra il guadagno termico dalla formazione del ghiaccio e la perdita di calore verso l’ambiente. Come risultato, quando il ghiaccio comincia a formarsi la temperatura aumenta rapidamente e rimane a questi valori fino a quando tutta l’acqua contenuta nella parete cellulare è congelata. Raggiunto tale punto la perdita di calore si arresta e la temperatura comincia di nuovo a scendere; la liberazione di calore durante la formazione del ghiaccio è la tecnica utilizzata quando le colture vengono spruzzate con acqua durante le brinate: fino a quando l’acqua continuerà a congelare fuori dalle cellule rilascerà calore che impedirà il congelamento intracellulare. La formazione del ghiaccio all’interno degli spazi intercellulari di cellule sensibili al gelo non è letale, ma quando vengono sottoposte a
periodi estesi di temperature gelide il vapore acqueo si sposta dal protoplasto non congelato verso la parete cellulare causando la crescita dei cristalli all’interno della parete; questa lenta disidratazione concentra i soluti nel protoplasto, abbassando il punto di congelamento di 2-3°C. Se la temperatura continua a scendere si può notare una seconda fase di liberazione di calore di fusione dell’acqua: questa fase implica una serie di piccoli eventi di congelamento, dove ogni picco di liberazione di calore rappresenta il congelamento dei protoplasti e coincide con la perdita in vitalità. La formazione dei cristalli di ghiaccio nelle pareti cellulari o nel protoplasto richiede la presenza di strutture definite punti di nucleazione del ghiaccio, nei quali i cristalli iniziano a crescere. In alcune specie l’acclimatazione conferisce l’abilità di sopprimere la nucleazione del ghiaccio nel protoplasto, permettendo un sovraraffreddamento spinto di molti gradi sotto il punto di congelamento, senza la formazione di ghiaccio: tale fenomeno comunque ha un limite inferiore di circa –40°C (temperatura della nucleazione spontanea dell’acqua: in realtà la temperatura omogenea di nucleazione delle gocce d’acqua pura sarebbe di –38,1°C ma la presenza di soluti la abbassa). A tale temperatura o a temperature più basse i cristalli di ghiaccio si formano anche senza i punti di nucleazione e diventano inevitabili il congelamento intracellulare e la morte della cellula.
L’abilità dei tessuti delle piante arboree di sovraraffreddarsi varia tra le specie: la “temperatura soglia” specie-specifica limita la loro distribuzione geografica a regioni dove tale temperatura minima non viene superata (Burke
et al., 1976). Tra le piante coltivate molte colture da frutto, tra le quali il pesco
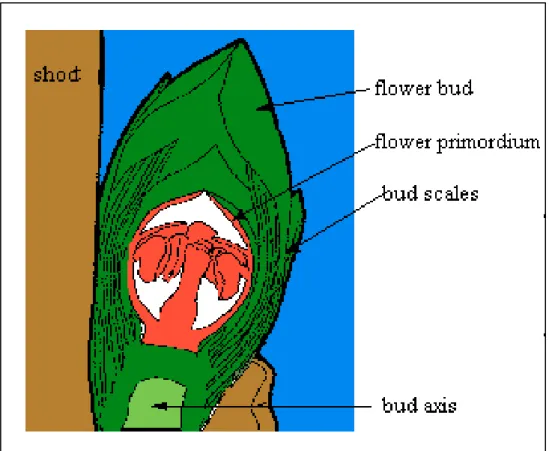
e la vite, tollerano le temperature invernali evitando il congelamento delle loro gemme fiorali, delle gemme vegetative e delle porzioni della corteccia o dei tessuti legnosi. In queste specie il ghiaccio si forma inizialmente nei tessuti che circondano la gemma e successivamente nell’asse gemmale alla temperatura di circa –5°C (Ashworth et al., 1989). Il primordio fiorale è circondato da un impenetrabile strato di cellule contenenti nella parete depositi di cere: questo
strato isola le cellule meristematiche del primordio da quelle dei tessuti esterni e dell’asse, evitando la penetrazione dei cristalli di ghiaccio.
Figura 1.9: rappresentazione schematica di una gemma fiorale di una pianta arborea.
All’interno del primordio sono assenti i siti di nucleazione del ghiaccio per cui, se la temperatura si abbassa, l’acqua del primordio tende a sovraraffreddarsi fino al raggiungimento della temperatura critica per la nucleazione del ghiaccio, che per il pesco è –16°C. Una volta che il ghiaccio si forma all’interno del primordio, lo squilibrio nei potenziali chimici è talmente accentuato che la gemma spontaneamente congela sia extra che intracellularmente e muore: questo significa che un albero di pesco con una temperatura critica di sovraraffreddamento di –16°C può sopravvivere ad una prolungata esposizione a –15,5°C senza subire danni cellulari ma viene devastata da una singola esposizione a –16,5°C.
1.4 Batteri e brina
Quando le foglie vengono raffreddate a temperature nella gamma dai −3 ai −5°C la formazione dei cristalli di ghiaccio sulla superficie (brina) viene accelerata dalla presenza di certi batteri che vivono naturalmente sulla superficie fogliare, come Pseudomonas syringae ed Erwinia herbicola, chiamati INA (ice nucleation active): questi batteri infatti fungono da punti di formazione del nucleo di ghiaccio. Quando vengono artificialmente inoculate con tali colture batteriche le foglie di specie sensibili alla brina congelano a temperature più alte di foglie non trattate con i batteri (Lindow et al., 1982): il ghiaccio di superficie si propaga velocemente verso gli spazi intercellulari all’interno della foglia, portando alla disidratazione cellulare e rendendo quindi le piante più suscettibili ai danni da congelamento. Ceppi batterici sono stati modificati geneticamente in modo tale da renderli inattivi nella formazione dei nuclei di ghiaccio, quindi sono stati utilizzati in spray fogliari su coltivazioni di valore per minimizzare il numero di punti potenziali di nucleazione attraverso la loro competizione con i ceppi locali.
