Unione Europea
UNIVERSITÀ DEGLI STUDI DI SALERNO
Dottorato di ricerca in
Biochimica e patologia dell’azione dei farmaci X ciclo nuova serie
2008-2012
BAG3 role in cardiomyocytes physiopathology
Dottoranda
Margot De Marco
Tutor Ch.mo Prof. Maria Caterina Turco
Index
i
Abstract Abstract ... ….1 Sommario Sommario... …2 Chapter I. Cardiomyocytes 1 Cardiomyocytes ... 31.1 Cardiac muscle organogenesis ... 4
1.1.1 Molecular signals in embryonic development of the heart ... 5
1.1.2 Myogenesis ... 6
1.2 Cardiomyopathies ... 7
1.2.1 Tako-tsubo cardiomyopathy ... 7
1.3 Extracellular signalling ... 8
1.3.1 Mechanisms of protein’s release in the extracellular space ... 9
1.3.2 Exosome secretion pathway ... 9
Chapter II. BAG3 protein 2 BAG3 protein ... 13
2.1 BAG3 and apoptosis ... 15
2.2 BAG3 in development ... 16
2.3 BAG3 role in cell adhesion and motility ... 19
2.4 BAG3 in autophagy ... 20
Chater III. Analisys of BAG3 role in cardiomyocytes physiopathology 3.1 Introduction ... 23
3.2 Results and Discussion ... 23
3.2.1 BAG3 role in process of cardiomyocytes differentiation ... 23
3.2.2 BAG3 induction in response to oxidative stress ... 26
3.2.3 Polymorphisms of BAG3 could have a role in the pathogenesis of Tako-tsubo cardiomyopathy ... 30
3.2.4 Release of BAG3 protein by stressed cardiomyocytes ... 32
3.2.5 Detection of soluble BAG3 and anti-BAG3 antibodies in patients with chronic heart failure ... 35
3.3 Conclusion ... 38
Chapter IV. Materials & Methods 4.1 Cell cultures ... 41
4.2 Blood samples ... 41
4.3 Antibodies ... 41
4.4 Western blot analysis ... 41
Index
ii
4.6 Purification of exocytic vesicles by differential ultracentrifugation ... 42
4.7 Quantitative RT-PCR ... 42
4.8 Genetic analysis ... 43
4.9 Plasmids construction and bacterial expression ... 43
4.10 Confocal microscopy ... 44
4.11 Dissociation of anti-BAG3 antibodies in human sera ... 44
4.12 Measurement of antibody titers by ELISA ... 45
4.13 NO2- assay ... 45
4.14 IL6 detection by ELISA ... 45
4.15 Statistical analysis ... 45 References
References………...………...I-XIII Acknowledgements
Abstract
- 1 -
Abstract
The anti-apoptotic protein BAG3 is expressed at high levels in skeletal and cardiac muscle in vivo. Our group recently focused its interest on BAG3 role in myocardiocyte proliferation, survival and response to stressful stimuli. We found that BAG3 is upregulated during the differentiation of cardiomyoblasts. Our results prompted us to verify whether bag3 silencing could affect the differentiation state of cardiocytes and we found that bag3 silencing resulted in highly reducing the levels of myogenin. Furthermore, we analyzed BAG3 expression and localization following cell exposure to oxidative stress. In particular, we found that epinephrine in vitro increases BAG3 expression in adult human cardiomyocytes. We evaluated whether BAG3 could be involved in the Tako-tsubo cardiomyopathy (or stress cardiomyopathy) pathogenesis that is characterized by left ventricular dysfunction, with symptoms that can mimic an acute coronary syndrome. The absence of significant cardiovascular risk factors in patients affected by stress cardiomyopathy suggested that it might be associated with a possible genetic etiology. Therefore, we sequenced bag3 gene to check for polymorphisms in 29 patients and 1043 healthy donors. Three polymorphism were highly represented among patients (R71Q, C151R, P407L).
We also showed for the first time that BAG3 protein is released from stressed cardiomyocytes and is found in chronic heart failure (HF) patients’ sera. Since anti-BAG3 antibodies are also present in patients’ sera, we developed an ELISA test for their specific detection. In serum samples from chronic HF patients, we found significantly higher values of anti-BAG3 antibodies respect to samples from healthy donors. The presence of anti-BAG3 antibodies in chronic HF patients’ sera and the availability of an ELISA test for their detection can contribute a novel tool for diagnostic and prognostic evaluations.
Sommario
- 2 -
Sommario
BAG3 è una proteina citoplasmatica di 74 kDa particolarmente concentrata nel reticolo endoplasmatico rugoso. In condizioni normali, l’unico tessuto che esprime significativi livelli di bag3 è il muscolo striato (muscolo scheletrico e cuore), dove la proteina è localizzata a livello dei dischi Z e sostiene la sopravvivenza e l’attività contrattile del muscolo, sebbene non sia essenziale per lo sviluppo dello stesso. Il nostro gruppo ha recentemente focalizzato la propria attenzione sul ruolo di BAG3 nella proliferazione, la sopravvivenza e la risposta allo stress dei cardiomiociti. Abbiamo dimostrato che BAG3 si esprime nel corso del differenziamento in vitro dei cardiomioblasti. I nostri risultati ci hanno spinto, quindi, a verificare se il silenziamento di bag3 potesse influenzare lo stato di differenziazione dei cardiociti ed abbiamo scoperto che esso causa riduzione dei livelli di miogenina. Inoltre, abbiamo analizzato l’espressione e la localizzazione cellulare di BAG3 dopo esposizione a stress ossidativo. In particolare, abbiamo trovato che l'adrenalina in vitro aumenta l’espressione di BAG3 in cardiomiociti umani. Abbiamo valutato se BAG3 potesse essere coinvolta nella cardiomiopatia Tako-tsubo (o cardiomiopatia da stress), la cui patogenesi è caratterizzata da una disfunzione del ventricolo sinistro e si manifesta con sintomi che possono simulare una sindrome coronarica acuta. L’assenza di significativi fattori di rischio cardiovascolare nelle pazienti colpite da questa forma di cardiomiopatia ci ha fatto ipotizzare che potesse essere associata ad un’eventuale causa genetica. Pertanto, abbiamo sequenziato il gene bag3 per verificare la presenza di polimorfismi in 29 pazienti e 1.043 donatori sani. Tre sono i polimorfismi più rappresentati tra i pazienti (R71Q, C151R, P407L). Abbiamo anche dimostrato per la prima volta che la proteina BAG3 viene rilasciata da cardiomiociti sottoposti a stress e può essere rilevabile nel siero di pazienti affetti da insufficienza cardiaca cronica. Dal momento che, nel siero dei pazienti è stata rilevata anche la presenza di anticorpi anti-BAG3, abbiamo sviluppato un test ELISA per la loro determinazione specifica. Nei campioni di siero dei pazienti abbiamo trovato valori significativamente più elevati di anticorpi anti-BAG3 rispetto a campioni di siero provenienti da donatori sani. La presenza di anticorpi anti-BAG3 nel siero dei pazienti affetti da scompenso cardiaco cronico e la disponibilità di un test ELISA per la loro individuazione possono rappresentare un nuovo strumento per le valutazioni diagnostiche e prognostiche di tale patologia.
Chapter I
Cardiomyocytes
- 3 -
1. Cardiomyocytes
Cardiomyocytes represent the largest population of cardiac muscle tissue, which is the most important structure of the heart, the myocardium, responsible for its contraction. Cardiocytes are elongated, generally about 100 m in length and 10-15
m thick. At the center of the cell is the nucleus, always unique, provided with one or two nucleoli, of ovoid shape, with the major axis disposed along the length of the cardiocyte. In the para position is observed a small-nuclear Golgi apparatus that plays an important role in the biogenesis of the sarcoplasmic reticulum. Mitochondria are bulky and numerous (occupy 30-40% of the volume of the sarcoplasm), have a shape roughly cylindrical and ridges highly developed, and they are arranged in a row between the myofilaments. The abundance of mitochondria indicates how the myocardial tissue is dependent from aerobic oxidative metabolism and, therefore, molecular oxygen. Under conditions of basal metabolic rate, the cardiomyocyte gets the energy it needs, in the form of ATP molecules, almost all processes of aerobic oxidative metabolism of fatty acids, glucose and lactic acid. Each cardiocyte possesses myofibrils organised and the alignment of the sarcomeres gives the cardiac muscle tissue a typical striated appearance. These cells are interconnected by means of junction specialised structures, known as intercalated disks or scalariform striae (fig. 1).
