Capitolo 3.
Studi in vitro e
meccanismo d’azione
3.1
Azioni
di
Echinacea
su
fagociti
polimorfonucleati
Il principale uso dell’Echinacea a livello mondiale è dovuto proprio a questa attività [84][85]. Riguardo alle azioni che l’Echinacea svolgerebbe sul sistema immune, secondo alcuni Autori, vi sarebbe una stimolazione aspecifica delle reazioni difensive, in particolare delle risposte dei monociti/macrofagi, dei neutrofili e delle cellule NK [86]. Una conferma a questa ipotesi verrebbe da studi recenti che hanno dimostrato in vitro un’efficace modulazione delle risposte immuni dei macrofagi da parte di Echinacea [87].
In particolare, i macrofagi producono grandi quantità di ossido nitrico (NO), una molecola biologicamente attiva a concentrazioni nanomolari [88], importante nella difesa contro i patogeni e sintetizzata dall’ossido nitrico sintetasi (NOS) [89]. Questo enzima è rappresentato da tre isoforme di cui una, la iNOS, non è costitutivamente presente, ma viene indotta da stimoli infiammatori, quali ad esempio il lipopolisaccaride (LPS), attraverso la via del fattore nucleare κB (NF-κB) [90][91]. Anche il TNF-α attiva l’NF-κB, inducendo così la trascrizione di citochine pro infiammatorie, quali TNF-α, interleuchina-1 (IL-1) e
interleuchina-6 (IL-6), e di altre molecole importanti in questo processo [87].
Stevenson e coll. [87] hanno studiato l’effetto degli estratti alcolici dell’Echinacea e di alcuni suoi componenti standardizzati, quali ad esempio l’acido cicorico e la frazione alchilamidica, sull’attività dei macrofagi in vitro, dimostrando una riduzione dei livelli dell’NF-κB indotto da LPS e una riduzione della produzione del TNF-α e dell’NO indotti da LPS [87].
Utilizzando un protocollo che simulasse la digestione, Rininger e coll. [85] hanno investigato l’effetto di diverse preparazioni di E. purpurea sui macrofagi murini e sulle cellule monocitarie del sangue periferico
(PBMC) umane (analizzando vitalità dei monociti, produzione di NO, attività antiossidante e antiinfiammatoria nei macrofagi). Gli Autori hanno dimostrato che l’intero estratto della pianta e la polvere di radice possono agire come immunostimolanti attraverso un incremento della vitalità delle PBMC e un’induzione dell’attivazione dei macrofagi in vitro, come mostrato dall’incremento della produzione di TNF-α, NO, IL-1, IL-6 dose-dipendente. Gli estratti chimicamente standardizzati in contenuto di acido fenolico o echinacoside e il succo fresco pressato, al contrario, non hanno effetti immunostimolanti, ma mostrano proprietà antiinfiammatorie e antiossidanti a vari gradi [92].
Burger e coll. [68] hanno invece dimostrato che il succo fresco pressato di Echinacea, contenente fruttofuranosidi, è attivo a concentrazioni nanomolari e molto più potente dell’LPS nello stimolare la sintesi delle citochine nei macrofagi [92].
Alcuni Autori riportano, inoltre, un aumento della chemiotassi di macrofagi e neutrofili [43][60] e dell’azione ossidante dei neutrofili [87][88]. Si riscontra, inoltre, un aumento della proteina C reattiva, probabilmente dovuto ad una induzione della produzione di IL-6 da parte di monociti e macrofagi [44].