Figura 1.10: eventi di nucleazione del ghiaccio in dischi fogliari di mais
1.5 Cambiamenti metabolici
Durante l’acclimatazione al freddo il metabolismo delle piante cambia drasticamente e cambiano di conseguenza i livelli dei metaboliti e le attività enzimatiche: è ipotesi comune che la tolleranza alle basse temperature sia associata con l’accumulo di specifici metaboliti crioprotettivi, che proteggono cioè la cellula durante il ciclo congelamento-scongelamento; risulta tuttavia molto difficoltoso fare una distinzione tra questi cambiamenti metabolici per determinare quali siano critici nell’acquisizione della tolleranza al freddo e quali siano semplici adattamenti per favorire la crescita alle basse temperature, quali siano attivati come reazione alle lesioni e quali siano gli effetti primari e quelli secondari. Sono stati effettuati molti studi con la finalità di comprendere a pieno tale fenomeno e di trovare le giuste correlazioni tra acclimatazione e cambiamenti metabolici; dai risultati ottenuti si evince che (Levitt, 1980):
• la concentrazione osmotica aumenta con l’acclimatazione; i più importanti cambiamenti nel potenziale osmotico riguardano l’accumulo di zuccheri, i quali possono abbassare il punto di congelamento di un tessuto, funzionare da riserve energetiche, alterare le proprietà fisiche delle membrane e infine agire come molecole crioprotettive nei confronti della struttura e della funzione delle proteine;
• il contenuto d’acqua di un tessuto è inversamente proporzionale alla tolleranza al congelamento: l’acclimatazione promuove contemporaneamente la perdita di acqua e l’accumulo di amido e proteine, che non sono osmoticamente attivi. La perdita d’acqua ha risvolti positivi poiché comporterà una minore formazione di ghiaccio e, di conseguenza, una minor espansione degli spazi intercellulari;
• i lipidi si accumulano durante l’acclimatazione e gli acidi grassi tendono ad un maggior grado di insaturazione;
• c’è una stretta correlazione tra il contenuto di proteine solubili e la tolleranza al congelamento: questa infatti è associata ad un incremento della quantità di mRNA, polisomi e tRNA richiesti per la sintesi proteica;
• i livelli dei regolatori di crescita cambia in quanto aumenta la concentrazione dell’acido abscissico mentre diminuisce quella delle gibberelline: in effetti la tolleranza al freddo è strettamente associata alla quiescenza o alla dormienza (indotte dall’ABA) e viene ostacolata dalla promozione della crescita dovuta alle gibberelline;
• si è osservata una relazione inversa tra tasso respiratorio e tolleranza al congelamento, anche se questo può dipendere dalla durata del periodo di acclimatazione;
• sono necessari luce e CO2 in modo da garantire un processo
fotosintetico ottimale per sostenere l’accumulo di zuccheri, proteine ed altri soluti; comunque anche in questo caso la relazione acclimatazione-fotosintesi è complessa e può dipendere dalla specie e dallo stadio di sviluppo della pianta.
1.6 Limitazione della formazione di ghiaccio
extracellulare
1.6.1 Lipidi
Le membrane vegetali sono costituite da un doppio strato lipidico contenente proteine e steroli: le proprietà fisiche dei lipidi hanno una grande influenza sull’attività delle proteine integrali di membrana, comprese le proteine formanti canali che regolano il trasporto degli ioni e di altri soluti. In piante sensibili al raffreddamento i lipidi del doppio strato posseggono un’alta percentuale di catene di acidi grassi saturi e questo tipo di membrana tende a solidificare in uno stato semi-cristallino a temperature ben sopra 0°C; a mano a mano che le membrane diventano meno fluide le loro componenti proteiche smettono di funzionare normalmente, così l’attività della H+-ATPasi è inibita,
il trasporto di soluti fuori e dentro le cellule è indebolito, la trasduzione dell’energia diminuisce e vi è una forte influenza sul metabolismo dipendente dagli enzimi. Inoltre le foglie sensibili al freddo esposte ad alti flussi fotonici e a temperature di congelamento subiscono la fotoinibizione, con conseguente danno grave all’apparato fotosintetico.
I lipidi di membrana di piante resistenti al freddo spesso posseggono una proporzione maggiore di acidi grassi insaturi (in particolare di-insaturi come la fosfatidilcolina e la fosfatidiletanolammina) rispetto a quelli di piante sensibili. Durante il processo di acclimatazione alle basse temperature aumenta la percentuale di lipidi insaturi (Williams et al., 1988; Palta et al., 1993; Uemura e Steponkus, 1997): tale processo abbassa le temperature alle quali i lipidi di membrana cominciano un cambiamento graduale di fase da quella liquida a quella semi-cristallina e quindi impartisce una determinata protezione contro i danni da freddo. In queste piante temprate le membrane modificate sono più in grado di mantenere la fluidità e le loro funzioni alle basse temperature.
L’importanza dei lipidi di membrana per la tolleranza alla basse temperature è stata dimostrata da studi con mutanti e caratteri transgenici in cui l’attività di particolari enzimi portava a cambiamenti specifici nella composizione dei lipidi di membrana indipendentemente dall’acclimatazione. Il grado di saturazione degli acidi grassi del fosfatidilglicerolo (PG) è particolarmente importante per la resistenza al freddo dei cloroplasti: in
Arabidopsis la trasformazione con un gene di Escherichia coli che aumenta la
proporzione di specie molecolari con alto punto di fusione causa la morte delle piante quando esposte alla luce e alla temperatura di 2°C (fotoinibizione letale) (Nishida e Murata, 1996). Allo stesso modo i mutanti fab1 di Arabidopsis posseggono quantità superiori di acidi grassi saturi, particolarmente 16:0 (acido palmitico): durante un periodo di 3−4 settimane a temperature fredde la fotosintesi e l’accrescimento erano gradatamente inibiti, culminando con la distruzione dei cloroplasti e la cessazione del processo fotosintetico; a temperature non fredde i mutanti crescevano come le piante wild-type (Wu et
al., 1997).
1.6.2 Proteine antigelo
Come trattato precedentemente la tolleranza al congelamento può essere ottenuta attraverso una maggiore stabilità delle membrane cellulari grazie all’aumento del grado di insaturazione dei componenti lipidici; sono state tuttavia identificate numerose proteine specializzate che possono essere d’aiuto per la limitazione della formazione dei cristalli di ghiaccio tramite un meccanismo non colligativo, il cui effetto cioè non dipende dalla presenza di soluti (che causerebbe l’abbassamento del punto di congelamento dell’acqua): tali proteine antigelo (anti-freeze proteins, AFPs) sono indotte dalle basse temperature e si legano alla superficie dei cristalli di ghiaccio, prevenendone o rallentandone l’ulteriore crescita all’interno della cellula (Wisniewski et al., 1999). Le AFPs hanno una composizione amminoacidica simile fra loro e con i
membri di tre classi di proteine apoplastiche associate alla patogenesi
(PR-proteins): endochitinasi, endo-beta-1,3-glucanasi e proteine tipo-taumatina
(Griffith e Yaish, 2004; Hon et al., 1995). La quantità di tali proteine nelle piante non acclimatate è minima e sono totalmente assenti nelle piante sensibili, mentre si ha un notevole accumulo nelle piante acclimatate (Thomashow, 1998), dove ne sono state individuate diverse isoforme. Le distribuzioni delle proteine AFP e delle proteine PR sono molto simili e possono riflettere la comune via biosintetica: studi di immunolocalizzazione (mediati cioè dall’uso di anticorpi) hanno permesso infatti di individuare nell’epidermide delle foglie e nelle pareti delle cellule del mesofillo i siti caratterizzati da una consistente presenza di tali proteine.