Chapter I
Cardiomyocytes
- 4 -
The gap junctions are scattered along the transverse section of the striae scalariform, but are much larger in size along the longitudinal section. It is in virtue of these areas that the passage of the pulse by another cardiocyte occurs. For this same reason, from the functional point of view, the myocardium is considered a syncytium. To the spread of the electrical pulse to the entire cardiac muscle mass then follows the mechanical event, that is, the contraction of its cavity, before those atrial and, immediately after, those ventricular.
1.1 Cardiac muscle organogenesis
The genesis of the heart in vertebrate embryos begins immediately after gastrulation when three germ layers are trained in the primitive streak: ectoderm, mesoderm, and endoderm. (Garcia-Martinez & Schoenwolf, 1993) (fig. 2).
Figure 2 – The three embryonic germ layers.
The cells of the germ layers can be defined primitive multipotent, because each of them can give rise to a number of different cell populations, characterised by its own morpho-functional physiognomy. By means progressive modifications, cells may further narrow their multipotency, to reach the unipotency, which coincides with the differentiated stage. The heart derives from mesoderm, which is divided into three portions: paraxial, intermediate and lateral. Mesothelial membranes originate own from the lateral mesoderm, the cardiac and smooth muscle that constitutes visceral muscles. Cardiac progenitor cells are mostly localised inside the primitive streak, in the same order antero-posterior wherein after are found in the heart (Garcia-Martinez & Schoenwolf, 1993). As development proceeds, the precursor population of precardiac mesoderm emigrates from the streak in an antero-lateral direction, giving rise to the heart-forming fields on either side of the streak (Filipczyk et al., 2007). The
Chapter I
Cardiomyocytes
- 5 -
real differentiation of primitive cardiomyocytes begins almost simultaneously with the merger of the endocardial tube. The cardiac tube, in humans, starting from the twentieth day, begins to contract in a non-synchronous, but peristaltic. When forming this tube, primitives myocardial cells begin to express genes that encode for characteristic proteins of the myocardium as myosin, actin, troponin and other components of the contractile apparatus. In muscle cells developing start to appear the sarcomeric structures and then the myofibrils, so the cardiac muscle tissue begins to contract rhythmically.
Despite being physically and electrically coupled through the intercellular junctions, cardiac cells differentiated, unlike the skeletal muscle cells, do not merge into multinucleated myotubes, but retain their individuality. In addition, cardiac muscle cells already differentiated continue to divide during fetal development, coming out of the cell cycle after birth. This greatly differs from what happens in the development of skeletal muscle, in which differentiation, characterised by the activation of muscle-specific genes, coincides with the exit from the cell cycle (Standring et al., 2009).
1.1.1 Molecular signals in embryonic development of the heart
The process of formation of the heart into an embryo developing requires some signals, some of which have their origin outside the pre-cardiac mesoderm; others have origin into the heart developing. Three families of peptide growth factors have been studied most intensely for their positive (and negative) effects on the establishment of cardiac cell identity. These are the bone morphogenetic proteins (BMPs), members of the superfamily of transforming growth factor (TGF-β), protein Wnts and fibroblast growth factors (FGFs) (Olson & Schneider , 2003). The BMPs control some of the major cardiac-specific transcription factors, including Nkx 2-5 (Brand, 2003; Liberatore, 2002; Lien , 2002). With regard to Wnt, were initially considered suppressive proteins of the heart training, but has been reported both an action of induction that of suppression: canonical Wnt 1, 3A and 8 appear to inhibit cardiac differentiation, while Wnt 11 does not seem to increase cardiac differentiation (Olson & Schneider 2003). Finally, some studies of chicks and zebrafish have indicated a role cardio-inductive by FGFs (Lough et al., 1996; Barron et al., 2000; Alsan & Schultheiss 2002). In Drosophila have been found two receptors for FGF and only one is expressed in mesoderm during gastrulation and early differentiation: it is therefore required for normal cardiac development (Narasimha & Leptin 2000, Michelson et al., 1998).
These three signaling pathways activated by ligands BMP, Wnt and FGF are essential not only for the primary cardiogenesis, but are also involved in secondary cardiogenesis (Brand, 2003).
Once the cells of the mesoderm have received appropriate signals, are activated a series of cardiac transcription factors that act on downstream genes involved in the pathway heart. The transcription factor Nkx2.5 (Lints et al., 1993) and the T-box protein TBX5 (Bruneau et al, 1999, Horb & Thomsen 1999) are among the first markers of cardiac lineage and are activated immediately after the cells have formed cardiac tissues. Nkx2.5 and TBX5 are associated with zing finger transcription factors, members of the GATA family and with the serum response factor (SRF), to activate genes of cardiac structures such as actin, the light and heavy chain of myosin, troponin and desmin (Bruneau et al., 2001, Habets et al., 2002). Even the transcription factors of the family Myocyte Enhancer Factor 2 (MEF2) play a key role
Chapter I
Cardiomyocytes
- 6 -
in the differentiation of cardiomyocytes (Wang et al., 2001). So there are multiple interactions between different transcription factors to control differentiation and maturation of early cardiomyocytes.
1.1.2 Myogenesis
Myogenesis is the process that leads to the formation of a mature muscle fiber, starting from undifferentiated precursors. During myogenesis, the precursors are transformed into myoblasts which in turn merge to become cells multinucleated said myotubes. Subsequently, the myotubes differentiate into real muscle fibers (Taylor & Jones, 1979). Although it is expected that in the future are identified the cardiac counterparties of transcription factors involved in the differentiation of skeletal muscle, the corresponding factors in cardiac myogenesis are still unknown (Standring et al., 2009). To date, four transcription factors have been cloned that regulate myogenesis: Myf5, MyoD, myogenin and MRF4 (fig.3).
Figure 3 – Illustration of skeletal muscle differentiation. Myoblasts, proliferative, mononucleated cells committed to becoming a skeletal muscle cell, differentiate to myotubes, long, multinucleated, tube-like cells responsible for muscle contraction.
These factors, for their secondary structure, belong to the superfamily of proteins bHLH (basic helix-loop-helix). The HLH domain is responsible for the dimerization of these proteins with other transcription factors, proteins E (E12, E2, E47), while the basic domain interacts with consensus sequences E-box (CANNTG) contained in the promoter of muscle-specific genes such as myosin, and creatine kinase (Perry et al., 2000). While MyoD and Myf5 are the factors involved in the early stages of myogenesis, in particular in the transition from undifferentiated precursors to myoblasts (Rudnicki et al. 1993), the myogenin and MRF4 regulate the late phases, respectively, the passage from myoblasts to myotubes and from myotubes to mature fibers (Edmonson & Olson , 1993; Olson & Klein, 1994). The myogenin is in turn regulated by another transcription factor MEF2 (muscle enhancer-binding factor 2), (Olson, 1992) already mentioned before among the factors that regulate embryonic development of the heart.
Chapter I
Cardiomyocytes
- 7 -
1.2 CardiomyopathiesThe cardiomyopathies are defined as a group of diseases in which the primary abnormality is one of the heart muscle. They are idiopathic by definition, and must be distinguished from the specific heart muscle diseases that have an identifiable cause, such as a systemic hypertension.
