1. INTRODUZIONE
1.1. APPRENDIMENTO E MEMORIA
Il comportamento di qualunque essere vivente dipende dall’interazione di fattori genetici e ambientali. I più importanti meccanismi, con cui l’ambiente influisce sul comportamento, sono l’apprendimento e la memoria. L’apprendimento è il processo mediante il quale vengono acquisite informazioni dal mondo circostante; la memoria è il processo mediante il quale tale conoscenza viene codificata, conservata e successivamente utilizzata.
Sono descritte due forme di memoria: 1) una memoria implicita o non dichiarativa, molto rigida, la quale riguarda le capacità percettive e le abilità motorie e che viene richiamata alla mente in modo non cosciente; sono esempi di memoria implicita l’addestramento all’esecuzione di compiti, motori o percettivi di tipo riflesso; 2) una memoria esplicita o dichiarativa, molto duttile che riguarda il ricordo di persone, luoghi e oggetti e richiede un ricordo consapevole (Squire, 1992) (vedere Fig.1 pag. 2).
La memoria,in generale, è il frutto di quattro distinti processi qui di seguito riportati e descritti:
1. La codificazione: processo mediante il quale le nuove informazioni sono acquisite, analizzate e trasformate in tracce mnemoniche. Dall’efficienza della codificazione, data dall’attenzione e dalla motivazione, dipende il grado di conservazione della traccia mnemonica.
2. Il consolidamento: fase in cui i processi di modifica delle nozioni appena acquisite, ancora labili, sono resi più stabili e durevoli nel tempo. Il consolidamento richiede l’ espressione di geni e la sintesi di proteine ex- novo.
3. La conservazione: processo mediante il quale la memoria viene mantenuta nel tempo. Tale capacità di conservazione sembra essere illimitata.
4. Il recupero:l’insieme dei processi che consentono di richiamare alla mente un ricordo. E’ la fase di ricostruzione di tutte le informazioni immagazzinate, che sono state “archiviate” in zone diverse del neocortex.
Fig.1:Sistemi multipli di memoria (modificata da Squire and Zola-Morgan, 1991).
Le memorie implicita ed esplicita coinvolgono aree cerebrali distinte. Le informazoni conservate come memorie esplicite vengono inizialmente elaborate in una o più delle cortecce associative polimodali (cortecce prefrontale, limbica e parieto-temporo-occipitale) che sintetizzano le
informazioni di natura visiva, uditiva e somatica. Da qui le informazioni vengono trasferite, in serie, alle cortecce paraippocampica e peririnale, e quindi alla corteccia entorinale, al giro dentato, all’ippocampo, al subiculum e infine di nuovo alla corteccia entorinale. Da quest’ultima le informazioni vengono rinviate alle cortecce paraippocampica e peririnale e infine ancora alle cortecce associative polimodali della neocortex (Squire, 1992).
La memoria implicita è custodita nei circuiti percettivi, motori e limbici. Paradigmi sperimentali, condotti su animali per studiarne la memoria implicita, hanno messo in evidenza che tale tipo di memoria può essere distinta in forme non associative e forme associative. L’apprendimento non associativo si manifesta quando il soggetto è esposto ripetutamente ad uno stimolo e impara a distinguerne le proprietà. Le due forme di apprendimento non associativo più comuni sono l’abitudine e la sensitizzazione. L’apprendimento associativo si manifesta, invece, quando il soggetto apprende quali siano le relazioni che intercorrono fra due stimoli o fra uno stimolo ed una risposta comportamentale. L’apprendimento associatvo si può distinguere ulteriormente in due forme: il condizionamento classico e il condizionamento operante.
Il condizionamento classico (CC) fu scoperto verso la fine del secolo scorso dal fisiologo russo Ivan Pavlov (Pavlov, 1927). Egli dimostrò che in un soggetto sperimentale (per esempio un cane), uno stimolo neutro che normalmente non produce risposte apprezzabili (ad esempio un suono), definito “stimolo condizionato” (CS), presentato ripetutamente in stretta associazione temporale con la presentazione di uno stimolo (come la vista del cibo), definito “incondizionato” (US) in grado di indurre una certa risposta innata (come l’aumento della salivazione), è in grado di produrre,
anche se presentato da solo, la risposta innata definita risposta indotta, appresa o “condizionata”.
