1
1.INTRODUZIONE
1.1 IL FENOMENO DELLA FLUORESCENZA
La fluorescenza è la proprietà che hanno alcune sostanze di emettere luce quando vengono colpite da radiazioni luminose (raggi ultravioletti o luce visibile) (Peter Atkins; Julio De Paula, Elementi di Chimica Fisica, Terza edizione italiana, Bologna, Zanichelli, 2007; Walter J. Moore, Chimica Fisica, Padova, Piccin-Nuova Libraria, 1990). Il termine fluorescenza deriva dalla fluorite, un minerale composto da calcio e fluoro, grazie a cui è stato scoperto il fenomeno. La fluorescenza si distingue dalla fosforescenza, altro fenomeno che comporta l'emissione di luce, in quanto i materiali fluorescenti cessano di essere luminosi al cessare dello stimolo che ne determina la luminosità, invece nei materiali fosforescenti la luce continua ad essere emessa per un certo periodo dopo la fine dello stimolo.
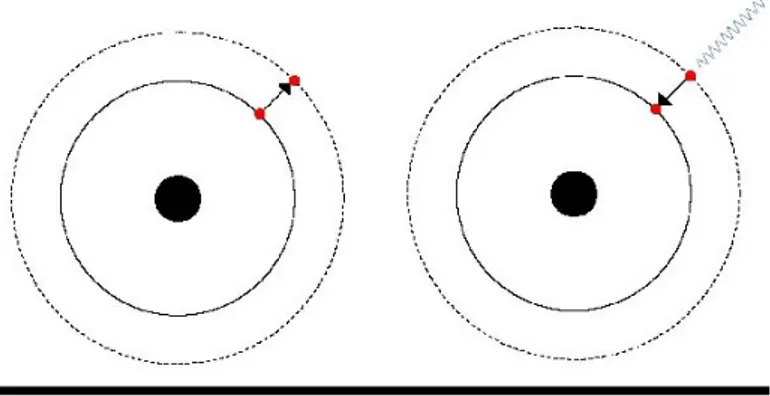
Il fenomeno della fluorescenza è determinato dall'assorbimento di energia da parte degli atomi del materiale: in seguito a tale assorbimento energetico gli elettroni si spostano da un livello energetico inferiore ad uno superiore (orbitale più esterno) (Figura 1).
2 La permanenza al livello energetico superiore è brevissima (miliardesimi di secondo) e quando gli elettroni tornano al livello energetico originario liberano l'energia assorbita sotto forma di radiazioni elettromagnetiche. Poiché la resa energetica non è mai totale, le radiazioni liberate hanno lunghezza d'onda superiore rispetto a quella dell'energia eccitante: ad esempio se l'eccitazione è stata ottenuta con raggi ultravioletti le radiazioni liberate sono sotto forma di luce visibile o di raggi infrarossi (figura 2).
Fig. 2 Schema di assorbimento e restituzione di energia da parte di fluorocromi
Sulla base di questa transizione elettronica è possibile anche distinguere i processi di fluorescenza e fosforescenza:nella fluorescenza la radiazione è generata in virtù di transizioni degli elettroni tra stati con la stessa molteplicità di spin (per esempio S1→S0), mentre nella fosforescenza la transizione coinvolta comporta variazione della molteplicità di spin (transizioni tripletto-singoletto).
Esempi di fluorescenza sono visibili in tutti i materiali che contengono pigmenti fluorescenti, come ad esempio nell'inchiostro degli evidenziatori, nelle vernici fluorescenti, ma anche in sostanze naturali come la clorofilla.
Quando i materiali colpiti da radiazioni UV emettono naturalmente fluorescenza si parla di fluorescenza primaria. La maggior parte delle sostanze emette fluorescenza solo dopo marcatura con fluorocromi e in questo caso si parla di fluorescenza secondaria o indotta.
3 Ogni fluorocromo ha una precisa lunghezza d’onda di eccitazione e di emissione e per la sua determinazione si fa uso di un fluorimetro dotato di filtri di eccitazione e di emissione compatibili con le caratteristiche del fluorocromo prescelto. Quanto più intensa è la luce utilizzata per illuminare un campione e tanto maggiore sarà il numero di molecole di fluorocromo che vengono eccitate e quindi il numero di fotoni emessi. Se la luce di eccitazione ha una lunghezza d’onda ed un’intensità costanti (ad esempio un laser con sorgente luminosa) il numero di fotoni emessi e quindi l’intensità della luce fluorescente prodotta, saranno in funzione del numero di molecole di fluorocromo presenti.
I fluorocromi si differenziano per l’intensità di fluorescenza che sono capaci di emettere (brightness). Tale proprietà può essere influenzata da fattori fisici e chimici come la temperatura, il pH e la forza ionica. Un ulteriore fattore che può condizionare l’efficienza di un fluorocromo è il fenomeno della foto-distruzione (photobleaching) causato da luce intensa, che comporta la perdita permanente della capacità di fluorescere.
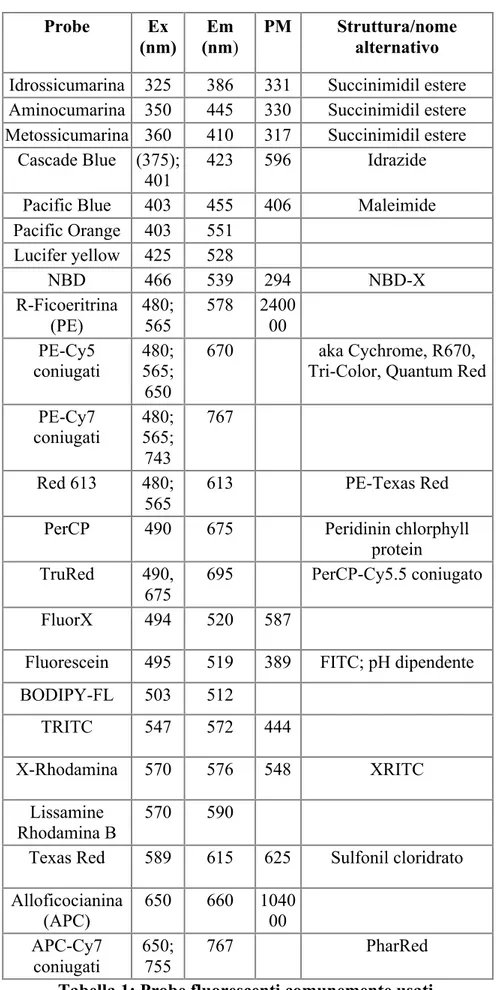
La fluorescenza è influenzata anche dalla struttura della molecola. Ad esempio le molecole rigide che presentano sistemi di doppi legami coniugati si prestano molto bene alla fluorescenza: in particolare molecole dove vi sono strutture aromatiche, nelle quali per il fenomeno di risonanza i doppi legami risultano sparsi per tutta la struttura, se eccitate danno luogo a transizioni π→π* e quindi favoriscono la fluorescenza. Esempi di fluorocromi che vengono maggiormente utilizzati nell'ambito delle tecniche biologiche sono l’isotiocianato di fluoresceina, conosciuto anche con l'acronimo FITC, l’isotiocianato di tetrametilrodamina (TRITC), la curarina e la cianina (tabella 1).
4 Probe Ex (nm) Em (nm) PM Struttura/nome alternativo Idrossicumarina 325 386 331 Succinimidil estere Aminocumarina 350 445 330 Succinimidil estere Metossicumarina 360 410 317 Succinimidil estere
Cascade Blue (375); 401
423 596 Idrazide
Pacific Blue 403 455 406 Maleimide Pacific Orange 403 551 Lucifer yellow 425 528 NBD 466 539 294 NBD-X R-Ficoeritrina (PE) 480; 565 578 240000 PE-Cy5 coniugati 480; 565; 650 670 aka Cychrome, R670,
Tri-Color, Quantum Red PE-Cy7 coniugati 480; 565; 743 767 Red 613 480; 565 613 PE-Texas Red
PerCP 490 675 Peridinin chlorphyll
protein TruRed 490,
675
695 PerCP-Cy5.5 coniugato
FluorX 494 520 587
Fluorescein 495 519 389 FITC; pH dipendente
BODIPY-FL 503 512 TRITC 547 572 444 X-Rhodamina 570 576 548 XRITC Lissamine Rhodamina B 570 590
Texas Red 589 615 625 Sulfonil cloridrato Alloficocianina (APC) 650 660 1040 00 APC-Cy7 coniugati 650; 755 767 PharRed
5
1.1.1 Fluorocromi
L’impiego di fluorocromi come sonde per lo studio di proteine si è rilevato una valida alternativa ai metodi tradizionali di studio e di ricerca in campo biologico. Essi forniscono un’ampia gamma di informazioni sui recettori a cui sono affini, sulla loro localizzazione, sui movimenti e l’interazione dei recettori stessi in cellule vitali. Inoltre i ligandi fluorescenti possono rappresentare un’alternativa utile rispetto ai ligandi radiomarcati grazie alla loro elevata sensibilità, sicurezza, nonché ai costi piuttosto limitati e alla disponibilità di un alto numero di composti fluorescenti.