The cardiomyopathies are categorised according to their morphologic and haemodynamic characteristics. There are five groups: hypertrophic, dilated, arrhythmogenic, restrictive or unclassified cardiomyopathies. They all cause significant morbidity and mortality, and are among the most common genetically transmitted cardiac disorders.
Hypertrophic cardiomyopathy (HCM) is inherited. It is characterised by disorganised cardiac myocytes and unexplained left ventricular (LV) hypertrophy due to mutations in the genes encoding sarcomeric proteins, such as cardiac beta-myosin heavy chain gene, troponin and alpha-tropomyosin.
Dilated cardiomyopathy (DCM) is characterised by an LV ejection fraction <45 per cent (normally 55-70 per cent) with increased (dilated) LV dimension. The condition is hereditary in 30-40 per cent of cases (usually autosomal dominant); however, it can be caused by acute viral (usually entero-/adenoviruses) myocarditis leading to chronic inflammation, ventricular remodelling and dysfunction.
Restrictive cardiomyopathy (RCM) is sub-classified into primary (Loeffler's endocarditis, endomyocardial fibrosis) and secondary (infiltrative causes: amyloidosis, sarcoidosis; storage disorders: haemochromatosis, glycogen storage disorder, Fabry's disease; post-radiation).
Arrhythmogenic right ventricular cardiomyopathy (ARVC) is caused by fibro-fatty replacement of right ventricular (RV) myocytes due to apoptosis, inflammation (definite causes for either mechanism are as yet unknown) or a genetic cause (familial in 30-50 per cent usually with autosomal dominant inheritance).
Unclassified cardiomyopathy includes left ventricular non-compaction (LVNC) and Tako-tsubo cardiomyopathy.
LVNC is caused by embryogenic arrest of normal myocardial maturation causing a loose meshwork of non-compacted myocardial fibres with deep recesses communicating with the LV cavity.
Tako-tsubo cardiomyopathy predominantly affects women and is due to catecholamine surges from physical or emotional stress, causing coronary vasospasm and severe apical, mid-LV dysfunction (Elliott et al., 2008).
1.2.1 Tako-tsubo cardiomyopathy
Tako-tsubo cardiomyopathy (TTC) or Stress-cardiomyopathy is an acute cardiac condition characterised by transient and reversible myocardial stunning leading to systolic left ventricular apical ballooning in absence of obstructive coronary artery disease. Patients often present with chest pain, have ST-segment elevation on electrocardiogram, and elevated cardiac enzyme levels consistent with a myocardial
Chapter I
Cardiomyocytes
- 8 -
infarction. Generally, it occurs in post-menopausal women and is triggered by emotional or physical stress (Citro et al, 2012; Kurisu et al., 2002; Tsuchihashi et al., 2001). It was first described by Satoh et al. (Satoh et al., 1990) and was named “Takotsubo”-shaped cardiomyopathy due to its unique “short neck round-flask”-like LV apical ballooning resembling the tako-tsubo (Japanese for octopus trap) of Japan (fig. 4).
Figure 4 – "Tako-tsubo" is the japanese name for octopus traps that fishermen still use to catch octopus. In
this syndrome, the left ventricle during systole shows apical ballooning akinesis with basal hyperkinesis, taking the shape of an octopus trap (tako-tsubo).
The long-term prognosis of TTC patients is favourable due to spontaneous recovery of myocardial function within days or weeks. Up to date the pathogenetic mechanism is not well known (Akashi et al., 2005). Coronary microvascular dysfunction in TTC has already been demonstrated by using both invasively and non-invasively methods (Litvinov et al., 2009). Furthermore women with a history of TTC
demonstrated an impaired endothelium-dependent vasodilation, excessive
vasoconstriction, and augmented sympathetic activation after experiencing acute mental stress compared with age-matched post-menopausal controls and patients with previous myocardial infarction (Martin et al., 2010).
The association of this syndrome with an acute stressful event, particularly in older women, has suggested to investigators a pathophysiologic role for the sympathetic nervous system (Ueyama et al., 2002). Indeed, substantial elevations of plasma catecholamines (epinephrine, norepinephrine, and dopamine) have been reported in patients with stress cardiomyopathy (Wittstein et al., 2005). Single nucleotide polymorphisms of the beta1 and alpha 2c adrenergic receptors result in enhanced myocyte receptor function and enhanced synaptic norepinephrine release and theoretically could result in harmful sympathetic nervous system overactivity (Small et al., 2002). It is therefore reasonable to consider such genetic variation within the sympathetic nervous system as a predisposing factor for this cardiomyopathy.
1.3 Extracellular signalling
A key process in the development, growth, differentiation, function and potentially the repair and regeneration of tissues is cell communication via paracrine,
Chapter I
Cardiomyocytes
- 9 -
autocrine and endocrine signalling (Prabhu, 2004). Such intercellular signalling is of particular importance in the heart, where it is required for normal cardiac development and function (Kakkar and Lee, 2010) and where it plays a central role in remodelling and potential repair of damaged and diseased myocardium (Lionetti, et al. 2010).
Cell-to-cell communication is enabled in many different ways via direct cell-cell contact and soluble mediators. However, another way for cell-cells to communicate is via the release of membrane enclosed vesicles that transport signals and information, protected from the extracellular environment, from donor- to recipient cell locally and/or at a distance.
1.3.1 Mechanisms of protein's release in the extracellular space
The best known mechanism for the release of proteins by cells is the classical endoplasmic reticulum (ER)-Golgi pathway: secreted eukaryotic proteins utilize an N-terminal signal peptide (SP) to direct their co-translation on ER-bound ribosomes into the ER lumen, after which they progress through the endomembrane system and are ultimately exported to the extracellular environment, or cell surface. In addition to this, there are other non-classical or alternative pathways that have been reviewed in detail recently (Nickel, 2003) and are typical of "leaderless" proteins such as the heat shock proteins. In some cases the release of these proteins involves cell lysis and can happen both in pathological conditions that lead to necrosis, both during the release of cytokines, physiologically regulated (Wewers , 2004). A second pathway involves the release of intracellular proteins by secretory vesicles (MacKenzie et al., 2001). The heat shock proteins such as HSP27, HSP90 and HSP70/HSC70 can evidently be issued within the lumen of exosomes, when, for example, B cells are exposed to a heat shock (Clayton et al., 2005). A third secretion pathway involves the entry of the leaderless protein into secretary lysosomal endosomes, migration of these organelles to the cell surface and release of the contents of the endolysosome into the extracellular space (Baraldi et al., 2004).
1.3.2 Exosome secretion pathway
Exosomes are membrane vesicles released into the extracellular environment upon exocytic fusion of multivesicular endosomes with the cell surface (Février & Raposo, 2004), (fig. 5).
Chapter I
Cardiomyocytes
- 10 -
Figure 5 – Schematic representation of the endocytic pathway.
The release process appears to require, first, sorting into the small vesicles contained in endosomes and, second, the fusion of the limiting membrane of these endosomes with the cell surface, resulting in the release of these small vesicles into the extracellular space (Harding et al., 1984; Pan et al., 1985; Johnstone et al., 1987). These small vesicles, which measure about 50–90 nm in diameter, are termed ‘intraluminal vesicles’ (ILVs) while they are contained within multivesicular endosomes and ‘exosomes’ when they are released into the extracellular environment. Multivesicular endosomes, commonly called multivesicular bodies (MVBs) on the basis of their morphology (Sotelo, 1959), have well-known functions as intermediates in the degradation of proteins internalised from the cell surface or sorted from the trans Golgi network: so proteins destined for degradation are sorted into the ILVs of the newly forming MVB, which then fuse with pre-existing lysosomes (Futter et al., 1996). Pioneering studies on reticulocytes demonstrated that MVBs and their ILVs can also, as an alternative fate, fuse with the plasma membrane to eliminate ‘obsolete’ proteins that do not follow an intracellular degradation pathway (Johnstone et al., 1991).
The identification of vesicles as exosomes is based on both morphological and biochemical criteria. Given their small size, exosomes can only be visualised by electron microscopy.