Le AFPs, oltre ad essere responsabili dell’inibizione della nucleazione, sono coinvolte in due fenomeni particolari:
• isteresi termica: ad alte concentrazioni le AFPs abbassano il punto di congelamento di una soluzione senza influenzare il punto di fusione; • inibizione della ricristallizzazione: a basse concentrazioni riescono ad
impedire durante la ricristallizzazione delle soluzioni parzialmente ghiacciate la formazione di grossi cristalli, responsabili di pericolosi danni fisici alla cellula (Griffith et al., 2005; Smallwood et al., 1999); oltre alla loro importanza relativa alla tolleranza verso il congelamento nelle piante, queste proteine sono oggetto di studio anche per il loro possibile utilizzo nell’industria alimentare come additivo per prevenire la ricristallizzazione del ghiaccio nei cibi congelati e nei gelati.
1.6.3 Zuccheri
Oltre ad alcune proteine indotte dal freddo si ritiene che anche gli zuccheri e gli amminoacidi (prolina, arginina, alanina) abbiano effetti crioprotettivi
andando a stabilizzare le membrane durante la disidratazione causata dall’abbassamento termico. Il glucosio predomina tra gli zuccheri solubili associati alla tolleranza al gelo ma in alcune specie assolvono la stessa funzione il saccarosio e il fruttosio (Sasaki et al., 1996), oppure il raffinosio e lo stachiosio (Castonguay et al., 1995), il sorbitolo o il mannitolo: questi si accumulano nelle pareti cellulari e limitano la crescita dei cristalli. Studi di ingegneria genetica effettuati su tabacco (pianta che non si acclimata) hanno permesso la sovraespressione di un gene batterico codificante la pirofosfatasi, che ha portato all’accumulo di carboidrati solubili: le foglie dei cloni mostravano un aumento di 1,2°C nella resistenza al gelo se comparati con il
wild-type (Hincha et al., 1996).
Altro zucchero particolarmente importante è il maltosio, la cui concentrazione tende ad aumentare durante gli shock termici e contribuisce alla protezione del trasporto elettronico fotosintetico e a quella delle proteine dello stroma (Kaplan e Guy, 2004): studi con RNA interferenziale hanno dimostrato che le linee con livelli ridotti di beta-amilasi (enzima responsabile della degradazione dell’amido con conseguente formazione del maltosio) riducono rispetto al wild-type l’accumulo di maltosio in risposta alle basse temperature e mostrano un danno maggiore alle componenti del trasporto elettronico, come evidenziato dalla riduzione della fluorescenza della clorofilla.
1.6.4 Acido abscissico
L’acclimatazione in natura è indotta dall’abbassamento di temperatura e dalla riduzione del fotoperiodo nel tardo autunno e la resistenza che la pianta acquisisce viene rapidamente persa nel periodo primaverile; di solito occorrono diversi giorni di esposizione alle temperature fredde per indurre completamente la resistenza al gelo ma, se portate a temperature superiori, le
piante perdono la loro acclimatazione in meno di 24 ore e diventano di nuovo suscettibili al gelo.
Fin dalla metà degli anni ’80, quando i cambiamenti dell’espressione genica in risposta alle temperature di congelamento furono osservate per la prima volta in spinacio, sono stati individuati numerosi geni che rispondono ai trattamenti al freddo. Sfortunatamente per molti di questi geni non è stato ancora determinato un chiaro ruolo funzionale. Molti dei geni indotti dalle basse temperature sono indotti anche dalla sola carenza idrica o da trattamenti con ABA esogeno senza esposizione al freddo (Guy, 1990): questi comportano cambiamenti nella modalità di sintesi di nuove proteine, che possono essere separate su gel bidimensionali. Alcuni cambiamenti sono unici per un particolare trattamento (freddo o ABA) ma una determinata quantità delle nuove proteine sintetizzate indotte dal freddo appare la stessa di quelle indotte dall’ABA o da deboli deficit idrici. L’isolamento dei geni di queste proteine indotte dalle basse temperature indica che esse mostrano omologia con la famiglia proteica RAB/LEA/DHN: i membri di questa famiglia sono sintetizzati durante la maturazione dei semi sotto controllo dello sviluppo ma sono anche indotti in cellule vegetative da deficit idrico e dall’esposizione all’ABA. Queste proteine ricche in glicina, le deidrine, sono molto idrofile e posseggono la proprietà inconsueta di essere stabili a temperature di ebollizione; si ritiene che proteggano le cellule disidratate dalla mancanza d’acqua o dal gelo, stabilizzando le membrane o altre proteine (Allagulova et
al., 2003).
L’ABA sembra avere un ruolo nell’indurre la tolleranza al congelamento: in Arabidopsis, specie resistente al freddo, deboli deficit idrici ma anche l’esposizione a basse temperature sono fattori che aumentano le concentrazioni di ABA endogeno nelle foglie. Prove del ruolo dell’ABA come segnale nell’acclimatazione sono evidenti nei mutanti aba1 di Arabidopsis, incapaci di sintetizzare l’ormone: sebbene questi vengano esposti a basse temperature, non riescono ad acclimatarsi e non sviluppano la tolleranza al congelamento; solo l’esposizione all’ABA esogeno ridona a questi mutanti ABA-deficienti la
capacità di sviluppare tolleranza al gelo (Heino et al., 1990; Mantyla et al., 1995). D’altra parte non tutti i geni indotti dalle basse temperature sono ABA-dipendenti e non è chiaro se la loro espressione sia critica per il competo sviluppo della tolleranza al gelo.
1.7 Cascata del segnale
La trasduzione del segnale è il mezzo attraverso il quale le cellule elaborano una risposta ad un stimolo; avviene grazie alla presenza di una rete di interazioni tra le componenti delle cellule e tra le cellule stesse. Come descritto in precedenza le piante sono costantemente sottoposte a innumerevoli stimoli esterni e interni, responsabili di variazioni nel metabolismo, nella crescita e nello sviluppo delle cellule.
Ormoni Ferite Zuccheri Peptidi Tensioni meccaniche della parete Trasmissione Cambiamenti nel flusso ionico Regolazione delle vie metaboliche Regolazione dell’espressione genica Cambiamenti del citoscheletro
Cellula
Luce Amminoacidi Stato idrico Temperatura Patogeni Minerali Amplificazione Divergenza verso molteplici bersagliFigura 1.11: rappresentazione schematica della percezione e della trasduzione dei segnali nella cellula,
con conseguenti alterazioni della crescita o del metabolismo (rivisitazione da Buchanan et al., 2000).
