Capitolo 3
In letteratura sono riportati svariati lavori che mirano a valutare le caratteristiche elettriche dei costituenti della cellula. Questo problema è stato affrontato tramite metodi diretti di simulazione molecolare per i compartimenti di struttura minore [22], come i canali proteici di membrana, o indiretti per quelli maggiori, come, ad esempio, la membrana cellulare [23, 24].
In particolare, il procedimento seguito con i metodi indiretti consiste nella misura, tramite tecniche di spettroscopia [23], del comportamento dielettrico delle cellule con concentrazione, forma e dimensioni note; inoltre questi metodi sono applicati a differenti modelli dielettrici al fine di ottenere una stima della costante dielettrica degli elementi che formano la sospensione cellulare.
In seguito, gli studi hanno posto l’attenzione su adeguate metodologie per la valutazione del campo elettromagnetico indotto nelle strutture biologiche esposte. Sono state proposte sia soluzioni analitiche [25] che tecniche numeriche [26]-[28]. Queste ultime consentono una accurata predizione della distribuzione di campo elettromagnetico indotto qualora sia disponibile un preciso modello numerico della sorgente. Nella pratica però, non è sempre facile fornire un modello per la sorgente, sia perché questa non è nota con precisione, sia perché la sua posizione non è facilmente deducibile.
Nei paragrafi successivi saranno esposti gli studi eseguiti per il calcolo del TMP e della frequenza di rilassamento tramite metodi numerici effettuati da parte di Stuchly e Fear [29].
Capitolo 3
3.1. Modelli e metodo di analisi
Stuchly e Fear hanno analizzato, nel loro lavoro [29], la tensione di transmembrana indotta (TMP) in una configurazione cellulare particolare: i raggruppamenti di cellule elementari connessi tra loro tramite gaps, ossia i cluster.
Il modello di ogni singola cellula è quello a strati, visto nel capitolo 1 (figura 1.1): il citoplasma (σcito =0.5S/m, εr =80) circondato da una sottile membrana, poco conduttiva, ( =10−7 S/m
mem
σ , εr =11.3) è immerso in un mezzo conduttivo esterno (σext =1S/m, εr =80). Il diametro cellulare, in genere variabile tra 1 e 200 µm, è stato fissato a 1µm, mentre lo spessore della membrana, che tipicamente è di circa 10 nm, per semplificare la complessità della struttura, è stato preso di 30 nm. Tale approssimazione è accettabile visto che lo spessore della membrana non influisce significativamente sul calcolo del TMP.
I gaps, come tutti gli elementi della cellula, sono caratterizzati a livello dielettrico: la conducibilità varia da 10-7 a 1 S/m ed, essendo modellati come canali riempiti di citoplasma, ne hanno la stessa permittività (εr =80). La dimensione massima del canale dei gaps è di 1.5-2 nm, perciò la lunghezza dei gaps stessi, che varia dai 20 ai 40 nm, è ragionevolmente assimilabile ad un cluster di gaps.
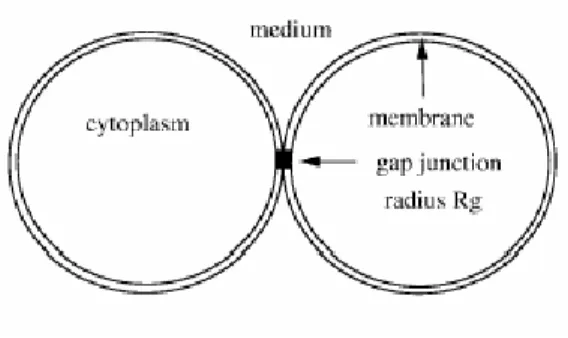
Figura 3.1 Modello sferico.
La figura 3.1 mostra un modello di concatenamento cellulare di tipo sferico, mentre la figura 3.2 quello di tipo cilindrico. Il modello sferico non rappresenta una
situazione reale, per cui quello che viene analizzato è il modello cilindrico che risulta, nella pratica, più simile alla struttura delle cellule biologiche.
