Dipartimento Bioscienze e Territorio
Dottorato di ricerca in
Scienze agro-forestali, delle tecnologie agro-industriali e del territorio
rurale. I sistemi forestali
XXVIII ciclo
TESI S.S.D. AGR/05
Dinamiche di crescita, sensitività
climatica ed efficienza d’uso dell’acqua
in specie forestali
Tutor
Chiar.mo Prof. Roberto Tognetti
Coordinatore
Chiar.mo Prof. Marco Marchetti
Dottorando
Emanuele Conte
Riassunto
Il clima sta cambiando e questo ha conseguenze a livello mondiale. L’impatto dei cambiamenti climatici sulle foreste implica alterazioni nell’accrescimento e produttività, e in modificazioni della composizione e distribuzione delle specie. Divengono quindi fondamentali le informazioni sulla risposta del patrimonio forestale al cambiamento climatico. L’individuazione e la comprensione delle dinamiche dei sistemi forestali più resistenti e resilienti ai repentini cambiamenti climatici sono importanti per l’elaborazione di politiche forestali idonee a favorirne l’adattamento come altrettanto rilevante risulta la capacità di predire gli impatti dei cambiamenti climatici sull’evoluzione delle specie forestali.
Il lavoro è strutturato in tre capitoli in cui si sono indagate: i) le differenze in termini di crescita, efficienza d’uso dell’acqua e sensitività al clima in popolamenti puri e misti di faggio e pino silvestre; ii) le differenze in termini di produttività, efficienza d’uso dell’acqua ed interazioni nella consociazione tra farnia e ontano napoletano rispetto alla monocoltura di farnia; iii) le analisi degli impatti del cambiamento climatico su 34 siti di faggio con l’obiettivo di contribuire alla creazione di un ambiente modellistico che possa aiutare a predire gli effetti dei cambiamenti climatici su specie vegetali in vari contesti territoriali.
I risultati della prima ricerca hanno evidenziato un trend di crescita più elevato e un uso dell’acqua più efficiente nei plot puri rispetto al plot misto. Tuttavia, bisogna tener conto che le risposte degli alberi a periodi di condizioni climatiche meno favorevoli alla crescita possono essere distorte da altri fattori, che giocano un ruolo altrettanto decisivo. In particolare, nel nostro caso, gli interventi di gestione (diradamenti e/o utilizzazioni parziali) hanno interferito con gli effetti puramente climatici, ed inoltre, la magnitudo dello stress climatico non è stata tale da risultare in variazioni evidenti nell’andamento delle cronologie.
La seconda ricerca ha evidenziato i benefici della consociazione tra farnia ed ontano napoletano in termini di crescita, produttività ed efficienza d’uso dell’acqua. Le caratteristiche azotofissatrici dell’ontano si sono rivelate un vantaggio per la farnia, che ha registrato performance superiori nel misto rispetto alla monocoltura. Tuttavia le interazioni tra le due specie si sono dimostrate mutevoli e dinamiche nel tempo a causa di cambiamenti nella disponibilità delle risorse e modificazioni delle condizioni del sito (strutturali e/o ambientali).
Nella terza ricerca si è esplorata una metodologia per la stima degli impatti climatici a scala regionale con l’utilizzo di modelli per la valutazione dell’indice di siccità più rappresentativo ed implementazione di test predittivi di crescita. Lo studio, ancora in corso, consentirà, una volta testati e validati i risultati, di costituire un SIS (Sectorial Information System) con tools interattivi che guideranno l’utente nell’esplorazione degli scenari climatici futuri e degli impatti sulle foreste.
Il lavoro, oltre a un incremento conoscitivo, contiene elementi che possono contribuire al miglioramento dell’efficacia delle politiche di adattamento e mitigazione dei cambiamenti climatici, facilitando l’adozione di una gestione forestale resiliente.
Abstract
Climate change alters forest ecosystems’ growth and productivity, and modifies species composition and distribution. Information on responses of forest resources to climate change is fundamental. Detecting and understanding the dynamics of most resilient and resistant forest ecosystems to climate change is important to prepare suitable forest policies towards improving adaptation. Similarly, predicting the impacts of climate change on tree species evolution over the time is extremely important.
The present work is structured in three chapters focusing on: (i) assessing the differences in terms of growth, water use efficiency, and climate sensitivity between pure and mixed beech and scots pine stands; (ii) evaluating the differences in terms of growth, water use efficiency, and interactions between mixed pedunculate oak-alder and pedunculate oak pure stands; (iii) analyzing the impacts of climate change in 34 beech forest stands as a basis to develop a model framework able to predict the effects of climate change in different contexts.
The results from the first work proved a higher growth trend and a more efficient use of water in pure than in mixed plot. However, the responses of trees to periods of less favorable weather conditions for growth may be distorted by other factors that play an equally decisive role. In particular, in our case, forest management (namely thinning) would have interfered with the purely climatic effects, and in addition, the magnitude of the climatic stress has not been as such to result in evident variations of the tree ring chronologies.
The second work highlighted that mixing pedunculate oak with alder positively influenced the growth, productivity, and water use efficiency. The nitrogen-fixing characteristics of alder were demonstrated to be an advantage for the pedunculate oak, which registered higher performances in comparison with the monoculture. However, the interactions among the
species were found to be variable and dynamic over the time due to changes in resources’ availability, and modifications of the site conditions (structural and/or environmental).
In the third work, a methodology to estimate climate impacts at regional scale by using models to evaluate the most representative drought index and implement prediction growth tests, was explored. This methodology will be the basis to develop a Sectorial Information System (SIS), containing interactive tools that will help users to explore future climate scenarios, and impacts on forests (e.g. beech forests).
Further than an increased knowledge, the present work contributes to improve the effectiveness of mitigation and adaptation policies, through facilitating the adoption of a resilient forest management.
Indice
Riassunto ... i Abstract ... iii Indice ... v Introduzione Generale ... 1 § Introduzione ... 2§ Obiettivi e struttura della tesi ... 11
§ Bibliografia ... 13
Capitolo 1: Growth dynamics, climate sensitivity and water-use efficiency in pure vs. mixed pine and beech stands in Trentino (Italy)... 19
1.1 Abstract ... 20
1.2 Introduction ... 21
1.3 Materials and Methods ... 24
1.3.1 Study area ... 24
1.3.2 Meteorological data and drought index ... 24
1.3.3 Tree sampling and ring-width chronologies ... 25
1.3.4 Stable isotope analysis ... 26
1.3.5 WUEi ... 27
1.3.6 Data treatment and statistical analysis ... 28
1.4 Results ... 30
1.4.1 Tree-Ring, BAI, WUEi , d13C and d18O ... 30
1.4.2 Drought index, growth-climate correlations and GLMM ... 31
1.5 Discussion ... 32 1.6 Conclusion ... 36 1.7 Acknowledgments ... 37 1.8 List of Tables ... 38 1.9 List of Figures ... 40 1.10 References ... 46 Capitolo 2: Effects of associating Quercus robur L. and Alnus cordata Loisel. on plantation productivity and water use efficiency ... 54
2.1 Abstract ... 55
2.2 Introduction ... 56
2.3 Materials and methods ... 59
2.3.2 Plantation design ... 59
2.3.3 Sampling approach ... 60
2.3.4 Soil samples ... 61
2.3.5 Stable C and O isotope analysis ... 61
2.3.6 Calculation of WUEi from C isotope ratios ... 62
2.3.7 Statistical analysis ... 63
2.4 Results ... 64
2.4.1 Tree-ring parameter trends ... 64
2.4.2 Climate influences ... 65
2.5 Discussion ... 66
2.5.1 N-fixing species affects the balance between A and gs ... 66
2.5.2 N interactions between species changed through time ... 69
2.6 Conclusion ... 71 2.7 Acknowledgements ... 72 2.8 List of Tables ... 73 2.9 List of Figures ... 74 2.10 References ... 77 Capitolo 3: Il Progetto AgriCLASS ... 87 3.1 Introduzione ... 87 3.2 AgriCLASS ... 91
3.2.1 Purpose and Scope ... 91
3.2.2 Sampling sites ... 92
3.2.3 Available data ... 92
3.2.4 Models ... 98
3.2.5 Conclusion and future work ... 103
3.2.6 References ... 105
Conclusioni ... 106
Elenco Pubblicazioni ... 110
Appendice ... 112
§ Effects of associating Quercus robur L. and Alnus cordata Loisel. on plantation productivity and water use efficiency. Paper pubblicato su Forest Ecology and Management
§ From one- to two-phase sampling to reduce costs of remote sensing-based estimation of land-cover and land-use proportions and their changes. Paper pubblicato su Remote Sensing of Environment
§ Introduzione
Il clima sta cambiando e questo ha conseguenze a livello mondiale. Eventi naturali e attività antropiche hanno contribuito ad incrementare le temperatura media globale, causata essenzialmente dall'aumento di emissioni “greenhouse gases” come l'anidride carbonica (CO2). L’atmosfera e gli oceani si sono riscaldati, il livello dei mari continua ad aumentare,
i ghiacciai si ritirano e l’intensità degli eventi estremi diventa sempre più elevata. Secondo il quinto rapporto IPCC, il trentennio 1983-2012 è stato probabilmente il più caldo in assoluto degli ultimi 1400 anni. Come certificato da NASA e Agenzia federale Usa per la meteorologia (NOAA), il 2016 è stato l’anno più caldo dal 1880. Un aumento della temperatura globale sulla terraferma e sulla superficie oceanica di 0.94° rispetto alla media del ventesimo secolo. Dal 1880 ad oggi ben nove dei dieci anni più caldi sono stati registrati a partire dal 2000. A fine XXI secolo, secondo l’IPCC, l’aumento della temperatura superficiale globale sarà di 1.5 °C, rispetto al periodo 1850-1900, in tutti gli scenari RCP (Representative Concentration Pathways), eccetto RCP2.6. Situazione ancor più preoccupante per gli scenari RCP6.0 (emissioni di gas serra stabili) e RCP8.5 (emissioni di gas serra molto alte) che prevedono un aumento di oltre 2 °C. Il contrasto e le differenze nelle precipitazioni tra le regioni umide e secche, e tra le stagioni umide e secche sono destinate ad aumentare (IPCC, 2013).
