Neurofisiologia del sistema nervoso centrale - 8 -
NEUROFISIOLOGIA DEL SISTEMA
NERVOSO CENTRALE
Il sistema nervoso è un complesso di formazioni interconnesse, che consentono di mettere in comunicazione fra loro le parti costituenti un organismo e l’organismo stesso con l’ambiente che lo circonda.
Nell’espletare le proprie funzioni il sistema nervoso è coadiuvato dall’apparato endocrino, apparato in grado di fornire stimoli lenti e ciclici di natura chimica. L'asse ipotalamo-ipofisario rappresenta la regione dove i due sistemi si incontrano, sia anatomicamente che funzionalmente.
Nel sistema nervoso possiamo individuare tre sottosistemi:
• sistema nervoso centrale, costituito da encefalo e midollo spinale
• sistema nervoso periferico, costituito da formazioni adatte alla trasmissione degli impulsi nervosi (nervi spinali, nervi encefalici, gangli)
• sistema nervoso vegetativo, costituito da formazioni nervose analoghe a quelle del sistema periferico, ma in grado di controllare la funzioni involontarie.
Il sistema nervoso centrale (SNC) è composto da due classi di cellule: le cellule della glia, che oltre a svolgere una funzione di sostegno sono responsabili dei fenomeni di sincronizzazione che si presentano tra le diverse aree della corteccia cerebrale, e le cellule nervose (neuroni), che rappresentano le unità fondamentali dell’intero sistema.
Il neurone
Il neurone è una cellula eccitabile in grado di ricevere, elaborare e trasmettere informazioni alle cellule adiacenti per mezzo di impulsi elettrici (potenziali d’azione o spike).
Strutturalmente il neurone presenta un corpo cellulare o soma, dal quale hanno origine delle arborizzaioni chiamate dendriti e un lungo prolungamento chiamato assone.
I dendriti sono delle brevi ramificazioni e costituiscono il corpo recettore principale della cellula, in quanto su oguno di essi, in regioni di contatto specializzate (sinapsi), terminano i prolungamenti delle altre cellule nervose.
Neurofisiologia del sistema nervoso centrale - 9 - L’assone ha invece come scopo principale la trasmissione dell’impulso nervoso, che si propaga al suo interno con una velocità che può raggiungere i 100m/s. Le proprietà di conduzione dell’assone sono legate esclusivamente alla caratteristica della membrana, rivestita in alcuni tratti da una guaina lipidica detta mielina.
Una possibilità per classificare i neuroni è analizzarne le caratteristiche morfologiche: i neuroni bipolari, ad esempio, hanno un corpo cellulare dal quale dipartono due soli prolungamenti, mentre i neuroni multipolari presentano un gran numero di arborizzazioni.
Figura 1.1: Neurone bipolare e neurone multipolare
Architettura del sistema nervoso centrale
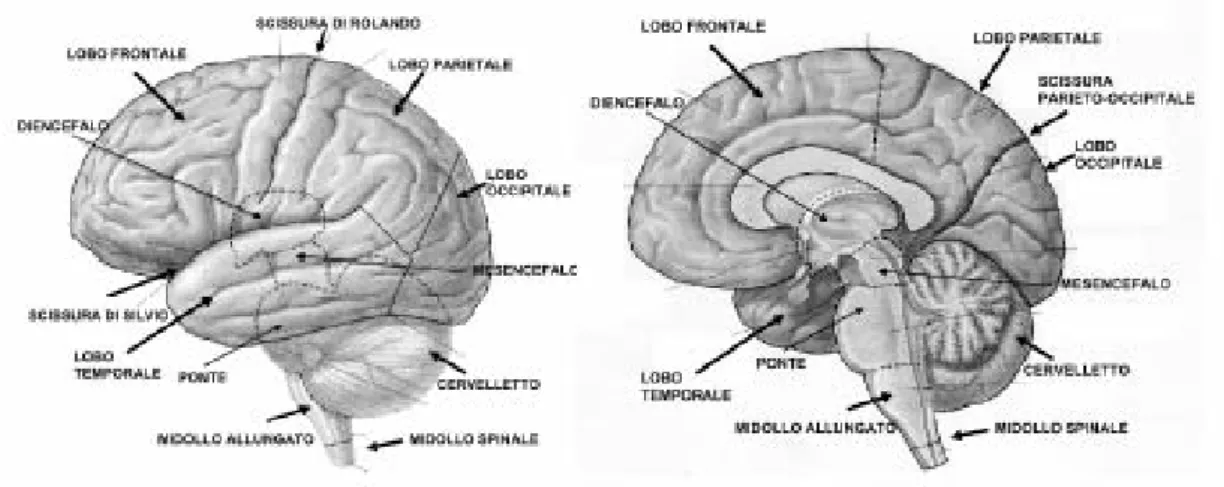
Come mostrato in figura 1.2, il sistema nervoso centrale è formato da un lungo cordone, genericamente indicato come asse nervoso, sormontato da tre formazioni nervose dette organi soprassiali. Antero-superiormente all'asse è situato il diencefalo, che non fa parte nè dell'asse nervoso nè degli organi soprassiali. I quattro quinti del diencefalo sono costituiti dal talamo, mentre nella parte inferiore del diencefalo troviamo l’ipotalamo
. L'asse nervoso percorre il canale midollare col nome di midollo spinale e prosegue per un breve tratto nella scatola cranica col nome di tronco encefalico. I tre organi soprassiali sono il cervelletto, la lamina quadrigemina e il telencefalo, suddiviso nei due emisferi cerebrali.