For further morphological and biochemical characterization, exosomes can be isolated from the cell culture medium by differential ultracentrifugation. The supernatants of viable cell cultures are centrifuged at low speed to remove cell debris and other cellular contaminants due to cell lysis. Exosomes, like any small membrane vesicle, are pelleted at 100 000 g. (Wubbolts et al., 2003). To remove soluble proteins that may associate aspecifically with exosomes, additional purification can be accomplished using the flotation of the membranes pelleted through a continuous gradient of sucrose (Raposo et al., 1996). Exosomes float at a density around 1.13 g / ml, but this may vary from cell to cell depending on the exosome protein content (Stoorvogel W et al., 2002). Recently an alternative method for obtaining highly purified exosomes has been developed, based on a process of ultrafiltration and centrifugation in sucrose-deuterium oxide cushions (Lamparski et al., 2002). Only
Chapter I
Cardiomyocytes
- 11 -
after purification, exosomes can be characterised morphologically and by the identification of protein markers.
About the biochemical characteristics of exosomes, the protein composition reflects that of the cells from which they are released. About the biochemical characteristics of exosomes, the protein composition reflects that of the cells from which they are released. Among the glycolipid and protein components of exosomes we can find: MHC class II and class I molecules, adhesion molecules (integrins and tetrasparine), transport and membrane fusion molecules (RAP1B/RABGDI, Rab 7, Rab 2 and annexin), heat shock proteins (Hsc70, HSP84/90), cytoskeletal proteins (actin, cofilin, tubulin, myosin), raft-associated proteins and glycolipids (flotilin, CD55, CD59, GM1, GM3, Gi2α), enzymes (pyruvate kinase, alpha-enolase) (Février & Raposo, 2004).
Recent studies have led to the hypothesis that exosomes may represent a novel method of intercellular communication (Stoorvogel et al., 2002; Thery et al., 2002). Nonetheless, there are as yet no experimental indications of how exosomes interact with their target cells. Different modes of interaction can be envisioned for different cell types, and these may be directly related to their functions: exosomes could fuse with the plasma membrane or they could be endocytosed via an as yet unknown mode of internalization. They are often released as small aggregates that could be taken up by neighbouring cells via a phagocytic mechanism. It cannot be excluded that exosomes simply bind to the cell surface, conferring new properties to the target cell (Denzer et al., 2000). Also unknown are the mechanisms regulating MVB fusion with the cell surface and thereby the secretory process itself. The small GTPases of the Rab superfamily are obvious candidates to regulate MVB docking and fusion, such as Rab 11 in reticulocytes or Rab 27 and Rab 3 (Savina et al., 2002; Tolmachova et al., 2004).
Exosomes may have regulatory functions in the immune system and their application in cancer immunotherapy is promising. Several studies, in fact, have shown that exosomes display functional MHC class II and class I as well as co-stimulatory and cell adhesion molecules, bringing out a particular interest in their use as acellular vehicles for stimulation of anti-tumoral immune responses in vivo (Chaput et al., 2004). It has also been suggested the possible use of exosomes as a vehicle to enter the cells, by some pathogens such as retroviruses and in particular HIV (Pornillos et al., 2002).
In the cardiovascular system extracellular vesicles with different cellular origins have been identified. It is now known that both cardiac fibroblasts (Bang et al., 2012) and cardiomyocytes (Gupta & Knowlton, 2007) possess the ability to release and communicate via extracellular vesicles. In cardiovascular disease smooth muscles cells in the vascular system release extracellular vesicles that may be involved in vascular calcification (Kalra et al., 2012). It has also been shown that exosomes derived from mesenchymal stem cells have been found to reduce infarction size in a mouse model of myocardial ischemia (Lai et al., 2010).
Chapter II
BAG3 protein
- 13 -
1. BAG3 protein
BAG proteins are a family of co-chaperones, that interact with the ATPase domain of the heat shock protein (Hsp) 70 through a specific structural domain known as BAG domain (110-124 amino acids) (Takayama et al.,1999).Members of the family are found throughout evolution, in yeast (Saccharromyces cerevisiae, Schizosaccharromyces pombe), invertebrates (Caenorhabditis elegans, Ciona intestinalis, Drosophila), amphibians (Xenopus laevis), mammals (humans, mice), plants (Oryza sativa, Arabidopsis thaliana) (Takayama et al, 1999; Takayama and Reed, 2001; Coulson et al., 2005; Doukhanina et al., 2006; Colinet and Hoffmann, 2010), suggesting a fundamental biological role.
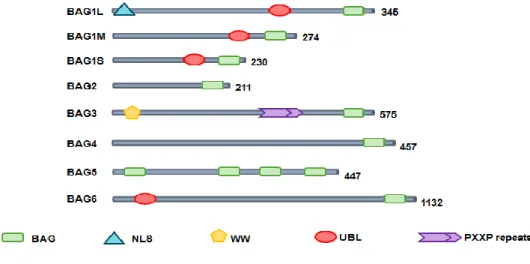
BAG3 was originally identified by yeast two-hybrid screening using the ATPase domain of the heat shock protein (Hsc/Hsp) 70 as bait (Takayama et al., 1999). In addition to the BAG domain, BAG3 contains a WW domain and a proline-rich repeat (PXXP) (fig. 6) that mediate binding to other partners (Takayama et al., 1999; Doong et al., 2000; Beere, 2005).
Figure 6 – Human BAG family proteins. All six reported BAG proteins contain a BAG domain at their C-terminus. Some BAG proteins contain other domains including nuclear localization signal (NLS), ubiquitin-like (UBL) domain,WWdomain and proline-rich regions (PXXP). Numbers next to the linear peptide sequence indicate amino acid number in the proteins.
Two BAG3 forms have been described so far: one is the full-length product of the bag3 gene having an apparent mass of 74 kDa; the other one is a shorter BAG3 protein found in association to neuronal synaptosomes (Rosati et al., 2011; Bruno et al., 2008) (fig. 7).
Chapter II
BAG3 protein
- 14 -
Figure 7 – Schematic representation of the full-length BAG3 protein and the shorter BAG3 protein found to be associated to synaptosomes.
The BAG3 full length protein is localized in the cytoplasm, mainly concentrated in the rough endoplasmic reticulum; upon cell exposure to stressors, a slightly different molecular weight variant of this form can also be observed, and both co-exist in some cell types and run as a doublet in a standard Western Blot. The origin of this doublet is currently unknown, but it could derive from post-traslational modifications as phosphorylations, indeed BAG3 protein contains several serine- rich motifs and ten tyrosine residues. Tyrosine phosphorylation of BAG3 occurs upon EGF stimulation in human breast cancer cell lines (Young et al., 2008). This post-traslational variant of BAG3 protein appears also to be associated to Btk protein in Defew cells upon oxidative stress induction (Rosati et al., 2009). Recently a shorter form of BAG3 (40 kDa) has been characterized by immunoprecipitation from neural synaptosomes homogenates and successive mass spectrometry (Bruno et al., 2008), again it is yet unknown if this form derives from alternative splicing or proteolitic processing.
In humans, bag3 gene expression is constitutive in myocytes, a few other normal cell types and several primary tumours or tumour cell lines (lymphoid or myeloid leukemias, lymphomas, myeloma, neuroblastoma, pancreas, thyroid, breast and prostate carcinomas, melanoma, osteosarcoma, kidney, colon and ovary cancers, glioblastoma) (Rosati et al., 2007; Pagliuca et al., 2003; Rosati et al., 2007b; Rosati et al., 2009; Chen et al., 2004; Ammirante et al., 2010). It is instead induced in different normal cell types (leukocytes, epithelial and glial cells, retinal cells) by a variety of stressors, such as oxidants, high temperature, serum deprivation, heavy metals, HIV-1 infection, ELF exposure, electrophile stress, haemodialysis treatment, pulsed ultrasound, retinal light damage, kanaic acid-induced seizure, transient forebrain ischemia (Rosati et al., 2007; Pagliuca et al., 2003; Rosati et al., 2007; Rosati et al., 2009; Ammirante et al., 2010; Basile et al., 2011, Festa et al., 2011). The expression of stress- responsive genes is regulated by the heat shock transcription factors (HSFs), including HSF1, that is required for tumour initiation and maintenance in a variety of cancer models (Franceschelli et al., 2008). Other known transcription factors that are known to regulate bag3 expression are: Egr1 (Gentilella et al., 2008), AibZIP (Androgen-Induced bZIP) (Ben et al., 2007), and WT1 (Wilms' tumor suppressor gene) (Cesaro et al., 2010).