Questi stimoli vengono percepiti dalla cellula per mezzo di recettori proteici o attraverso cambiamenti del potenziale di membrana; sebbene alcuni siano localizzati nel citosol o in altri compartimenti cellulari, la maggior parte dei recettori noti si trova sul plasmalemma, come i recettori associati alle proteine G, quelli associati agli enzimi o quelli associati a canali ionici, i quali possono anche essere localizzati nelle membrane interne della cellula, cioè nel tonoplasto e nella membrana del reticolo endoplasmatico. Tra i più importanti
bisogna citare il recettore per l’inositolo 1,4,5-trifosfato (IP3; è un secondo
messaggero in quanto viene sintetizzato dalla fosfolipasi C una volta attivata da una proteina G): questo recettore è composto da quattro subunità, ognuna delle quali contiene quattro domini che attraversano la membrana; quando l’IP3 si lega al recettore, i cambiamenti conformazionali che ne derivano
determinano il movimento di due delle sue subunità. La distribuzione di cariche positive e negative stabilizza la conformazione aperta del canale e permette l’influsso di Ca2+ nel citoplasma dal vacuolo e dal reticolo
endoplasmatico, ognuno dei quali contiene concentrazioni di Ca2+ di molti
ordini di grandezza maggiori rispetto a quelle del citosol. Insieme all’IP3 anche
il Ca2+ si è dimostrato un importante secondo messaggero, che interviene nella
trasduzione del segnale originatasi in seguito all’esposizione alle basse temperature (White e Broadley, 2003).
1.8 Ruolo del calcio
Per acclimatarsi una pianta deve percepire un segnale e tradurlo in risposte biochimiche; il segnale deve stimolare cambiamenti osservabili del [Ca2+] o
del flusso di Ca2+ attraverso le membrane che a loro volta devono precedere le
risposte fisiologiche: la trasmissione del segnale del Ca2+ dipende dai gradienti
chimici transmembrana dell’elemento attraverso la membrana plasmatica e le membrane intracellulari. Per facilitare i processi di trasmissione dei segnali le cellule mantengono concentrazioni basali di Ca2+ molto basse (da 100 a 200
nM); il vacuolo ed il reticolo endoplasmatico rugoso costituiscono grandi riserve di Ca2+ intracellulare (tipicamente circa 1 mM) che possono essere
immobilizzate dall’IP3 e da altri segnali prodotti dalla membrana plasmatica,
cellulare, dove il Ca2+ è utilizzato come una molecola strutturale, si stima che
la sua concentrazione sia da 0,5 a 1 mM; anche i mitocondri, i cloroplasti e persino il nucleo possono funzionare da riserve di Ca2+. Quando le cellule
ricevono i segnali, i canali del Ca2+ si aprono temporaneamente e la sua
concentrazione aumenta rapidamente: questa non può essere tollerata a lungo dalla cellula, poiché il Ca2+ può interferire con il metabolismo competendo con
il Mg2+ per l’ATP, quindi vengono attivate alcune proteine carrier (Ca2+
-ATPasi dei vari organelli, l’antiporto di Ca2+/H+ del tonoplasto o l’uniporto di
Ca2+ della membrana mitocondriale interna), fornendo ai meccanismi di
trasporto il controllo della tossicità del Ca2+. Con l’apertura dei canali vengono
attivate anche numerose proteine di legame al Ca2+, tra cui le protein chinasi
Ca2+-dipendenti o la calmodulina: si tratta di una piccola proteina altamente
conservata, recettore primario del calcio nelle cellule sia vegetali sia animali, che può trovarsi nei compartimenti citoplasmatici e nucleari e può essere legata alla membrana plasmatica. La molecola presenta quattro regioni o anse di legame per il Ca2+ e la presenza di due eliche su entrambi i lati delle anse dà origine al termine di proteina elica-ansa-elica; in seguito al legame con il Ca2+
la calmodulina va incontro a un cambiamento sostanziale della sua struttura terziaria esponendo una regione molto idrofobica, riconosciuta poi dai domini delle proteine bersaglio, che si legheranno e verranno attivate.
Il Ca2+ e la fosforilazione proteica da esso dipendente giocano un ruolo
importante nella regolazione dell’espressione dei geni cold-responsive e nell’acquisizione della tolleranza al gelo (Tahtiharju et al., 1997; Monroy et
al., 1993): è stato dimostrato, infatti, che gli shock termici provocano un
immediato aumento della concentrazione citosolica di Ca2+ nelle piante di
Arabidopsis resistenti alle basse temperature e ciò farebbe supporre che i flussi
di Ca2+ siano coinvolti nell’acclimatazione (Knight et al., 1996). Altri studi
hanno invece dimostrato che attraverso trattamenti con inibitori della calmodulina e con bloccanti dei canali del calcio (EGTA) si verificano nella cellula dei cambiamenti nello stato di fosforilazione di alcune proteine.
1.9 Trasduzione del segnale e acclimatazione
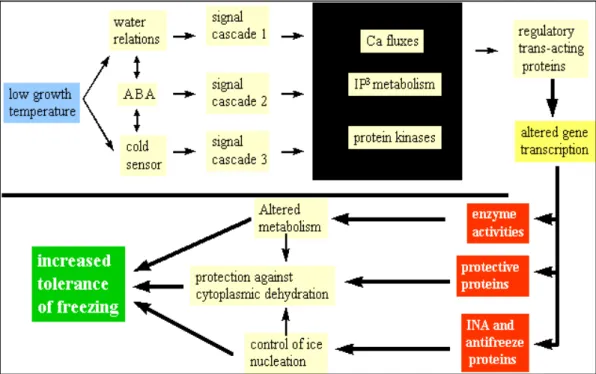
L’acclimatazione alle basse temperature coinvolge la sintesi di nuovi peptidi che, attraverso la loro attività enzimatica o le loro proprietà strutturali, conferiscono ai tessuti la tolleranza al congelamento. Nel 1970 Weiser propose che l’acclimatazione richieda l’attivazione trascrizionale di un pool di geni normalmente non espressi nelle cellule in condizioni non acclimatanti: secondo la sua teoria le basse temperature ed il fotoperiodo corto sono i responsabili dell’attivazione di una cascata di segnali che comporta delle modifiche nella trascrizione genica con conseguenti alterazioni delle attività enzimatiche e sintesi di proteine crioprotettive per permettere la sopravvivenza della pianta.
Figura 1.12: modello dei processi coinvolti nella cascata dei segnali, responsabile dell’acclimatazione e
dell’aumento della tolleranza al congelamento.