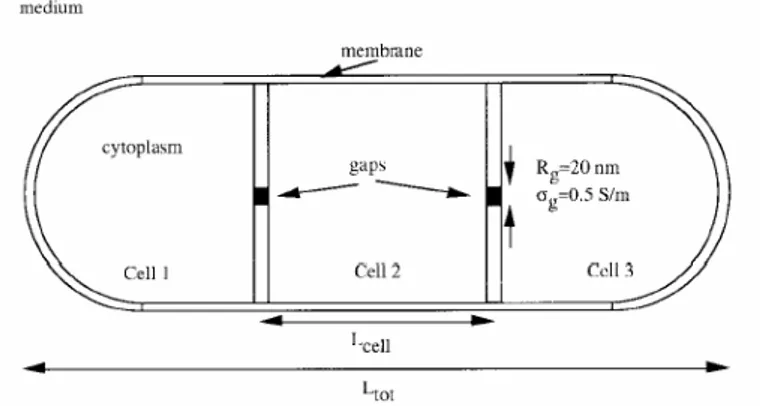
Figura 3.2 Modello cilindrico.
Quest’ultima struttura è il punto di partenza delle simulazioni di Fear e Stuchly. In tabella 3.1 sono riassunti i vari modelli di concatenamento cellulare simulati, dove la lunghezza della cellula è presa pari a Lcell =2R, se non diversamente specificata, e i cluster sono di 2x2x2 cellule (figura 3.3). In figura 3.4 è raffigurato un cluster 2x2x4.
Capitolo 3 Cluster Numero di cellule Raggio µm Lung. Tot. µm
Raggio dei gap (nm) 2 0.5 5 2 20 20 15 10 200 3-15 0.5 3 - 15 20 Da 10 a 78 (L = 10) 0.5 97 - 776 100 8 5 80 20 7 (L=15) 7.5 15 22.5 100 105 20 50 200 400 600 4000 5 7.5 75 300 3 (L=25) 7.5 75 300 10 (L=25) 5 250 20 200 300 9 (L=100) 5 900 20 100 Tabella 3.1
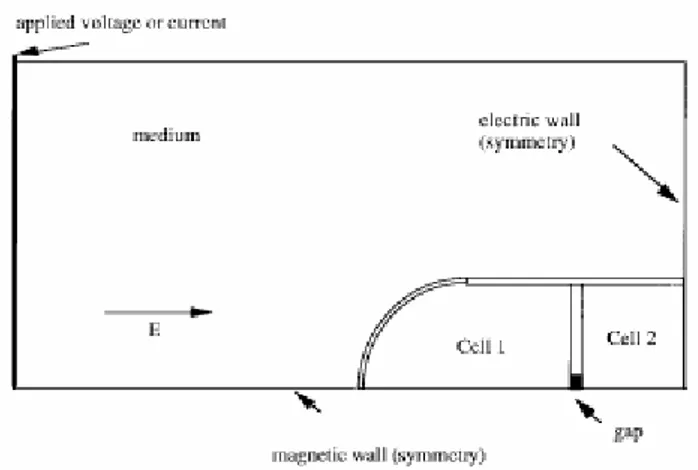
Per ridurre le dimensioni del problema, se ne crea una versione simmetrica rispetto agli assi delle cellule: un esempio di concatenamento a tre cellule, del tipo in figura 3.2, è mostrato in figura 3.5. Per sfruttare la simmetria del sistema, vengono posti sul bordo della struttura troncata, come condizioni al contorno, due schermi, uno magnetico ed uno elettrico. Tali condizioni vengono posizionate ad una distanza di 2.25 celle dal centro della struttura vera e propria.
Le cellule sono sottoposte ad un campo elettrico di 1 V/m, ottenuto tramite o una tensione in continua (dc) o una corrente alternata (ac); nel primo caso la tensione applicata lungo il contorno dipende dalle dimensioni spaziali del problema, nel secondo la corrente applicata dipende dalla conducibilità del mezzo.