A number of drugs - proteasome inhibitors, TNF-related apoptosis-inducing ligand (TRAIL), fludarabine, cytosine arabinoside, etoposide gemcitabine– increase BAG3 protein levels, this in turn contributes to cell resistance to therapy, and indeed bag3 silencing improves neoplastic cell apoptotic response to drugs (Romano et al.,
Chapter II
BAG3 protein
- 15 -
2003; Chiappetta et al., 2007; Liu et al., 2009; Ammirante et al., 2010, Rosati et al., 2012).
2.1 BAG3 and apoptosis
A number of studies in tumour cell lines of different origin have shown that bag3 silencing or hyperexpression results in, respectively, enhancing or inhibiting spontaneous or drug- induced apoptosis (Doong et al., 2002; Chiappetta et al., 2007; Rosati et al., 2007; Liu et al.2009; Jacobs et al., 2009; Wang et al., 2009; Lee et al., 2002; Ammirante et al., 2010; Jung et al., 2010; Cesaro et al., 2010).
BAG3 seems to influence cell survival by interacting with different molecular partner thus activating multiple pathways. A first demonstrated mechanism of BAG3 anti-apoptotic activity is mediated by its role, as a co-chaperone, in protein delivery to the proteasome. Indeed, while another member of BAG family, i.e. BAG1, positively cooperates with Hsp70 and CHIP (C-terminus of the Hsc70-interacting protein) to direct, through its ubiquitin-like domain (Fig.1 – Figure 9), client proteins to proteasome; BAG3 can interfere with this process by competing with BAG1 (Rosati et al., 2007; Coulson et al., 2005; Gentilella et al., 2008; Du et al., 2008). Indeed, in osteosarcoma and melanoma cells, BAG3 protects IKKgamma from proteasome delivery and these results in sustained NF-kB activation and cell survival (Ammirante et al., 2010). A different mechanism has been observed in glioblastoma cells, where BAG3 retains BAX protein in the cytosol, preventing its mitochondrial translocation (Festa et al., 2011). Both mechanisms rely on an interaction between BAG3 and Hsp70 (Festa et al., 2011; Ammirante et al., 2010).
We can speculate that through its binding to Hsp70, BAG3 might also positively or negatively modulate folding of other apoptosis- regulating proteins, and expect that future research will disclose a very complex regulative mechanism mediated by this protein. Moreover, since HSP70 can bind to AU-rich elements (AREs) in the 3’-untranslated regions, regulating expression of a number of proteins, including the well-known pro-apoptotic, BH3 only protein Bim (Matsui et al., 2007), BAG3 might be expected to regulate Hsp70 ability to stabilize Bim mRNA and possibly other mRNAs, involved in various cell functions. Finally, we can envisage functions of BAG3 that are independent of Hsp70, since it could also bind some client proteins through its WW- or PXXP domain, directly influencing their stability, localisation or activity. It is likely that the availability of the different partners underlies the different mechanisms through which BAG3 exerts its anti-apoptotic activity in different cell types (fig. 8).
Chapter II
BAG3 protein
- 16 -
Figure 8- Mechanism of BAG3 activity – models. (Rosati et al., 2011).
2.2 BAG3 in development
In some cell types, bag3 expression appears to be developmentally regulated. In developing rat central nervous system (CNS), there is a transient expression, detectable by immunohistochemistry, of BAG3 in the cerebral cortex and hippocampus, while a considerable expression is maintained in the rostral migratory stream and the subventricular zone of the lateral ventricle; there is an abrupt increase of BAG3- positive neurons in the cortex and hippocampus during the first postnatal week, which declines thereafter. Two specific populations of BAG3- positive neurons can be identified in the developing rat forebrain (Choi et al., 2006). Furthermore, recent results indicated that BAG3 is expressed in neural progenitors and sustains proliferation, mainly in response to FGF2, in those cells (Gentilella et al., 2010). In addition, an early transient expression of BAG3 was observed in midline radial glia in
Chapter II
BAG3 protein
- 17 -
the developing rat brainstem and spinal cord (Choi et al., 2009). Together, these pieces of evidence indicate a role for BAG3 in the development of both the neuronal and glial components of central nervous system. In agreement with the hypothesis of BAG3 involvement in CNS development, an altered BAG3 expression was observed in the cerebellum of hypothyroid juvenile mice and was suggested to contribute to impaired development of the hypothyroid brain.
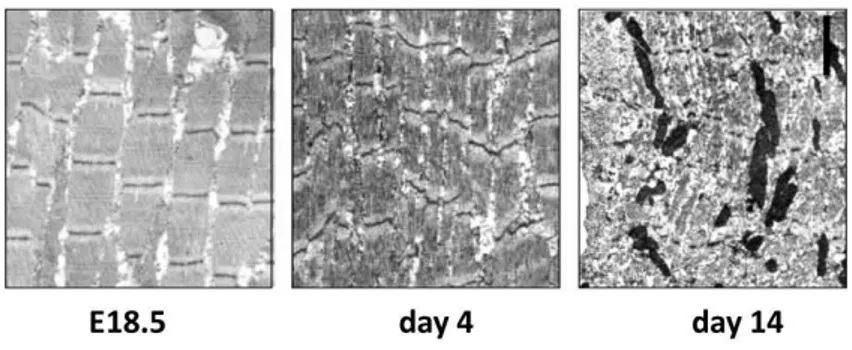
Mice with homozygous disruption of the bag3 gene developed normally but deteriorated postnatally with stunted growth evident by 1 to 2 weeks of age and death by 4 weeks (fig. 9).
Figure 9- Z-disk alterations, degenerated myofibrils, and apoptotic nuclei in muscle of bag3 -/- mice.
Muscle was analyzed by transmission electron microscopy. A: Diaphragm from bag3 -/- mice at E18.5, P4, and P14 (original magnification, _11,000). E18.5 diaphragm showed no abnormalities. By postnatal day 4, Z-disk streaming and disorganization of myofibrils were observed, but apoptosis was not present. By day 14, muscle degeneration was present, with numerous pyknotic nuclei. (Homma et al., 2006).
BAG3-deficient animals developed a fulminant myopathy characterized by non-inflammatory myofibrillar degeneration with apoptotic features (Homma et al., 2006). These data suggesting that, although BAG3 is not required for muscle development, this co-chaperone appears to be critically important for maintenance of mature skeletal muscle. Moreover, in humans, a heterozygous p.Pro209Leu mutation in BAG3 protein was recently recognised to be responsible for a severe muscular dystrophy with cardiomyopathy and severe respiratory insufficiency (Selcen et al., 2009; McCollum et al., 2009) (fig. 10).
Chapter II
BAG3 protein
- 18 -
Figure 10- (A) Scheme of the genomic structure of BAG3 and the identified mutation. (B) Alignment of
amino acid sequences of human, chimpanzee, dog, bovine, rat, mouse, chicken, and zebra fish. (Selcen et al., 2009).