1.1.2 Utilizzo dei fluorocromi per la diagnosi tumorale
Recenti studi condotti utilizzando composti fluorescenti, capaci di legare specificamente quei recettori la cui densità risulta alterata nei tumori, hanno evidenziato la possibilità di utilizzare tali ligandi come nuove sonde per l’imaging diagnostico. Il successo di tali osservazione dipende anche dalla capacità del probe di legare specificatamente il recettore a basse concentrazioni (nanomolare) per le tecniche di tomografia a emissione di positroni (PET) e di tomografia a emissione di fotone singolo (SPECT). Le stesse sonde, se usate in tecniche quali la tomografia computata (CT) e la risonanza magnetica tomografica (MRT) richiedono invece concentrazioni più elevate (millimolari).
In genere ligandi usati come agenti di imaging devono avere un’alta affinità per il recettore e tale recettore deve risultare sovraespresso nelle cellule tumorali. Inoltre deve essere possibile eliminare il ligando in eccesso dal tessuto in esame mediante semplici lavaggi quando. Questo può rappresentare un limite per gli studi perché esperimenti condotti su tessuto hanno evidenziato che i lavaggi previsti nelle tecniche impiegate per la visualizzazione spesso causano un’alterazione dell’equilibrio chimico con conseguente distacco del ligando fluorescente dal TSPO. Infine il probe deve dare una
6 buona risoluzione di immaging anche a basse concentrazioni (Emanuela Sega, Philip Low, 2008).
Recentemente è stato proposto come target di imaging diagnostico tumorale una proteina denominata Traslocatore Proteico mitocondriale (TSPO). Il TSPO in effetti riveste un ruolo chiave nella regolazione della proliferazione cellulare, controllando la morte cellulare programmata. Inoltre ha una specifica funzione anche nella sintesi degli steroidi, che a loro volta possono regolare l'omeostasi cellulare. La densità di tale proteina è stata riscontrata elevata in differenti tipologie di tumori ed in particolare è risultata direttamente proporzionale all'aggressività della neoplasia. Nei gliomi, ad esempio, i livelli di espressione del TSPO sono stati correlati con la malignità del tumore (Jason R. Buck et al., 2011) e per tale motivo sono stati sviluppati nuovi radioligandi, specifici al TSPO, come sonde di imaging per la diagnosi di tumori (Doorduin, Venneti 2006).
1.2 IL TRASLOCATORE PROTEICO MITOCONDRIALE
1.2.1 Struttura e localizzazione del TSPO
Il TSPO, precedentemente denominato recettore periferico delle benzodiazepine (PBR, Periferic Benzodiazepine Receptor) fu inizialmente scoperto come sito di legame per la benzodiazepina diazepam (CBR, Central Benzodiazepine Receptor). Il TSPO è una proteina transmembrana con massa molecolare pari a 18 KDa costituita da 169 amminoacidi, altamente idrofoba e ricca di triptofano.
La proteina TSPO è soprattutto espressa sulla membrana mitocondriale, ma livelli più bassi si ritrovano anche in altri compartimenti subcellulari, come la membrana plasmatica e la frazione nucleare.
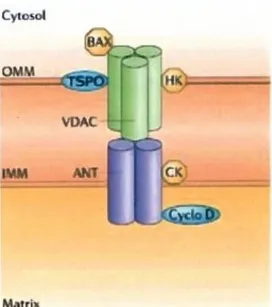
7 Questo recettore può essere strettamente associato in un complesso proteico formato dal canale anionico voltaggio-dipendente (VDAC, voltage dipendent anion channel) e da un trasportatore di nucleotidi adeninici (ANT). L’associazione di queste tre proteine forma il poro di transizione della permeabilità mitocondriale (MPTP) (Fig. 3)
Fig. 3 Struttura del complesso
La localizzazione subcellulare del TSPO è stata studiata in vari organi: i siti auotradiografici di alcuni ligandi specifici al TSPO avevano una distribuzione istochimica simile a quella della citocromo ossidasi e delle monoammino ossidasi (MAO), markers per i mitocondri. Il fatto che un’analoga correlazione non sia stata trovata con i markers per i nuclei, i lisosomi, i perossisomi ed il reticolo endoplasmatico, ha suggerito che il TSPO è associato con la frazione mitocondriale. In particolare, studi successivi hanno dimostrato che il TSPO è associato alla membrana esterna del mitocondrio poiché la sua densità, per mg di proteina, risulta maggiore nella membrana esterna rispetto al mitocondrio nel suo insieme. Dal momento che è stata evidenziata la presenza del TSPO anche sulla membrana mitocondriale interna è stato
! "!
30 KDa (ANT adenine nucleotide translocator).Insieme, queste tre proteine, vanno a formare sulla membrana mitocondriale il poro di transizione della permeabilità mitocondriale (MPTP, mitochondrial permeability transition pore).
8 suggerito che probabilmente tale recettore sia localizzato sui siti di contatto tra le due membrane mitocondriali.
Per quanto riguarda la localizzazione tissutale, il TSPO è stato evidenziato in quasi tutti i tessuti periferici di mammifero.
Esso è espresso in particolar modo nei tessuti coinvolti nella steroidogenesi (ovaio, testicolo, surrene, placenta e tessuto gliale), ma si trova abbondantemente anche nel rene, nel fegato, nell’epitelio nasale, nei polmoni, nel cuore, nelle piastrine, nei linfociti e negli eritrociti. Al contrario è poco espresso nel tratto gastrointestinale. Nel sistema nervoso è espresso principalmente a livello delle cellule gliali.
1.2.2 Funzioni del TSPO
Il TSPO è capace di legare il colesterolo e favorire il suo trasporto dal citosol ai mitocondri, dove inizia la sintesi degli ormoni steroidei. Esso è inoltre implicato in meccanismi quali la respirazione cellulare, l’immuno- modulazione, il trasporto di porfirine, la biosintesi dell’EME, la modulazione dei canali del calcio, la regolazione del differenziamento, della proliferazione cellulare e la modulazione dei processi alla base della vita/morte cellulare (Gavish et al., 1999 Casellas et al., 2002). In particolare, il TSPO risulta fondamentale nei processi apoptotici in cui l’evento iniziale è la dissipazione del potenziale transmembrana del mitocondrio, causata dalla formazione e apertura dell’MPTP di cui il TSPO fa parte. Il periodico e transiente aumento della permeabilità mitocondriale è un evento fisiologico che porta alla sintesi di ATP. Tuttavia l’apertura prolungata del MPTP può innescare una serie di eventi che hanno importanti conseguenze sull’equilibrio bioenergetico della cellula. In particolare è stato osservato che l’apertura prolungata del MPTP provoca l’ossidazione del NADPH e del glutatione ridotto, la deplezione di ATP, la dissipazione del potenziale transmembrana e il disaccoppiamento della fosforilazione ossidativa. Inoltre, tale apertura influenza la
9 concentrazione degli ioni intracellulari, favorendo il rilascio di calcio dalla matrice e l’equilibrio colloido-osmotico, portando al rigonfiamento della matrice mitocondriale, fenomeno definito “swelling” mitocondriale.
L’apertura del poro favorisce anche l’efflusso dal mitocondrio di proteine e molecole, tra cui il fattore inducente l’apoptosi (AIF) e il citocromo c. La flavoproteina AIF traslocando nel nucleo, induce la condensazione della cromatina nucleare e la frammentazione del DNA. Nel citosol il citocromo c, interagendo con APAF-1 (fattore di attivazione dell’apoptosi e con la caspasi 9 porta all’attivazione della caspasi 3. Quest’ultima attiva numerosi enzimi importanti per indurre il riarrangiamento strutturale del nucleo, del citoscheletro e della membrana plasmatica, eventi caratteristici del processo apoptotico (Fig.4).