Secondo Bauer e coll. [93], il principio attivo responsabile dell’attività immunostimolante sembra risiedere nella frazione lipofila piuttosto che nella frazione polare, mentre secondo altri Autori [43], i responsabili di tale attività sono i polisaccaridi ad alto peso molecolare, come ad esempio l’arabinogalattano, i quali favoriscono la mobilità spontanea dei granulociti polimorfo-nucleati. Gli arabinogalattani attivano inoltre i macrofagi, i quali producono TNF-α, IL-1 e interferone-β2 (IFN-β2) e rilasciano fattori stimolanti per le cellule NK [43][60][63][92][94][96][97]. In contrasto con quanto riportato, un recente studio condotto nel 2008 da Tamta e coll. [98] dimostra invece che l’attivazione in vitro di monociti/macrofagi con estratti di Echinacea, è prevalentemente determinata dalla presenza nelle preparazioni di LPS e lipoproteine
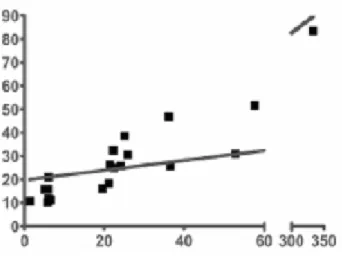
macrofagici tra quelle testate) [99]. Per dimostrare quanto asserito, un estratto di Echinacea è stato saggiato contemporaneamente con lisato di amebociti di Limulus (LAL) test, per stimare la quantità di LPS in un campione vegetale, e con Polimixina B, per verificare quanta dell’attività rilevata fosse imputabile al LPS. L’esito del test ha mostrato una forte correlazione tra il contenuto antigenico e l’attivazione macrofagica, che veniva inibita dal trattamento con la Polimixina B.
Figura 10: Correlazione tra l’attività stimolante i macrofagi RAW 264.7 ed i livelli di endotossina negli estratti di Echinacea. Le ascisse rappresentano il numero di unità di endotossina (EU) presente in ogni estratto espresso per milligrammo di materiale
vegetale macinato essiccato (EU/mg). Le ordinate
rappresentano l’attività complessiva ottenuta da 12 g di materiale vegetale macinato essiccato determinato usando cellule RAW 264.7 LPS sensibili.
Da Tamta et al., 2008
Gli Autori hanno, quindi, condotto un secondo studio sugli estratti di
Echinacea per valutare effettivamente quanto le componenti vegetali
incidessero sull’attivazione dei macrofagi: l’attività riscontrata inizialmente si è ridotta drasticamente quando i prodotti in esame sono stati trattati con una lipoproteina lipasi, per inattivare le lipoproteine, e con polimixina B come inibitore dei LPS [99].
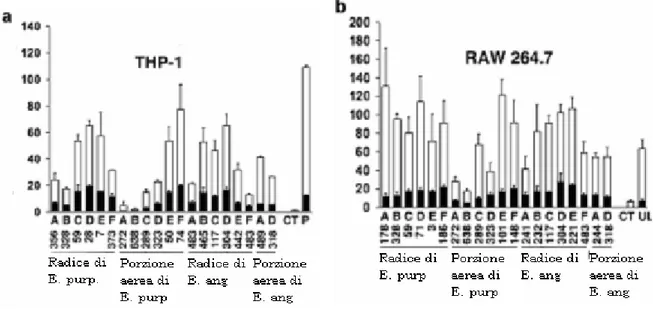
Figura 11: La maggior parte dell’attività stimolante di estratti ottenuti dalla totalità dell’Echinacea mostrata in vitro su monociti/macrofagi è eradicata ridotta fortemente dal trattamento con lipoprotein lipasi e Polimixina B. Le materie prime, radici e porzioni aeree, sia di E. purpurea che di E. angustifolia sono ottenute da sei produttori commerciali (A-F). (a)Nelle cellule THP-1 lipoproteine sensibili, le barre nere rappresentano estratti aggiunti alla coltura cellulare dopo un’incubazione a 37°C per 16h con lipoprotein lipasi (1 mg/mL), 10 M ASBEF un cocktail di inibitori delle proteasi (Sigma), 1% octilglucoside, e 0.2% di albumina serica bovina (BSA). Le barre bianche rappresentano gli estratti di controllo, in cui non è stata effettuata l’addizione di reagenti. (b) Nelle cellule RAW 264.7 LPS sensibili gli estratti sono stati aggiunti alle colture in presenza (barre nere) o assenza (barre bianche) di Polimixina B (100 g/mL). La Polimixina B è stata aggiunta 30 minuti prima di quella del campione. (CT) si riferisce alle cellule non trattate, (UL) si riferisce a 100 ng/ml di E. coli LPS ultra puro e (P) ri riferisce a 0.5ng/ml di lipoproteina sintetica Pam3CSK4. Da Tamta et al., 2008