Inevitabilmente, in questo contesto di global climate change e global warming, le foreste assumono un ruolo sempre più rilevante nella mitigazione del clima come bacini di assorbimento del carbonio, e per le problematiche legate all’adattamento degli ecosistemi terrestri. Grazie alla loro capacità di assorbire e stoccare il carbonio, contribuiscono a ridurre la concentrazione di CO2 presente nell’aria, fungendo da serbatoi di anidride carbonica
climatici in atto mette in serio pericolo la funzionalità e la sopravvivenza stessa degli ecosistemi forestali (es., Borghetti et al., 2012; Rustad et al., 2012). Comprendere come il cambiamento climatico si riflette sul territorio e in che modo influenza ed influenzerà la distribuzione delle specie forestali e le interazioni tra esse, è la sfida che sta impegnando i ricercatori di tutto il mondo.
L’impatto dei cambiamenti globali (clima e uso del suolo) influenza il paesaggio e gli ecosistemi, in particolare quelle delle aree marginali (montane e interne, per il nostro Paese), ma anche le dinamiche a livello socio-economico (spopolamento, invecchiamento della popolazione e abbandono delle terre). I processi di abbandono hanno innescato a loro volta processi di successione ecologica, che hanno portato dapprima all’invasione da parte di arbusti, per poi lasciare spazio alla colonizzazione da parte delle superfici boscate su circa 470 mila ettari in Italia. Si tratta di superfici importanti, e le aree agricole perse nelle aree interne corrispondono alla metà del totale perso a livello nazionale, l’aumento delle superfici boscate corrisponde a più del 70% del totale aumento del bosco in tutta Italia. Queste nuove aree forestali richiedono linee guida per gestire i boschi di neo formazione in modo sostenibile e per sfruttare al meglio le potenzialità che il bosco porta in termini di servizi ecosistemici, in un contesto di produzione attenta ai temi della mitigazione e adattamento.
Un importante strumento per studiare gli effetti del cambiamento climatico sugli ecosistemi forestali, e sulle risposte degli alberi alle variazioni ambientali, è rappresentato dalla dendrocronologia. La dendrocronologia è la scienza che studia l'accrescimento delle piante arboree nel tempo, le modalità con cui si sviluppa e i fattori esterni che lo influenzano (Schweingruber, 1988). Si inizia a parlare di dendrocronologia come scienza già dai primi del 1900 grazie all’astronomo Andrew E. Douglass (in Arizona), ma solamente nel 1974 assume i caratteri di disciplina autonoma (Barefoot e Coll., 1974). Più correttamente quando la dendrocronologia viene utilizzata per individuare i parametri climatici che influenzano
l’accrescimento delle piante, si parla di dendroclimatologia. Gli alberi, rispondono ai cambiamenti ambientali con variazioni che si riscontrano nel loro accrescimento (Trenard, 1982). La dendrocronologia risulta quindi molto utile per studiare le risposte degli alberi dopo fattori di disturbo consentendo di definirne l’impatto e collocarli temporalmente.
Gli anelli legnosi costituiscono una vera e propria “memoria naturale” e con opportune tecniche si possono ottenere informazioni sempre più dettagliate. Lo studio degli isotopi stabili del carbonio (13C e 12C), ad esempio, consente di valutare le risposte fisiologiche delle piante a stress ambientali (McCarroll and Loader, 2004). Gli isotopi stabili sono presenti in natura secondo proporzioni specifiche e la loro concentrazione nei composti chimici è soggetta a variazioni (frazionamenti isotopici) in relazione ai processi chimico-fisici in cui sono coinvolti. I rapporti isotopici (rapporto tra le concentrazioni dell’isotopo raro e dell’isotopo più abbondante) vengono misurati con lo spettrometro di massa e ogni rapporto viene confrontato con quello di uno standard di riferimento (standard int. per il carbonio: VPDB – Vienna Pee Dee Belemnite) mentre le differenze dei rapporti vengono indicate con la notazione delta δ (in unità per mille ‰):
dove:
Rc = rapporto isotopico del campione,
Rstd= rapporto isotopico dello standard di riferimento.
A seguito di un frazionamento, si ha una diminuzione oppure un arricchimento dell’isotopo più pesante. Le piante contengono meno 13C rispetto a quello della CO2 atmosferica, perché
sia i processi enzimatici che fisici discriminano l’isotopo più pesante, in favore di quello più
1000
)
1
(
1000
)
(%
0=
-
´
=
-
´
std c std std cR
R
R
R
R
d
leggero. Questo processo di discriminazione isotopica (Δ) è fortemente influenzato da variazioni della capacità fotosintetica e della conduttanza stomatica nonché dalle variabili climatiche che influenzano il rapporto tra concentrazione intercellulare (ci) e atmosferica (ca)
di CO2 (Farquhar et al., 1982; Switsur and Waterhouse, 1998). Uno stress dovuto ad un
periodo di siccità o altre variazioni ambientali possono indurre la chiusura stomatica e causare una diminuzione del rapporto ci/ca, che si associa ad un incremento del δ13C della
pianta. Infatti, in condizioni di necessità (“fame di carbonio”) la pianta tende a far entrare maggiori quantità di 13C, in proporzione. Ne deriva che la composizione isotopica degli anelli di accrescimento risulta particolarmente adatta per indagare cambiamenti nell’efficienza d’uso dell’acqua (Perez-Harguindeguy et al., 2013); quantità di CO2
assimilata in rapporto alla quantità di H2O consumata. Discriminazione isotopica del
carbonio (Δ) ed efficienza d'uso dell'acqua (WUE) sono, infatti, legate da una relazione inversa che può essere utilizzata per studiare le interazioni pianta-ambiente e valutare la resistenza delle piante a stress idrici (Brugnoli and Farquhar, 2000; Duquesnay et al., 1998). La WUE può essere calcolata seguendo diverse formule. Frequentemente viene stimata come rapporto tra assimilazione e traspirazione (instantaneous water use efficiency) oppure come rapporto tra assimilazione e conduttanza stomatica (intrinsic water use efficiency) (Ehleringer et al., 1993). E può essere misurata su matrici fogliari, per investigare effetti dell’ambiente sugli scambi gassosi nel breve periodo (durante la stagione vegetativa), o su matrici legnose, per misurare retroattivamente tali effetti.