Fig.4 Processo di morte cellulare
L’MPTP quindi risulta un elemento critico nel processo della morte cellulare e rappresenta il punto di non ritorno dell’intero evento apoptotico. Il TSPO, che risulta far parte di questo poro, sembra possa regolarne le funzioni e, quindi, avere la capacità di
! ""!
Fig. 4 Processo di morte cellulare
1.6 TSPO come target terapeutico e marker diagnostico
Un’alterata espressione del TSPO può essere indice di diverse patologie tra cui, danni al cervello dovuti a varie forme di infiammazione e lesioni,alcune forme di epilessia, patologie neurodegenerative, come la sindrome di Huntigton e l’Alzheimer, ischemia. (Batarseh e Papadopoulos, 2010). Soprattutto però, un’alterata espressione di tale recettore, risulta evidente in due aree terapeutiche: la neuropsichiatria e l’oncologia.
Per quanto riguarda la neuropsichiatria, i ligandi del TSPO possono esercitare una modulazione, tramite neurosteroidi, sul complesso recettoriale del GABAA nelle cellule della glia.
10 indurre apoptosi. Ciò suggerisce che il TSPO possa assumere un importante ruolo nella crescita cellulare e nella progressione dei tumori.
1.2.3 Ligandi del TSPO
Numerosi ligandi affini al TSPO ed appartenenti a diverse classi chimiche sono stati progettati e sintetizzati al fine di influenzare la funzionalità del TSPO soprattutto per quanto concerne steroidogenesi e apoptosi. Da alcune di queste classi sono anche derivati i ligandi radiomarcati e fluorescenti, quali sonde del TSPO.
1.2.3.1 Modello farmacoforico proposto per il TSPO
Molte molecole, con struttura chimica diversa da quella delle benzodiazepine, come le isochinoline o le imidazopiridine, possono legarsi al TSPO con alta affinità. L’andamento parallelo delle SAR (relazione struttura-attività) di diversi derivati suggerisce che essi vadano ad interagire con il TSPO in modo del tutto analogo.
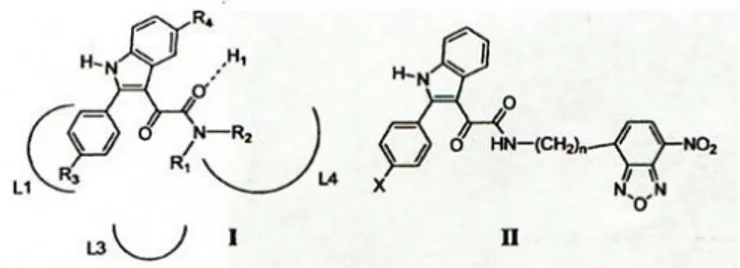
È stato proposto un modello farmacoforico che include tre regioni lipofile (siti L1, L2, L4) ed un gruppo donatore di idrogeno (sito H1) (Fig.5). È possibile ipotizzare che i punti fondamentali di interazione siano:
• Il carbonile ammidico che forma legami ad idrogeno con il sito donatore H1;
• Il sistema eterociclico azotato ed il gruppo 2-arilico, che vanno ad occupare le aree lipofile L1 ed L3;
11 Fig.5 Modello farmacoforo
1.2.3.2 Ligandi endogeni
Negli anni sono stati identificati diversi tipi di ligandi endogeni per il TSPO tra cui DBI (inibitore del legame del diazepam o endozepina) e i suoi metaboliti, porfirine (Snyder et al., 1987) e molecole benzodiazepino-simili (Rothstein et al., 1992). Il DBI è un polipeptide di 9 KDa, capace di spiazzare il [3H]diazepam sia dal recettore centrale per le benzodiazepine che dal TSPO (Costa E, Guidotti A, 1991). Le porfirine, le coproporfirine e le uroporfirine sono ligandi più potenti, mentre la biliverdina e la bilirubina, prodotti del catabolismo delle porfirine, legano debolmente il traslocatore proteico. Inoltre recentemente è stato scoperto che il colesterolo è un ligando endogeno ad alta affinità.
1.2.3.3 Ligandi di sintesi
I primi ligandi di sintesi dimostratisi affini per TSPO sono state le benzodiazepine ed in particolare all’interno di questa classe è stato possibile stabilire una gradualità nei valori di affinità e selettività per i siti di legame centrali e periferici. Il diazepam, ad esempio, si lega con elevata affinità al TSPO e al recettore centrale delle benzodiazepine, mentre il 4’-cloro derivato del diazepam, Ro 5-4864, si lega con affinità nanomolare al TSPO e con un’affinità micromolare al recettore GABAA (recettore initropo per il GABA),
! "!
Fig.2 Modello farmacoforo
1.4 Ligandi del TSPO
1.4.1 Ligandi endogeni
Negli anni sono stati identificati diversi tipi di ligandi endogeni per il TSPO tra cui DBI (inibitore del legame del diazepam o endozepina) e i suoi metaboliti, porfirine (Snyder et al., 1987) e molecole benzodiazepino-simili (Rothstein et al., 1992).
Il DBI è un polipeptide con un PM di 9 KDa, capace di spiazzare il [3H]diazepam sia dal recettore centrale per le benzodiazepine che dal TSPO
12 inoltre l’interazione del Ro 5-4864 con il TSPO varia a seconda della specie. Negli anni l’interesse farmacologico e chimico nei confronti del TSPO ha portato alla sintesi di ligandi strutturalmente diversi dalle benzodiazepine e dotati di una maggiore selettività. Tra questi ligandi troviamo i derivati isochinolincarbossammidici, dei quali il capostipite è il 1-(2-clorofenil)-N-(1- metilpropil)-3-isochinolincarbossiammide meglio conosciuto come PK 11195, che hanno dimostrato un’affinità nanomolare selettiva per il TSPO, i derivati indoloacetamidici come FGIN 1-27, i derivati pirisanoindoli (SSR 180575) ed i derivati fenossifenilacetamidi (DAA 1106 e DAA 1097) (Papadopoulos et al., 2006; Casellas et al., 2002; Manning et al., 2002).
Le caratteristiche che presentano sia le benzodiazepine che le isochinoline nell’interazione con il TSPO suggeriscono che i loro domini di legame siano sovrapposti o allostericamente accoppiati. Inoltre sulla base della dipendenza dall’entropia o dall’entalpia dell’interazione ligando-recettore, il PK 11195 è stato classificato come antagonista mentre il Ro 5-4864 come agonista (Le Fur et al., 1983). Tuttavia entrambi i ligandi possono indurre simili effetti in condizioni fisiologiche analoghe.
Fig.6 Ligandi del TSPO
! "!
Le caratteristiche che presentano sia le benzodiazepine che le isochinoline nell’interazione con il TSPO suggeriscono che i loro domini di legame siano sovrapposti o allostericamente accoppiati. Inoltre sulla base della dipendenza dall’entropia o dall’entalpia dell’interazione ligando-recettore, il PK 11195 è stato classificato come antagonista mentre il Ro 5-4864 come agonista (Le Fur et al., 1983). Tuttavia entrambi i ligandi possono indurre simili effetti in condizioni fisiologiche analoghe.
FGIN-1
13
1.2.3.4 Ligandi radiomarcati e fluorescenti
Ligandi radiomarcati sono stati progettati come sonde con lo scopo di valutare l’espressione e la distribuzione del TSPO nei tessuti ed essere quindi utilizzati anche a scopo diagnostico.
Tali sonde sono in genere derivanti da ligandi già noti di cui è mantenuta l’affinità e la specificità per il TSPO e a cui sono aggiunti gruppi marcanti radioattivi.
Numerosi di questi ligandi radioattivi, portanti 11C ed 8F, sono stati progettati per essre utilizzati a scopo diagnostico nella tecnica PET, ma hanno un limite per la loro breve emivita (Denora et al., 2011). Per tale motivo attualmente gli studi si stanno rivolgendo verso l’uso di ligandi fluorescenti che risultano essere una valida alternativa poiché sono anche composti che richiedono tecniche di studio più sicure per l’operatore, veloci e meno costose.