Per ottenere risposte apprese devono verificarsi condizioni ben definite: i due stimoli CS e US devono essere presentati al soggetto sperimentale contemporaneamente, oppure il CS deve precedere di poco l’US; i due stimoli CS ed US non devono essere forniti insieme ad altri stimoli che potrebbero ostacolare la loro associazione; la latenza tra CS e US deve essere costante nelle varie presentazioni; l’associazione CS-US deve essere ripetuta un numero sufficiente di volte (Pavlov, 1927).
1.2. STUDIO DEI MECCANISMI DI BASE DELLA MEMORIA: TECNICHE A CONFRONTO
La memoria può portare a plasticità neuronale ovvero a cambiamenti della biochimica e della struttura delle sinapsi.
In relazione al tempo necessario per indurre questi cambiamenti plastici si distinguono due tipologie di memoria: una memoria a breve termine, che dura da pochi minuti ad un massimo di 24 ore, ed una memoria a lungo termine, che perdura per giorni, settimane o addirittura per tutta la vita di un organismo. Lo studio dei meccanismi alla base dei processi mnemonici, sia a breve che a lungo termine, può essere effettuato su vari modelli animali e utilizzando tecniche sperimentali diverse.
Per lo studio della memoria a breve termine, vengono usate soprattutto tecniche di elettrofisiologia, basate sulla registrazione dell’attività dei singoli neuroni coinvolti nel consolidamento delle tracce mnemoniche. Tramite l’utilizzo di queste tecniche in modelli sperimentali come gli invertebrato, quali Aplysia californica e Hirudo medicinalis, è stato possibile studiare in dettaglio le forme di apprendimento non associativo.
Tali modelli sperimentali presentano il vantaggio di avere sistemi nervosi semplici, di facile accesso per lo sperimentatore e di essere costituiti da circuiti neuronali complessi e tuttavia di presentare atti comportamentali complessi.
È stato dimostrato che negli invertebrati le forme di apprendimento non associativo determinano modificazioni della efficacia sinaptica (Castellucci et al.,1986; Scuri et al., 2007).
Tecniche elettrofisiologiche sono largamente utilizzate anche in slices di vertebrato per lo studio dei meccanismi di base dell’apprendimento associativo.
Studi effettuati su slices di ippocampo di ratto hanno dimostrato infatti che neuroni della formazione ippocampale posseggono capacità plastiche durevoli nel tempo simili a quelle richieste per la conservazione delle tracce mnemoniche. L’applicazione di una breve scarica di stimoli ad alta frequenza nelle vie eccitatorie ippocampali, determina un aumento dell’ampiezza dei potenziali postsinaptici eccitatori dei neuroni bersaglio. Questo aumento dell’efficacia sinaptica viene definito long term potentation (LTP) (Bliss and Lomo., 1973).
L’LTP può essere suddiviso in due fasi temporalmente distinte ciascuna caratterizzata da meccanismi cellulari e molecolari. La fase precoce (early LTP o E-LTP) è indotta da una singola stimolazione tetanica, dura poche ore ed è legata esclusivamente a modificazioni a livello di proteine già preesistenti. La ripetizione della stimolazione tetanica determina invece la comparsa di una fase duratura dell’LTP, mantenuta per almeno 24 ore, che viene definita LTP tardivo (late LTP o L-LTP) e che richiede modulazione genica e sintesi di proteine ex-novo.
È ormai noto che la memoria a lungo termine richiede la modulazione dell’espressione genica e quindi il coinvolgimento di specifici meccanismi molecolari (Matthies, 1989).
Risulta per tanto che il solo approccio elettrofisiologico non sia appropriato per studiare questi meccanismi genetici coinvolti nel consolidamento delle tracce mnemoniche. Sono state quindi adottate tecniche di biologia molecolare specifiche per determinare modificazioni a livello sia del mRNA (tecniche di trascrittomica) sia e a livello della sintesi di proteine (tecniche di proteomica).
Lo studio del trascrittoma prevede l’utilizzo di varie tecniche, le più conosciute sono il microarray (chip genici) e l’ SSH.
I chip genici sono una tecnica relativamente semplice che permette di analizzare più di due campioni contemporaneamente e che richiede concentrazioni minime di mRNA. Rispetto all’SSH, informazioni sulle sequenze da esaminare ed, inoltre, risulta essere molto più costosa. L’SSH rispetto alle altre tecniche di analisi di patterns di espressione genica presenta i seguenti vantaggi:
• richiede concentrazioni minime di mRNA (1-2 µg) • segue un protocollo semplice e veloce
• riusce ad isolare anche sequenze rare o poco espresse
L’SSH presenta tuttavia lo svantaggio di confrontare solamente due campioni per volta.