In effetti una grande quantità di geni sovra-regolati da trattamenti di acclimatazione sono stati recentemente isolati in specie sia monocotiledoni che
dicotiledoni. Gli studi molecolari hanno confermato che l’acclimatazione al congelamento è un processo complesso e, sebbene in alcune specie siano stati identificati geni con un effetto maggiore, solitamente la tolleranza alle basse temperature è un fenomeno multigenico. La reazione agli stress termici è diversa a seconda delle famiglie multigeniche: la temperatura alla quale i geni sono sovraespressi e quella responsabile della massima risposta variano, infatti, tra le specie, anche se la risposta tende gradualmente a crescere con l’abbassarsi delle temperature.
Nell’espressione di questi geni sono coinvolti controlli sia trascrizionali che post-trascrizionali; nel frumento lo sviluppo della tolleranza al congelamento è stato associato con l’espressione di specifici geni regolata a livello trascrizionale: Danyluk e Sarhan, infatti, hanno notato durante l’esposizione alle basse temperature la presenza di particolari mRNA indotti ed espressi ad alti livelli nelle cultivar tolleranti il congelamento ma non in quelle sensibili; con l’innalzamento delle temperature, il livello di espressione di tali mRNA ritorna a valori normali, mostrando una correlazione positiva con la capacità dei vari genotipi di sviluppare la tolleranza al congelamento (Danyluk e Sarhan, 1990).
Anche in Arabidopsis è stata individuata una regolazione trascrizionale di specifici geni coinvolti nell’acclimatazione: Thomashow e colleghi hanno isolato quattro geni COR i cui livelli di mRNA sono incrementati vertiginosamente durante le prime 4 ore di trattamento a 5°C; i livelli rimangono alti per l’intera durata del periodo di acclimatazione per cominciare a decrescere al momento dell’innalzamento della temperatura (Thomashow et
1.10 COR
I geni COR (cold-regulated), anche conosciuti con i nomi LTI (low
temperature-induced), KIN (cold-inducible), RD (responsive to desiccation) e
ERD (early dehydration-inducible), appartengono a quattro famiglie geniche, ognuna delle quali è composta da due geni che sono fisicamente associati nel genoma, avendo una disposizione a tandem; almeno un gene di ogni coppia è indotto in risposta alle basse temperature o altre condizioni associate a deficit idrici, come la siccità, l’alta salinità e l’ABA. Questi geni sono differenzialmente regolati; un esempio è rappresentato da COR15a e COR15b: sono due geni omologhi, aventi l’82% di sequenze nucleotidiche uguali; il livello dei loro trascritti aumenta in risposta alle basse temperature e in seguito alla somministrazione di ABA esogeno, ma solo quello di COR15a si accumula in risposta alla siccità. Discorso diverso vale per i geni LTI, che sono fortemente indotti in piante esposte alla temperatura di 4°C: in particolare i trascritti di LTI45 e di LTI30 si accumulano sia in risposta alle basse temperature che alla somministrazione di ABA e alla disidratazione. La vicinanza di questi geni nel genoma, la loro organizzazione a tandem e la quasi totale omologia nelle sequenze nucleotidiche fanno presupporre che questi geni si siano originati da un evento di duplicazione.
COR15a codifica una piccola proteina idrofilica di 15 kDa, codificata nel nucleo e indirizzata al cloroplasto; una volta importata nel cloroplasto viene processata per formare la proteina matura COR15am, un polipeptide di 9,4 kDa. Protoplasti ottenuti da piante transgeniche non acclimatate di
Arabidopsis, esprimenti costitutivamente il gene COR15a, mostrano una
maggiore tolleranza al congelamento sia in vivo che in vitro (Artus et al., 1996). I ricercatori hanno proposto che COR15a sia responsabile della stabilizzazione delle membrane andando a diminuire la loro propensione a formare lipidi in fase esagonale II in seguito al congelamento (Steponkus et
°C l’espressione di tale gene comporta un calo dell’incidenza della transizione dalla fase lamellare a quella esagonale II che avviene nelle regioni dove la membrana plasmatica entra in contatto con il cloroplasto a causa della disidratazione indotta dal gelo (Steponkus et al., 1998). L’inizio della transizione sembra essere determinato dalla membrana che ha la più forte propensione a formare la fase esagonale II: tra la membrana plasmatica e il cloroplasto l’“anello debole” è rappresentato proprio da quest’ultimo. Per spiegare il fenomeno relativo alla diminuzione del cambiamento tra una fase e l’altra si è ipotizzato che COR15am alteri la curvatura intrinseca della membrana del cloroplasto: COR15am presenta, infatti, delle regioni formanti α-eliche anfipatiche che stabilizzano le membrane contro la disidratazione associata al congelamento.
Il gene COR15a presenta un promotore che viene indotto dal freddo: la regione 5’ tra i nucleotidi -305 e +78 (in relazione all’inizio della trascrizione) contiene, infatti, uno o più elementi agenti in cis responsabili dell’espressione del gene regolata dalla temperatura (Baker et al., 1994). Questo promotore, che è debolmente attivo nella maggior parte dei tessuti e degli organi delle piante cresciute a temperature normali, viene fortemente attivato in risposta alle basse temperature, all’ABA e alla siccità.
1.11 CRT/DRE e CBF1
Molti dei geni regolati dal freddo contengono l’elemento CRT
(C-repeat)/DRE (drought responsive element), cioè un frammento di DNA
presente nei promotori dei geni COR e presumibilmente anche in quelli di geni
cold-regulated non ancora identificati. CRT/DRE è un elemento di regolazione
basse temperature e alla carenza idrica; a tale elemento si lega l’attivatore trascrizionale CBF1 (C-repeat binding factor; è anche detto DREBP, cioè DRE
binding protein): questo contiene un segnale di localizzazione nucleare (NLS),
una regione acida (che potenzialmente serve come dominio di attivazione) ed un dominio di legame al DNA di 60 amminoacidi che è stato trovato in altri fattori trascrizionali delle piante, come APETALA2, AINTEGUMENTA e TINY.
Numerosi studi hanno dimostrato l’appartenenza di CBF1 ad una famiglia genica codificante tre attivatori trascrizionali strettamente correlati: CBF1,
CBF2 e CBF3 (anche chiamati rispettivamente DREB1B, DREB1C e DREB1A) sono fisicamente associati in ripetizione sullo stesso cromosoma
(Gilmour et al., 1998). Il livello dei trascritti dei tre geni tende ad aumentare notevolmente nell’arco dei primi 15 minuti dopo l’esposizione delle piante alle basse temperature, mentre i trascritti dei geni COR si cominciano ad accumulare dopo circa due ore: Gilmour e colleghi hanno quindi proposto che l’espressione dei geni COR possa coinvolgere una cascata del segnale nella quale l’espressione del gene CBF rappresenta uno degli step iniziali; comunque l’induzione di CBF1 coinvolge a sua volta un attivatore definito ICE (inducer of CBF expression), che presumibilmente riconosce un elemento regolatore, l’“ICE Box”, presente nei promotori di ogni gene CBF. ICE solitamente è già presente nelle cellule alle normali temperature di crescita in uno stato inattivo, poiché è sequestrato nel citoplasma da una proteina regolatrice (HOS1) o poiché presenta una forma non in grado di legare il DNA o non in grado di attivare effettivamente la trascrizione; l’esposizione della pianta alle basse temperature implica una modificazione di ICE, che può quindi indurre l’espressione del gene CBF.