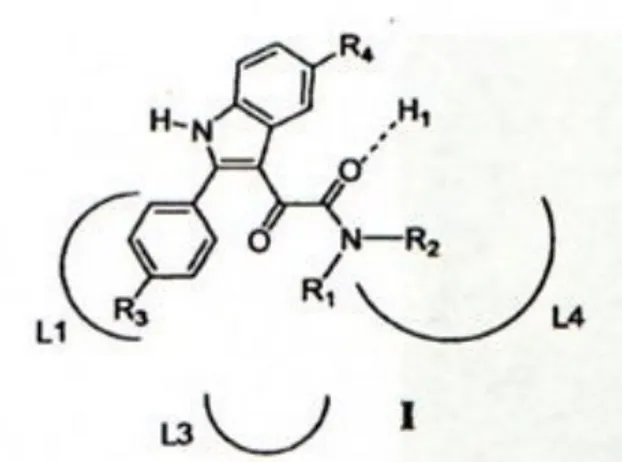
I ligandi fluorescenti specifici al TSPO, fino ad ora studiati, sono stati sintetizzati a partire dall’N,N-dialchil-2-fenilindol-3-ylgliossilamide, potente ligando del TSPO a cui è stato legato un gruppo fluorescente, 7-nitrobenzen-2-ossa-1,3-diazol-4-yl (NBD) che è stato scelto grazie alle sue piccole dimensioni che non influenzano l’affinità del ligando al recettore.
! "#!
1.2.3.4 Ligandi radiomarcati e fluorescenti
Ligandi radiomarcati sono stati progettati come sonde con lo scopo di valutare l’espressione e la distribuzione del TSPO nei tessuti ed essere quindi utilizzati anche a scopo diagnostico.
Tali sonde sono in genere derivanti da ligandi già noti di cui è mantenuta l’affinità e la specificità per il TSPO e a cui sono aggiunti gruppi marcanti radioattivi.
Numerosi di questi ligandi radioattivi, portanti 11C ed 8F, sono stati progettati per essere utilizzati a scopo diagnostico nella tecnica PET ma hanno un limite per la loro breve emivita (Denora et al., 2011). Per tale motivo gli studi si stanno rivolgendo verso l’uso di ligandi fluorescenti che risultano essere una valida alternativa poiché sono composti che richiedono tecniche di studio più sicure per l’operatore, veloci e meno costose. I ligandi fluorescenti specifici al TSPO, fino ad ora studiati, sono stati sintetizzati a partire dall’N,N-dialchil-2-fenilindol-3-ilgliossilamide (Fig.7), potente ligando del TSPO a cui è stato legato un gruppo fluorescente, 7-nitrobenzen-2-ossa-1,3-diazol-4-il (NBD) (Fig.8) che è stato scelto grazie alle sue piccole dimensioni che non influenzano l’affinità del ligando al recettore.
Fig. 7 N,N-dialchil-2-fenilindolgliossilamidi I nel modello farmacoforo/topologico e formula generale dei ligandi fluorescenti
14 Fig. 7 Struttura del NBD
Molti di questi composti portanti il gruppo NBD dimostrano una bassa fluorescenza in soluzioni acquose, ma danno alta emissione in solventi non polari o quando si legano alle membrane o a proteine idrofobe (Taliani et al., 2007).
15
2. SCOPO DELLA TESI
Fino ad ora numerosi radioligandi, capaci di misurare il livello di espressione del TSPO, sono stati progettati e testati sia sugli animali che sull’uomo. Per quanto riguarda i ligandi fluorescenti gli studi sono ancora in fase di sviluppo.
Sulla base del fatto che la presenza contemporanea di un gruppo chemioreattivo, capace di legare irreversibilmente il recettore, e di un gruppo fluoroforo può offrire una molteplicità di applicazioni, in questo lavoro di tesi sono stati saggiati alcuni composti con tali caratteristiche quali ligandi al TSPO. Questi derivati, appartenenti alla classe delle 2-fenilindol-3-ilgliossilamidi, presentavano un gruppo chemioreattivo (isotiocianato) capace di legare il recettore in maniera irreversibile e covalente. Inoltre ad uno di questi composti era anche legato un gruppo fluoroforo (NBD).
Tali caratteristiche, presenti su una singola molecola, potrebbero permettere di ottenere numerosi vantaggi sia sulle tecniche di purificazione, isolamento e caratterizzazione della proteina TSPO, che nei metodi di visualizzazione e determinazione della densità del recettore stesso.
16
3. MATERIALI E METODI
3.1 I LIGANDI DEL TSPO
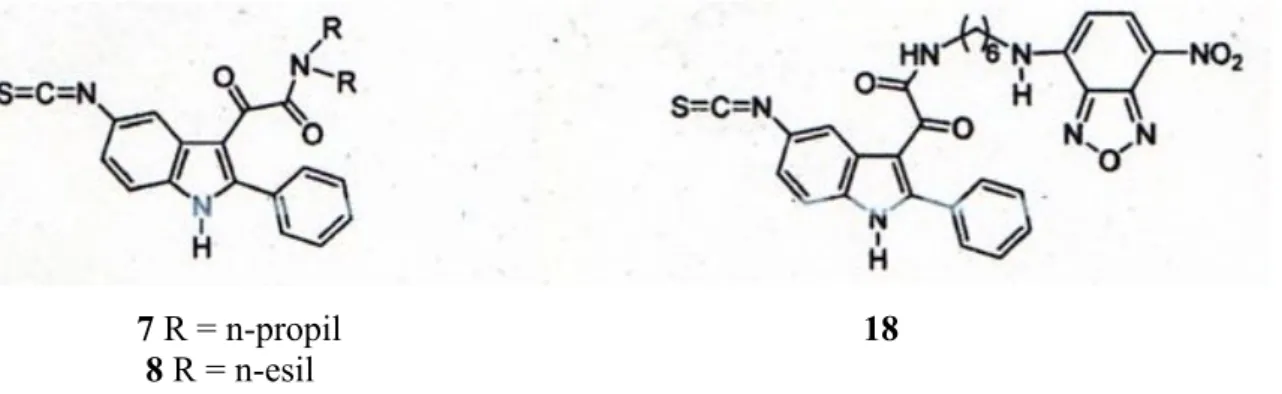
Nel presente lavoro di tesi sono stati saggiati 3 composti progettati e sintetizzati nel laboratorio diretto dal Prof. Da Settimo che legano selettivamente il TSPO con formula generale I descritta in fig. 9, derivati N,N-dialchil-2-fenilindol-3-ilgliossilamidi. Tali derivati sono stati sintetizzati a partire da una molecola di indolgliossilamide, riportato in letteratura come agonista parziale per i recettori BzR.
17 7 R = n-propil 18
8 R = n-esil
Fig. 10 Strutture dei ligandi per il TSPO
I ligandi caratterizzati in questo lavoro sono i composti 7, 8 e 18 (Fig.10).
I tre composti portano una molecola di isotiocianato e solo il composto 18 contemporaneamente all’isocianato presenta il gruppo NBD.
3.2 PREPARAZIONE DI MEMBRANE DI RENE DI RATTO
La preparazione delle membrane mitocondriali di rene di ratto è stata effettuata come descritto da Campiani et al.( 1996 ) e Trapani et al.( 1997 ).
Il rene è stato sminuzzato e sospeso 1:10 nel tampone Tris-HCl 50 mM, pH 7.4 (T50), contenente saccarosio 0.32 M, EDTA 1 mM e inibitori delle proteasi ( benzamidina 160 µg/ml, bacitracina 200 µg/ml, inibitori della tripsina 20 µg/ml).
La sospensione ottenuta è stata omogeneizzata e quindi centrifugata a 600 x g per 10 minuti alla temperatura di 4°C. Il sovranatante ottenuto (S1) è stato nuovamente centrifugato a 10000 x g per 10 minuti a 4°C.
Il precipitato ottenuto da quest'ultima centrifugazione (P2) è stato risospeso in rapporto 1:20 con il tampone T50 e ricentrifugato a 10000 x g per 10 minuti a 4°C. Dalla centrifugazione si è ottenuto un pellet (P3) contenente le membrane mitocondriali che sono state conservate a -20°C fino al momento del loro utilizzo.