Gli esiti ottenuti portano Tamta e coll. a speculare sulle differenze in termini di quantità o tipologia batterica come veri responsabili della sostanziale variazione osservata (anche 200 volte), ipotizzando come potenziali contaminanti batteri endofiti, batteri naturalmente presenti nell’ambiente, contaminazione dopo la raccolta oppure una combinazione di queste fonti. Il primo test condotto dagli Autori ha previsto l’analisi del contenuto di LPS in germogli di Echinacea fatti crescere in mezzi sterili, ma i livelli antigenici rilevati erano sostanzialmente equivalenti a quelli riscontrati nei campioni di
Echinacea forniti da altri produttori. Un secondo test ha mostrato,
invece, la presenza di alti livelli di componenti batteriche in campioni ottenuti da stoccaggio controllato. Le materie prime raccolte, lavate più volte, liofilizzate immediatamente, macinate ed estratte subito dopo la raccolta, avevano mostrato un’attività immunomodulatoria paragonabile, anche in questo caso, a quella riscontrata in campioni forniti da altri produttori. Sulla base di questi esiti, Tamta e coll. hanno dimostrato che i batteri erano penetrati all’interno dei semi ed inoltre che il trattamento successivo alla raccolta del materiale non è stata la causa del contenuto di LPS e lipoproteine, confermando, così, che tali componenti erano già presenti.
Gli Autori confermano con il loro studio il ruolo immunomodulatorio dei contaminanti batterici con risultati simili a quelli ottenuti in studi precedenti non coinvolgenti Echinacea [100]-[102]. Nel primo, si indica che il consumo di alimenti contenenti batteri può avere un effetto benefico sul sistema immunitario [100]. Il secondo mostra che, in un trial in doppio cieco, l’assunzione di Bifidobacterium lactis HN019, per tre settimane da parte di volontari sani, ha migliorato significativamente la fagocitosi delle cellule tumorali da parte delle cellule NK [101], dimostrando, così, che molti di questi effetti non hanno bisogno di cellule batteriche vive, ma possono essere imitati dal consumo di organismi uccisi dal calore [102].
Il fatto che l’induzione dell’attività dei monociti/macrofagi sia imputabile al contenuto di lipoproteine batteriche e LPS è sostenuta anche da altri Autori. Pugh et al. [99] hanno riscontrato una potente attivazione delle cellule accessorie del sistema immunitario da parte di questi contaminanti antigenici, anche in assenza di piante usate come immunomodulatori quali Echinacea, Ginseng americano e germogli di
Erba medica.
Analogamente allo studio precedente, lipoproteine batteriche e LPS sono risultati i più potenti attivatori dei monociti/macrofagi nei test in vitro, rispetto ad altri contaminanti della stessa natura.
Lo stesso vale per i dati ottenuti dai test: la maggior parte dell’attività (85-98%) degli estratti da 8 vegetali immunostimolanti è stata eliminata dal trattamanto con lipoprotein lipasi e con polimixina B.
La verifica di questa tesi è stata riscontrata con l’attività di preparazioni ottenute da germogli di Erba medica seminati in un terreno sterile: gli estratti ottenuti contenevano sia lipoproteine che LPS, e gli effetti ottenuti sulle colture in vitro scomparivano al trattamento con antibiotici.
3.2 Effetti di Echinacea su cellule dendritiche
umane
Wang e coll. hanno caratterizzato l’espressione genica delle cellule dendritiche (DCs) ed hanno mostrato la loro capacità di processare segnali ambientali ed attivare diversi programmi di trascrizione in risposta a stimoli differenti [26].