Figura 3.5 Modello di 3 cellule simmetrico rispetto agli assi
Data la complessità delle strutture in esame, per il calcolo del TMP è stato usato “il metodo agli elementi finiti” (FEM) e nello specifico, per le simulazioni, il pacchetto commerciale EMAS (Ansoft, Pittsburgh, PA).
Questo tipo di approccio prevede una discretizzazione del problema in tante piccole regioni, chiamate elementi, che possono avere varie forme e dimensioni in modo che, combinati tra loro, riescono ad approssimare strutture geometriche molto complesse. In particolare, con elementi estremamente piccoli si riescono a rappresentare determinate aree d’interesse senza creare problemi a livello di risoluzione computazionale.
Il codice EMAS [30], per simulare il progetto reale (vale a dire l’energia elettrica e magnetica immagazzinata, l’energia dissipata, etc.) si serve di un funzionale che è espresso in termini di potenziale vettore magnetico A e di potenziale elettrico non convenzionale ψ , il quale risulta essere l’integrale nel tempo del convenzionale potenziale scalare. Sebbene i potenziali varino in maniera complessa nell’intero dominio, la loro variazione su ogni elemento si può facilmente descrivere tramite le funzioni ‘forma’, le quali vengono usate per esprimere il funzionale, che poi va integrato sia sul tempo che sul volume di ogni singolo elemento.
Capitolo 3 Combinando l’espressione del funzionale e la rappresentazione degli elementi finiti, EMAS crea una matrice per tutti gli elementi del sistema per calcolare il potenziale
A e ψ incogniti.
Il problema è che EMAS fornisce un risultato che tiene conto solo delle proprietà di conduzione dei materiali; questa approssimazione quasi-statica è appropriata per una analisi cellulare alle basse frequenze, ossia fino ad arrivare a poche centinaia di KHz. Per questo motivo sono state usate le solo proprietà statiche del mezzo, del citoplasma e della membrana. Inoltre, per maggiore semplicità, è stato tracurato sia il campo magnetico indotto che la corrente di spostamento.
I modelli e i dati d’uscita elaborati da EMAS sono generati da un software complementare, ARIES [31].
Tramite ARIES si costruisce il modello solido della struttura cellulare e si realizza la mesh, una per ogni regione del modello: ad esempio gli elementi della membrana vengono presi con una lunghezza pari a 5 nm, quelli del citoplasma a 100 nm ed infine quelli del mezzo a 400 nm.
Quindi usando una discretizzazione spaziale con elementi lineari del primo ordine, si è scelta per la membrana e i gaps una risoluzione piu’ fine rispetto a quella, piu’ grossolana, scelta per il mezzo e il citoplasma.
Il numero massimo di elementi dipende dalla memoria del computer usato e dal tempo computazionale. I tempi di calcolo variano molto, da circa un minuto per le singole celle, fino a cinque ore per modelli più grandi con circa 10000 elementi. Il modello più grande che sono riusciti a simulare contiene 48000 elementi, usando un HP 9000/735.
I risultati ottenuti sono validi al di sotto della frequenza di rilassamento, che per una cellula sferica viene espressa in termini di capacità di membrana Cm (in genere pari a
2 2 F/m
10− ) e di raggio cellulare R [3], come:
+ = i e m relax RC f σ σ π 1 2 1 2 1 . (3.1.1)
Calcolando il TMP a diverse frequenze e interpolandone i risultati, si ricava la frequenza alla quale l’ampiezza della tensione di transmembrana risulta essere 3 dB
al di sotto del valore continuo ( frelax). La frequenza di rilassamento della singola cellula è di 12.7 MHz, frequenza alla quale sono state calcolate le sue proprietà dielettriche.
3.2. Risultati e verifiche
Stuchly e Fear hanno dapprima posto la loro attenzione su un modello di cellula bidimensionale per poi passare a quello tridimensionale, cioè simmetrico rispetto agli assi delle cellule stesse ed infine alle catene di cellule.