18 3.2.1 Determinazione della concentrazione proteica
Per determinare il contenuto proteico, presente in un campione biologico, è stato effettuato un dosaggio proteico che prevede l'uso del radioattivo commerciale Bio-rad protein Assay (metodo colorimetro di Bradford).
Tale dosaggio utilizza il colorante Comassie Brilliant Blue G-250, che cambia colore in seguito al legame con i residui amminoacidici di cui principalmente aromatici o basici (es. arginina).
Il dosaggio è stato eseguito aggiungendo alle aliquote delle membrane 200 µl del reattivo e il quantitativo di acqua distillata necessario a raggiungere una colorazione tendente al blu, la cui intensità è direttamente proporzionale alla concentrazione delle proteine.
Tampone Membrane Acqua MQ Reattivo
(biorad) B B’ 20 µl ↓ ____ 780 µl ↓ 200µl ↓ M M’ ____ 20 µl ↓ 780 µl ↓ 200 µl ↓ Tab. 2 Preparazione saggio proteico
È stata quindi misurata l'assorbanza con uno spettrofotometro alla lunghezza d'onda di 595 nm. Dal valore dell'assorbanza si è ricavata la concentrazione di proteine presenti nel campione facendo riferimento alla retta di taratura ottenuta usando soluzioni di ϒ-globulina a concentrazioni crescenti (2.5 - 20 µg).
19
3.3 REAZIONI DI BINDING DI TIPO REVERSIBILE
Il saggio di radiobinding è una tecnica biochimica che permette di stabilire, con l'ausilio di radioisotopi, la presenza di siti di legame (binding sites) specifici per il ligando utilizzato in cellule o in frazioni subcellulari.
Occorre fare una distinzione tra recettore e binding site: un recettore è un sito di competizione per l'agonista e l'antagonista. Lo stimolo prodotto dal legame dell’agonista al recettore, porta ad una risposta fisiologica. Al contrario un binding site è solo un sito accettore a cui non necessariamente è connessa la trasmissione di un segnale e quindi una risposta cellulare. Il binding site può essere relazionato al recettore di legame solo quando sono soddisfatte le correlazioni, da un punto di vista sperimentale, tra affinità del ligando in vitro e potenza farmacologica in vivo, specificità tissutale, stereospecificità, saturabilità, reversibilità ed alta affinità.
La tecnica di binding radioattivo recettoriale prevede l'incubazione dell'omogenato di tessuto opportunamente diluito, contenente il recettore in esame con un ligando specifico per tale recettore marcato con un isotopo radioattivo. L'incubazione deve essere eseguita temperatura, tempo e pH sufficienti al raggiungimento dell'equilibrio di legame. Durante l'incubazione una certa quantità di ligando (L) lega il recettore (R) e forma il complesso ligando (RL). La formazione del complesso recettore-ligando avviene grazie ad una reazione reversibile che segue la legge d'azione di massa:
K1
[ L ] + [ R ] [ RL ] K2
K1 x [ L ] x [ R ] = velocità di associazione iniziale
20 All'equilibrio le due velocità assumono lo stesso valore, per cui :
K1 x [ L ] x [ R ] = K2 x [ RL ]
e quindi
[ RL ] / [ L ] x [ R ] = K1 / K2
da cui si ottiene la costante di associazione: Ka = K1 / K2
Per determinare la quantità di complesso recettore-ligando formatasi all'equilibrio è necessario terminare l'incubazione con una procedura che permetta di separare il ligando legato al recettore da quello ancora libero e presente nel mezzo di incubazione. Il metodo utilizzato per terminare la reazione di binding è quello della filtrazione che consiste nel filtrare il mezzo di incubazione, contenente quindi sia il complesso recettore-ligando che il ligando libero, sottovuoto attraverso filtri in fibra di vetro, su cui si deposita il tessuto con il ligando legato al recettore. Prima della filtrazione si dispensa velocemente il tampone freddo nel mezzo di incubazione per poter bloccare la reazione e impedire la dissociazione del complesso recettore-ligando, dopo la filtrazione si effettua un ulteriore lavaggio per allontanare dai filtri la radioattività non legata.
Dalla misurazione della radioattività si ricava quindi il legame specifico (Ls) del radioligando interessato, mentre il legame aspecifico (La) corrisponde alla quantità di radioligando che si lega alle strutture non specifiche recettoriali lo si determina incubando le membrane con il radioligando in presenza di concentrazioni saturanti di un ligando specifico e selettivo per lo stesso sito recettoriale. Questo ligando è detto "spiazzante" poiché compete con il radioligando per lo stesso sito recettoriale spiazzandolo. Il legame aspecifico deve essere il più basso possibile (20-30% del totale).
21 membrane con il solo radioligando.
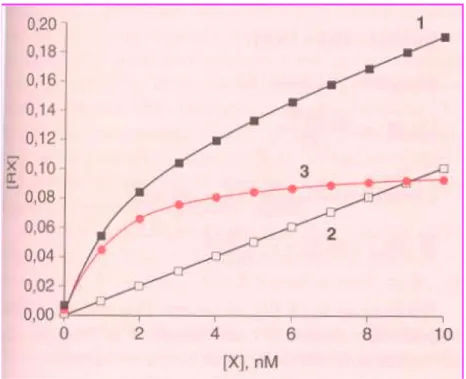
Il binding specifico è quindi calcolato come la differenza tra il legame totale e quello specifico. Quando la radioattività rappresenta il legame ad un recettore (R) saturabile con una singola affinità per il ligando (L) si ottiene come grafico un'iperbole rettangolare, riportando nelle ascisse la concentrazione del radioligando libero [L], espressa in nanomolarità, e nelle ordinate il complesso [ LR ], espressa in fentomoli o picomoli per mg di proteine.
Fig. 10 Grafico del legame specifico (curva rossa), del legame non specifico (retta quadrati vuoti) e del totale (curva quadrati neri)
La fig. 10 mostra che il legame specifico, essendo un fenomeno saturabile, aumenta linearmente in funzione della concentrazione del ligando fino a giungere ad un plateau (steady-state). Ciò è dovuto al fatto che il legame aspecifico è dato dall'interazione del ligando con siti non recettoriali, o più semplicemente a radioattività assorbita dal filtro. La misurazione della radioattività residua viene effettuata mediante uno scintillatore in fase liquida, che fornisce le disintegrazioni per minuto (dpm). Dal valore dei dpm è possibile risalire alla radioattività (µCi) presente nei campioni attraverso la seguente
22 relazione:
1µCi = 2.220.000 dpm
Infine conoscendo l'attività specifica del radioligando (A.S. = µCi/ nmoli) è possibile determinare le nanomoli di radioligando presenti come complesso radioligando-recettore all'equilibrio (BOUND = pmoli / mg proteine).
Attraverso questa tecnica, affiancata dal concetto di competizione è stato possibile determinare i parametri di binding dei composti presi in esame all'equilibrio.
In questo caso l'omogenato tissutale, rappresentato da membrane di rene di ratto, è stato incubato con una concentrazione costante di radioligando, che in questo caso è rappresentato dal [3H] Ro 5-4864, in presenza di concentrazioni crescenti del ligando competitore non marcato, rappresentato da uno dei composti in esame. I valori di legame del radioligando sono stati quindi riportati come percentuale del legame specifico, in funzione del logaritmo della concentrazione dell'agente competitore.
Per poter quantizzare la potenza del ligando nel competere per il sito recettoriale, dalla curva di competizione è stato possibile determinare la concentrazione di competitore in grado di inibire per il 50% il legame specifico del radioligando (IC50). L'IC50 determinata negli studi di binding viene trasformata in costante di inibizione (Ki) del competitore, mediante l'equazione di Cheng e Prussof:
Ki = IC50 / ( 1 + [ L ] / Kd )
dove: Ki = costante di inibizione del competitore Kd = costante di dissociazione del radioligand L = concentrazione del radioligando.
23
3.4 REAZIONI DI BINDING DI TIPO IRREVERSIBILE
Per la caratterizzazione di ligandi capaci di instaurare un legame irreversibile con il TSPO è statostudiato il carattere di irreversibilità nel tempo e l’andamento del legame dei nuovi composti al TSPO.