Le DCs giocano un ruolo importante nella promozione e nella regolazione sia dell’immunità innata che di quella acquisita [22]. In condizioni normali queste cellule si trovano in uno stato immaturo ma, indotte da segnali opportuni come infiammazione o antigeni ambientali, vanno in contro ad un rapido cambiamento ed iniziano una cascata di attività rivolte alla difesa dell’organismo. Inoltre le caratteristiche fenotipiche e funzionali delle DCs sono intimamente e dinamicamente correlate al loro stato di differenziazione e maturazione [37].
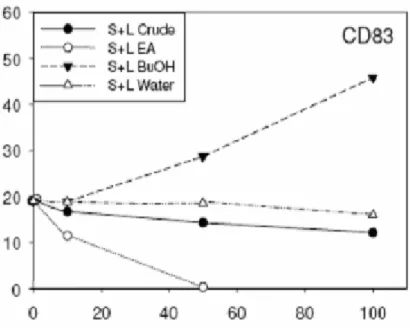
Con queste premesse, gli Autori hanno analizzato l’espressione del CD83, un marker di crescita presente sulle cellule dendritiche immature (iDCs), in risposta al trattamento con quattro sostanze. Soltanto il trattamento con la frazione butanolica di Echinacea purpurea (composta da estratti di foglie e di germogli detta [BF/S+L/Ep]) ha dato risultati rilevanti. Invece, l’esposizione delle iDCs all’acido cicorico, maggior componente della suddetta frazione, o ad una soluzione acquosa delle stesse porzioni vegetali non ha mostrato effetti significativi; infine, una frazione estratta con etilacetato ha mostrato attività biologiche opposte.
Figura 12: Bioattività specifica di tre subfrazioni di germogli e foglie (stem + leaf) di E. purpurea in cellule dendritiche umane immature. (S+L crude) Acido cicorico, (S+L EA) Etil acetato, (S+L BuOH) butanolo, (S+L Water) acquosa. Ascisse: Concentrazione g/ml. Ordinate: Attività relativa (% di cellule positive in FACS assay) Da Wang e t al., 2008.
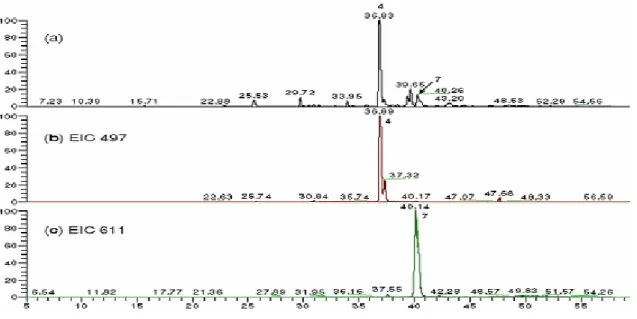
I risultati ottenuti sull’espressione della proteina CD83 suggeriscono che la frazione butanolica possa stimolare la maturazione delle iDCs. Per conoscere quali componenti della preparazione butanolica avessero tale capacità, gli Autori hanno analizzato l’estratto tramite HPLC, per caratterizzarne poi, con saggi bioguidati, i componenti principali o quelli più attivi. I dati riportano sette specifici fitocomponenti tra cui l’acido cicorico e la rutina, che sono stati successivamente quantificati ed utilizzati come componenti distintivi della frazione bioattiva.
Figura 13: Cromatogramma ottenuto con RP-HPLC della frazione butanolica a 330 nm. (a) Analisi su ESI-MS per ottenere un rapporto m/z dei componenti 4 e 7; (b) Rapporto m/z 497; (c) Rapporto m/z 611. Da Wang et al., 2008.
Lo studio dei geni che venivano espressi differenzialmente durante la maturazione delle DC indotta dalla frazione butanolica, ha messo in evidenza che geni codificanti per alcune chemochine, come CCL2, CCL5, e IL-8, venivano sovraespressi in maniera precoce [38].
Wang e coll. hanno, inoltre, identificato un’ipotetica rete di segnali che coinvolge i geni a risposta precoce attraverso l’attivazione delle vie del cAMP e delle PKC e la regolazione del calcio. Questi dati supportano studi precedenti che mettono in relazione la maturazione, la migrazione [39][41], la differenziazione e la capacità di presentare l’antigene [40] delle DCs, rispettivamente con cAMP, ioni calcio e PKC.