Il telencefalo è l’organo più voluminoso del sistema nervoso centrale, ed è costituito dai due emisferi cerebrali, di forma ovoidale, appaiati e separati dalla profonda scissura interemisferica.
Neurofisiologia del sistema nervoso centrale - 10 - Figura 1.2: Sistema nervoso centrale e alcune delle sue componenti
Gli emisferi cerebrali sono di colore grigio-rosato, essendo rivestiti in superficie da sostanza grigia, e sono collegati tra loro per mezzo del sistema di fibre trasversali che costituisce il corpo calloso, posto nella profondità della scissura interemisferica. Gli emisferi presentano una superficie irregolare per la presenza di rilievi e solcature, che separano parti funzionalmente molto importanti chiamate lobi (lobo frontale, parietale, temporale, occipitale, dell'insula e limbico): i rilievi più evidenti sono le circonvoluzioni, mentre le solcature più profonde sono chiamate scissure.
La corteccia cerebrale (figura 1.3) è un mantello di sostanza grigia che riveste la superficie degli emisferi. Si presenta ripiegata in giri o circonvoluzioni separate da solchi. I tre solchi principali sono la scissura centrale di Rolando, tra la corteccia frontale e quella
Neurofisiologia del sistema nervoso centrale - 11 - parietale, la scissura laterale di Silvio, tra la corteccia parieto-frontale e quella temporale e la scissura calcarina, sulla superficie mediale della corteccia occipitale.
Figura 1.3: Corteccia cerebrale e sistema nervoso centrale
A queste differenze morfologiche della corteccia fanno riscontro differenze funzionali: alle varie suddivisioni corticali è infatti possibile riconoscere una o più funzioni. In questo senso, la corteccia cerebrale è stata la protagonista di un notevole sviluppo storico, a partire da Gall fino ad arrivare all’attuale teoria dell’organizzazione modulare delle funzioni sensoriali.
Il sogno di capire cosa accade nella mente umana nasce infatti con lo studio della frenologia, teoria elaborata da F.G. Gall alla fine del XIX secolo, che individua sulla superficie del cervello una specie di mappa con specifiche protuberanze. Secondo tale corridoio gnoseologico, dall’esame morfologico della teca cranica era possibile risalire alla localizzazione delle funzioni psichiche di un individuo, quali l’istinto alla violenza, l’arguzia, l’istinto della proprietà. Quella di Gall fu una teoria molto seguita nell’Ottocento, tanto da comparire persino in un racconto di H. de Balzac, “Papà Goriot”.
Questa teoria, per quanto sicuramente non del tutto adeguata ed ormai completamente superata, spianò la strada al panorama delle localizzazioni cerebrali, e ad alcuni studiosi, quali P. Broca e C. Wernicke, individuarono sulla corteccia cerebrale le sedi del linguaggio parlato e scritto.