Tuttavia, un’analisi completa non può prescindere dallo studio della composizione isotopica dell’ossigeno (δ18O), altrettanto importante in un contesto di valutazione di resistenza a
stress ed effetti dei cambiamenti climatici. δ18O, non è fortemente influenzato dal tasso fotosintetico (come invece avviene per δ13C), che quindi può dipendere sia dall’apertura stomatica sia dall’efficienza biochimica, ma è funzione dalla composizione isotopica δ18O
della sorgente di acqua del suolo e dall’arricchimento traspirativo dell’acqua fogliare (>δ18O
rispetto a quello del suolo) (Scheidegger et al., 2000). Lo studio di δ18O consente di valutare il grado di apertura stomatica e le risposte delle piante alle variazioni climatiche che influenzano i consumi idrici. L’analisi integrata di δ13C e δ18O permette quindi di valutare separatamente gli effetti della variazione della conduttanza stomatica da quelli della capacità fotosintetica (Barnard et al., 2012; Tognetti et al., 2014).
Analisi di questo tipo sono molto utili anche in ricerche comparative, ad esempio per confrontare la capacità, resistenza e adattamento della stessa specie in contesti diversi (naturali o artificiali), e per confrontare le capacità di adattamento di specie diverse. In particolar modo, nell’ultimo decennio si sono moltiplicati gli studi sui boschi misti in termini di crescita, sensitività e resilienza al clima e produttività rispetto ai boschi puri. Alcuni ricercatori hanno evidenziato la capacità dei boschi misti di essere più produttivi rispetto ai boschi puri (Brassard et al., 2013; Forrester and Albrecht, 2014; Toïgo et al., 2015), di fornire livelli più alti di servizi ecosistemi (Gamfeldt et al., 2013; Loreau et al., 2001), ed essere più resistenti ad eventi estremi (Pretzsch et al., 2013). Altri studi hanno, invece, dimostrato che non sempre i boschi misti sono più produttivi e stabili (Epron et al., 2013; Grossiord et al., 2014; Merlin et al., 2015). Questi risultati discordanti inducono ad approfondire le relazioni intercorrenti tra le specie forestali all’interno di boschi misti, soprattutto in ecosistemi particolarmente fragili come quelli delle montagne dei paesi che si affacciano sul Mediterraneo. Comprenderne le dinamiche è molto importante per poter definire misure di gestione proattive volte ad enfatizzare il processo di adattamento sostenibile e mitigazione del climate change (Condés and del Río, 2015).
In questo contesto, gli eventi climatici estremi (siccità, alluvioni, eruzioni vulcaniche ecc.) rapprensentano degli snodi cruciali che evidenziano le reazioni dei sistemi forestali e consentono lo studio delle relazioni clima-accrescimenti. In particolare, la NOAA (National
Oceanic and Atmospheric Administration), ha mostrato che la regione del Mediterraneo è
affetta da una crescente siccitá. Secondo la NOAA, tutta la regione sta soffrendo dagli anni '70 di un regime secco che si manifesta con minori precipitazioni durante la stagione invernale, proprio quando le riserve idriche si ricaricano. Nell'area si sono verificati 10 degli ultimi 12 inverni piú secchi del pianeta. Il monitoraggio di questi fenomeni è importante per confermare la "diagnosi" che vede nell'area del Mediterraneo una regione particolarmente critica per le conseguenze che potrebbe avere il cambiamento climatico sugli ecosistemi forestali e per i servizi da loro forniti. Un indice di siccità particolarmente efficace è lo SPEI (Standardized Precipitation Evapotranspiration Index Vicente-Serrano et al., 2010). Nasce come estensione del più diffuso SPI (Standardized Precipitation Index; Mckee et al., 1993) e considera, oltre alle precipitazioni mensili, anche l´evapotraspirazione potenziale di riferimento (ET0). I principali vantaggi rispetto agli altri indicatori consistono nella semplicità di calcolo e nel carattere multiscalare che consente di individuare i periodi siccitosi e la loro intensità a diverse scale temporali. Si rivela proficuo, come vedremo in seguito, anche come fattore per stime previsionali di crescita.
L’integrazione fra analisi dendrocronologiche, isotopiche (δ13C, δ18O, e WUE) e indici
climatici, permette di estendere le riflessioni sulle risposte ai cambiamenti del clima, dalle
problematiche di adattamento degli ecosistemi terrestri a quelle della mitigazione tramite piantagioni artificiali. In questo caso, la maggior parte delle ricerche sono rivolte ad aumentare la produttività, la qualità dell’habitat e la fertilità del suolo. Le consociazioni tra specie azotofissatrici e non azotofissatrici incrementano la produttività e le performance ecofisiologiche rispetto alle monocolture (es., Clark et al., 2008; Corazzesi et al., 2010;
Nebout, 2008; Tani et al., 2007). La stima di WUE e in generale l’analisi isotopica può essere
determinante per comprendere le dinamiche d’uso delle risorse (azoto e acqua, in primis) e le ripercussioni sull’efficienza d’uso dell’acqua (Altieri et al., 2015; Battipaglia et al., 2016;
Dawson and Ehleringer, 1993). Inoltre, esplorare come variano questi processi nel corso del tempo, consente di mettere in atto azioni volte a massimizzare e velocizzare la produttività delle piantagioni, mitigando l’effetto dei cambiamenti climatici con risvolti socio-economico importanti.
Il passato, come abbiamo visto, è depositario di informazioni fondamentali per indagare gli aspetti che governano gli ecosistemi forestali. Ma l’obiettivo e la sfida che sta coinvolgendo i ricercatori di tutto il mondo, è riuscire a predire l’impatto dei cambiamenti climatici sulle specie forestali. Grazie ai moderni sistemi di calcolo e di modellistica, e alla rete di organizzazioni di meteorologia a scala mondiale, sono disponibili scenari climatici futuri basati su sofisticati algoritmi di Numerical Weather Prediction (NWP) e software Weather
Generator (http://www.rothamsted.ac.uk/mas-models/larswg). Ciò consente di simulare il
clima per aree specifiche e a scala globale, in modo da prevedere le risposte degli ecosistemi terrestri (adattamento) e le capacità di compensare le emissioni di gas serra (mitigazione), e di produrre occupazione.
Le tecniche dendrocronologiche abbinate a questi scenari, opportunamente modellizzati, possono fornire previsioni attendibili sulla crescita ed evoluzione delle specie forestali. Come precedentemente accennato, per veicolare e sintetizzare le relazioni che intercorrono tra le componenti (ambientali ed endogene) in un modello previsionale, in ambiente Mediterraneo, può risultare molto utile l’uso di un indice sintetico di siccità. Negli eventi estremi, infatti, le specie forestali tendono ad enfatizzare le reazioni, consentendo una migliore comprensione delle dinamiche clima-crescita. Tuttavia, l’incertezza che ancora governa tali interazioni e la generazione degli scenari climatici, rende difficile realizzare modelli totalmente affidabili e generalizzati.
Gli argomenti e le problematiche finora introdotti sono stati affrontati durante il percorso di dottorato, cercando di apportare ulteriore conoscenza su aspetti ancora poco esplorati. Le specie forestali che hanno caratterizzato la ricerca sono state: Fagus sylvatica L., Pinus
sylvestris L., Quercus robur L. e Alnus cordata Loisel. Tali specie sono importanti
ecologicamente e economicamente non solo a scala nazionale ma anche per l’intero continente, e localmente nel caso dell’ontano.
Il faggio (Fagus sylvatica L.) è una delle più importanti specie forestali europee, sia per l’estensione del suo areale, sia per la qualità del legname. È una specie tipicamente mesofila, amante cioè di climi né troppo caldi e secchi né troppo freddi e umidi; predilige esposizioni luminose purché la luce non sia eccessiva. Viene considerato un buon indicatore ambientale. Si tratta di una latifoglia maestosa, alta fino a 40 metri e che può vivere fino a 500 anni (Piovesan et al., 2005), raggiungendo diametri superiori al metro. L’areale è molto ampio e copre l'Europa occidentale, dalla Scandinavia meridionale fino alla Sicilia meridionale. In Italia domina le foreste della fascia montana dalle Alpi alla Sicilia. La corteccia è liscia e sottile di color grigio-cenere con striature orizzontali spesso colonizzata da licheni. Le foglie sono ellittiche, arrotondate alla base e appuntite all’apice, di colore verde superiormente, più biancastre nella pagina inferiore. Il frutto, chiamato faggiola, è di colore rossastro incluso in una cupula legnosa.