In one of the three studied families carrying this mutation, an axonal neuropathy was also present. This observation is intriguing in view of the reported localization of a 40 kD form of BAG3 in synaptosomes (Bruno et al., 2008) and of the above discussed role of BAG3 in CNS.Finally a recent paper reports a role of BAG3 in the development of the haematopoietic system, showing that mice with a targeted disruption of bag3 exhibit a loss of hematopoietic stem cells and defective B-cell development, due to a microenvironmental defect, i.e. an alteration in the vascular stem cell niche (Selcen, 2010). The Authors observed a defective growth of stromal progenitor cells in colony forming unit-fibroblasts, a defect in sinusoidal endothelium, and the loss of stromal cells expressing CXCL-12 or IL-7 in the bone marrow (Selcen, 2010). The molecular mechanisms underlying those perturbations could, once identified, disclose novel prospects in the understanding of the haematopoietic process.
Last but not least, a recent evidence indicate that BAG3 is also implicated in regulating the general metabolic state of the organism, since bag3- deficient mice were reported to show significant hypoglycemia, a decrease in triglyceride and cholesterol levels and growth retardation, and died by 3 wks after birth (Kwon et al.,2010).
bag3-knockout animals have been studied in two papers (Homma et al. 2006; Youn et al. 2008). Both of them report homozygous animal death by 3–4 weeks after birth, although the observed phenotypes appear different. Indeed, Homma et al. report a fulminant myopathy with apoptotic features, while Youn et al. evidence a
Chapter II
BAG3 protein
- 19 -
metabolic deterioration. At present, the precise reasons for the differences in the phenotypes of both models are not entirely clear. The method used for gene targeting may contribute to the different phenotypes observed. Homma et al., describe a bis-deficient model developed with ES clones that had been mutagenized by retroviral insertion; Youn et al., instead, report a bis-deficient mice model developed by precise deletion of exon 4 of the bis gene with a Cre-loxP system. Although Homma et al. do not describe which part of the bis gene was disrupted by retroviral insertion, partial disruption of the bis gene may have resulted in the expression of truncated Bis protein products, and these may have retained some function. Youn et al. did not detect, in their system, any full-length or truncated Bis protein by Western blotting. Another possible explanation for the discrepancy in the reported phenotypes of bis-deficient mice may be the extent of homogeneity in the genetic background. For the generation of homozygous bis-/- mice, Youn et al. have used heterozygous mice that were backcrossed with C57BL6 more than eight generations. It is also possible that the metabolic disturbances observed in his study using biochemical and ultrastructure assays were not noticeable in the histological examinations performed by Homma et al. (Youn et al. 2008)
However, altogether these data show a fundamental role of BAG3 in regulating essential physiological events.
2.3 BAG3 role in cell adhesion and motility
It has been shown that bag3 silencing reduces adhesion and/or motility of epithelial (breast, prostate) tumour cells (Iwasaki et al., 2007; Kassis et al., 2009; Kassis et al., 2006; Fontanella et al., 2010). In MDA435 human breast cancer cells, BAG3 over-expression resulted in a decrease in migration and adhesion to matrix molecules; the decrease was reversed upon deletion of the BAG3 proline-rich (PXXP) domain, indicating that an interaction of BAG3 with a SH3 domain- containing protein was involved (Kassis et al., 2006). Expression array studies showed that indeed BAG3 regulated, in a PXXP- dependent manner, the expression of CCN (Cyr61, Connective tissue growth, NOV) 1, a matricellular signaling protein that promotes cell adhesion through integrins and heparan sulfate-containing proteoglycans (Kassis et al., 2009). In addition BAG3 seems to regulate cell adhesion through binding to guanine nucleotide exchange factor 2 (PDZGEF2). This protein induces the activation of Rap1, a regulator of cell-cell junction formation and remodeling, and increases integrin-mediated cell adhesion. The PPDY motif at the C-terminus of PDZGEF2 was shown to bind to the WW domain of BAG3 and PDZGEF2- knockdown reduced BAG3 ability to induce cell adhesion in Cos7 cells (Iwasaki et al., 2010).
Moreover, recently has been reported that BAG3, through its interaction with the cytosolic chaperonin CCT (Chaperonin Containing TCP-1), regulates actin folding. This property of BAG3 highlights its involvement in cytoskeleton organization, possibly influencing not only cell survival and migration, but also membrane trafficking and organellar dynamics.
Similarly to what we described above for apoptosis, the ability of BAG3 to regulate cell adhesion appears to rely on multiple interactions of this protein through different structural domains.
Chapter II
BAG3 protein
- 20 -
2.4 BAG3 in autophagy
The role played by BAG3 in cytoskeleton remodelling and membrane trafficking suggests the possibility that it might be involved in autophagy. With this term we refer to a set of non-specific bulk degradation processes in which cells deliver cytoplasmic substrates for lysosomal degradation (Moreau et al., 2010).
There is a chaperone-mediated autophagy (CMA), selective for cytosolic proteins that contain a pentapeptide motif: this motif is recognised by the chaperone heat shock cognate 70 (HSC70), which transfers protein substrates to lysosomes (Moreau et al., 2010). Since BAG3 is a Hsc/Hsp 70 co-chaperone, it is plausible to imagine its involvement in CMA, but we can also envisage a role in the other two types of autophagy, namely micro- and macro- autophagy. In macroautophagy, cells shape double-membrane vesicles, called autophagosomes, around a portion of cytoplasm; autophagosomes are trafficked along microtubules and fused with lysosomes, resulting in degradation of their contents. Microautophagy is a process in which lysosomes directly engulf cytoplasm (Moreau et al., 2010). Like heat shock proteins and BAG3, autophagy is upregulated under stress. Indeed, the autophagic process is important in promoting cell survival in several conditions, such as protein aggregate formation, nutrient and growth factor deprivation, ER stress and pathogen infection. Defective autophagy is associated with diverse diseases, including neurodegeneration, lysosomal storage diseases, muscular dystrophies, cancers, and Crohn's disease (Moreau et al., 2010).
BAG3 participates, along with HspB8, a member of the HspB family of molecular chaperones, in the degradation of misfolded and aggregated proteins via macroautophagy. Indeed HspB8 forms a stable complex with BAG3 in cells and the formation of this complex is essential for the HspB8- mediated degradation of the polyglutamine protein Htt43Q (HunTingTin exon 1 fragment with 43 CAG repeats), a pathogenic form of huntingtin prone to aggregation (Carra et al., 2009a; Carra et al., 2009b). HspB8 and BAG3 induce, in a Hsp70- independent manner, the phosphorylation of the alpha-subunit of the translation initiator factor eIF2; this in turn causes a translational shut-down and stimulates autophagy. The mechanism by which the BAG3/HspB8 complex induces eIF2 phosphorylation is not completely understood (Carra et al., 2009a; Carra et al., 2009b). BAG3 binding to HspB8 is mediated by two conserved IPV (Ile-Pro-Val) motifs located between the WW- and the Pro-rich- domains of the co-chaperone (fig. 11); deletion of these motifs suppresses HspB8 activity in Htt43Q degradation (Fuchs et al., 2009).
Chapter II
BAG3 protein
- 21 -
Interestingly, the p.Pro209Leu mutation responsible for dystrophy is in one of the two IPV motifs (Selcen et al., 2009; Fuchs et al., 2009). This suggests that the disease pathogenesis could involve defective autophagy. Through the same protein region, BAG3 can bind also to another chaperone HspB6. This is particularly intriguing in view of the cardioprotective property of HspB6/Hsp20 (Fan et al., 2005) and its role in myocyte contractility (Dreiza et al., 2010).