Oggi abbiamo molte più informazioni sulla corteccia; sappiamo che essa è costituita da circa 10 miliardi di neuroni e diversi miliardi di sinapsi e sulla sua superficie, sia
Neurofisiologia del sistema nervoso centrale - 12 - nell’emisfero destro che in quello sinistro, sono state individuate particolari aree funzionali: corteccia motoria, corteccia somatosensoriale, centri che ricevono informazioni relative alla vista, all’udito, al gusto e all’olfatto. A ciascuna di queste aree sensoriali è collegata un’area di associazione, ed è dalla cooperazione tra centri riceventi e relative aree di associazione che si produce la nostra percezione sensoriale.
Esistono poi aree di associazione che sono la sede delle più sofisticate attività mentali, come per esempio l’area di associazione frontale, che raccoglie le informazioni provenienti da molte altre aree del cervello e produce valutazioni, giudizi, progetti, ciò che comunemente chiamiamo 'pensiero'.
Un’altra interessante peculiarità del nostro cervello è rappresentata dalla diversità funzionale tra i due emisferi: mentre nell’emisfero sinistro si trovano i centri del linguaggio e, più in generale, le capacità logiche e matematiche, l’emisfero destro è sede dell’astrazione e delle capacità artistiche e musicali, vale a dire la creatività.
Il premio Nobel R. Sperry ha effettuato negli anni Sessanta un gran numero di ricerche su individui con emisferi separati: si trattava di pazienti epilettici a cui veniva reciso il corpo calloso, cioè la struttura di collegamento tra i due emisferi. Contrariamente a quanto si possa pensare, questa operazione determinava un netto miglioramento delle condizioni di questi pazienti per ciò che riguardava l’epilessia; tuttavia, sottoponendo questi individui a semplici test, essi si comportavano come se fossero state due persone distinte. A seconda del tipo di stimolo veniva attivato l’uno o l’altro dei due emisferi e si osservavano perciò risposte diverse: se per esempio questi pazienti venivano bendati e si chiedeva loro di riconoscere al tatto una chiave e di usarla, questi sapevano usare la chiave se si trovava nella mano sinistra, ma non riuscivano a darle un nome. Tutto ciò è spiegabile se si pensa che l’informazione proveniente dalla mano sinistra va all’emisfero destro, che è muto, in quanto il centro del linguaggio è situato nell’emisfero sinistro.
In realtà con il progredire degli studi sul cervello si è visto che esso possiede una forma di plasticità, che gli permette di riorganizzare e ridistribuire le funzioni corticali: si è osservato, per esempio, che dopo l’amputazione di una parte del corpo, l’area corticale collegata a essa stabilisce nuove connessioni con le altre parti.
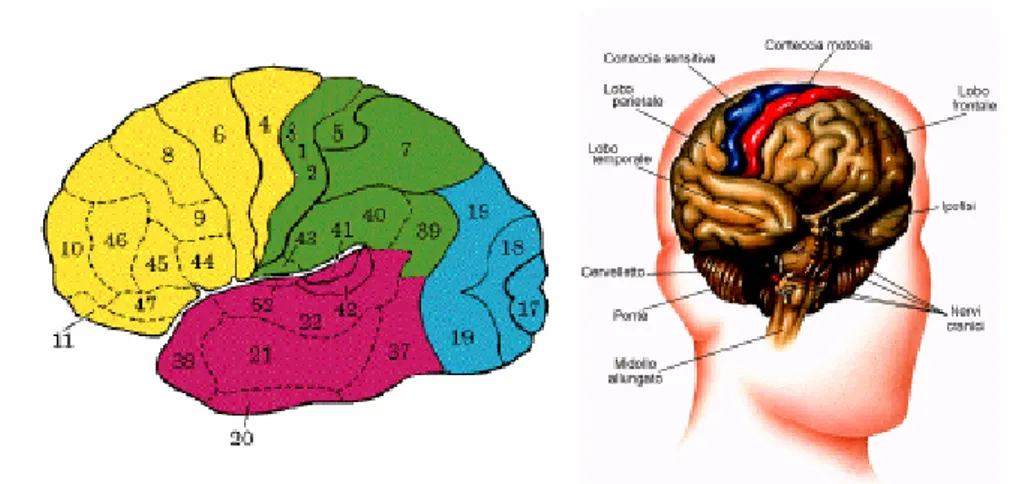
Agli inizi del secolo Brodman (figura 1.4) localizzò sulla superficie corticale circa 50 aree funzionali, e tale suddivisione è tuttora in uso. Come vedremo in seguito, le aree vengono divise in tre grandi gruppi, motorie, sensitive e associative. Tuttavia, studi recenti hanno
Neurofisiologia del sistema nervoso centrale - 13 - dimostrato come non sia possibile parlare di vera e propria segregazione funzionale all’interno di queste aree.