Per l’analisi dell’espressione genica si possono utilizzare la PCR tradizionale o la PCR real-time. Rispetto alla PCR tradizionale, la real-time PCR permette di monitorare in tempo reale l’andamento della reazione ed
effettuare una quantizzazione del prodotto di amplificazione, in quanto esiste una relazione quantitativa tra l’ammontare del templato iniziale e l’ammontare del prodotto di PCR nella fase esponenziale della reazione. I prodotti non richiedono ulteriori processamenti dopo l’amplificazione, riducendo così il rischio di contaminazione
Per analizzare se i trascritti individuati siano fisiologicamente funzionali si possono utilizzare tecniche di proteomica come il western blot, che permette l’identificazione di una proteina di interesse mediante la reazione con il suo specifico anticorpo. Il western blot presenta i seguenti vantaggi:
• è un test molto sensibile e specifico;
• l’identificazione avviene in base al peso molecolare della proteina. Nel mio internato di tesi ho avuto la possibilità di conoscere ed applicare le tecniche di PCR real-time e western blot per analizzare, rispettivamente, i geni la cui espressione viene modificata in seguito a cambiamenti fisiologici di base ottenuti tramite CFC e la modulazione di proteine specifiche in seguito a trattamento cronico con acetil-L-carnitina (ALC), sostanza capace di modulare le performance comportamentali. Queste due tecniche possono venire integrate l’una con l’altra per ottenere risultati più completi nell’indagine sui meccanismi molecolari legati alla memoria e all’apprendimento.
1.3. IL PARADIGMA DEL CONTEXTUAL FEAR CONDITIONING Le risposte condizionate comprendono anche quelle indotte dalla presenza di un pericolo, le quali vengono definite come “risposte condizionate di paura”. L’apprendimento consente ad ogni individuo di rispondere in maniera tendenzialmente più efficace a stimoli nocicettivi. Questa forma di
apprendimento è essenziale per la sopravvivenza, perché consente di associare stimoli che indicano la presenza di un pericolo con il pericolo stesso. Essendo intimamente legato alla sopravvivenza, il condizionamento alla paura viene rapidamente appreso e mantenuto per lungo tempo. Il fear conditioning è un paradigma comportamentale di condizionamento classico usato per studiare le basi neurali della memoria emozionale. Al soggetto viene ripetutamente presentato uno stimolo, ad esempio uno stimolo acustico (stimolo condizionato), che non provoca alcuna reazione, appaiato con uno shock elettrico (stimolo incondizionato), che provoca una reazione di paura. L’acquisizione dell’associazione fra i due stimoli viene valutata a intervalli temporali diversi ed è rappresentata dal manifestarsi di una forte reazione di paura: l’animale si blocca (freezing), il suo battito cardiaco accelera e la pressione arteriosa sale. La risposta di freezing è definita come la completa assenza di mobilità del corpo ad eccezione dei movimenti respiratori (Fanselow, 1990; LeDoux, 1995).
La struttura nervosa cruciale per il manifestarsi del condizionamento alla paura è l’amigdala ed in particolare il nucleo basolaterale dell’amigdala stessa (Le Doux, 2000).
Nel contextual fear conditioning (CFC) è l’ambiente, o contesto, a funzionare come CS, di conseguenza, la sola presentazione dell’ambiente senza US sarà in grado di determinare risposte di paura.
Nel CFC, quindi, lo stimolo condizionato è rappresentato dall’apparato sperimentale di condizionamento, mentre lo stimolo incondizionato, come per il condizionamento classico alla paura, è rappresentato da una scossa elettrica alle zampe (LeDoux, 1995). E’ noto che, dopo essere stati sottoposti al paradigma comportamentale del CFC, i soggetti addestrati apprendono con maggior facilità e mostrano di associare l’ambiente in cui si trovano agli stimoli avversivi (US) forniti loro durante l’esplorazione
dell’apparato di condizionamento. I ratti addestrati mostrano freezing ogni volta che sono reinseriti nell’apparato di condizionamento (retrieval test). Tale reazione non è rilevabile quando gli stessi soggetti, precedentemente sottoposti alla sessione di condizionamento, sono inseriti in un ambiente diverso da quello in cui sono stati forniti gli stimoli avversivi. Pertanto il freezing espresso nel retrieval test rappresenta una risposta condizionata dovuta all’associazione tra quello specifico ambiente e gli stimoli avversivi. Inoltre il freezing non è rilevabile quando i soggetti esplorano l’apparato di condizionamento senza che siano forniti loro stimoli avversi (exploration procedure) oppure quando gli animali vengono inseriti nell’apparato di condizionamento e sono loro forniti stimoli avversi di pari intensità ma in un intervallo di tempo minore, rispetto al protocollo del CFC, così da impedire l’associazione fra questi stimoli e il nuovo ambiente (shock-only procedure) (Milanovic et al., 1998). . Numerosi studi hanno dimostrato che questo tipo di condizionamento è legato all’attivazione sia dell’amigdala che dell’ippocampo (Rodrigues et al., 2001). L’ippocampo ha un ruolo fondamentale nella formazione e nel consolidamento delle relazioni tra stimoli ed eventi (Rudy et al., 2004).