Figura 1.13:
rappresen-tazione schematica della cascata del segnale per l’acquisizione della tol-leranza al congelamento; SNOW è una proteina partner di ICE1.
Il gene CBF1 è stato clonato e la sua espressione costitutiva in Arabidopsis ha dato come risultato un aumento dell’abbondanza di tutti i trascritti dei geni COR in piante non acclimatate, che hanno poi mostrato un aumento della tolleranza al congelamento. La sovraespressione di CBF1 ha aumentato la tolleranza alle basse temperature più di quanto non facesse la sovraespressione del solo COR15a (Jaglo-Ottosen et al., 1998): da qui si deduce che CBF1 sia un importante regolatore dell’acclimatazione alle basse temperature, controllando il livello di espressione dei geni COR che, a loro volta, promuovono la tolleranza al congelamento. L’induzione del regulone CRT/DRE comporta dunque una più facile sopravvivenza della pianta se esposta alle basse temperature ma anche un incremento della tolleranza alla siccità (Liu et al., 1998). Tali risultati indicano ancor di più il coinvolgimento di geni regolati dal freddo nello sviluppo della tolleranza alle basse temperature ed aprono la via a ulteriori ricerche riguardanti la sovraespressione di fattori trascrizionali al fine di proteggere dal congelamento le piante coltivate.
1.12 Identificazione di geni per la tolleranza al
gelo con l’uso di mutanti
Sono stati compiuti molti studi sui mutanti di Arabidopsis sia per cercare di comprendere la funzione dei geni conosciuti sia per poterne individuare di nuovi: Warren e colleghi (1996) hanno screenato pool di semi M3 derivati da
piante M2 chimicamente mutagenizzate nel tentativo di individuare linee che
non mostrassero effetti negativi durante l’acclimatazione ma che poi non raggiungessero i normali livelli di tolleranza. Questi studi hanno permesso l’identificazione dei geni SFR (sensitivity to freezing) che sembrano avere un ruolo significativo nell’acclimatazione al freddo, in particolare SFR1, 2, 4, 5 e 6: in effetti analisi di electolyte leakage dimostrano che piante mutanti per tali geni risentono dei danni dovuti all’esposizione a basse temperature, in particolare quelli che intaccano la criostabilità della membrana plasmatica. Tali mutazioni comportano, infatti, la riduzione dell’accumulo di zuccheri (saccarosio e glucosio), antocianine e acidi grassi 18:1 e 18:2; la mutazione
sfr6 impedisce inoltre l’attivazione dei promotori CRT/DRE anche se
l’espressione dei geni CBF rimane invariata: questo suggerisce che il gene SFR6 aumenti l’espressione dei geni COR, essenziali per la tolleranza al congelamento, andando a modificare i geni CBF (McKown et al., 1996). Da questi dati si ipotizza che i geni SFR abbiano un ruolo importante nella regolazione dell’attivazione dell’acclimatazione al freddo.
Il mutante hos2 (high expression of osmotically responsive genes), così come sfr6, indebolisce la tolleranza al congelamento dopo l’acclimatazione alle basse temperature (Lee et al., 1998); si pensa che il gene HOS2 sia un regolatore negativo della cascata del segnale che si origina in seguito all’esposizione alle basse temperature, cioè eserciti un controllo a feedback che blocca la via: mutazioni recessive nel locus HOS2 accrescono, infatti, l’induzione dei geni responsabili dell’acquisizione della tolleranza al congelamento.
Anche Xin e Browse (1998) hanno sfruttato un approccio mutazionale in
Arabidopsis per identificare geni con importanti ruoli nell’acclimatazione alle
basse temperature, andando ad analizzare 800.000 piante M2 mutate
chimicamente alla ricerca di mutanti che mostrassero una costitutiva tolleranza al congelamento. Grazie a questi studi è stato identificato il gene eskimo1 (esk1): a differenza di piante wild-type, nelle quali la mortalità del 50% della popolazione (LT50) si aveva quando essa veniva esposta a temperature intorno
a −5,5°C, nelle piante mutanti esk1 non acclimatate tale valore si manifestava intorno a −10,6°C. La base molecolare dell’incremento della tolleranza al congelamento favorita dalla mutazione esk1 non è ancora certa, anche se si pensa che agisca come un regolatore negativo; quello che è certo è che in queste piante la concentrazione della prolina, molecola crioprotettiva, è più alta di trenta volte rispetto al wild-type acclimatato, mentre quelle degli zuccheri liberi di due. Xin e Browse hanno proposto l'esistenza di molteplici vie di trasduzione del segnale coinvolte nell’attivazione di differenti aspetti dell’acclimatazione al freddo (per esempio la via di CBF1 e quella di ESK1) e che una via può permettere una considerevole tolleranza al congelamento senza l’attivazione delle altre.
1.13 Proteomica
Il termine proteoma è stato coniato nel 1995 dall’australiano Marc Wilkins per indicare l’intero equivalente in proteine di un genoma: esso rappresenta, infatti, l’insieme di tutti i possibili prodotti proteici espressi in una cellula, incluse tutte le isoforme e le modificazioni post-traduzionali. Il proteoma è dinamico nel tempo poiché varia in risposta a fattori esterni e differisce sostanzialmente tra i diversi tipi cellulari di uno stesso organismo. La
proteomica diventa dunque la disciplina che studia il proteoma, il
complemento tempo-specifico e cellulo-specifico del genoma: tale disciplina riguarda lo studio su grande scala della proteina, in particolare delle sue strutture e delle sue funzioni. Il termine proteomica è stato coniato in analogia al termine genomica, campo rispetto al quale la proteomica rappresenta il passo successivo, essendo molto più complessa: infatti il proteoma differisce da cellula a cellula ed è in continua evoluzione nelle sue innumerevoli interazioni con l’ambiente e con il genoma, entità pressoché costante. Con la genomica si studia come il DNA di una cellula sottoposta a determinati stimoli possa esprimere i suoi geni in un contesto globale, non singolo: un fattore esterno induce nella cellula una serie di geni correlati a cascata e, di conseguenza, la sintesi di proteine che risponderanno a questa nuova situazione. Lo schema di reazione della cellula nei confronti di questi stimoli è DNA Æ mRNA Æ proteine: l’mRNA sintetizzato a partire dal DNA si traduce in proteine, le quali si esprimeranno in maniera diversa e in tempi diversi a seconda delle condizioni ambientali, dello stato fisiologico e della fase fenologica della pianta o degli organi in esame; la proteomica interviene in questo stadio, quando bisogna determinare quali, quante ed in che tempi le proteine si esprimono in relazione ad un determinato stimolo, ed è complementare alla genomica in quanto si focalizza sul prodotto del gene.