Il pino silvestre (Pinus sylvestris L.)è una specie sempreverde della famiglia delle Pinaceae. Questa conifera può essere alta fino a 40 metri; sopporta bene i periodi siccitosi ed è resistente al freddo; riesce a vivere anche su terreni difficili come i ghiaioni e gli accumuli detritici. L'areale di questo pino è il più vasto della specie e si spinge dall'Europa sud-occidentale fino all'Asia-orientale, al nord arriva in Norvegia, poi tutta la Siberia fino alla
Manciuria rimanendo tangente il circolo polare artico. In Italia si trova, non uniformemente distribuito, dall'Appennino ligure-emiliano, al bordo meridionale delle Alpi e Prealpi. Ha fusto cilindrico e può raggiungere il metro di diametro. La corteccia, nella parte inferiore, è fessurata in placche irregolari con profonde scanalature longitudinali, mentre nella parte alta del fusto tende a sfaldarsi in scaglie lamellari donando il caratteristico colore arancione-ocraceo. Le sue foglie sono aghiformi; il frutto, la pigna, è di consistenza legnosa e di colore grigio-marrone.
La farnia (Quercus robur L.) è una specie a foglie decidue appartenente alla famiglia delle Fagacee. Essa è caratterizzata da grande longevità, crescita iniziale lenta, dimensioni imponenti e dall’eccellente qualità del legno. Può raggiungere i 40 metri di altezza, eccezionalmente anche 50. La Farnia è una specie eliofila, mesofila, adatta a terreni ricchi, profondi e umidi. Esigente in luce, la farnia tollera per pochi anni l'ombreggiamento. L’areale è il più vasto di tutte le querce europee. Si estende fino alla Scandinavia, alle isole britanniche, all'Anatolia e al Caucaso. In Italia, è presente su tutto il territorio eccetto Sardegna e Puglia. E’ una specie dal portamento maestoso, con una chioma molto ampia e di forma globosa ed irregolare. Il fusto è diritto e robusto e i rami con il passare del tempo divengono più massicci e contorti. Le foglie, lunghe in media 10 cm, sono caduche, hanno picciolo molto breve e una forma obovato-oblunga con margini lobati. Assumono un colore verde medio nella pagina superiore mentre sono più chiare in quella inferiore. I frutti, chiamati ghiande, hanno una forma ovale allungata, fino ai 4 cm e sono ricoperti (per circa ¼) da una cupula ruvida e squamata.
L’ontano napoletano (Alnus cordata Loisel) è una specie endemica del Sud Italia, appartenente alla famiglia delle Betulaceae. Il suo areale originale è limitato all’Appennino meridionale e alla Corsica ma è attualmente diffuso in quasi tutte le regioni italiane. Si tratta di una specie molto importante poiché gli apparati radicali ospitano batteri azotofissatori simbionti, per cui la pianta fertilizza naturalmente il suolo. A differenza degli altri ontani sopporta bene la carenza idrica e la sua crescita è favorita in terreni acidi. Ha un’altezza media di 15m con chioma piramidale, fusto slanciato e rami glabri; in età giovane la corteccia è liscia grigio-verde e coperta da una sostanza biancastra chiamata pruina. Le foglie caduche sono semplici e alterne, da obovate a sub-orbicolari, con margine dentato e picciolo di 1-2 cm.Il colore è verde lucido sulla pagina superiore e verde più chiaro nella pagina inferiore. Le infruttescenze hanno un tipico aspetto legnoso e non si disintegrano a maturità.
§ Obiettivi e struttura della tesi
L’obiettivo principale dello studio è stato quello di indagare le dinamiche di crescita, la sensitività climatica e l’efficienza d’uso dell’acqua in popolamenti puri e misti (naturali e artificiali) delle specie forestali menzionate sopra. Inoltre, fornire, in un contesto di climate
change e global warming, gli strumenti per modellizzare correttamente le relazioni
clima-crescita al fine di predire gli impatti del cambiamento climatico sugli ecosistemi forestali a scala locale, nazionale e continentale. Lo studio vuole testare l’ipotesi che i boschi misti e le piantagioni plurispecifiche consentano un miglior sfruttamento delle risorse (acqua e nutrienti, oltre che luce ecc.) e che, quindi, siano più resilienti come risposta a disturbi ambientali, quali episodi siccitosi ricorrenti, in aumento come conseguenza dei cambiamenti climatici, e abbiano maggiori capacità di mitigazione.
Nel primo lavoro è stato verificato: i) se vi fossero differenze in termini di WUEi (intrinsic
water use efficiency) e risposta a stress climatici tra popolamenti puri e misti di faggio e pino
silvestre (sotto ipotesi: >WUEi in popolamenti misti); ii) se l’interazione tra le due specie
consentisse un aumento di produttività; iii) se vi fossero differenze in termini di sensitività al clima ipotizzando il faggio come specie più sensitiva rispetto al pino. Analisi dendrocronologica, analisi isotopica eseguita sugli anelli di accrescimento, e indice di siccità (SPEI) sono stati la base metodologica dello studio.
Anche nel secondo lavoro sono state esaminate le differenze tra ambiente monospecifico e plurispecifico, non più in boschi bensì in piantagioni. Si è valutata se la consociazione tra farnia (non azotofissatrice) e ontano napoletano (azotofissatrice) producesse benefici in termini di produttività e WUEi. Inoltre, le interazioni tra le due specie sono state monitorate
nel tempo allo scopo di comprendere se, a causa di cambiamenti nella disponibilità di risorse, stress climatici o composizione del sito, l’ipotizzato vantaggio della consociazione si tramutasse in interazione competitiva.
Nel terzo capitolo sono state concentrate le conoscenze acquisite sulle relazioni che intercorrono tra dinamiche di crescita e clima allo scopo di implementare un sistema in grado di sintetizzare tali interazioni in parametri utili a fini predittivi. Su 34 punti di rilievo in Molise, caratterizzati dalla dominanza di faggio, è stato effettuato uno studio dendrocronologico multi-sito volto ad evidenziare gli impatti climatici sugli accrescimenti. Lo SPEI è stato correlato con le curve di accrescimento dei diversi siti mediante GLMM (Generalized Linear Mixed Models, Zuur et al., 2013). Per individuare quale valore di SPEI (mese e scala) rappresentasse il miglior indice di siccità, sono stati implementati tre modelli alternativi. Il GLMM con le migliori performance è stato quindi utilizzato per effettuare un test predittivo in base a diversi scenari climatici futuri. Una volta testati e validati i risultati, l’obiettivo finale è quello di contribuire alla creazione di un ambiente modellistico che possa
aiutare a predire gli effetti dei cambiamenti climatici su specie vegetali (nel caso in questione il faggio) in vari contesti territoriali (es., Molise), costituendo un vero e proprio SIS (Sectorial Information System) accessibile al pubblico e ai portatori d’interesse istituzionali (es.: servizi regionali alla programmazione territoriale e forestale).
§ Bibliografia
Altieri, S., Mereu, S., Cherubini, P., Castaldi, S., Sirignano, C., Lubritto, C., Battipaglia, G., 2015. Tree-ring carbon and oxygen isotopes indicate different water use strategies in three Mediterranean shrubs at Capo Caccia (Sardinia, Italy). Trees - Struct. Funct. 29, 1593–1603.
Barefoot, A.C., Woodhouse, L.B., Hafley, W.I., Wilson, E.H., 1974. Developing a dendrochronology for Winchester, England. Journal of the Institute of Wood Science, VI (5), 34-40.
Barnard, H.R., Brooks, J.R., Bond, B.J., 2012. Applying the dual-isotope conceptual model to interpret physiological trends under uncontrolled conditions. Tree Physiol. 32, 1183–1198.
Battipaglia, G., Savi, T., Ascoli, D., Castagneri, D., Esposito, A., Mayr, S., Nardini, A., 2016. Effects of prescribed burning on ecophysiological, anatomical and stem hydraulic properties in Pinus pinea L. Tree Physiol. 36, 1019–1031.
Borghetti, M., La Mantia, T., Menozzi, P., Piotti, A., 2012a. The likely impact of climate change on the biodiversity of Italian forests. For. - Riv. di Selvic. ed Ecol. For. 9, 245– 250.
Borghetti, M., La Mantia, T., Menozzi, P., Piotti, A., 2012b. Probabili impatti del cambiamento climatico sulla biodiversità delle foreste italiane. iForest - Biogeosciences For. 9, 245–250.
Brassard, B.W., Chen, H.Y.H., Cavard, X., Laganière, J., Reich, P.B., Bergeron, Y., Paré, D., Yuan, Z., 2013. Tree species diversity increases fine root productivity through increased soil volume filling. J. Ecol. 101, 210–219.