Figura 1.4: Schema della superficie dell’emisfero sinistro umano, con la suddivisione in lobi e l’indicazione numerica delle aree funzionali di Brodman.
Le funzioni che caratterizzano l’uomo e lo differenziano dagli altri animali, dipendono proprio dalle interazioni esistenti tra queste parti della corteccia cerebrale e tra la corteccia e le altre regioni del cervello.
A livello istologico una via sensoriale può essere considerata semplicemente come un insieme di neuroni disposti in serie. Si può così riconoscere una suddivisione della corteccia in sei strati cellulari (figura 1.5) con spessori differenti.
Dalla superficie verso l'interno possiamo così distinguere lo strato molecolare (o plessiforme), lo strato granulare esterno, lo strato piramidale esterno, lo strato granulare interno (caratterizzato da piccoli neuroni dal soma sferoidale), lo strato piramidale interno, lo strato polimorfo.
In tutti gli strati sono presenti neuroni associativi che collegano le cellule sia all'interno dei singoli strati che tra i diversi strati. Si deve a questo punto ricordare che i granuli sono le cellule che ricevono le informazioni da altre aree corticali o sottocorticali, mentre le cellule piramidali e le cellule polimorfe sono i neuroni il cui neurite esce nella sostanza bianca, andando a formare i fasci di associazione (tra le diverse aree corticali), i fasci di correlazione (tra organi soprassiali diversi), i fasci motori.
Neurofisiologia del sistema nervoso centrale - 14 - Figura 1.5: Comparazione tra le distribuzioni dei diversi tipi cellulari,
identificati con l’impregnazione argentica e con colorazione per la mielina
Mantenendo un certo schematismo descrittivo, si può affermare che le aree sensitive hanno molto sviluppato lo strato dei granuli, a differenza delle altre aree: ad esempio la corteccia della circonvoluzione precentrale è agranulare, ma per converso possiede un ben sviluppato strato di cellule piramidali giganti.
Le aree corticali
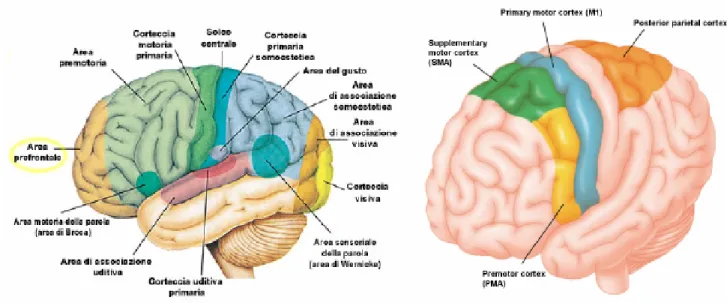
Nella corteccia cerebrale si riconoscono alcune unità anatomiche e funzionali denominate "aree corticali" (figura 1.6). Esse si distinguono in aree sensitive (primarie e secondarie), aree motorie (primarie e secondarie) e aree associative. Le aree sensitive primarie sono le seguenti:
• area sensitiva somatica: corrispondente alla circonvoluzione postcentrale, dove gli impulsi esterocettivi vi vengono proiettati con organizzazione somatotopica (omuncolo sensitivo)
• area visiva: corrispondente ai margini superiori e inferiori della scissura calcarina, a livello della faccia interemisferica dei lobi occipitali
Neurofisiologia del sistema nervoso centrale - 15 - • area acustica: corrispondente ad una regione a livello del margine inferiore della scissura laterale di Silvio (lobo temporale), che comprende anche l’area di Wernicke per la comprensione del linguaggio parlato
• area olfattiva: corrispondente alla corteccia piriforme del lobo temporale
Figura 1.6: Aree corticali
Un'area motoria primaria è invece l’area motrice somatica, dalla quale dipartono con una distribuzione somatotopica (omuncolo motorio) le fibre del sistema piramidale. L'area motrice somatica è una tipica corteccia agranulare e comprende i centri motori del linguaggio: l'area di Exner per il linguaggio scritto e l'area del Broca per il linguaggio parlato.