La proteomica ha compiuto un grosso balzo in avanti grazie alla tecnologia ed alla ricerca applicata nello studio di tecniche o strumentazioni, attraverso le quali negli ultimi anni è stato possibile rilevare quantità di proteine dell’ordine di picogrammi così come riuscire a separare quantità di proteine dell’ordine di milligrammi. Il termine proteomica, quindi, non fa riferimento ad una specifica tecnica di rilevazione, ma si avvale di più tecniche correlate le une alle altre per comprendere il prodotto finale, la proteina. Tra le tecniche di rilevazione bisogna ricordare l’elettroforesi bidimensionale, metodica di separazione delle proteine mediante un campo elettrico che, anche se utilizzata già dal 1970, rimane una tecnica di eccellenza. Una volta ottenuta questa mappatura, la
proteina o le proteine di interesse vengono eluite dal gel di corsa ed analizzate con l’aiuto della spettrometria di massa per determinare la loro identità; anche questa tecnica, relegata alla chimica stretta fino a qualche tempo fa, ha subito un notevole progresso tecnologico in questi ultimi anni. Oltre a queste due tecniche se ne utilizzano altre che consentono anch’esse il riconoscimento delle proteine e della loro struttura, come l’elettroforesi capillare o il sequenziatore di amminoacidi.
Naturalmente in un singolo esperimento si ottiene generalmente una consistente quantità di dati, la cui analisi computazionale (bioinformatica) è un passaggio essenziale per l’identificazione delle proteine, risultando dunque parte integrante degli studi proteomici: in questo senso un balzo in avanti è stato dato dalla creazione di enormi data-base, a cui i ricercatori possono rivolgersi per confrontare i loro dati, ed ottenere dei risultati probabilistici vicini al 100%.
1.14 Spettrometria di massa
La spettrometria di massa è una tecnica analitica di delucidazione strutturale basata sulla ionizzazione di una molecola e sulla sua successiva frammentazione in ioni di diverso rapporto massa/carica (m/z); a differenza delle tecniche spettroscopiche, però, questo è un metodo d’analisi distruttivo poiché la molecola non rimane intatta, e soprattutto non si basa sull’interazione tra radiazioni e materia.
Il principio su cui si basa consiste nella ionizzazione di una molecola per espulsione di un elettrone; il catione radicalico che si forma (ione molecolare) in parte si frammenta dando molecole e/o radicali neutri (che lo strumento non
rileva), in parte genera cationi. I vari ioni che si originano per frammentazione vengono discriminati sulla base del loro rapporto massa/carica e rivelati da un detector: si ottiene così lo spettro di massa, diagramma che riporta l’abbondanza relativa degli ioni in funzione del loro rapporto m/z, tipico di ogni composto in quanto direttamente correlato alla sua struttura chimica ed alle condizioni di ionizzazione cui è stato sottoposto.
Il campione viene ionizzato in un’apposita camera di ionizzazione, in cui il fascio di elettroni viene prodotto da una sorgente ionica che varia a seconda della tecnica utilizzata (impatto elettronico, ionizzazione chimica o electrospray). In genere gli elettroni sono emessi da un filamento caldo di tungsteno o renio e passano attraverso un condotto che crea il raggio, nella parte centrale della camera che contiene il campione gassoso.
La frazione di elettroni che non urta contro le molecole è raccolta da una trappola per gli elettroni, le molecole che non sono ionizzate sono allontanate mentre quelle ionizzate sono accelerate e convogliate verso l’analizzatore: questo consente di differenziare gli ioni generati in base al loro rapporto m/z. I più comuni sono l’analizzatore magnetico, quello a doppia focalizzazione, quello a quadrupolo e infine quello a tempo di volo (TOF): il principio su cui si basa quest’ultimo analizzatore è che ioni di differente valore massa/carica hanno uguale energia ma differente velocità dopo l’accelerazione subita nella camera di ionizzazione; ne deriva che il tempo che ciascuno mette per attraversare l’analizzatore è differente.
Come collettore e rivelatore degli ioni si usa comunemente un moltiplicatore elettronico, costituito da una serie di elettrodi in cascata: quando uno ione arriva sul primo elettrodo questo emette un fascio di elettroni che vanno a colpire il secondo elettrodo, il quale a sua volta emette una quantità maggiore di elettroni e così via: il risultato è una forte amplificazione del segnale che viene poi digitalizzato ed elaborato infine dal calcolatore dello spettrometro per la presentazione dello spettro di massa; l’uso dei calcolatori permette inoltre di combinare rapidamente la gestione dei parametri dello strumento con
la ricerca bibliografia in librerie di spettri in formato elettronico, in modo da automatizzare l’identificazione.
1.15 Arabidopsis thaliana
Arabidopsis thaliana è una pianta infestante molto diffusa sulla Terra; si
tratta di una piccola “erbaccia” annuale appartenente alla famiglia delle
Brassicaceae che cresce in quasi tutto l’emisfero boreale, dall’Artico
all’Equatore; è una pianta che tollera le basse temperature e che può crescere e maturare anche a 4-5°C (Schneider et al., 1995).
E’ stata scoperta in Germania sulle montagne Harz da Johannes Thal nel XVI secolo: il suo nome originario è Pilosella siliquosa ma successivamente fu denominata Arabidopsis thaliana in onore del suo scopritore (Meyerowitz, 2001).
A. thaliana è una comune specie erbacea dicotiledone priva di alcuna
importanza agronomica, tuttavia negli ultimi anni è diventata la base della ricerca sperimentale nei campi della genetica e della fisiologia vegetale molecolare (Meyerowitz and Somerville, 1994; Meinke et al., 1998).