Brugnoli, E., Farquhar, G.D., 2000. Photosynthetic Fractionation of Carbon Isotopes. Photosynth. Physiol. Metab. 399–434.
Clark, J., Hemery, G., Savill, P., 2008. Early growth and form of common walnut (Juglans
regia L.) in mixture with tree and shrub nurse species in southern England. Forestry
81, 631–644.
Condés, S., del Río, M., 2015. Climate modifies tree interactions in terms of basal area growth and mortality in monospecific and mixed Fagus sylvatica and Pinus sylvestris forests. Eur. J. For. Res. 134, 1095–1108.
Corazzesi, A., Tani, A., Pelleri, F., 2010. Effetto della consociazione e del diradamento in un impianto di arboricoltura da legno con latifoglie di pregio dopo oltre 20 anni dall'impianto positivo. Ann. Silvic. Res. 36, 37–48.
Dawson, T.E., Ehleringer, J.R., 1993. Isotopic enrichment of water in the “woody” tissues of plants: Implications for plant water source, water uptake, and other studies which use stable isotopic composition of cellulose. Geochim. Cosmochim. Acta. 57, 3487-3492.
Duquesnay, A., Bréda, N., Stievenard, M., Dupouey, J.L., 1998. Changes of tree-ring δ13C
and water-use efficiency of beech (Fagus sylvatica L.) in north-eastern France during the past century. Plant, Cell Environ. 21, 565–572.
Ehleringer, J.R., Hall, A. E., Farquhar, G.D., 1993. Stable Isotopes and Plant Carbon/Water Relations. Academic Press, San Diego, 555 pp.
Epron, D., Nouvellon, Y., Mareschal, L., Moreira, R.M. e, Koutika, L.S., Geneste, B., Delgado-Rojas, J.S., Laclau, J.P., Sola, G., Gonçalves, J.L. de M., Bouillet, J.P., 2013. Partitioning of net primary production in Eucalyptus and Acacia stands and in mixed-species plantations: Two case-studies in contrasting tropical environments. For. Ecol. Manage. 301, 102–111.
Farquhar, G., O’Leary, M., Berry, J., 1982. On the Relationship Between Carbon Isotope Discrimination and the Intercellular Carbon Dioxide Concentration in Leaves. Aust. J. Plant Physiol. 9, 121.
Fischlin, A., Buchter, B., Matile, L., Hofer, P., T.R., 2006. Serbatoi di carbonio nell’economia forestale: conteggio dell’assorbimento e delle fonti di emissione nel contesto del Protocollo di Kyoto. Stud. sull’ambiente n. 0602 2, 1–17.
Forrester, D.I., Albrecht, A.T., 2014. Light absorption and light-use efficiency in mixtures of Abies alba and Picea abies along a productivity gradient. For. Ecol. Manage. 328, 94–102.
Gamfeldt, L., Snall, T., Bagchi, R., Jonsson, M., Gustafsson, L., Kjellander, P., Ruiz-Jaen, M.C., Froberg, M., Stendahl, J., Philipson, C.D., Mikusinski, G., Andersson, E., Westerlund, B., Andren, H., Moberg, F., Moen, J., Bengtsson, J., 2013. Higher levels of multiple ecosystem services are found in forests with more tree species. Nat. Commun. 4, 1340.
Grossiord, C., Granier, A., Ratcliffe, S., Bouriaud, O., Bruelheide, H., Chećko, E., Forrester, D.I., Dawud, S.M., Finér, L., Pollastrini, M., Scherer-Lorenzen, M., Valladares, F., Bonal, D., Gessler, A., 2014. Tree diversity does not always improve resistance of forest ecosystems to drought. Proc. Natl. Acad. Sci. 111, 14812–14815.
Loreau, M.; Naeem, S.; Inchausti, P.; Bengtsson, J.; Hector, A.; Hooper, D.U.; Huston, M.A.; Raffaelli, D.; Tilman, D.; Wardle, D.A.; Grime, J.P.; Schmid, B., 2001. Biodiversity and Ecosystem Functioning: Current Knowledge and Future Challenges. Science. 294, 804–808.
McCarroll, D., Loader, N.J., 2004. Stable isotopes in tree rings. Quat. Sci. Rev. 23, 771– 801.
Mckee, T.B., Doesken, N.J., Kleist, J., 1993. The relationship of drought frequency and duration to time scales. AMS 8th Conf. Appl. Climatol. 179–184.
Merlin, M., Perot, T., Perret, S., Korboulewsky, N., Vallet, P., 2015. Effects of stand composition and tree size on resistance and resilience to drought in sessile oak and Scots pine. For. Ecol. Manage. 339, 22–33.
Nebout, J., 2008. Plantation et éducation des chênes dans les recrû naturel. Forêt Entrep. 178, 17-20
Pérez-Harguindeguy, N., Díaz, S., Garnier, E., Lavorel, S., Poorter, H., Jaureguiberry, P., Bret-Harte, M. S., ... & Urcelay, C., 2013. New handbook for standardised measurement of plant functional traits worldwide. Aust. J. Bot. 61, 167–234.
Piovesan, G., Di Filippo, A., Alessandrini, A., Biondi, F., Schirone, B., 2005. Structure, dynamics and dendroecology of an old-growth Fagus forest in the Apennines. J. Veg. Sci. 16, 13–28.
Pretzsch, H., Schütze, G., Uhl, E., 2013. Resistance of European tree species to drought stress in mixed versus pure forests: Evidence of stress release by inter-specific facilitation. Plant Biol. 15, 483–495.
Rustad, L., Campbell, J., Dukes, J.S., Huntington, T., Lambert, K.F., Mohan, J., Rodenhouse, N., 2012. Changing Climate, Changing Forests: The Impacts of Climate Change on Forests of the Northeastern United States and Eastern Canada. Gen. Tech. Rep. NRS-99. Newtown Square, PA: U.S. Department of Agriculture, Forest Service, Northern Research Station. 48 p.
Scheidegger, Y., Saurer, M., Bahn, M., Siegwolf, R., 2000. Linking Stable Oxygen and Carbon Isotopes with Stomatal Conductance and Photosynthetic Capacity : A Conceptual Model. Oecologia 125, 350–357.
Schweingruber, F.H., 1988. Tree Rings Basics and Applications of Dendrochronology, Kluwer Academic Publishers. Journal of Tropical Ecology, 5(03), 352-352.
Stocker, T.F., D. Qin, G.-K. Plattner, M. Tignor, S.K. Allen, J. Boschung, A. Nauels, Y. Xia, V.B. & P.M.M. (eds.), 2013. IPCC, 2013: Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge Univ. Press. Cambridge, United Kingdom New York, NY, USA, 1535 pp.
Switsur, R., Waterhouse, J., 1998. Stable isotopes in tree ring cellulose. Stable Isot. 303– 321.
Tani, A., Maltoni, A., Mariotti, B., 2007. Noce da legno e specie azotofissatrici. Sherwood-Foreste ed Alberi Oggi 15–17.
Tognetti, R., Lombardi, F., Lasserre, B., Cherubini, P., Marchetti, M., 2014. Tree-ring stable isotopes reveal twentieth-century increases in water-use efficiency of Fagus sylvatica and Nothofagus spp. in Italian and Chilean Mountains. PLoS One 9 (11).
Toïgo, M., Vallet, P., Perot, T., Bontemps, J.D., Piedallu, C., Courbaud, B., 2015. Overyielding in mixed forests decreases with site productivity. J. Ecol. 103, 502–512. Trenard, Y., 1982. Making wood speak: an introduction to dendrochronology. Forest
Products Abstracts V, 267-297.
Vicente-Serrano, S.M., Beguería, S., López-Moreno, J.I., 2010. A multiscalar drought index sensitive to global warming: The standardized precipitation evapotranspiration index. J. Clim. 23, 1696–1718.
Zuur, A.F., Hilbe, J.M., Ieno, E.N., 2013. A beginner’s Guide to GLM and GLMM with R: a frequentist and Bayesian perspective to ecologists. Highland Statistics Ltd., Newburgh.
Capitolo 1.