1.4. IL CIRCUITO NERVOSO DELLA PAURA
Il circuito nervoso che sottende alle risposte di paura è stato studiato per la prima volta, tramite esperimenti di condizionamento alla paura. Tramite l’applicazione di uno stimolo condizionato uditivo sono state analizzate le modalità con cui tale stimolo raggiunge il cervello. Seguendo questo percorso è stato scoperto che la via uditiva passa dal talamo per giungere poi la corteccia uditiva. Danneggiando le aree corticali non si ottiene alcuna conseguenza nella manifestazione delle risposte alla paura. Se
invece vengono danneggiate le aree del talamo, allora le manifestazioni della paura scompaiono (LeDoux, 1995). È stato quindi ipotizzato che il talamo proietti anche in altre zone cerebrali. L'area determinante, senza la quale le reazione alla paura non si instaurano, è l'amigdala. L’interruzione delle connessioni tra talamo e amigdala inibisce il condizionamento alla paura. In particolare esperimenti farmacologici condotti al livello del nucleo basolaterale dell’amigdala hanno evidenziato un blocco dell’induzione duratura della traccia mnemonica (LeDoux, 2000). Il nucleo basolaterale dell’amigdala proietta al nucleo centrale, che costituisce la principale via di “uscita” dall’amigdala. Le proiezioni dal nucleo centrale dell’amigdala sono dirette all’ipotalamo e alle strutture del tronco dell’encefalo coinvolte nel controllo sia del sistema nervoso autonomo che del sistema endocrino, soprattutto dell’asse ipotalamo-ipofisi-surrene (vedere Fig. 2). L’ipotalamo regola l’omeostasi dell’organismo, in quanto integra le risposte del sistema nervoso autonomo con quelle del sistema endocrino tramite la manifestazione delle seguenti risposte comportamentali: il controllo della temperatura corporea, della frequenza cardiaca, dell’assunzione di acqua e cibo e della pressione arteriosa (Le Doux, 2007).
I circuiti coinvolti nelle risposte emozionali sono divisi in due vie: la via “alta”, corticale a conduzione lenta; la via “bassa”, sottocorticale più rapida (vedere Fig. 3). La via alta prende inizio a livello dal talamo sensoriale e termina a livello della corteccia sensoriale, dove viene elaborato lo stimolo che inseguito raggiungerà l’amigdala.
Fig.2: Schema delle regioni cerebrali coinvolte nell’apprendimento alla paura (tratto da
LeDoux, 2007)
La via bassa invece porta direttamente le informazioni dal talamo sensoriale all’amigdala. Questa via, essendo più veloce, anticipa quella alta, fa sì che le risposte di paura non coinvolgano la corteccia e quindi l’elaborazione cosciente (LeDoux, 1995).