La metodica seguita è essenzialmente quella descritta da Martini C. e Lucacchini A. (1987). Brevemente le membrane di rene di ratto sono state preincubate con ciascun composto (in un range di concentrazione tra 1 nM e 1µM) a differenti tempi ( 0, 10, 30, 60, 90 e 180 minuti) in tampone Tris-HCl buffer pH=7 contenente 2 mM di MgCl2 e
inibitori delle proteasi a 4°C.
La preincubazione è stata quindi interrotta, ai tempi stabiliti, con l'aggiunta di 1 ml di Tris-HCl freddo pH≅7.4. Tale aggiunta,provocando una diluizione delle concentrazioni del ligando è capace di influenzare un legame reversibile perturbando l’equilibrio, ma non influenza un legame irreversibile. Successivamente i campioni sono stati centrifugati per 15 min a 13000 x g alla temperatura di 4°C.
I pellets ottenuti sono stati lavati due volte con tampone freddo. In seguito è stato effettuato un dosaggio proteico per ogni campione al fine di determinarne la concentrazione proteica.
Dopo la preincubazione ed il lavaggio dei pellets, da ognuno di essi sono stati prelevati circa 65 µg di proteine e incubati con 1nM [3H] Ro 5-4864 in tampone in un volume finale di 500 µl per 60 min a 0°C.
Infine è stata effettuata la filtrazione e la lettura della radioattività come descritte precedentemente.
24
3.4.1 Calcolo delle costanti K
2e K
obsIn accordo con Kitz e Wilson una reazione di binding di tipo irreversibile segue tale relazione: Kd K2 L + R RL
→
L*R dove: L: ligando irreversibile R: recettoreLR: complesso recettore-ligando reversibile L*R: complesso irreversibile
Kd: costante di dissociazione del complesso reversibile
K2: costante di velocità di formazione del complesso irreversibile
Se l’eccesso del ligando irreversibile viene rimosso o ridotto prima del saggio e [ L ] >> [ R ] allora:
1/Kobs = Kd/K2 x 1/[ L ] + 1/K2
(Harmut W. M. Lueddens et al., 1986).
3.5 LINEA CELLULARE U87MG DI GLIOBLASTOMA
UMANO
I composti sono stati anche testati sulla linea cellulare U87MG, derivante da glioma maligno umano di III grado con morfologia epiteliale. Questa linea cellulare è stata ottenuta dall'Istituto Nazionale per la Ricerca del Cancro (ICLC) di Genova.
25
3.5.1 Condizioni di coltura cellulare
Le cellule sono state piastrate e mantenute nell'appropriato mezzo completo sterile costituito da:
• RPMI-1640 addizionato di 10% di siero fetale bovino (FBS); • L-glutammina 2 mM;
• 100 U/ml di penicillina e 100 mg/ml di streptomicina; • 1% di soluzione di aminoacidi non essenziali (NEAA).
La coltura è stata mantenuta nell'incubatore ad una temperatura di 37°C e un'atmosfera umidificata costituita da 5% di CO2 e 95% di O2. Le cellule sono state staccate ogni 4
giorni e ripiastrate con una diluizione 1:10 ad ogni passaggio.
3.5.2 Procedura di congelamento
Per il congelamento di questa linea cellulare è stata preparata un'appropriata soluzione sterile costituita da:
• 50% di RPMI-1640
• 20% di siero fetale bovino ( FBS ) • 10% di dimetilsolfossido ( DMSO )
Per ottenere tale composizione sono state preparate due distinte soluzioni: ▪ Soluzione A: RPMI-1640 (6 ml) + FBS (4 ml)
▪ Soluzione B: RPMI-1640 (8 ml) + DMSO (2 ml)
Le cellule presenti nella fiasca da 75 cm2 e sono state centrifugate a 100 x g per 5 minuti. Il pellet ottenuto è stato sospeso prima in 2 ml della soluzione A e dopo si sono aggiunti molto lentamente 2 ml di soluzione B.
Le U87MG sospese sono state poi contate utilizzando una camera di Burker.
26 una quantità di cellule tra 5x105 e 1x106. Le criovials sono state mantenute per 24 ore ad una temperatura di - 80°C e poi conservate in azoto liquido ( - 196°C )
3.6.3 Procedura di scongelamento delle U87MG
Le cellule, conservate in azoto liquido in criovials, sono state velocemente scaldate nel bagnetto a 37°C e raccolte. In seguito, è stato aggiunto molto lentamente il mezzo completo sterile, caldo, fino ad avere un volume appropriato. Il tutto è stato poi centrifugato a 100 x g per 5 minuti. Il pellet ottenuto è stato poi risospeso in un volume adeguato di mezzo completo riscaldato. La sospensione ottenute è stata deposta poi in una fiasca da 75 cm2, contenente mezzo completo, in modo da ottenere 10 ml totali
.
3.6.4 Tecnica di splitting
Questa tecnica è l’operazione che consente il distacco delle cellule eucariotiche aderenti ad un substrato solido. Lo splitting avviene mediante l'utilizzo di una soluzione contenente l'enzima tripsina addizionata di EDTA. La tripsina è una proteasi che agisce demolendo le proteine delle giunzioni che creano delle interconnessioni tra le cellule. L’EDTA che ad alte concentrazioni va a chelare il Ca2+, indispensabile per l'adesione cellulare.
Il procedimento sperimentale prevede l'uso del mezzo di coltura, della soluzione di Tripsina-EDTA e di soluzione fisiologica, che devono essere precedentemente scaldate a 37°C.
Il mezzo di coltura dalla fiasca è stato aspirato mediante l'uso di una Pasteur di vetro collegata ad una pompa a vuoto, poi è stato effettuato un lavaggio con la soluzione fisiologica, che in seguito è stata aspirata; successivamente si è aggiunto un volume idoneo di soluzione contenente tripsina e si è lasciato in incubazione a 37°C per qualche
27 minuto.
Dopo qualche minuto si sono introdotti circa 3-4 ml di mezzo di coltura, si è prelevato quindi la sospensione lavando bene le superfici della fiasca con il mezzo e si è trasferito il tutto in una falcon da 15 ml.
La sospensione cellulare è stata poi centrifugata a 1000 rpm per 5 minuti a 18°C. Dopo aspirazione del sovranatante e il pellet ottenuto è stato risospeso in un volume idoneo di mezzo di coltura mediante omogenizzazione accurata.
3.6.5 Conta delle cellule
Al fine di contare il numero di cellule si è fatto uso della camera di Burker. Tale camera consta di una lastra di vetro rettangolare, che presenta al centro un rilievo, delimitato ad ambo i lati da due solchi o scanalature parallele ed un corto solco trasversale che divide questo rilievo centrale in due metà, ciascuno delle quali porta inciso un reticolo di conta. I due reticoli vengono coperti da un unico vetrino coprioggetto tenuto aderente da due morsetti laterali metallici.
La conta è stata effettuata prelevando 10 µl di sospensione cellulare, derivante da un pellet di cellule diluito in un opportuno volume di mezzo. La quantità prelevata poi è stata posta tra i due vetrini e con l'uso del microscopio, si è operata la conta. Ogni reticolo di conta che costituisce la Camera di Burker, è diviso in 9 griglie e ognuna di queste è formata da 16 quadrati (Fig.11).
Fig. 11 Schema della camera di Burker • Cellule che vengono contate
28 Le cellule contenute in 3 o 4 delle 9 griglie sono state contate per ottenere un valore medio.. Ciascun quadrato ha un volume pari a 0.1mm3 10-4 cm3 (1 cm 3 equivale a circa 1 ml), quindi:
numero di cellule / ml = conta media del quadrato per 104 (fattore di Burker).
Il numero totale di cellule contenute in un certo volume si determina moltiplicando il numero di cellule / ml ottenuto per i ml della sospensione iniziale.
3.6 STUDI IN VITRO DEL LIGANDO 18
Con questi studi è stata valutata la capacità del composto 18 di legarsi al TSPO in cellule di glioma umano, misurata con l’ausilio della spettroscopia a fluorescenza. Le cellule U87MG sono state seminate in pozzetti di multi-well da 96 a fondo nero per una quantità di circa 4 x 104 cellule/well sospese nel mezzo RPMI 1640 completo. Una
volta effettuata la semina, le cellule sono state fatte aderire ai pozzetti per 24 ore a 37°C. Successivamente le cellule sono state incubate con differenti concentrazioni del composto 18 (100, 250 e 500 nM) in mezzo di coltura per 20 e 90 minuti, al 5% di CO2
e 37°C. In parallelo alcuni campioni sono stati incubati con Ro 5-4864 (10 µM) prima dell’aggiunta del composto 18.