In relazione alle condizioni al contorno, alla dimensione della membrana e al numero di elementi con cui si è stato discretizzato il dominio, si riportano in tabella 3.2 i valori di TMP calcolati tramite FEM, con il relativo errore rispetto alla soluzione analitica, che per il modello 2D risulta essere di 1µV [3].
Membrana d (nm)
Numero di celle dal centro alle condizioni al contorno
Numero di elementi TMP (µV) Errore (%) 10 1.5 5708 (lineare) 0.915 8.5 30 3 4929 0.964 3.5 30 4.5 5316 0.985 1.5 30 2.25 3488 0.959 4.1 30 2.25 1807 (quadr.) 0.961 3.9 10 2.25 5449 (lineare) 0.957 4.3 Tabella 3.2
Nel caso tridimensionale il TMP è 0.745 µV e differisce dell’1% dal risultato esatto. Questo errore tende ad aumentare con la frequenza, infatti, si arriva a circa il 9 % alla frequenza di 20 MHz. Per quanto riguarda la frequenza di rilassamento, questa è calcolata a 11.8 MHz; rispetto alla soluzione analitica si commette un errore, pari al 7%, dovuto all’interpolazione lineare dei dati.
Capitolo 3
3.2.1. Catene e cellule allungate
Il TMP viene valutato per cellule e catene di cellule (connesse da gaps) di vari diametri ed estensioni, esposte a campi elettrici applicati lungo i loro assi maggiori. Si considera sia il caso di catene lunghe che quello di catene corte, la cui lunghezza dipende dalla lunghezza d’onda λ , calcolata tramite la seguente relazione:
m i e i m Rd r r r σ σ λ 2 ≈ + = , (3.2.1)
dove r è la resistività della membrana, m r la resistività del citoplasma e i r la e resistività del mezzo.
Le catene connesse da gap hanno una lunghezza d’onda λ più corta rispetto alle cellule equivalenti (insieme di più cellule non divise tra loro dalla membrana) a causa dell’incremento della resistività del citoplasma. Per considerare le giunzioni di gap, la resistività del citoplasma viene calcolata come segue:
(
)
(
)
− + − + = 2 2 2 2 , 1 2 1 g m g g g i cell tot gap i R R R d N R L N L r σ π σ π σ π , (3.2.2)in cui Nè il numero delle cellule nella catena, L la lunghezza di una singola cell cellula, L la lunghezza totale della catena ed infine tot R e g σg, rispettivamente, il raggio e la conducibilità del gap.
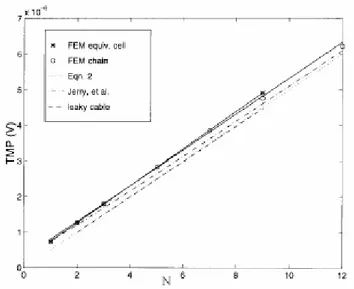
In figura 3.6 si riportano i valori del TMP, ottenuti con il FEM, nel caso di cellule connesse da gap e cellule equivalenti. Questi risultati sono confrontati con quelli raggiunti tramite il modello del cavo con perdite [32] e la soluzione analitica per cellule sferiche [33] e sferoidali. Per quest’ultimo tipo di cellule, si usa l’approssimazione di Pauly e Bernhardt [17], in cui la tensione si ricava con un’equazione che tiene conto sia della lunghezza complessiva del raggruppamento cellulare L che dell’intensità del campo applicato E : tot
tot EL TMP 2 1 = (3.2.3)
Figura 3.6 TMP al variare del numero di cellule connesse tra loro.
Usando, invece, l’approssimazione ai minimi quadrati, il TMP per le catene cellulari si può esprimere in funzione del numero totale N di cellule formanti la catena e della tensione di transmembrana della singola cellula Vm,s:
) 4 . 0 66 . 0 ( , + =V N Vm ms . (3.2.4)
Le equazioni (3.2.3) e (3.2.4) forniscono dei risultati molto simili tra loro, e la differenza si riduce sempre più all’aumentare della dimensione della catena.