Growth dynamics, climate sensitivity and water-use
efficiency in pure vs. mixed pine and beech stands in
Trentino (Italy)
EMANUELE CONTE1, FABIO LOMBARDI4 GIOVANNA BATTIPAGLIA3, CATERINA PALOMBO1, SIMONA ALTIERI3, NICOLA LA PORTA2,5, MARCO MARCHETTI1, ROBERTO TOGNETTI1,2
1Dipartimento di Bioscienze e Territorio, Università degli Studi del Molise, Contrada Fonte Lappone, I-86090 Pesche (IS), Italy
2The EFI Project Centre on Mountain Forests (MOUNTFOR), 38010, San Michele a/Adige (TN), Italy
3Department of Environmental, Biological and pharmaceutical sciences and Technologies, Second University of Naples, via Vivaldi 43, I-81100 Caserta, Italy
4Dipartimento di Agraria, Università Mediterranea di Reggio Calabria, Località Feo di Vito, I-89122 Reggio Calabria, Italy
1.1 Abstract
Understanding to what extent the species mixture modifies the tree growth under drought-derived stress conditions, in comparison with monospecific stands, is demonstrated to be important to support specific adaptation and mitigation strategies, despite being still poorly explored. Trees are able to record and store many information useful to identify, assess and reconstruct the environmental factors influencing their growth and adaptation capacity. This paper reports a case study realized with the aim of (i) define the tree growth trends, then analysing possible correlations with the climatic trends (Standardised
Precipitation-Evapotranspiration Index; SPEI), and for (ii) evaluate the intrinsic Water Use Efficiency
(WUEi) of two tree species, common across the Europe, occurring in pure and mixed forest
stands through the support of isotopic analysis.
The study area is located on the Italian southern Alps (Ranzo, Tn), at an altitude of about 1000 m s.l.m.. Three plots with similar pedo-climatic conditions were identified, as hereinafter described. The first two plots are characterized by monospecific stands, respectively dominated by E. beech (Fagus sylvatica L.) and S. pine (Pinus sylvestris L.). The third plot refers to a mixed stand of both the previous species. In particular, we analysed
δ13C in tree rings in order to assess the annual changes in WUE
i. The influence of the
stomatal conductance was investigated through δ18O data. Our results indicated that: (i)
Basal Area Increments (BAI) consistently increased in all stands (except for pure stand of S. pine) in the 1994÷2003 period; (ii) SPEI highlighted a drought period between 1991 and
2007; and (iii) the WUEi values resulted generally higher in pure than in mixed stands. The
divergence between BAI and SPEI values in the ’90 and early ’00 may be explained by anthropogenic activity. Indeed, our findings showed that past forest management (namely thinning) realized in the study area resulted more influent on tree growth than the climatic conditions (temperature and precipitation).
1.2 Introduction
European forests play a very important role as providers of ecosystem services, functions and goods (Stenger et al., 2009). Forest management has to deal with uncertainties related to the adaptation of forest ecosystems to environmental changes. In particular, climate change exerts a growing pressure on mountain regions and, therefore, there is a need to emphasize forest systems that are resilient to climate change and natural disturbances. Mixed-species forests are considered more resistant to disturbances and extreme events than mono-specific stands (H. Pretzsch et al., 2013) and may provide higher levels of multiple ecosystem services (Gamfeldt et al., 2013; Loreau and Hector, 2001). These results could be attributed to the niche complementarity, which reduces the competition for resources in mixed stands (Morin et al., 2011), and in favourable interaction between the two populations of plants within a mixed stand (Yachi and Loreau, 2007). Differences between species in shade tolerance, crown phenology, canopy structure and root depths have been associated to increased productivity in mixed stands (Brassard et al., 2013; Forrester and Albrecht, 2014; Toïgo et al., 2015). However, mixed-species forests are not always more productive and stable than mono-specific stands (Epron et al., 2013; Grossiord et al., 2014; Merlin et al., 2015), which suggests the need for carefully considering the reciprocity of species assemblages and the specificity of site conditions (del Río et al., 2016). Understanding dynamics (growth, resistance, productivity, etc.) of mixed forests is very important to define proactive management measures towards sustainable adaptation to and mitigation of climate change (Condés and del Río, 2015). In particular, a better understanding of the exceeding/decreasing productivity of mixed stands in comparison with pure stands is essential for both ecology and management of forest ecosystems.
Although climatic signals are often confounded with management effects, warming temperatures are likely to induce changes in species distribution and community composition
in mountain regions by altering physiological thresholds and disturbance regimes (Gonzalez et al., 2010). In this context, extreme climatic events are very useful pointers to highlight the reactions of forest ecosystems to disturbance patterns and to allow in-depth study of growth-climate relationships (H. Pretzsch et al., 2013). Drought, in particular, is one of the most relevant indicators for response analyses of forest ecosystems in terms of tree growth (Pretzsch and Dieler, 2011), allometric modification (Hans Pretzsch et al., 2012), water-use efficiency (Tognetti et al., 2014) and plant mortality (McDowell et al., 2008). Tree responses to drought depend on species, age, and size, as well as within-stand competition for resources and other issues associated with stand structure (Cavin et al., 2013; Lloret et al., 2011; Pretzsch and Dieler, 2011; Zang et al., 2012). The sensitivity of plants to drought varies according to the severity and duration of stress, and the combination with other stress factors (Niinemets, 2010). Increasing drought exposure could alter the competitive hierarchy between tree species with distinct drought tolerance, resulting in shifts in tree species composition in mixed forest stands (Cavin et al., 2013). Stable carbon and oxygen isotopes
from tree rings (d13C, d18O) provide a continuous, annually resolved record of environmental
conditions and show stronger correlations between tree individuals and environmental variables than annual growth rings that are laid down during each growing season
(McCarroll and Loader, 2004). Tree-ring d13C is controlled by the balance between stomatal
conductance (i.e., supply of CO2) and photosynthetic rate (i.e., demand for CO2), and is often
used to calculate intrinsic water-use efficiency (WUEi) and differences in water use between
trees (McCarroll and Loader, 2004). Restrictions in water availability generally reduce
stomatal conductance and intercellular CO2 concentration (Ci), leading to an increase in
d13C, though light limitation of photosynthesis increases Ci. Conversely, tissue d18O is not
plant water exchange, so that combined measurements of d13C and d18O allow stomatal and
photosynthetic effects on d13C to be teased apart (Barnard et al., 2012; Tognetti et al., 2014).
Below-ground and above-ground mixing effect can change over time, productivity and stability advantages of mixed vs. pure stands becoming weaker, due to changes in resource acquisition and species interactions with stand development (Cavard et al., 2011). In this study, we considered two important tree species, S. pine (Pinus sylvestris L.) and E. beech (Fagus sylvatica L.), which are widely distributed throughout Europe (Pretzsch et al., 2015). S. pine and E. beech growing in mono-specific and mixed-species stands allow for inferring the effects of admixture on functional traits and tree growth, comparing an early and light-demanding conifer and a late-successional and shade-tolerant broad-leaves (del Río et al., 2016). This mixture was found to show significant mixing effects in terms of productivity and structural heterogeneity (Pretzsch et al., 2016, 2015).
Our main objectives were: (i) to investigate whether there exist differences in WUEi of S.
pine and E. beech from mixed-species and mono-specific stands, and their response to drought periods; (ii) to elucidate if mixing species of contrasting traits increases stand
productivity; (iii) to detect the seasonal climatic signals that control tree ring d13C and d18O
in the studied species. Aiming at better understanding the inter-specific dynamics of E. beech and S. pine under a global warming scenario, we hypothesized that: (i) differences in productivity between mixed-species and mono-specific stands decreases over time, due to the temporal dynamics of resource partitioning and the species differences in functional
traits; (ii) WUEi is higher in mixed-species than in mono-specific stands, because of
1.3 Materials and Methods
1.3.1 Study area
The study area is located near the village of Ranzo (Trento, Northern Italy), at an altitude of about 1000 m a.s.l.. The site morphology is flat or gently sloping, with moderately deep soil, high organic matter content in the surface and bedrock mainly dominated by limestone. Three plots were identified, as hereinafter described. The first two plots are characterized by monospecific stands, dominated respectively by E. beech (Plot PF) and S. pine (Plot PP).
The third plot refers to a mixed E. beech-S. pine stand (Plot MFP). The plots have similar site conditions in terms of geographical location, topography, aspect, climate, soil substrate, and soil type. The pure stands are used as the reference for the mixed stands and for the quantification of mixing effects.
The pure stands were selected when the corresponding species represented ̴ 90% of the stand basal area. The mixed stand was defined as the stand in which the two species of interest together represented at least ̴ 80% of the total stand basal area, and the sum of the basal area of other species was lower than that of each of the two species studied.