Dopo l’incubazione sono stati effettuati 4 lavaggi con cloruro di cisteina 3g/l precedentemente preparata per rimuovere l’eccesso di composto non legato, in quanto la cisteina è in grado di reagire immediatamente con il gruppo isotiocianato.
Al termine del quarto lavaggio sono stati aggiunti 100µl di PBS 1x in ogni well e si è effettua la lettura della fluorescenza al lettore di piastre Victor Wallac 2 ( filtro di eccitazione pari a 485nm e filtro di emissione pari a 535nm 0.1 sec.). (Fig.12)
29 Fig. 12 Victor Wallac 2
30
4. RISULTATI E DISCUSSIONE
Nel presente lavoro di tesi sono stati caratterizzati biologicamente alcuni composti di nuova sintesi appartenenti alla classe delle 2-fenilindol-3-ilgliossilamidi ed è stata valutata sia la loro capacità di legare il TSPO che la loro attitudine a poter essere impiegati nello studio su cellula.Gli studi biologici dei nuovi ligandi hanno avuto come primo step la valutazione dei parametri di binding all’equilibrio, seguiti da studi sulla potenziale irreversibilità del loro legame alla proteina recettoriale. Il ligando irreversibile fluorescente inoltre è stato saggiato in esperimenti sulla linea cellulare U87MG di glioma umano attraverso la spettroscopia fluorescente.
Nel corso degli anni sono stati progettati numerosi radioligandi per il TSPO in grado di misurare il livello di espressione di questa proteina sia negli animali che nell’uomo. Tuttavia nella ricerca biomedica, i ligandi fluorescenti rappresentano un’alternativa più sicura, efficace e meno costosa dei radioligandi.
A tale scopo sono stati recentemente studiati nuovi ligandi fluorescenti derivati N,N-dialchil-2-fenilindol-3-ilgliossilamidi. Il gruppo responsabile della fluorescenza che è stato adottato come fluoro cromo è il 7-nitrobenz-2-ossa-1,3-diazol-4-il (NBD). I valori di massima eccitazione ed emissione per l’NBD sono rispettivamente 470 e 545 nm. Numerosi studi riportano l’NBD come un buon marcatore per la valutazione del sito di legame e per lo studio delle interazioni ligando-recettore.
I derivati indolgliossilamidi N,N di sostituiti fluorescenti hanno un’affinità intorno al nanomolare per il TSPO poiché vanno ad interagire con i siti L1, L2 e L4 del recettore e
31 fenilico in posizione 2 può essere coinvolto nell’interazione π-stacking con il sito L1 del
TSPO grazie all’anello aromatico ricco di elettroni; questa interazione può essere rafforzata dalla presenza in posizione 4’ (R3) di un gruppo elettronattrattore (Taliani et
al., 2007).
Nonostante la buona affinità di legame di questi composti,esperimenti condotti su tessuto hanno evidenziato che i lavaggi previsti nelle tecniche impiegate per la visualizzazione, spesso causano un’alterazione dell’equilibrio chimico con conseguente distacco del ligando fluorescente con dal TSPO. Per non andare incontro a questo fenomeno sono stati sintetizzati, presso il laboratorio diretto dal Prof. Da Settimo, nuovi ligandi in grado di legare covalentemente il recettore. Utile gruppo in grado di stabilire un legame irreversibile con il TSPO risulta essere l’isotiocianato altamente reattivo nei confronti dei gruppi amminici e sulfidrilici, mentre poco reattivo nei confronti dell’acqua e delle funzioni idrossiliche. Questo gruppo è capace di alchilare l’istidina, amminoacido presente nel dominio del TSPO, reagendo con il suo gruppo imidazolico e dando un’interazione irreversibile.
In questo lavoro di tesi sono stati quindi saggiati dei nuovi ligandi al TSPO N,N-dialchil-2-fenilindol-3-ilgliossilamidi derivati irreversibili (composti 7 e 8) e irreversibili fluorescenti (composto 18).
4.1 RISULTATI DEI RADIOBINDING
Per lo studio biologico dei nuovi ligandi irreversibili 7 8 e 18 ci siamo avvalsi della tecnica di binding all’equilibrio, seguita da studi sulla potenziale irreversibilità di questi ligandi al recettore.
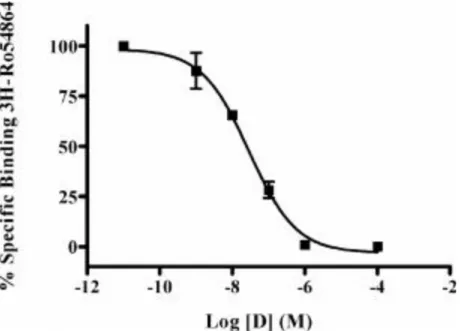
La tecnica di binding all’equilibrio, eseguita su membrane di rene di ratto, si basa sulla competizione dei ligandi in esame nei confronti del [3H] Ro 5-4864.
32 Di seguito sono riportate le curve di spiazzamento del [3H] Ro 5-4864 da parte dei composti in esame (Fig. 14, 15 e 16).
Fig. 14 Grafico di spiazzamento del [3H] Ro 5-4864 da parte del composto 7 in
membrane mitocondriali di rene di ratto.
Fig. 15 Grafico di spiazzamento del [3H] Ro 5-4864 da parte del composto 8 in membrane mitocondriali di rene di ratto.
33 Fig. 16 Grafico di spiazzamento del [3H] Ro 5-4864 da parte del composto 18 in
membrane mitocondriali di rene di ratto.
I dati relativi al binding all’equilibrio dei composti 7,8 e 18 mostrano valori di affinità nel range del nanomolare, comparabili quindi ai valori dello standard Ro 5-4864 (Ki da 37.6 nM a 49.5 nM). (Tab. 3)
Tab. 3 Valori di IC50 e Ki dei composti 7,8 e 18 ottenuti dai saggi di binding all’equilibrio attraverso l’equazione di Cheng-Prusoff.
34 Per esaminare la potenziale irreversibilità del legame dei composti 7,8 e 18 sul TSPO è stato eseguito un binding in cinetica con particolari accorgimenti. I campioni sono stati diluiti 1:10 con tampone freddo al termine della loro incubazione con il radioligando e si è osservato che i dati ottenuti dopo tali diluizione non erano differenti da quelli ottenuti nei campioni non diluiti. Ciò indicava che l’interazione TSPO-ligando era irreversibile e questo escludeva immediatamente la possibilità di applicare l’equazione di Cheng-Prusoff nell’analisi dei parametri di binding , perchè valida solo nel caso di ligandi reversibili.
I risultati ottenuti hanno dimostrato che, pretrattando le membrane mitocondriali con i nuovi ligandi per il TSPO, questi erano in grado di inibire il legame del [3H] Ro 5-4864 con il recettore. Questa inibizione era di tipo tempo- e concentrazione- dipendente come si può osservare nelle cinetiche di legame relative ai composti 7,8 e 18 (Fig. 17, 18 e 19).
Fig. 17 Comportamento nel tempo del binding del composto 7 al TSPO. Rappresentazione semilogaritmica della diminuzione del binding del [3H] Ro 5-4864 al TSPO con pretrattamento del composto 7 (0, 10, 30, 60, 90 e 180
min.) a varie concentrazioni. I punti rappresentano il valore medio ±SEM derivanti da tre esperimenti indipendenti eseguiti in doppio.
35 Fig. 18 Comportamento nel tempo del binding del composto 8 al TSPO.
Rappresentazione semilogaritmica della diminuzione del binding del [3H] Ro 5-4864 al TSPO con pretrattamento del composto 8 (0, 10, 30, 60, 90 e 180
min.) a varie concentrazioni. I punti rappresentano il valore medio ± SEM derivanti da tre esperimenti indipendenti eseguiti in doppio.
Fig. 19 Comportamento nel tempo del binding del composto 18 al TSPO. Rappresentazione semilogaritmica della diminuzione del binding del
[3H] Ro 5-4864 al TSPO con pretrattamento del composto 18 (0, 10, 30, 60, 90 e 180
min.) a varie concentrazioni. I punti rappresentano il valore medio ± SEM derivanti da tre esperimenti indipendenti eseguiti in doppio
36 Nei grafici sopra riportati B indica il binding dopo pretrattamento delle membrane a diverse concentrazioni dei ligandi ai diversi tempi (0, 10, 30, 60, 90 e 180 min) mentre B0 rappresenta il caso in cui la concentrazione di ligando è pari a 0.