I cosiddetti omuncoli motorio e sensitivo (figura 1.7) sono due diagrammi utili per rappresentare l'estensione delle aree motorie e sensitive sulle circonvoluzioni pre e postcentrali del telencefalo: ambedue le strisce di corteccia mostrano una rappresentazione somatotopica della periferia, cioè ogni parte dell'apparato locomotore e della superficie corporea si proietta in una precisa porzione della circonvoluzione.
Tuttavia, poichè l'ampiezza delle diverse porzioni corticali non corrisponde alle dimensioni delle corrispondenti parti periferiche, ma è legata all’entità della loro innervazione, disegnando il diagramma di esse, si ottiene un corpo umano mostruoso, con mano e testa (soprattutto labbra e lingua) smisuratamente grandi.
Neurofisiologia del sistema nervoso centrale - 16 - Figura 1.7: Omuncolo motorio e sensitivo
E’ importante notare come l’attività mentale è integrata e coordinata con l’attività cinesica: mentre una persona parla, pensa a ciò che vuole dire, al fine di ottenere una risposta coerente ed olista.
Il legame causale tra attivazione neurale e responso mentale è il frutto di un meccanismo complesso di correlazione ed elaborazione fra le diverse strutture cerebrali. Tutto ciò mette in evidenza quanto sia importante la connettività, le connessioni anatomiche tra le varie aree corticali, e la memoria. La memoria infatti consente di rispondere in modo adeguato ad una nuova esperienza, ricordando il passato e associando esperienze diverse (memoria associativa). Tutto questo può esplicarsi in modo cosciente o non cosciente: la memoria dell’andare in bicicletta è ad esempio eterogenea, motoria, vestibolare, dal momento che non facciamo alcuno sforzo per ricordare le dinamiche motorie e di equilibrio che utilizziamo.
Immaginazione motoria ed emozioni
Quando un essere umano pensa di eseguire un movimento corporeo senza svolgerlo, effettua un compito di immaginazione mentale, che porta all’attivazione della stessa area motoria che si accenderebbe nel caso dell’espletazione di un movimento reale.
Neurofisiologia del sistema nervoso centrale - 17 - Se ad esempio facciamo compiere ad una persona un particolare movimento velocemente e ripetiamo l’operazione, chiedendo però di riflettere sul movimento da compiere, confrontando i due pattern di “accensione” neurale relativi ai vari segmenti cerebrali, osserveremo che nel secondo caso si ha un incremento di attività nell’emisfero coinvolto nel meccanismo motorio, con un conseguente responso dell’azione più accurato.
Consideriamo invece la programmazione motoria di un tennista mentre aspetta la battuta dell’avversario: è possibile notare l’attivazione della corteccia visiva, ma soprattutto dell’amigdala e dell’ipotalamo, che rivestono un ruolo importante nelle emozioni; tutto ciò dimostra che il tennista attiva delle strutture emotivamente coinvolte nella pianificazione dell’atto motorio. Le emozioni ed il movimento sono infatti intimamente correlati.
La corteccia prefrontale è sede della formazione e pianificazione degli obiettivi, dell’insight, cioè dei fenomeni dell’ideazione; la corteccia orbito-frontale è invece sede delle capacità introspettive. Essa è suddivisa in corteccia orbito-frontale-mediale, che si occupa del processamento degli stimoli emotivi, e corteccia orbito-frontale-laterale, che abbina il processamento emotivo a quello cognitivo.
Queste regioni della corteccia sono estremamente importanti nel controllo sociale: nei criminali non pentiti, nei serial killer o negli hoolygans, l’attivazione della corteccia orbito-frontale-mediale è spesso alterata a causa di lesioni, e provoca così responsi comportamentali a livello sociale inadeguati. L’aumento di aggressività può essere poi associato ad una mancata modulazione da parte dell’amigdala. È dimostrato infatti che l’amigdala si attiva associativamente con una serie di psicopatologie, quali l’ansia, l’agorafobia, la claustrofobia, l’aracnofobia, gli attacchi di panico.
L’amigdala è un piccolo nucleo nervoso a forma di mandorla che fa parte del sistema limbico; un insieme di nuclei e fibre nervose coinvolti nelle emozioni e situati al di sotto della corteccia cerebrale. È comunque sempre la corteccia cerebrale che analizza le informazioni complesse, codifica memorie e apprendimenti, dà significati alle esperienze e le integra in una trama concettuale. Il sistema limbico, invece, ha un’origine più antica in termini di storia naturale, è un retaggio del passato che ci accomuna, per modalità di reazione, alle emozioni di altre specie animali.