1.3.2 Meteorological data and drought index
The climatic data available for the study area cover only a short period and they are also spatially fragmented. For this reason, interpolated climatic data were here used, deriving them from a spatial downscaling of European climate data. In particular, a downscaled version of E-OBS in combination with WorldClim climate surfaces to obtain a 0.0083° (about 1 × 1 km grid) resolution climate data set (Moreno and Hasenauer, 2015) was used. Monthly minimum, mean and maximum temperatures, as well as total monthly precipitation
from 1951 to 2012 were considered.
In order to assess the water availability for tree growth, but also to detect and monitor the drought episodes, a multi-scalar drought index (Standardized Precipitation Evapotranspiration Index; SPEI) based on climatic data was calculated (
https://climatedataguide.ucar.edu/climate-data/standardized-precipitation-evapotranspiration-index-spei). More in detail, the SPEI uses the monthly difference between precipitation and potential evapotranspiration (PET) at different time scales (Di =
Pi -PETi) (Vicente-Serrano et al., 2010). It merges the sensitivity of Palmer Drought Severity
Index (PDSI) (Palmer, 1965) to changes in evaporation demand with the advantages of the Standardized Precipitation Index (SPI; Mckee et al. 1993). The multi-scalar features of SPEI allow to identify different drought types, their duration and intensity, as well as the time-lag
of drought episodes (http://sac.csic.es/spei/home.html).
1.3.3 Tree sampling and ring-width chronologies
We inventoried the plots in order to derive the dendrometric state variables at the stand level. Table 1 gives an overview of the field measurements. Furthermore, 20 dominant trees were sampled, and two increment cores at breast height were collected from each tree. In order to avoid the effect of any wood alteration and of exogenous disturbances on ring growth, only trees without abrasion scars or other visible evidence of injury were selected. The summary of structural data are reported in the Table 1.
Ring widths were measured with a resolution of 0.01 mm using the LINTAB measurement equipment (Frank Rinn, Heidelberg, Germany) fitted with a Leica MS5 stereoscope (Leica Microsystems, Germany); tree ring widths were visually cross-dated and statistically verified with the software TSAP software package and COFECHA (Holmes, 1983). Once
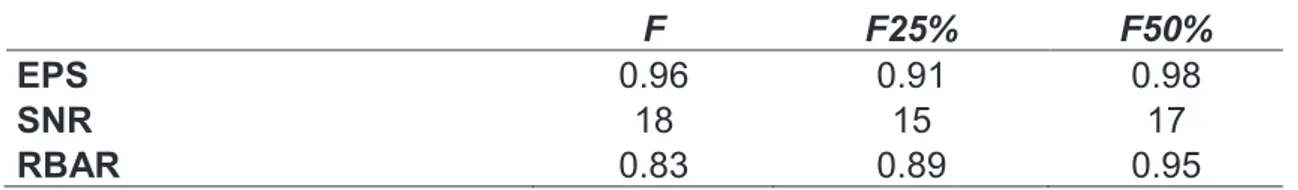
all measurement series had been validated, tree-ring chronologies were developed for the three stands. The ARSTAN program was used to process the tree ring data into final chronologies (Cook et al., 1990) and the standard deviation (SD), the expressed population signal (EPS) and the mean RBAR, were calculated for the three chronologies (Table 2). Ring width was converted into tree basal area increment (BAI) according to the following standard formula:
BAI~ π (r2n- r2 (n-1)) (1)
where r is the radius of the tree and n is the year of tree-ring formation. In order to examine the mean growth trend for the dominant trees, BAI for each year was averaged over all the individuals, to remove variation in radial growth attributable to increasing circumference.
We worked with mean not-standardized BAI values to preserve the long-term cumulative effects of climatic events on tree growth.
1.3.4 Stable isotope analysis
The five best cross-dated trees per site and species (i.e. those showing the highest correlation with the residual ring-width chronology) were selected for carbon and oxygen isotope measurements. Rings corresponding to the same year were pooled into a single sample before analysis. Every ten years, however, tree rings were individually handled and analysed (1951, 1961, etc.) to have an estimate of between-tree variability in the isotopic signal. The resulting samples were homogenized to a fine powder with a ball mixer mill (Retsch MM301, Haan, Germany).
The stable isotope compositions (C and O) were measured at the IRMS Lab (SUN, Caserta, Italy) by continuous-flow isotope ratio mass spectrometry (Delta V plus Thermo electron corporation, Bremen Germany), using 0.06 mg of dry matter for 13C measurements, 0.3-0.6 mg for 18O determinations.
The δ13C series were corrected for the Suess Effect (decrease in δ13C of atmospheric CO2
since the beginning of industrialization), resulting from the emission of fossil carbon dioxide, which is depleted in 13C (Francey et al., 1999; McCarroll and Loader, 2004). The isotopic compositions are expressed in delta notation (‰) relative to accepted reference standards: Vienna Pee Dee belemnite (VPDB) for carbon isotope values and Vienna Standard Mean Ocean Water (VSMOW) for oxygen isotope values. The standard deviation for the repeated analysis of an internal standard (commercial cellulose) was better than 0.2‰ for carbon and better than 0.4 ‰ for oxygen.
1.3.5 WUEi
We used the d13C measured in tree rings to calculate the intrinsic WUE since: d13C
plant = d13C atm-a-(b-a)·(ci/ca) (2)
where ci is the internal leaf CO2 concentration, ca is the atmospheric CO2 concentration, a is
the fractionation factor due to CO2 diffusion through stomata (4.4‰) and b is the
fractionation factor due to Rubisco enzyme during photosynthesis (27‰).
Deriving ci from equation 2, the formula reported by Ehleringer and Cerling (1995) was used
to calculate WUEi:
WUEi= A/gs= (ca-ci)/1.6 (3)
The carbon isotope discrimination D represents the difference between the carbon isotopic ratio of atmospheric CO2 (d13C air) and of plant organic matter (d13C plant), calculated as:
D= (d13C
air – d13C plant)/(1+d13C plant) (4)
We used d13C
air estimated values for the period 1960–2003 from McCarroll and Loader
(2004), and measured values for the period 2004-2012 available online (http://www.esrl.noaa.gov/gmd/), while d13C
plant are the values measured in the rings of our
samples.
Combining equation 2 and equation 4, we can derive ci as:
ci = ca [(D – a) / (b – a)] (5)
Inserting ci from equation 5 into equation 3, we obtain:
WUEi = (ca-ci)/1.6 = [ca – ca (D-a / b-a)]1/1.6 = ca [(1-(D-a / b-a)) 1/1.6] (6)
As said before, a and b are known values and ca is the concentration of CO2 in the
atmosphere, estimated for each year and obtained by NOAA (http://www.esrl.noaa.gov/, Mauna Loa station).
1.3.6 Data treatment and statistical analysis
In order to extract the climatic data within each sampling plot, a multi-step method was developed starting from the European climate data grid developed by Moreno and Hasenauer (2015).
At first, a R-script was settled to automatically identify, for each inventory plot (X, Y coordinates), the corresponding 1 km2 cell from the climatic data grid. The climatic
information at local scale were extracted from the grid and then organized using a specifically built C# program. Finally, a R-script was developed using the R-Package ‘SPEI’
for calculating the SPEI from the climatic data sets within each plot.
The BAI and stable isotope values were than correlated with the monthly precipitation and temperature. In order to simplify and explore the large amount of data, a PCA approach was applied. Multiple correlation analyses were then used and the relations between the considered variables were highlighted using the Pearson’s correlation coefficients.
SPEI was used as composite index of climate data (temperature and precipitation) in order to further clarify the relations between growth-climate variables. SPEI was calculated for the period ranging from October of the previous year to September of the current year (SPEI12sep). This methodological approach avoids the correlate the tree growth-trend to climate parameters in the months following the cessation of the seasonal growth. The correlation were realized within the same species (beech and pine), occurring both in the pure and mixed stands.
A generalized linear mixed model (GLMM) was used taking into consideration the differences in terms of stand and species. The GLMM makes it possible, incorporating the random effects in the model and allowing the non-independence of the observations (Zuur et al., 2013). Moreover, the intercept and slope of the model were analysed considering the site’s random effect. Values were considered significant at a probability level of 5% and R-squared values were used to assess the impact of random effects (Nakagawa et al., 2013). Afterwards, correlation and time series analyses were carried out using XLSTAT 2013 (Microsoft Corp., Seattle, WA, USA), while the GLMM was carried out using the R software.