Infine, gli Autori hanno rivelato che le iDCs trattate con la preparazione di Echinacea o acido cicorico, avevano sovraespresso enzimi antiossidanti di difesa e proteine del citoscheletro in un tempo compreso da 12 a 24 ore. I primi giocano un ruolo importante nella sopravvivenza [42], nella capacità di indurre le cellule T e nel regolare la polarità della risposta immunitaria [43] delle DCs; le seconde sono quelle che
permettono l’espressione di componenti necessari alla presentazione dell’antigene [44], mentre le proteine del citoscheletro che legano l’actina sono importanti per la motilità delle cellule dendritiche.
Gli Autori concludono i loro studi ritenendo che specifici fitocomponenti in miscela possono avere significativi effetti immunomodularori su una determinata tipologia di cellule immunitarie.
3.3 Azioni di Echinacea sul sistema cannabinoide
In letteratura sono riportati sull’argomento tre lavori condotti dal 2004 al 2008 [74][75][103]. Nel primo studio effettuato nel 2004, Gertsch e coll. [75] hanno valutato gli effetti di rilevanti concentrazioni di Echinaforce™ (Ech) (10-25 g/ml) su fagociti polimorfonucleati umani. La preparazione già in precedenza aveva prodotto benefici nel trattamento del raffreddore [104]; inoltre è ottenuta da pianta fresca, standardizzata, registrata e con un contenuto di endotossina batterica minore di 0.5 EU/ml. Queste caratteristiche consentono agli Autori di valutare il potenziale immunomodulatorio dell’Echinacea, senza incorrere nell’erronea interpretazione dei risultati dei test.
Nel tentativo di chiarire la sua attività sul sistema immunitario, Gertsch e coll. ne hanno studiato gli effetti sull’espressione genica del TNF- sui monociti/macrofagi, che risultavano upregolati [105]. Sulla base degli esiti riscontrati, sono state condotte analisi di co-incubazione con Actinomicina D, un inibitore della trascrizione, per verificare se la sovraespressione riscontrata fosse dovuta alla sintesi de novo (trascrizione) o alla stabilizzazione dei trascritti all’interno delle cellule. I dati ottenuti hanno dimostrato che l’Echinacea aveva causato la sintesi
de novo di TNF-
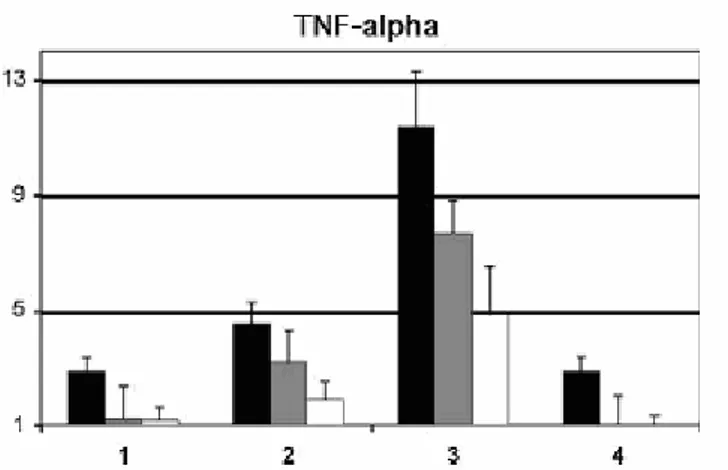
Figura 14: Concentrazioni ed effetti tempo-dipendenti dell’ Echinaforce™ sul TNF- mRNA nei monociti/macrofagi
supplemantati con PBMCs. 2 x 10*6 cellule sono state trattate con 25g/ml (barre nere), 15 g/ml (barre grigie), e 10 g/ml (barre bianche) di Ech ed il TNF-mRNA è stato quantificato con una RT-rt-PCR. L’ esperimento è stato anche eseguito con 4 ore di co-incubazione di ActD (2l). L’ ActD è stato aggiunto un’ora prima della stimolazione con Ech. I dati (+ S.E.) rappresentano 3 esperimenti indipendenti eseguiti in doppio con cellule ottenute da diversi donatori di sangue. I valori a Ct del TNF- sono stati normalizzati con GAP-DH. Una differenza minore o uguale a 2 volte risulta significativa. L’asse della ascisse riporta i quattro esperimenti. L’asse delle ordinate riporta i livelli di mRNA in rispetto al veicolo di controllo. Da Gertsch e coll. (2004).