Chapter III
Analisys of BAG3 role in cardiomyocytes physiopathology
- 23 -
3.1 Introduction
The human BAG family of proteins comprises six family members (BAG1-6) that function as molecular chaperone regulators. BAG proteins, that share the BAG domain interacting with Hsp70, are involved in a number of cellular processes, including proliferation and apoptosis. Among these proteins, BAG3 is receiving increased attention due to its high levels in several disease models. It has been recently shown that, in human primary lymphoid and myeloblastic leukemias and other neoplastic cell types, BAG3 expression sustains cell survival and underlies resistance to therapy, by influencing apoptosis. BAG3 expression is stimulated during cell response to stressful conditions, such as exposure to high temperature, heavy metals, and certain drugs. It is instead constitutive in myocytes, a few other normal cell types and several tumours (leukemia and lymphoma, myeloma, pancreas and thyroid carcinomas, melanoma, osteosarcoma, etc.). Furthermore, it has been recently shown that expression of the gene encoding BAG3 is especially high in skeletal and cardiac muscle in vivo. However, relatively little is known about levels of BAG3 under normal conditions, and the physiological role of BAG3 has not previously been defined, even if it is seen that knockout of the BAG3 gene in the mouse leads to a fulminant myopathy and an early lethality.
Our group recently focused its interest on BAG3 role in myocardiocyte proliferation, survival and response to stressful stimuli. We found that BAG3 is upregulated during the differentiation of cardiomyoblasts and seems to sustain myogenin expression. Furthermore, we analyzed BAG3 expression and localization following cell exposure to oxidative stress. In particular, we found that epinephrine in vitro increases BAG3 expression in adult human cardiomyocytes. We evaluated whether BAG3 could be involved in the Tako-Tsubo cardiomyopathy (or stress cardiomyopathy) pathogenesis that is characterized by left ventricular dysfunction, with symptoms that can mimic an acute coronary syndrome. Therefore, we sequenced bag3 gene to check for polymorphisms in 29 patients and 1043 healthy donors.
We also showed for the first time that BAG3 protein is released from stressed cardiomyocytes and is found in chronic heart failure (HF) patients’ sera. Since anti-BAG3 antibodies are also present in patients’ sera, we developed an ELISA test for their specific detection.
3.2 Results and discussion
3.2.1 BAG3 role in processes of cardiomyocytes differentiation
In the program of cardiomyoblast differentiation, induction of proteins that protect cardiomyocytes from stretch and load stress- induced apoptosis has a relevant role. Indeed, cardiomyocytes are intrinsically resistant to apoptosis, due in part to high levels of endogenous caspase inhibitors (Siu et al., 2005), anti- apoptotic Bcl-2 proteins (Dorn, 2009) and the pro-survival kinase Akt (Matsui et al., 2003).
To investigate the role of anti-apoptotic BAG3 protein in cardiac myocytes, we first analyzed BAG3 levels in undifferentiated H9c2 myoblasts, an embryonic rat-derived cloned line able to undergo differentiation when cultured in medium with reduced (1%) fetal bovine serum (FBS) (Kee et al., 2007), and compared them to
Chapter III
Analisys of BAG3 role in cardiomyocytes physiopathology
- 24 -
levels detectable in differentiated H9c2 cells. H9c2 myoblasts, cultured in medium supplemented with 10% FBS, displayed a typical roundish phenotype; when cultured for 4 days in medium supplemented with 1% FBS, the cells (myocytes) appeared thin and elongated, often fused to form multinucleated syncytia (fig. 12).
Figure 12 – Left panel: H9c2 cells were seeded at a density of 5x105 cells/ 100 mm plate and cultured at 37°C in a 5% CO2 humidified atmosphere, in Dulbecco’s Modified Eagle’s Medium (DMEM) supplemented with 100 U/ml penicillin, 100 mg/ml streptomycin, and 10% FBS (high-serum, growth-promoting medium). Right panel: H9c2 cells in a subconfluence (70–80%) state were cultured in DMEM medium supplemented with 1% FBS (low-serum, differentiation-promoting medium) for 4 days.
We analyzed by Western blotting cell lysates from the two types of cultures and found that BAG3 levels were markedly higher in differentiated myocytes. In parallel it was observed an increase for myogenin, a marker of cardiomyocyte differentiation (Kee et al., 2007) (fig. 13).
Figure 13 – H9c2 myoblasts were cultured in high (10% FBS)- or low (1% FBS)- serum medium for 4 days,
Chapter III
Analisys of BAG3 role in cardiomyocytes physiopathology
- 25 -
antibody AC-1 (Enzo Biochem, New York, NY). Myogenin was also analyzed, as a marker of the differentiated state, using a monoclonal antibody purchased from BD Biosciences, San Diego, CA. An anti- lamin-A polyclonal antibody (Sigma-Aldrich, St. Louis, Missouri) was used to monitor equal protein levels.Therefore BAG3 levels appear to increase during myoblast differentiation, suggesting that its biological role is relevant for differentiated myocytes and not for immature cells. This is in agreement with the observation that bag3 deletion causes a lethal cardiopathy not in embryos, but postnatally in mice (Homma et al., 2006). We then verified whether bag3 silencing could affect the differentiation state of cardiocytes. For this purpose, we analyzed primary human cardiac myocytes, HCMa line, in which we reduced BAG3 levels by using a specific small interfering (si) RNA for 48h. bag3 siRNA- transfected cells did not display a significant degree of cell death - possibly because of the incomplete down-modulation of the protein, nor an appreciable change in morphology. On the other hand, surprisingly, we found that bag3 silencing resulted in highly reducing the levels of myogenin (fig. 14) (De Marco et al., 2011).
Figure 14 – Human primary cardiac myocytes (HCM, Sciencell Research Laboratories, San Diego, CA) at
30% confluency were transfected with a bag3- specific small interfering (si) RNA () or a control scrambled RNA. After 48 h, whole-cell extracts were obtained and analyzed by immunoblotting with the anti-BAG3 monoclonal antibody AC1, anti-myogenin antibody or, as a control, an anti-α-tubulin monoclonal antibody (Santa Cruz Biotechnology, Santa Cruz, CA).
These findings indicate an involvement of BAG3 protein in late heart development and are in keeping with the role of BAG3 in the survival and myofibrillar integrity in cardiocytes.
Chapter III
Analisys of BAG3 role in cardiomyocytes physiopathology
- 26 -
3.2.2 BAG3 induction in response to oxidative stress
Phenethyl isothiocyanate (PEITC) is an inducer of oxidative stress belonging to the family of isothiocyanates It is well known that ITCs can induce cellular oxidative stress by rapidly conjugating and thus depleting cells of glutathione (GSH). As electrophiles, ITCs readily form conjugates with thiols, including the thiols in GSH and cellular proteins; this conjugation reaction is reversible (Hall, 1999).
In order to analyse the effect oxidative stress on BAG3 protein expression, H9c2 cells were treated with PEITC at concentrations of 10, 25, 50 microM for 30 minutes, 2, 4, 8, 16 and 24 hours. After treatments, apoptosis levels were evaluated, through flow cytometry, analysis;: the highest concentrations lead to a strong induction of cell death, while that corresponding to 10 uM not is significantly harmful to the cells. For this reason, the following experiments were conducted using this concentration. After treatment with PEITC, the cells were harvested and lysed and proteins were analyzed by Western blot analysis in order to detect changes in BAG3 protein cytoplasmic levels Data analysis (fig. 15) reveals a peak of expression after 16 hours of stimulation.
Figure 15 – Subconfluent serum-starved cardiomyocytes, H9c2 line, (4.5x105 cells) were treated with PEITC 10 µM for indicated times. A: Western blot analysis of the intracellular BAG3 protein. An antibody recognizing GAPDH was used to monitor equal loading conditions. B: cell apoptosis was then analyzed by cell permeabilization and PI staining in flow cytometry.
Then, to evaluate the effect of PEITC on differentiated cardiac cells, HCMa cells were treated at the same concentrations for the same times. Even in this cell line, only the concentration corresponding to 10 microM allows to highlight an increase in the BAG3 levels in the absence of a significant cell death, this level peak, however, is reached after only 4 hours of culture with the inductor (fig. 16). Long times of exposure to PEITC are all harmful to cells.
Chapter III
Analisys of BAG3 role in cardiomyocytes physiopathology
- 27 -
Figure 16 – Subconfluent serum-starved human cardiomyocytes, HCMa line, (4.5x105 cells) were treatedwith PEITC 10 µM for indicated times. Western blot analysis of the intracellular BAG3 protein. An antibody recognizing GAPDH was used to monitor equal loading conditions.