Secondo Antonio Damasio, gran parte delle emozioni dipendono soprattutto dall’amigdala e non da altri nuclei del sistema limbico: emozioni di base come paure e rabbia sarebbero legate a questa piccola formazione nervosa che reagisce ai segnali di allarme
Neurofisiologia del sistema nervoso centrale - 18 - provenienti dagli organi di senso, scatenando reazioni nell’organismo e bombardando di stimoli la corteccia cerebrale, che viene così coinvolta solo in seconda battuta.
La peculiarità dell’amigdala, come indicano numerosi studi sul comportamento animale e umano, deriva dalle caratteristiche delle sue connessioni: questo nucleo nervoso riceve infatti numerose fibre di recettori uditivi e visivi ed è quindi in grado di rispondere immediatamente ai messaggi che denunciano una situazione di pericolo, producendo reazioni di paura o rabbia. Queste reazioni sono molto più rapide di quelle che dipendono dalla corteccia, dal momento che quest’ultima viene informata solo in un tempo successivo. Gli stimoli che generano paura, come un richiamo di allarme o un urlo di terrore, vengono infatti istradati verso la corteccia dopo esser pervenuti all’amigdala ed è a questo punto che la corteccia può analizzare, paragonare, razionalizzare.
La conferma è emersa da una serie di sperimentazioni che hanno coinvolto situazioni di pericolo tra gatti e topi: le emozioni primarie come la paura, sono fondamentalmente governate dall’amigdala, cosicché in alcuni pazienti colpiti da lesioni di questa formazione nervosa, la nozione di pericolo è quasi inesistente, proprio come avviene negli animali in cui l’amigdala è stata messa fuori gioco.
Joseph LeDoux, professore di neurobiologia della New York University, osserva: “Si tende a confondere emozioni e sentimenti. Le emozioni sono funzioni biologiche che si sono evolute per permettere agli animali di sopravvivere in un ambiente ostile e riprodursi. I sentimenti invece sono il prodotto della coscienza, etichette soggettive che l’uomo attribuisce alle emozioni inconsce”. Il neuroscienziato conclude che occorre evitare i sentimenti, impossibili da studiare oggettivamente, per concentrarsi invece sulle emozioni e sulla loro base biologica, i cui circuiti neurali sono tangibili quanto quelli dei meccanismi sensoriali.
Secondo LeDoux, quindi, “l’amigdala è specializzata nel reagire ad un particolare stimolo e nel provocare una risposta fisiologica…. Molto diverso invece è il sentimento cosciente della paura, attivato da un secondo percorso che collega “l’orecchio” dell’amigdala alla corteccia cerebrale. Qui il segnale minaccioso viene analizzato in dettaglio, usando informazioni provenienti anche da altre parti del cervello…”.
Quali sono le conseguenze di tutto questo? Se si tratta di un falso allarme, la corteccia cerebrale cercherà di arrestare la sensazione di paura, anche se il soggetto avrà pur sempre avvertito l’iniziale stato di allerta: è per questo motivo che una persona sobbalza alla vista di
Neurofisiologia del sistema nervoso centrale - 19 - un serpente in un sentiero. Se però il serpente si rivela un semplice pezzo di legno, ecco che la paura subito svanisce.
Gli input provenienti dalla corteccia cerebrale aiutano quindi a vincere l’istintivo timore: tuttavia, come sottolinea LeDoux “le connessioni neurali di ritorno dalla corteccia all’amigdala sono molto meno sviluppate di quelle di andata, dall’amigdala alla corteccia. È dunque maggiore l’influenza dell’amigdala sulla corteccia, che non viceversa, e perciò spesso stentiamo a controllare razionalmente le nostre emozioni”.
Concludendo, ispirandoci alle idee oliste di Damasio, non possiamo parlare di segregazione delle funzioni all’interno delle mente umana, non esiste una vera e propria area specifica con funzioni specifiche. Il cervello non è solo un atlante geografico di continenti differenziati: i confini tra emozione e cognizione sono in realtà lievi e le dinamiche funzionali globali del cervello non possono essere studiate tassonomicamente come entità individuali e discrete cablate in un insieme di connessioni sinaptiche.