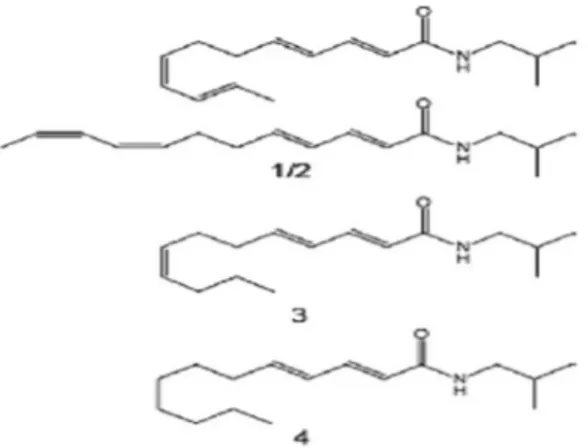
Inoltre studi cinetici dell’effetto immunomodulatorio sull’espressione della citochina sono stati condotti prima con il test ELISA e poi con la colorazione intracellulare FACS. Gli esiti hanno mostrato che, dopo stimolazione con Ech, le proteine del TNF- risultavano soltanto trascritte; dopo quella con LPS, contrariamente alle aspettative, l’espressione proteica risultava inibita e che in seguito ad una co-stimolazione con entrambi i reagenti venivano modulati sia trascritti che tradotti. Quest’ ultimo risultato suggerisce che alla base degli effetti modulatori ci sia un meccanismo d’azione molecolare in relazione con specifici principi bioattivi della tintura, finora non dimostrati. Per capire quali fossero i componenti dell’Echinacea coinvolti nella modulazione del TNF-Gertsch e coll. hanno studiato individualmente i suoi componenti, ed hanno mostrato che l’isobutilammide dell’acido dodeca-2E, 4E, 8Z, 10E/Z-tetraenoico (Figura 15: strutture1/2), trienoico (Figura15: struttura 3) ed i derivati dell’acido dienoico (Figura15: struttura 4), sono responsabili sia dell’upregolation del TNF-mRNA che dell’inibizione della sintesi proteica di TNF-stimolata dal LPS. Questi dati sono confermati dai risultati ottenuti con gli altri componenti della preparazione, acido cicorico, acido clorogenico e frazione polare, che risultavano inattivi (Figura 16).
Figura 15: struttura degli alchilammidi isolati. (1/2) coppia di isomeri dell’ isobutilammide dell’ acido
dodeca-2E,4E, 8Z,10E/Z-tetraenoico (1/2), dell’
isobutilammide dell’ acido dodeca-2E,4E, 8Z
trienoico (3) e l’ isobutilammide dell’ acido dodeca-2E,4E-dienoico. Da Gertsch e coll. (2004)
Figura 16: induzione del TNF- mRNA nei monociti/macrofagi da parte di ½, 3, 4, acido cicorico, acido clorogenico e Frazioni polari che contengono residui oligosaccaridici di Echinaforce misurati con RT-rt-PCR. Differenze > di due volte sono significative. Barre bianche: 0.5 M (frazione polare 0.5 ppm) dei componenti del test; barre grigie: 5M (frazione polare 5 ppm) incubati con 2 x 10*6 cellule per 22 ore prima dell’analisi con RT-rt-PCR. I dati rappresentano valori medi +/- S.E. di due esperimenti indipendenti. Le ascisse rappresentano i singoli esperimenti. Le ordinate rappresentano i livelli di TNF-alpha mRNA in rapporto al veicolo di controllo. Da Gertsch e coll. (2004)
Le somiglianze strutturali e funzionali tra le alchilammidi dell’Echinacea, l’anandammide (arachidonil-etanolammina) ed il 2-AG (arachidonoil-glicerolo) degli endocannabinoidi, hanno fatto ipotizzare agli Autori che i recettori CB2 potessero essere il bersaglio di queste componenti. Tali recettori giocano un ruolo prominente per gli effetti