Riportando le grandezze di B e B0 in scala logaritmica in funzione del tempo si può
osservare come la diminuzione del valore ln B/B0 sia lineare e quindi costante nel tempo
(fig. 19, 20 e 21). Secondo queste osservazioni, si può parlare quindi di reazione di cinetica di primo ordine definita dalla seguente relazione:
ln B/B0 = - Kobs x t
dove Kobs è la costante di velocità apparente per le reazioni di primo ordine,
caratterizzanti il binding irreversibile. Matematicamente Kobs è definita dalla pendenza
delle diverse rette presenti nelle fig. 20-22.
Questa relazione matematica permette di calcolare la costante di dissociazione (Kd) del complesso reversibile recettore-ligando e la pseudocostante di velocità della reazione di primo ordine (K2) implicata nella formazione dell’interazione irreversibile tra il
recettore ed il ligando secondo la seguente equazione:
1/Kobs = Kd/K2 x 1/[ D ] + 1/K2
Il grafico in cui sono riportati i valori di 1/Kobs in relazione ai valori di 1/D (dove D è la
concentrazione del ligando) mostra una retta la cui intercetta sull’asse delle ordinate rappresenta il valore di 1/K2, mentre l’intercetta sull’asse delle ascisse dà il valore di
37 Fig. 20 Relazione matematica per calcolare i valori di K2 e Kd del ligando 7
38 Fig. 22 Relazione matematica per calcolare i valori di K2 e Kd del ligando 18
Nella seguente tabella sono riportati i valori della K2 e della Kd dei composti 7,8 e 18;
come si può notare i valori delle Kd sono all’interno del range del nanomolare.
COMPOUND K2 (nM) Kd (nM)
7 0.003 ± 0.001 3.17 ± 0.31
8 0.015 ± 0.002 5.07 ± 0.61
18 0.0012 ± 0.002 8.31 ± 0.89
Tab. 4 Valori di K2 e Kd dei composti 7,8 e 18
Si è potuto quindi concludere che l’interazione dei ligandi testati con il TSPO è di tipo irreversibile e segue tale relazione:
Kd K2
39
4.2 STUDI IN VITRO DEL COMPOSTO 18
La capacità del composto 18 di legare specificamente ed irreversibilmente il TSPO è stata valutata sulla linea cellulare U87MG di glioma umano usando la spettroscopia fluorescente.
Per valutare la capacità del composto 18 di permeare attraverso la membrana cellulare sono state anche studiate le sue proprietà chimico-fisiche (tab. 5) dimostrando la sua buona capacità di permeare.
Tab.5 Proprietà chimico-fisiche del composto 18
a coefficiente di ripartizione n-ottanolo acqua. b Totale superficie del solvente. c
Superficie polare. d Numero di atomico. e Numero di legami ruotabili. f Peso molecolare.
Per la valutazione in vitro, le cellule sono state incubate a differenti concentrazioni del composto 18 e a due tempi diversi (20 e 90 minuti, tempo necessario per la formazione del legame calcolato sostituendo le costanti K2 e Kd nelle equazioni precedentemente
descritte). Dopo 20 minuti, i valori d’intensità di fluorescenza ottenuti erano più bassi dei valori ottenuti dopo 90 minuti, come mostrato anche dai grafici riportati in figura 23.
40 Fig. 23 Cellule di glioma umano trattate con il ligando irreversibile fluorescente 18 per il TSPO. Le cellule sono state incubate con il composto 18 a due diversi tempi,
20 minuti (A) e 90 minuti (B) in assenza o presenza di Ro 5-4684.
È interessante notare che dopo 20 minuti di incubazione, il Ro 5-4864 è stato capace di spiazzare il composto 18 con la stessa entità per tutte le concentrazioni saggiate, dimostrando così la specificità del composto 18 per il TSPO. Viceversa negli esperimenti eseguiti in tempi più lunghi, il Ro5-4864 si è dimostrato incapace di spiazzare il composto 18, mostrando così l’irreversibilità del legame con il recettore.
41
4.3 CONCLUSIONI
In questo lavoro di tesi è stato possibile definire l’ordine di reazione e le costanti cinetiche dell’interazione tra i ligandi saggiati ed il TSPO. I dati sperimentali ottenuti dimostrano che tutti i ligandi presi in esame sono caratterizzati da un’interazione irreversibile con il recettore.
Inoltre dati ottenuti in cellule U87MG su cui sono stati condotti esperimenti di spettroscopia fluorescente eseguiti per il composto 18 rivelano la sua alta affinità, specificità e capacità di dare legami covalenti con il TSPO. I nuovi ligandi irreversibili al TSPO possono rappresentare mezzi utili sia per lo studio molecolare del recettore stesso che per la sua caratterizzazione e misurazione dei suoi livelli di espressione in ambito diagnostico.
In particolare il composto fluorescente 18 potrebbe essere usato come marcatore per valutare l’espressione del TSPO in biopsie di pazienti affetti da patologie dove si abbia un’alterata densità del TSPO.
42
BIBLIOGRAFIA
Atkins P., De Paula J., Elementi di Chimica fisica, Terza edizione italiana,
Bologna, Zanichelli. 2007.
Buck J. R., McKinley E. T., Hight M. R., Fu A., Tang D., Smith R. A., Tantawy M.
N., Peterson T. E., Colvin D., Ansari M.S., Baldwin R. M., Zhao P., Guleryuz S., and Mannin H. C. Quantitative, preclinical PET imaging of TSPO expression in glioma using [18F]PBR06. J Nucl Med., 2011, 52(1): 107-114.
Casellas P., Galiegue S., Basile A.S. Peripheral Benzodiazepine Receptorsand
Mitochondrial Function. Neurochemestry International, 2002, 40 (6): 475-486.
Denora N., Laquintana V., Trapani A., Suzuki H., Sawada M., Trapani G.. New
Fluorescence Probes Targeting the Mitochondrial Located Translocator Protein 18 KDa (TSPO) as Activated Microglia Imaging Agents. Pharm Res. ,2011, 28:2820- 2832.
Gavish M., Bachman I., Shoukrun R., Katz Y., Veenman L., Weisinger G., Weizman.
Enigma of the Peripheral Benzodiazepine Receptor. Pharmacological Reviews , 1999, 51(4): 630-646
43 Harmut W.M. Lueddens, Amy Hauck Newman, Kenner C. Rice, Phil Skolnick. AHN
086: An irreversible Ligand of “Peripheral” Benzodiazepine Receptor. Molecular Pharmacology, 1986, 29: 540-545.
Martini C., Lucacchini A. Affinity Labeling of Adenosine A1 Binding Sites.
Neurochemestry, 1987, 49 (3): 681-684.
Moore W. J., Chimica Fisica, Padova, Piccin-Nuova Libreria. 1990.
Papadopoulos V., Lecanu L., Brown R.C., Han Z., Yao Z.X. Peripheral-Type
Benzodiazepine Receptor in Neurosteroid Biosynthesis, Neuropathology and Neurological Disorders. Neuroscience, 2006, 138 (3): 749-756.
Sega E. I., Low P. S., Tumor detection using folate receptor-targeted imaging
agents. Cancer Metastasis Rev, 2008, 27:
Taliani S., Simorini F., Sergianni V., La Motta C., Da Settimo F., Cosimelli B.,
Abignente E., Greco G., Novellino E., Rossi L., Gremigni V., Spinetti F., Chelli B., Martini C. New Fluorescent 2-Phenylindolglyoxylamide Derivates as Probes Targeting the Peripheral-Type Benzodiazepine Receptor: Design, Synthesis and Biological Evaluation. J. Med. Chem. 2007, 50: 404-407.
![Fig. 15 Grafico di spiazzamento del [ 3 H] Ro 5-4864 da parte del composto 8 in membrane mitocondriali di rene di ratto](https://thumb-eu.123doks.com/thumbv2/123dokorg/7535258.107511/32.892.177.788.674.1069/fig-grafico-spiazzamento-parte-composto-membrane-mitocondriali-ratto.webp)