Fig.3 Schema riassuntivo del circuito della paura di Le Doux (modificato da Le Doux,
Nel ratto l’informazione sensoriale richiede circa dodici millesimi di secondo per percorrere la via bassa e il doppio del tempo per percorrere quella alta. Il vantaggio evolutivo è più che evidente, per la sopravvivenza è meglio reagire a delle circostanze potenzialmente pericolose come se lo fossero davvero piuttosto che non reagire affatto. L’amigdala ha connessioni con le reti della memoria a lungo termine, tra cui il sistema ippocampale e le aree corticali che interagiscono con l’ippocampo per immagazzinare in maniera durevole le informazioni. Tutti questi circuiti possono contribuire all’attivazione dei ricordi a lungo termine rilevanti per le implicazioni emotive degli stimoli immediati. Negli esperimenti sul condizionamento alla paura, è stato preso in esame un altro aspetto cruciale: il ruolo del contesto in cui l’addestramento viene condotto. Nel condizionamento classico non è soltanto lo stimolo condizionato a provocare le reazioni di paura, ma anche il contesto in cui l'esperimento di svolge. Lesioni dell’ippocampo prima o subito dopo il condizionamento contestuale comportano perdita delle risposte di freezing, mentre lesioni eseguite parecchi giorni dopo l’addestramento non compromettono l’acquisizione delle risposte condizionate (Burgess et al., 1999). Questa suggerisce che l’ippocampo possa essere coinvolto nell’acquisizione e nel mantenimento temporaneo della rappresentazione del contesto (che in questo ccaso funziona da CS), mentre l’amigdala sarebbe coinvolta nell’associazione US-CS, nell’elaborazione delle singole entrate sensoriali, nell’attribuzione ad esse di un significato emotivo e nella produzione delle risposte di paura (Maren and Fanselow, 1996). Oltre all’ ippocampo e all’amigdala, è noto che anche la corteccia cerebrale è coinvolta nell’apprendimento delle risposte condizionate alla paura (Lacroix et al., 2000). Tramite lesioni irreversibili sono state individuate, le aree corticali coinvolte nell’apprendimento alla paura: l’area prefrontale (PFC)
(Vouimba et al., 2000), l’area frontale (FC) (Rosen et al., 1992), l’area parietale (PAC) (Suzuki, 1996) e l’area peririnale (PC) (Corodimas and Le Doux, 1995). Studi successivi, hanno evidenziato che la lidocaina (LIDO) blocca le attività neuronali in maniera reversibile in PFC, FC, PAC e PC. Tali aree giocano ruoli contemporanei ma distinti nella acquisizione della traccia mnemonica del freezing dato da US accoppiato a un CS acustico o contestuale (Sacchetti et al., 2002). L’inattivazione di PC indebolisce la ritenzione della memoria a breve termine di entrambe le tracce mnemoniche in quanto il freezing di tali soggetti ha durata inferiore rispetto ai controlli. Allo stesso intervallo, l’inattivazione di PFC migliora la ritenzione di entrambe le tracce in quanto i soggetti sperimentali mostrano freezing più duraturo dei controlli. PFC è connessa con l’amigdala, ed è stato supposto che eserciti un’azione inibitoria sulle attività dell’amigdala integra (Morgan, LeDoux, 1995). PAC e FC non giocano ruoli essenziali nel CFC, però giocano un ruolo importante nell’elaborazione della memoria a lungo termine, poiché 72 ore dopo il CFC sono danneggiate entrambe le tracce mnemoniche (Acquas et al., 1996). L’integrità funzionale di PC è rilevante durante la fase di consolidamento. Infatti la sua inattivazione con LIDO è seguita dal danneggiamento della memoria a breve e a lungo termine sia nel condizionamento acustico sia nel condizionamento contestuale. E’ stato mostrato che l’integrità funzionale di PC è necessaria per la memorizzazione di entrambe le tracce mnemoniche fino a 8 giorni dopo la fase di acquisizione (Sacchetti et al., 1999a;b). PC gioca un ruolo molto importante nella memorizzazione delle risposte alla paura ed ha un ruolo specifico nella memoria e nell’apprendimento avversivi. PC è una delle più importanti stazioni di connessione tra la neocortex e le strutture subcorticali; infatti, sia nei roditori che nei primati, è il punto in cui convergono le informazioni dalle aree associative e da
strutture sottocorticali, come mostrato da evidenze morfologiche (Suzuki, 1996), elettrofisiologiche (Vann et al., 2000) e comportamentali (Sacchetti et al., 1999a;b). Numerosi studi hanno dimostrato che l’assenza funzionale dell’amigdala durante l’acquisizione è seguita da amnesia al CS acustico e al contesto. Invece l’assenza funzionale dell’ippocampo induce amnesia solo per la ritenzione del contesto (Sacchetti et al., 1999a;b). Poiché ancora non è noto attraverso quali vie l’informazione contestuale arrivi all’ippocampo, l’inattivazione di specifiche aree cerebrali supporta l’ ipotesi corrente secondo la quale l’informazione contestuale potrebbe arrivare all’ippocampo dalla corteccia peririnale (LeDoux, 2000).
1.5. MODIFICAZIONI DELL’ECCITABILITÀ IPPOCAMPALE LEGATE A FEAR CONDITIONING
Gli effetti del CFC sull’ area CA1 dell’ippocampo sono stati analizzate applicando stimolazioni tetaniche (HSF) a livello delle sinapsi tra le collaterali di Schaffer e i dentriti dei neuroni del campo CA1 in slices di ippocampo di ratto adulto (vedere Fig.4).