delle alchilammidi, sono espressi soprattutto in periferia, specialmente nei monociti/macrofagi [106], come recettori accoppiati a proteine G (GPCRs) e correlati alle attività di messaggeri ed attività genica. La tesi di Gerstch e coll. è stata dimostrata dagli accertamenti effettuati sul CB2 per verificare che fosse realmente il sottotipo recettoriale coinvolto. Come per gli studi sull’espressione genica del TNF-, antagonisti dei CB1 e CB2, rispettivamente SR147778 e SR144528, sono stati usati in combinazione con l’alchilammide 1/2. I dati dimostrano che soltanto l’antagonista specifico dei CB2 ha soppresso la trascrizione del TNF-quindi le alchilammidi 1/2 sembrano mediare un segnale agonista tramite i recettori CB2. L’analisi di una frazione di monociti/macrofagi usata nei test con metodo western blot, conferma la presenza dei CB2 su queste cellule, anche se occorreranno ulteriori studi per valutare una diretta regolazione dell’espressione dei CB2 da parte delle alchilammidi. A sostegno della tesi di Gerstch e coll. l’upregolation del cAMP costitutivo da parte dei principi attivi dell’Ech (1/2), e la sua soppressione ad opera del PTX (tossina della pertosse), confermano di nuovo la partecipazione dei recettori CB2 accoppiati a proteina G.
Nel 2006, Raduner e coll. [74] hanno caratterizzato l’interazione delle alcamidi con il recettore CB2 umano rispetto all’affinità di legame, alla specificità del ligando e alle conseguenze funzionali concentrazione dipendenti, in linee cellulari macrofagiche. Gli studi di legame si sono basati su test di spiazzamento di un radioligando ad alta affinità per i cannabinoidi. E’ stato mostrato che i principali componenti dell’Echinacea, l’isobutilammide dell’acido dodeca-2E, 4E, 8Z, 10E/Z-tetraenoico (A1) e l’isobutilammide dell’acido dodeca-2E, 4E-dienoico (A2), si legano al recettore CB2 più saldamente ed a basse concentrazioni (CB2~60nM e CB1>1500nM) rispetto agli endocannabinoidi, ed ancora che tale legame si instaura generalmente sullo stesso sito scelto dal radioligando [107]. Questo esito, insieme alle analogie strutturali evidenti tra la alcamidi e l’anandammide, ha creato
Le opinioni degli Autori che hanno già condotto questo tipo di analisi sono discordanti poiché, mentre Chen e coll. [108] ritengono che le alchilamidi si dispongano in una estesa conformazione a doppia elica, McAllister et al. [109] suggeriscono che l’anandammide necessiti di una conformazione ad “U” per interagire con il recettore CB1. Questo perché l’endocannabinoide ha una catena acilica con quattro doppi legami non coniugati che le permettono una maggire flessibilità e, di conseguenza, la capacità di alloggiare nella tasca recettoriale. Raduner e coll. Hanno, quindi, condotto degli studi di simulazione di legame con l’isobutilammide dell’acido dodeca-2E, 4E, 8Z, 10E/Z-tetraenoico: la disposizione all’interno del recettore CB2 avviene attraverso un arrangiamento conformazionale che vede l’inserzione di una coda alchilica all’interno della tasca idrofobica formata dalla porzione aromatica dell’anello di una tirosina (Tyr 190), ed inoltre la formazione di due doppi legami in C-2 e C-4 che sono assenti con l’anandammide. Per meglio caratterizzare gli effetti funzionali del legame tra alcammide e CB2 è stata misurata la concentrazione intracellulare di ione calcio libero e lo studio è stato condotto in promielociti HL60, in cui il recettore cannabinoide risultava espresso positivi) o meno (CB2-negativi). Entrambi gli alcamidi A1 e A2 hanno provocato un aumento di Ca++ libero intracellulare soltanto nelle cellule che esprimevano il recettore [110].