1.4 Results
1.4.1 Tree-Ring, BAI, WUEi , d13C and d18O
Tree chronologies refer to the period 1886÷2014; however, since the climatic data were available for a shorter period, only the time-span 1951÷2012 was used for further analyses. Tree ages varied significantly among the investigated stands. In detail, as average (years ± standard deviation), the following results were obtained: 124 ±5 in the pure S. pine stand (PP) and 55 ±8 in the pure E. beech stand (PF); for the mixed stand (MFP), tree ages were of 86 ±31 and 63 ±9 for S. pine and E. beech, respectively. (Table 1).
The BAI curve showed that pure stands grew more than mixed stand, but with a few distinctions. In detail, the BAI curve of E. beech in the mixed stand (MFP) was constantly lower than BAI curve of E. beech (growing in the monospecific stand (PF)). Until mid-80s, S. pine revealed higher BAI values when growing in the pure stand then in the mixed one; later, an opposite trend was observed. Furthermore, a consistent increase of BAI in all stands
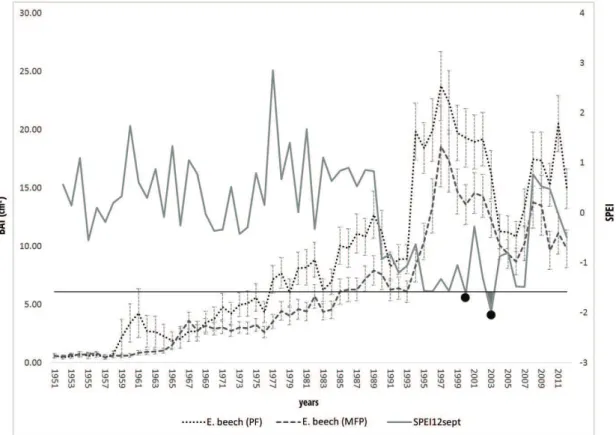
(except for pure S. pine stand) in the 1994÷2003 period was found (Fig. 1a and 1b).
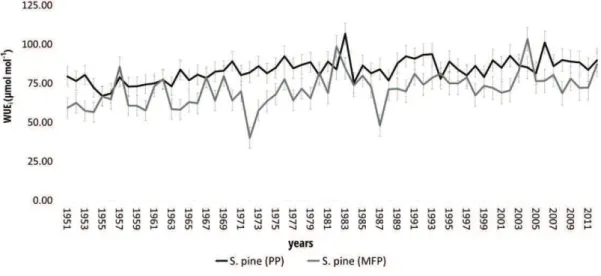
The WUEi values (Fig. 2) of pure stands (E. beech (PF) = 89±8 µmol mol-1; S. pine (PP) =
84± 7 µmol mol-1), derived from tree-ring d13C measurements (Fig. 3a and 3b), were higher than mixed stand (beech (MFP) = 67±10 µmol mol-1; S. pine (MFP) = 71±11 µmol mol-1)
for both the investigated species. Moreover, the WUEi showed a slight uptrend over the
investigated time-span for all the studied species combination.
d18O analyses for beech trees revealed similar values between pure and mixed stand (Fig. 4a). On the contrary, for the S. pine (Fig. 4b), the average d18O value was higher in the pure (d18O = 26.70± 2) than in the mixed stand (d18O = 25.20± 1).
1.4.2 Drought index, growth-climate correlations and GLMM
Exploratory climate data analysis for the three stands showed a period of low precipitation between 1991 and 2007 (period 1991÷2007 = 467 mm/year, on average; 1951÷2012 = 750 mm/year, on average). Details are reported in Figure 5. Temperature showed a significant increase from 80’s, both for maximum and minimum values, as observed in other studies across the Europe (Hansen et al., 2010).
SPEI index highlighted a drought period between 1991 and 2007, according to the lower amounts of precipitation recorded in the same period (Fig. 1a and 1b and Fig. 5).
Furthermore, multiple correlation analyses were applied for highlight the relations between
WUEi, BAI, SPEI and d18O. In the mixed stand, we found a significant positive correlation
between WUEi and d18O, with p-value <0.001 for E. beech (r=0.363) and p-value <0.005 for
S. pine (r= 0.351). In the pure stands, no significant correlations between WUEi and d18O
were found, for both the investigated species.
S. pine in the pure stand showed a negative correlation between WUEi and BAI (r=0.365;
p<0.005), while no correlations were found for E. beech growing in the monospecific stand. Mixed stand highlighted a significant positive correlation between WUEi and BAI only for
E. beech (r=0.449; p<0.001).
The SPEI obtained for E. beech showed a negative correlation with BAI both in the pure and mixed stand. S. pine growing in the mixed stand (MFP) revealed a negative correlation between SPEI and BAI. On the contrary, SPEI of S. pine (PP) showed a positive correlation with BAI (r=0.348). However, these correlations were not significant.
Finally, the GLMM model was used to consider the management variable and its effect on tree growth, together with the natural variability of the sites and the climatic conditions (SPEI12sep). Results highlighted a significant role of forest management in influencing the tree growth among the investigated years. In detail, forest management is significantly
related with growth (p<0.05), while the relation with the SPEI was not significant. The marginal R squared (R2m) values take into account the fixed effects, while the conditional ones (R2c) take into account the fixed effects plus the random effects. In our case, the site’s random effect increased the R squared (from 0.12 to 0.39), as showed in Table 3.
1.5 Discussion
The growth dynamics in our study showed that beech has not benefited from the presence of Scots pine. In contrast to other studies in which the mixture of species appears to be favorable in terms of productivity and radial growth (Morin et al., 2011; Hans Pretzsch et al., 2013; Vallet and Pérot, 2011), in our case the pure plot of beech turns out to have a consistently higher growth during the period. However, the same Pretzsch et al., (2015) stated that a generally overyielding by mixture species of beech and pine was reached, but also that there was a large variability in relative productivity from case to case. Thus, in our stands, the benefits caused by complementarity of ecological traits (Condés et al., 2013; Kelty, 1992) were are not found as the primary drivers of beech.
Growth trend of S. pine showed a different behavior, which result lower in the pure stand compared to the mixed stand since mid ’80.
To explore such trends we considered the climate in the analysis. The plots, because of their proximity, present similar climate conditions. We used an index of drought stress (i.e. SPEI) in order to understand the response to climatic variations in the timeframe concerned. Increasing resilience and resistance to climatic stress is demonstrated to derive by the interactions between beech and pine (Forrester, 2014). This is partial confirmed by our results, for which the mixture of the two species does not limit the growth, also in
unfavorable weather conditions. However, the pure beech stand growths even better than the mixed one.
On the contrary, the growth of S. pine in pure stand since ’90 seems to be influenced by the adverse weather conditions recorded by SPEI (1991-2007 period). According to drought classification of Mckee et al. (1993), SPEI (i.e. SPEI12sep) highlighted only two years as severely dry (i.e. 2000 and 2003), while most of remaining years of the period 1991-2007 was identified as moderately dry or mild dry. Sensitivity of European beech in terms of growth under climatic variations and water deficits was reported in many studies (Piovesan and Adams 2001; Nielsen and Jørgensen 2003; Rose et al. 2009; Metz et al. 2013; Cavin and Jump 2016). Nevertheless, other studies showed different results highlighting resistance to drought in pure or mixed stands of beech (e.g., Tegel et al., 2014). Moreover, as confirmed by our findings, the extreme drought of 2003 in Central Europe did not result in substantial reductions the radial growth of beech (Leuzinger et al., 2005) in both pure and mixed plots. Regarding the S. pine, our results showed that mixture with beech has facilitated a better response in terms of growth in the period of less favorable weather conditions. On the contrary, before the time-frame as identified by SPEI (1991-2007), the pure plot had higher growth performances in comparison with the mixed one. The only positive correlation between BAI and SPEI (despite not significant) was observed for the pure S. pine plot. Therefore, the growth-climate analysis highlights that the mixture of the two species had benefits in terms of growth, or lower impact of drought on growth, only for S. pine.
The complexity of the relationships between the two species and the conflicting nature of the results in comparison with other studies need further investigation. The analysis of BAI highlighted above-average growth during the 1994÷2003 period, but no for S. pine growing in pure conditions. These aspects induce to consider management as an additional variable in the analysis. Therefore, we used a GLMM to assess the impact of management on the