In order to confirm this finding ,an indirect immunofluorescence was conducted on same cells using the anti-BAG3 monoclonal antibody AC-1, which recognizes the N-terminal portion of the protein. Confocal microscopy analysis allowed to highlight the increase of BAG3 expression following treatment with PEITC 10 M for 4 hours (fig. 17).
Figure 17 – HCMa cells were cultured on coverslips in six-well plates to 60-70% confluence and then treated with for PEITC 10 µM 4 h. Cells were permeabilized for 10 min with Triton X-100 (0.1% v/v in PBS). Following incubation with a normal goat serum (NGS) (5% v/v in PBS) for 1 hour at RT, coverslips were incubated with monoclonal antibody BAG3 AC1. After incubation with a 1:500 dilution of goat anti-mouse IgG DyLigth 594-conjugated antibodies (Jackson ImmunoResearch, West Grove, PA, USA) at room temperature for 45 min, coverslips were again washed for three times in 1x PBS. Samples were analyzed using a confocal laser scanning microscope (Zeiss LSM confocal microscope, Germany).
Chapter III
Analisys of BAG3 role in cardiomyocytes physiopathology
- 28 -
It was also observed a significant increase in BAG3 protein in rat heart tissue after induction of heart attack through a temporary descending coronary artery occlusion. In fig. 18 are shown the results obtained by immunohistochemistry using a BAG3-specific monoclonal antibody in the control groups and MI.
Figure 18 – Male Sprague-Dawley rats (Charles River Laboratories, Italy),were anesthetized with an intraperitoneal injection of pentobarbital (60 mg per pound) and then orally intubated. After performing an anterior thoracotomy, the heart was exteriorized and subjected to a suture of the proximal anterior descending coronary artery. Control animals were subjected to the same procedure except for artery ligation. The day after surgery, rats were selected by transthoracic echocardiography for the presence of large infarcts involving at least 35% of the left ventricle (MI group). Finally, animals were sacrificed by standard procedures and the left ventricle treated with formalin for immunohistochemistry. In the figure are shown representative data of BAG3 expression in control and MI groups.
Oxidative stress leads to the catecholamines release by the nervous system; high concentrations of circulating adrenaline could be found after ischemic episodes. These findings have led us to assess whether the induction of BAG3 protein could also occur following treatment with adrenaline. For this purpose HCM cells were incubated with adrenaline 500 M for 30'-1-2-3 and 4h. After the treatments indicated, cells were harvested and lysed; proteic lysates obtained were analyzed by Western blot with the purpose of detecting a possible variation of the BAG3 protein expression. In our experimental conditions after a 2 hours induction with adrenaline occurs a significant increase in BAG3 cytoplasmic levels (fig. 19).
Chapter III
Analisys of BAG3 role in cardiomyocytes physiopathology
- 29 -
Figure 19 – HCMa cells at 80% confluence were incubated with epinephrine 500 uM for 2 hours, then totalcell lysates were obtained and analyzed by Western blotting with the anti-BAG3 TOS2 polyclonal antibody. An antibody recognizing GAPDH was used to monitor equal loading conditions.
To determine whether the increase in this expression correlates with a transcriptional increase, has been used the technique of Real Time-PCR, which has allowed us to demonstrate that increased levels of protein expression corresponds to the increase in the levels of BAG3 mRNA. Fig. 20 shows that BAG3 mRNA is positively regulated already after 1 hour of adrenaline's stimulation, compared to untreated cells.
Figure 20 – HCMa cells were treated with epithephrine 500 uM for indicated times, then were collected and total RNA was isolated by using TriZol Reagent (invitrogen), real time qPCR was performed by using bag3 gene targeting primers (hBAG3mRNA793fw CCAGCCTCCCACGGACCTGA and hBAG3mRNA906rw CTGGTGACTGCCCAGGCTGC) and gapdh targeting primers as housekeeping
gene (GAPDHmRNA107fw GAAGGTGAAGGTCGGAGT and GAPDHmRNA333rw
GAAGATGGTGATGGGATTTC). Analysis of relative gene expression data was calculated using the 2 e
-ΔΔCt
Chapter III
Analisys of BAG3 role in cardiomyocytes physiopathology
- 30 -
3.2.3 Polymorphisms of BAG3 could have a role in the pathogenesis of Tako-Tsubo cardiomyopathy
In the cardiac muscle tissue, BAG3 is expressed at high levels, it localizes at the Z-discs and supports the survival and the contractile activity of muscle (Arimura et al., 2011). Mutations of bag3 gene have been associated with some forms of myofibrillar myopathy and dilated cardiomyopathy (Hishiya et al., 2010; Villard et al., 2011). We therefore sought the presence of BAG3 mutations or single nucleotide polymorphisms (SNPs) in patients with Tako-Tsubo Cardiomyopathy (TTC).
Study population consisted of 29 patients (27 females, 94%; age 62±13; range 35–82) with TTC diagnosed according to the Mayo Clinic diagnostic criteria, compared with 1043 healthy donors (68% females, age 92±12; range 50–110). Isolated chest pain (59%), and dyspnea (20%) were the most prevalent presenting symptoms. Emotional (71%) and physical (29%) stress were identified as trigger event in 21 patients. Slight elevation of troponin I level was documented (3.42±6.29 ng/ml; normal value: 0-0.10 ng/ml). ECG at presentation was characterized by ST-elevation in 18 patients. Apical ballooning was more prevalent than variant forms (4 patients) and detected in 25 patients. Acute complications, such as cardiogenic shock (3 patients) and acute heart failure (6 patients) were reported.
The polymorphisms of BAG3 in TTC patients compared with healthy donors are listed in table I.
Table I. BAG3 polymorphisms: allele counts and frequency among patients and healthy donors.
Chapter III
Analisys of BAG3 role in cardiomyocytes physiopathology
- 31 -
Two TCC patients were heterozygous for the SNP rs35434411, BAG3 R71Q, conversely all (1043) analyzed controls were homozygous for the major allele (p =
0.0007). Other two TTC patients (7%) were homozygous for the SNP rs3858340
(P407L) missense mutation of BAG3, versus 1% of analyzed controls (p = 0.045). Instead, differences in frequency of polymorphism rs2234962 (C151R), between patients and control cases, was not statistically significant (fig. 21).
Figure 21 – Bag 3 mRNA sequence in green, the translated region in red (protein) , motifs and domain are illustrated in grey. Below the location of polymorphisms are indicated showing both, the nucleotide position on the mRNA sequence (up) and the aminoacidic position on the protein sequence (middle) as well as the SNP code as annotated in the databank (down).
The missense mutation R71Q is very close to the two IPV (Ile-Pro-Val) motifs that are crucial for the HspB8/HspB6–Bag3 interaction (Fuchs et al., 2009). HspB8 (HSP22) and HspB6 (Hsp20) use the Bag3 IPV motifs to form hetero-oligomers that are necessary for HspB8 and HspB6 chaperone activity. These two proteins protect against tachypacing-induced CaT reduction and F-actin stress fiber formation in cardiomyocytes, via inhibition of RhoA GTPase activity (Fuchs et al., 2009; Mymrikov et al., 2011). So, it is possible that this missense mutation can interfere with the cardioprotective role of those HSPB members. In particular HspB6 is constitutively expressed in cardiac muscle and skeletal muscle and its phosphorylation has been implicated in multiple physiological and pathophysiological processes including apoptosis (Fuchs et al., 2009; Mymrikov et al., 2011).
The missense mutation P407L is inside the fourth PXXP motif of BAG3 protein. Bag3 PXXP region is a SH3 binding domain, and this region binds the latent form of PLC- (Doong et al., 2000). The binding of PLC- to BAG3 limits the availability of PLC-, and the presence of P407L BAG3 missense mutation could influence positively or negatively PLC- availability and activation.