Altri studi [111] hanno mostrato che i livelli del Ca++ potrebbero essere modificati anche dall’interazione del CB2 con un altro endocannabinoide e che il 2-AG è da considerarsi il suo vero ligando endogeno.
Altri Autori [112] suggeriscono anche che la liberazione intracellulare di ione calcio avvenga con la mediazione di entrambe le molecole, delle quali il 2-AG, responsabile della risposta immediata (il rilascio di calcio avviene in 100 ms secondo una misurazione in tempo reale), e l’anandammide della risposta tardiva. Secondo Makriyannis e coll.
infatti, l’anandammide prima di dirigersi al sito recettoriale necessita di un primo legame al doppio strato fosfolipidico di membrana.
Nel 2008 Gertsch [3] ha condotto un nuovo studio in cui riesamina ed amplia i risultati raggiunti nel 2004 e ripropone le amidi degli acidi grassi (FAAs o alcamidi) come nuova classe di cannabinomimetici, cercando un collegamento tra regno vegetale ed animale. Queste sostanze non soltanto sono strutturalmente correlate agli endocannabinoidi, ma originano anche da precursori biosintetici simili [113]: nelle piante sono state individuate strutture di varia complessità, dalle più semplici N-aciletanolammine (NAEs) a isobutilamidi o dimetilbutilamidi polinsaturi, con catene alchiliche di lunghezza variabile da 6 a 24 atomi di carbonio. Inoltre le FAAs non sono state soltanto riconosciute nell’Echinacea [114]-[118], anche se è soprattutto nelle sue specie che si riscontra un potenziale farmacologico significativo [119][120].
Nonostante siano ancora sconosciuti il sito d’azione, l’esatta natura biosintetica ed il trasporto delle FAAs, Lopez-Bucio e coll. [120] ritengono che queste molecole agiscano come ormoni vegetali o regolatori della crescita, come confermato da altri Autori [121]-[123] che ritengono le alcamidi responsabili della modulazione dei processi cellulari. Altri Autori ancora ritengono che la produzione delle FAAs sia regolata dalle stesse piante, come dimostrano specifiche idrolasi delle ammidi degli acidi grassi (FAAHs), all’interno di un sistema bilanciato tra attivazione ed inibizione reciproca. Inoltre, vista la somiglianza strutturale tra FAAHs vegetali ed animali, gli Autori [124]-[127] ritengono possibile che alcune isobutilamidi, comprese quelle dell’Echinacea, possano parzialmente inibire le idrolasi animali. Cravatt [128] e Lin [129] sostengono questa ipotesi osservando che, negli animali, almeno una parte dell’effetto antinfiammatorio delle alcamidi potrebbe essere mediato proprio dal mantenimento dei livelli endogeni di anandammide. Inoltre le FAAs dell’Echinacea risultano più
stabilità risiede nelle differenze strutturali della catena acilica che, nell’anandammide, è costituita da una porzione arachidonica, mentre nelle alchilamidi deriva da acidi undeca- e dodecanoici insaturi.
In conclusione, basandosi sui dati di differenti gruppi di ricerca [130]-[132], Gertsch e coll valutano la possibilità di indirizzare il profilo dell’Echinacea come rimedio vegetale antiinfiammatorio, più che immunomodulatorio. La rilevanza dei FAAs come principi attivi della pianta è stata rivisitata anche da Bauer e Woelkart [133]: mentre il loro uso come analgesici ed antiinfiammatori è supportato da evidenze terapeutiche, quello per il trattamento della sindrome da raffreddamento mostra apparentemente risultati controversi [134], [135]. In ogni caso i sintomi che nascono dalle infezioni durante il raffreddore spesso coinvolgono dolore e infiammazione, quindi si potrebbe supporre un legame tra il sistema endocannabinoide e il raffreddore, poiché il sistema endogeno comprende tipicamente effetti diretti contro queste componenti.