È evidente che piccole catene possono modellarsi con semplici cellule equivalenti, poiché la presenza dei gaps è irrilevante: questo comporta un errore trascurabile, ad esempio di circa 1% per una catena di tre cellule e il 3% per una di sette. Con l’approssimazione di Pauly e Bernhardt, il TMP risulta sottostimato di circa il 20% in catene bicellulari e per meno del 10% in catene con sette cellule. Ciò nonostante, questa approssimazione può essere usata per stimare la tensione indotta nelle lunghe catene in quanto l’errore introdotto decresce all’aumentare della lunghezza.
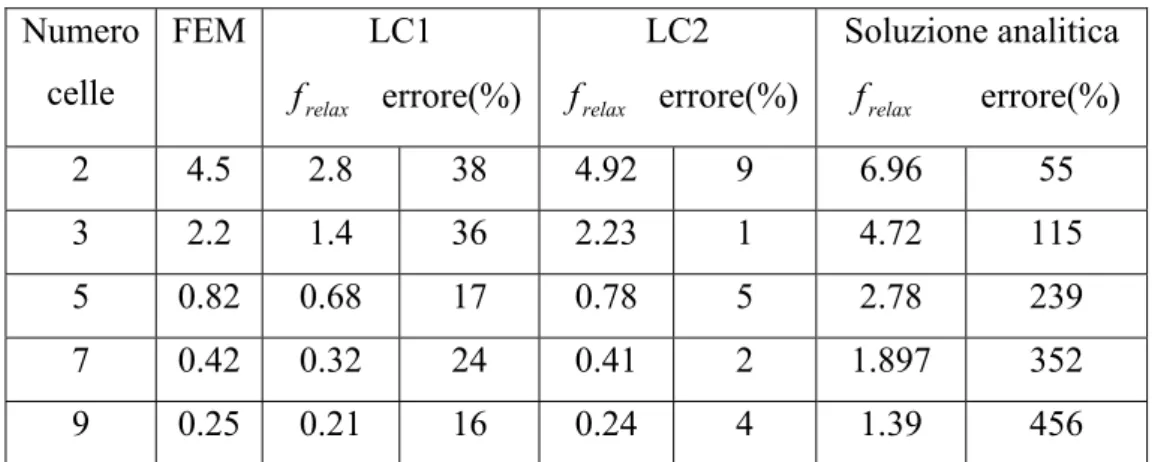
Per queste configurazioni, viene calcolata anche la frequenza di rilassamento, la quale viene rapportata a quella calcolata nel caso di cellule ellissoidali e nel caso del cavo con perdite (tabella 3.3). Per il cavo con perdite si considerano due casi, uno con gli “endcaps” di membrana, ossia una sorta di cappuccio che termina la cellula, LC1, e uno senza, LC2.
Capitolo 3 Numero celle FEM LC1 relax f errore(%) LC2 relax f errore(%) Soluzione analitica relax f errore(%) 2 4.5 2.8 38 4.92 9 6.96 55 3 2.2 1.4 36 2.23 1 4.72 115 5 0.82 0.68 17 0.78 5 2.78 239 7 0.42 0.32 24 0.41 2 1.897 352 9 0.25 0.21 16 0.24 4 1.39 456
Tabella 3.3 Frequenza di rilassamento in MHz per il modello ellissoidale.
LC1 indica il cavo con perdite con gli ‘endcaps’ e LC2 quello senza.
I risultati ottenuti per il modello ellissoidale non sono ragionevoli, poiché questo modello, geometricamente parlando, è abbastanza differente da quello cilindrico e tale differenza cresce sempre più all’aumentare della lunghezza delle cellule.
La tabella, inoltre, indica che il caso LC1 non fornisce accuratamente la frequenza di rilassamento delle cellule equivalenti per la presenza degli ‘endcaps’, che la fanno abbassare. Il modello LC2, al contrario, produce una buona stima della frequenza di rilassamento.