Il primo a considerare A. thaliana come una pianta modello è stato Laibach in Europa; successivamente la pianta venne studiata in dettaglio da Rédei negli USA), ma è nei primi anni ’80 che Arabidopsis acquisì sempre più importanza grazie alla creazione di una dettagliata mappa genetica (Koornneef et al., 1983) e grazie a numerosi studi sottolineanti il valore della pianta nella ricerca nei campi della biochimica e della fisiologia vegetale, poiché:
- presenta un genoma di piccole dimensioni, costituito da 115-125 milioni di basi (circa 26.000 geni contenuti in cinque cromosomi), completamente sequenziato;
- attraverso analisi RFLP (Nam et al., 1989; Chang et al., 1988) è stato possibile creare per tutti e cinque i cromosomi mappe fisiche e genetiche (mappe che mostrano le distanze tra i geni in funzione della lunghezza del DNA);
- il suo ciclo vitale è corto (circa 8 settimane dalla germinazione alla produzione dei semi);
- la produzione di semi è molto prolifica;
- si coltiva facilmente in spazi ristretti grazie alle sue dimensioni (35-50 cm in altezza);
- può essere efficacemente trasformata utilizzando Agrobacterium
tumefaciens (Chang et al., 1994; Bechtold et al., 1993);
- è caratterizzata da una grande quantità di linee mutanti, importanti per molti studi genetici (Koornneef et al., 2004).
Alcune di queste caratteristiche, come le dimensioni contenute, il tempo di crescita veloce e il genoma ridotto, rendono Arabidopsis facilmente gestibile nell’analisi genetica; inoltre esiste una notevole somiglianza di molti suoi geni con geni di specie distanti dal punto di vista filogenetico, quali la colza da olio, il grano o il riso: la scoperta della locazione e della funzione di tutti i geni di
Arabidopsis permette quindi di studiare efficacemente i geni che controllano
gli stessi processi in piante da coltura più complesse (Szekeres et al., 1996; Schaller and Bleecker, 1995; Chang et al., 1993). Inoltre il genoma di
Arabidopsis contiene informazioni essenziali non solo per la conoscenza della
biologia vegetale ma anche per interessanti confronti con la struttura del Dna umano (Wei et al., 1998; Newman et al., 1994).
1.16 Scopo
Man mano che la popolazione umana aumenta, l’agricoltura deve nutrire un numero sempre maggiore di persone, ma deve comunque competere con lo sviluppo urbano per la disponibilità di terre coltivabili: questa crescente domanda, insieme alla diminuzione delle risorse, ha alimentato l’interesse sia scientifico sia commerciale verso la comprensione dei meccanismi attraverso i quali le piante rispondono agli stress.
L’impatto dell’ambiente sulla produttività vegetale diventa palese quando si paragonano le rese ottimali con quelle medie: se le prime possono essere considerate come rappresentative della crescita delle piante in condizioni ideali, allora le perdite associate con gli stress possono ridurre la produttività media del 65-87% a seconda del tipo di coltura; la maggior parte di queste perdite sono attribuite alle caratteristiche sub-ottimali.
Molte piante riescono ad aumentare la loro tolleranza al congelamento in seguito all’esposizione a basse temperature, fenomeno conosciuto con il termine di acclimatazione al freddo. L’interesse nello studio e nella comprensione delle basi molecolari di tale acclimatazione è dovuto non solo al desiderio di comprendere i meccanismi che le piante hanno evoluto per tollerare gli stress ambientali ma anche e soprattutto alla prospettiva che questa conoscenza possa permettere lo sviluppo di nuove strategie atte a garantire una tolleranza agli stress da parte di importanti piante di interesse agronomico: l’applicazione di tecniche biotecnologiche e di miglioramento genetico classico potrebbe portare, infatti, allo sviluppo di colture vegetali tolleranti lo stress che aumentino le riserve mondiali di cibo e forniscano consistenti benefici economici.
A tal fine ad oggi sono stati compiuti numerosi studi relativi a questo argomento (vedi Introduzione) e in quest’ottica si inserisce anche il presente lavoro: lo scopo è stato quello di individuare, attraverso un approccio proteomico, i meccanismi che permettono il conferimento della tolleranza alle
basse temperature ed in particolare al congelamento in Arabidopsis thaliana nella previsione di comprendere le basi di eventuali interventi atti a modificare questi meccanismi al fine di aumentare la produttività vegetale di piante di interesse agrario in condizioni ambientali non ottimali.
La scelta di Arabidopsis non è stata casuale: ormai da diversi anni tale pianta è infatti considerata una pianta modello per le sue caratteristiche biologiche e molecolari che hanno permesso il suo vasto utilizzo nella ricerca sperimentale nei campi della genetica e della fisiologia vegetale molecolare.
Capitolo 2
Materiali e metodi
2.1 Analisi preliminari
Dal momento che questo progetto non si è basato su risultati di precedenti lavori è stato necessario effettuare alcune analisi preliminari per poter valutare diverse condizioni di crescita delle piante e quindi individuare le condizioni migliori da utilizzare nel proseguimento del progetto.
I parametri analizzati operando attraverso prove incrociate sono stati: I parametro: età (giorni);
II parametro: acclimatazione; III parametro: temperatura;
IV parametro: durata del trattamento (ore).
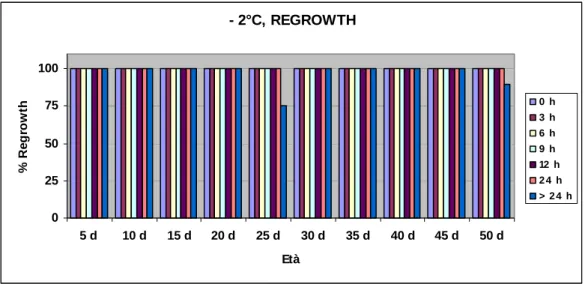
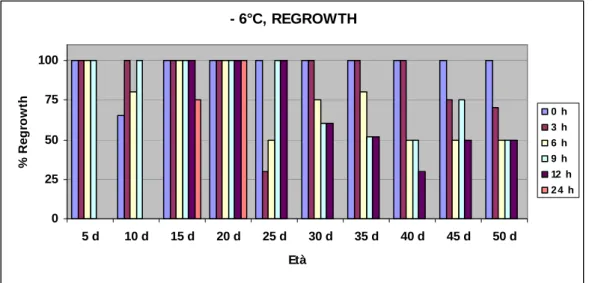
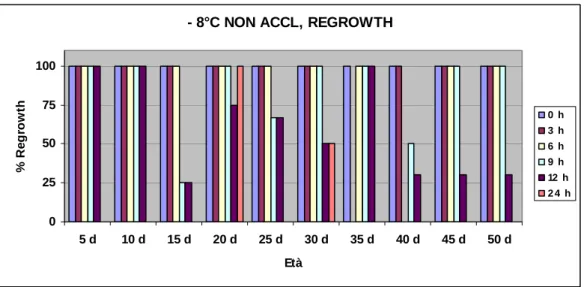
La variabilità dei risultati ottenuti è stata studiata effettuando analisi di
electrolyte leakage e di regrowth (tasso di sopravvivenza), oltre che analisi