In questa prospettiva assumono un ruolo di rilievo gli studi di connettività anatomica e connettività funzionale, volti proprio ad investigare le dinamiche organizzative spazio-temporali che possono essere direttamente associate alla coscienza dell’individuo.
I segnali bioelettrici
Per lo studio dell’attività elettrica cerebrale si utilizza come grandezza fisica di riferimento il potenziale elettrico.
Partiamo dal presupposto che in quasi tutte le cellule dell’organismo è possibile rilevare dei potenziali elettrici di membrana, prodotti dalla differenza di concentrazione degli ioni sodio, potassio e cloro tra citoplasma e mezzo extracellulare.
In condizione di riposo il potenziale transmembrana delle cellule nervose è di circa -70 milliVolt, e può essere calcolato sommando i potenziali nerstiani delle tre specie ioniche sopra citate.
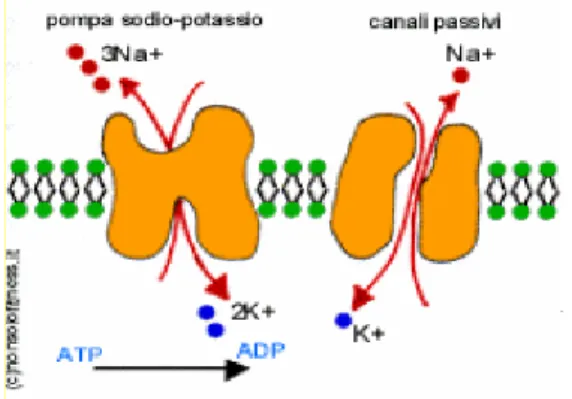
Questo potenziale ha origine per una serie di fattori, primo dei quali il contributo di una proteina pompa sodio-potassio, che si occupa della circolazione dei due ioni tra l’interno e l’esterno della cellula, mantenendo costante il gradiente di concentrazione (vedi figura 1.8).
Neurofisiologia del sistema nervoso centrale - 20 - La comunicazione tra le cellule nervose avviene mediante una variazione rapida del potenziale di membrana seguita dal ritorno ad una condizione di riposo (potenziale d’azione).
La forma e l’ampiezza del potenziale d’azione variano in base alle caratteristiche del tessuto eccitabile, ma durante la fase di propagazione vengono conservate per l’intera lunghezza della fibra e non sono proporzionali all’entità dello stimolo che le ha generate: si parla infatti di risposta tutto o nulla, a seconda se lo stimolo sia in grado o meno di scatenare quei meccanismi che generano il potenziale d’azione.
Figura 1.8: Pompa sodio potassio e canali passivi
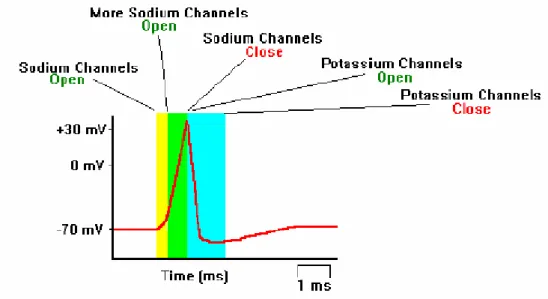
Gli impulsi di corrente che possono modificare il potenziale di membrana vengono classificati come depolarizzanti, se portano il potenziale ad un valore meno negativo, o iperpolarizzanti, se portano il potenziale ad un valore più negativo. Per innescare il meccanismo del potenziale d’azione è necessario uno stimolo in grado di alzare il potenziale di membrana a -60 mV.
Ciò che accade può essere così schematizzato:
• fase di depolarizzazione: la membrana della cellula diviene permeabile agli ioni sodio, che entrando provocano una depolarizzazione. Nel momento in cui viene superato un valore soglia di -60 mV, si ha un’inversione del potenziale fino a raggiungere +50 mV. Se la depolarizzazione è troppo lenta, la soglia può essere superata senza che si verifichi un potenziale d’azione; in questo caso si parla di accomodazione, durante la quale i primi canali sodio attivati si richiudono a causa del troppo tempo trascorso.