La figura 3.7 mette a confronto la frequenza calcolata per il modello a catena e quella delle cellule allungate, entrambe eccitate da un campo applicato in parallelo all’asse più lungo della cellula. Si nota che la frequenza di rilassamento di una catena è sempre più bassa rispetto a quella della cellula equivalente a causa proprio della presenza dei gaps.
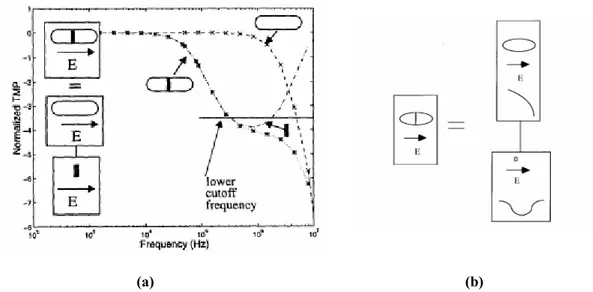
La figura 3.8 confronta la risposta in frequenza di una catena di due cellule parallela al campo applicato con quella di una cellula equivalente, il cui comportamento è simile a quello di un passa basso, con una certa frequenza di rilassamento. Tale frequenza dipende dalle proprietà totali della cellula, quali la dimensione e il materiale.
(a) (b)
Figura 3.8 Risposta in frequenza di una catena di due cellule e di una catena allungata (a);
Modello in cascata dei filtri passa basso ed elimina banda (b).
La catena ha due frequenze di rilassamento, di cui la prima è più bassa rispetto a quella della cellula equivalente.
Il comportamento frequenziale di entrambe le configurazioni va, comunque, a coincidere dopo il secondo ‘rilassamento’ della catena e questo può essere visto la cascata di due filtri (figura 3.8 b).
Il primo elemento della cascata è un filtro passa-basso, corrispondente al comportamento della cellula equivalente; il secondo rappresenta la risposta in frequenza della membrana e delle giunzioni di gap che separano le due cellule e corrisponde ad un filtro elimina banda.
La frequenza di rilassamento della catena corrisponde alla frequenza di cutoff del filtro elimina banda ed è più bassa di quella della cellula equivalente.
Capitolo 3
3.2.2. Catene lunghe di cellule
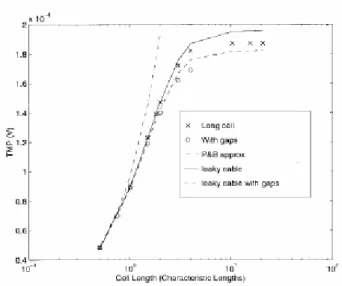
Per le catene di cellule più lunghe si affrontano gli stessi discorsi già visti per quelle corte. I risultati sono mostrati in figura 3.9 e meglio specificati in tabella 3.4.
Figura 3.9 Variazione del TMP con la lunghezza
delle catene lunghe e delle cellule allungate.
Per questo tipo di raggruppamento non sono state prese in considerazione le soluzioni analitiche e quindi i valori si sono confrontati semplicemente con il modello del cavo con perdite. Il caso LC1 prevede che la resistività del citoplasma sia calcolata tramite l’equazione (3.2.2) e il caso LC2 considera i gaps come una seconda sorgente.
Modello TMP (µV) # Cellule R (µm) L (µm) g R (nm) λ tot L FEM LC1 Errore (%) LC2 Errore (%) 8 5 10 20 2.4 22.7 26.4 16.3 27.2 19.8 7 7.5 15 20 50 200 3.0 1.2 0.4 26 38.3 52.4 28.6 45.4 51.7 10 18.5 -1.3 30 45.9 51.7 15.4 19.8 -1.3 5 7.5 15 300 0.2 42 37.3 -11.2 37.3 -11.2 3 7.5 25 300 0.2 42 37.3 -11.2 37.3 -11.2 10 5 25 20 200 300 4.8 0.9 0.8 40 101 110 30 114 117.2 -25 12.9 6.5 50.3 117.5 118.7 25.8 16.3 7.9 9 5 100 20 100 8.8 3.1 89.2 172.8 98.8 266.3 10.8 54.1 106.9 261.2 19.8 51.2 Tabella 3.4
Si osserva che la lunghezza totale in rapporto alla lunghezza d’onda (Ltot λ) diminuisce all’aumentare del numero di cellule e del raggio dei gaps. Inoltre, fissata la dimensione della catena e aumentando solo il raggio R , si riduce la differenza tra g i valori di TMP del cavo e quelli con FEM.