Neurofisiologia del sistema nervoso centrale - 21 - • fase di ripolarizzazione: alla richiusura dei canali sodio segue l’apertura dei
canali potassio, che facilitano l’uscita di tale ione. La membrana ritorna così verso i valori del potenziale di riposo con la stessa velocità con cui si era depolarizzata. • fase di iperpolarizzazione: terminata la fase di ripolarizzazione, a causa della
lenta cinetica di chiusura dei canali potassio, il potenziale di membrana raggiunge i -90 mV prima di tornare ai valori di riposo.
Figura 1.9: Potenziale d’azione
Grazie agli esperimenti condotti da Hodgkin e Huxley negli anni Cinquanta, si può ricondurre l’andamento del potenziale d’azione a quello delle conduttanze dei tre ioni precedentemente elencati.
Durante la fase di ascesa del potenziale, la conduttanza del sodio (g Na+ ) aumenta molto
rapidamente, raggiungendo il massimo circa nello stesso istante in cui il potenziale d’azione raggiunge il suo apice.
La conduttanza del potassio (g K+ ), aumenta invece in modo più graduale, raggiunge il
suo massimo circa alla metà della fase di ripolarizzazione e ritorna più lentamente ai livelli di riposo. La conduttanza del cloro, o più in generale del termine di lekeage, può essere infine considerata costante.
Neurofisiologia del sistema nervoso centrale - 22 - La durata complessiva del potenziale d’azione è di circa 6 millisecondi; tuttavia è possibile distinguere due intervalli temporali in cui la membrana si comporta in modo differente. Duranti i primi 3-4 ms, includenti la fase di depolarizzazione, la membrana si trova infatti in uno stato di refrattarietà assoluta, ovvero se pur sollecitata con altri stimoli non è in grado di generare un nuovo potenziale d’azione; negli ultimi 2-3 ms, coincidenti con la fase di ripolarizzazione, lo stato della membrana è di refrattarietà relativa ed essa può generare un nuovo potenziale d’azione solo se lo stimolo applicato è molto più intenso del precedente.
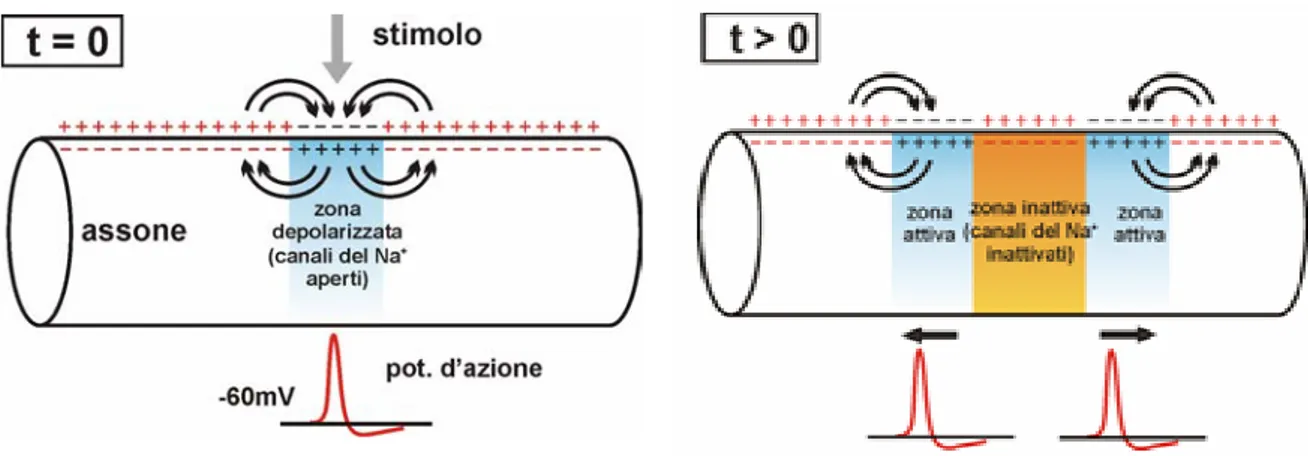
Una volta generato, il potenziale d’azione si propaga lungo il nervo o la fibra nervosa sfruttando i flussi ionici, mediante una conduzione di tipo elettrotonico. Questa conduzione ha inizio in una zona in cui vi sono delle differenze di potenziale, date da accumulo di carica negativa sulla superficie esterna della membrana. Come mostrato in figura 1.10, per compensare le variazioni locali di potenziale, si generano dei flussi di corrente, che provocano a loro volta la depolarizzazione delle aree contigue.