Interpolando i vari risultati si riescono ad avere i valori della frequenza di rilassamento riportati in tabella 3.5.
Come si può notare i valori della frequenza sono molto diversi, anche se si è usato, come termine di paragone, il caso di cavo con i gaps, considerati come una seconda sorgente. All’allungarsi della catena e al diminuire della dimensione dei gaps la frequenza diminuisce.
Capitolo 3 Modello frelax (Hz) # Cellule R (µm) L (µm) R (nm) FEMg LC 8 5 10 20 244 460 7 7.5 15 20 200 259 1149 360 9778 5 7.5 15 300 2996 29517 3 7.5 25 300 4769 36668 10 5 25 20 200 300 381 443 667 293 2374 2805 9 5 100 20 100 460 310 304 370 Tabella 3.5
Per capire meglio il comportamento delle lunghe catene di cellule si vedano le figure 3.10 e 3.11; la prima mostra la risposta in frequenza di due catene con la stessa lunghezza ma con un diverso numero di cellule (tre e cinque), la seconda, invece, il comportamento frequenziale di alcune catene con differenti raggi (stessa lunghezza).
(a) (b) Figura 3.10 Comportamento frequenziale di alcune catene con differente (a) numero di cellule e (b) raggio ma con la stessa lunghezza.
Se il numero di giunzioni non è elevato, la frequenza di rilassamento aumenta; inoltre, si ha un’attenuazione minore tra i due rilassamenti della stessa curva.
L’effetto del numero di giunzioni è legato alle caratteristiche del filtro elimina banda che rappresenta i gaps. Con un numero maggiore di giunzioni, la frequenza di cutoff più piccola si abbassa, portando all’allargamento della larghezza del filtro, mentre non si ha un cambiamento rilevante della frequenza più alta.
A differenza dei gaps, la variazione del raggio non influisce notevolmente sul comportamento frequenziale almeno fino alla prima frequenza di rilassamento. Dopo tale frequenza, le variazioni della curva dipendono dal diametro che diventa più largo rispetto alla lunghezza della catena.
3.2.3. Cellule e catene perpendicolari al campo
Sono stati ricavati dei risultati anche nel caso in cui il campo è applicato perpendicolarmente all’asse maggiore della catena. Per quanto riguarda il TMP, questo incrementa del 14% rispetto al TMP della cellula singola e ciò è in accordo con l’espressione di Pauly e Bernhardt per una cellula ellissoidale, la quale mostrava un incremento del 13.6%.
La frequenza di rilassamento per catene di due e tre cellule è riportata in tabella 3.6. Modello frelax (MHz) Catena di 2 cellule 10 Cellula equivalente 11.4 Catena di 3 cellule 10.8 Cellula equivalente 11.3 Tabella 3.6
Come si può vedere i risultati per le due configurazioni sono molto simili, quindi è possibile trascurare sia la membrana interna che i gaps, in quanto paralleli al campo applicato. In sostanza, le catene di cellule si possono modellare con cellule
Capitolo 3 equivalenti e sebbene questo introduca un certo errore, si ha un vantaggio a livello computazionale, infatti per il calcolo sono sufficienti poche giunzioni.
3.2.4. Cluster
Modello cluster
TMP
Sol. Num. Cell. Equiv. 2x1x1, E paral. 1.23 1.21 2x1x1, E perp. 0.84 0.85 2x2x1, E perp. 1.13 1.06 2x2x1, E parl. 1.68 1.5 Tabella 3.7
La tabella 3.7 raffronta i valori della tensione di transmembrana rilevati dal FEM con quelli delle cellule equivalenti. L’errore tra i risultati, anche se minimo, dipende dalla differente geometria, dalla discretizzazione del dominio e ancora dal piazzamento delle condizioni al contorno.
Quindi, un metodo computazionalmente meno costoso che modella i clusters è il seguente: ogni catena di cellule che costituisce il cluster viene sostituita con una cellula equivalente, posizionata nella direzione del campo applicato. In pratica vengono eliminate le membrane che dividono le varie cellule, le quali rimangono connesse tra loro da giunzioni, incorporate nella membrana interna e poste nella direzione perpendicolare la campo applicato (figura 3.11).
Figura 3.11 Sezione di un cluster 2x2x4
3.2.5. Proprietà dei gaps
La tensione di transmembrana e la frequenza di rilassamento si modificano anche al variare delle proprietà delle giunzioni, in particolare con la conducibilità. Nel caso in cui la conducibilità vari tra 10-7 e 10-5 S/m, il TMP si avvicina parecchio a quello di una singola cellula; quando poi raggiunge i 0.05 S/m è possibile approssimarlo con la tensione di una catena connessa da gaps riempiti di citoplasma.
Qualora la conducibilità assuma un valore intermedio tra i due casi limite, il TMP è calcolato mediante la seguente espressione:
(
)
(
( )
)
[
]
7 . 3 log 5 *TMP= single+ elongated −single − σgap (3.2.5)
in cui single si riferisce al TMP della cellula singola ed elongated a quello della cellula equivalente (figura 3.12).
Capitolo 3
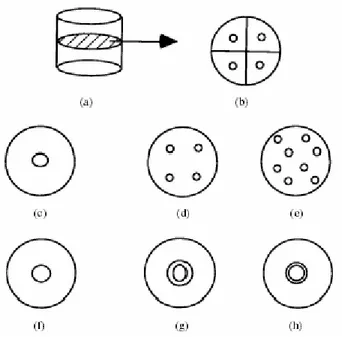
(a) (b) Figura 3.12 TMP al variare della conducibilità dei gaps.
In ogni modo, la presenza delle giunzioni influenza il TMP, anche se esse hanno delle dimensioni più piccole rispetto all’area superficiale della singola cellula.
La frequenza di rilassamento calcolata, ad esempio per una catena di due cellule (tabella 3.8), si riduce al diminuire sia della dimensione che della conducibilità delle giunzioni (figura 3.12 b). Tuttavia, tale diminuzione non è lineare, in altre parole la frequenza non si dimezza quando l’area si riduce della metà.
Gap R (nm) g frelax (MHz) Rapporto aree Rapporto frelax
G1 20 0.23 G1:G2 = 1.78 G1:G3 = 4 1.55 2.78 G2 15 0.15 G2:G3 = 2.25 1.8 G3 10 0.082 - - Tabella 3.8
La catena di due cellule, per intenderci, è modellata con semplici cilindri connessi da un gap centrale di area 0.01 µm2 (figura 3.12 a). La frequenza di rilassamento di questa configurazione è comparata con quella di altre strutture connesse da una distribuzione di quattro e otto giunzioni (fig.3.12 b-e). Per un singolo gap la frequenza è 0.53 MHz, per quattro è 0.77 MHz ed infine per otto 0.85 MHz.
Una ulteriore valutazione della frequenza si fa per due cilindri connessi da un singolo gap centrale e da due gaps di forma diversa ma con la stessa area (fig. 3.12 f-h). La frequenza per la struttura base è di 0.038 MHz, per il disco con un raggio più largo è di 0.15 MHz ed infine di 0.1 MHz per un disco di raggio piccolo.
Figura 3.12 Varie configurazioni di gaps
In pratica la forma dei gaps può significativamente variare il comportamento in frequenza della struttura: più i gaps sono distribuiti, più la frequenza aumenta