Come soggetti sperimentali sono stati utilizzati ratti condizionati, che esibivano freezing duraturo quando posti nuovamente nello stesso apparato di condizionamento (retrieval test); come soggetti di controllo sono stati utilizzati: soggetti exploration, ratti che avevano liberamente esplorato l’apparato senza ricevere scosse elettriche;
Fig.4: Slices di ippocampo dove viene mostrata la posizione dei due elettrodi,
stimolante e registrante (modificata da Sacchetti et al., 2001)
soggetti shock-only, ratti che avevano ricevuto lo stesso numero di scosse ad un’uguale intensità pari a quella dei condizionati, ma con le scosse compresse temporalmente così che i ratti non potevano associare US al contesto e soggetti naive, ratti mai entrati nell’apparato di condizionamento. Tutti gli animali di controllo, non esibivano la risposta condizionata di freezing (Sacchetti et al., 2002). Tali evidenze hanno portato ad investigare il ruolo del CFC sulla risposta sinaptica ippocampale. Infatti HFS ha prodotto una diminuzione nell’ampiezza del LTP nei ratti condizionati, rispetto ai valori dei soggetti di controllo naive. LTP è risultata minore nelle slices dei gruppi “condizionati” e exploration dopo 15 minuti dopo il condizionamento. A distanza di 1 giorno dal CFC, solo le slices dei “condizionati” hanno mostrato una diminuzione dell’LTP. Infine a 7 giorni di distanza LTP è risultata essere invariata anche nei soggetti “condizionati” (vedere Fig.5).
Fig.5: Modificazioni dell’LTP nella regione CA1 in slices di ippocampo di ratto
registrate a partire da 0 a 7 giorni dopo il CFC.
Questi risultati mostrano che solo la presentazione contemporanea di durata sufficiente del contesto CS e dello stimolo nocicettivo US è in grado di apportare modificazioni del comportamento spontaneo e dare apprendimento. Infatti nei soggetti shock-only l’assenza di modificazioni dell’eccitabilità sinaptica, ha evidenziato che l’emozionalità, l’attività muscolare e gli stimoli nocicettivi di per se non sono sufficienti a dare inizio al processo di apprendimento e ad innescare le variazioni nell’eccitabilità ippocampale connesse all’apprendimento stesso (Sacchetti et al., 2001 ). I dati ottenuti sui soggetti exploration, sono in linea con altri dati, i quali mostrano che ratti esposti a un ambiente “arricchito” esibiscono una diminuzione dell’LTP (Waters et al., 1997). L’affermazione, che il processo associativo mnemonico è la causa principale del decremento nell’induzione di LTP, poggia sull’ipotesi che LTP si basi su meccanismi i quali portano all’elaborazione specifica dell’apprendimento (Rioult-Pedotti et al., 2000).
I meccanismi biochimici e molecolari di consolidamento, che hanno luogo nell’ippocampo, avvengono entro le 24 ore seguenti la sessione di
acquisizione (Ambrogi et al., 1998) . Perciò, i processi di induzione dell’LTP potrebbero essere correlati alle modificazioni dell’eccitabilità dovute al consolidamento mnemonico. 7 giorni dopo l’acquisizione, l’LTP non mostra più modificazioni. Nell’ippocampo quindi avvengono modificazioni della risposta sinaptica precisamente misurabili durante il processo di memorizzazione.
1.6. MODIFICAZIONI DELL’ESPRESSIONE GENICA LEGATA A
FEAR CONDITIONING
Il fear conditioning può essere utilizzato per studiare gli eventi molecolari alla base dell’acquisizione e del consolidamento di tracce mnemoniche. Numerosi studi hanno dimostrato che meccanismi biomolecolari alla base della late-fase del LTP, sono necessari per il consolidamento della memoria alla paura nell’amigdala. Infusioni intraamigdaloidee di inibitori della sintesi di mRNA distruggono la memoria a lungo termine per la paura uditiva e contestuale (Bailey et al., 1999). Inoltre infusioni intra-amigdaloidee di inibitori della sintesi proteica o della proteina chinasi A (PKA) danneggiano il consolidamento della memoria a lungo termine alla paura ma non quella a breve termine. Questo studio dimostra che il consolidamento della memoria alla paura richiede l’espressione genica PKA-dipendente e sintesi proteica nell’amigdala (Abel et al., 1997). La somministrazione dose-dipendente intraventricolare di tre inibitori delle chinasi attivate da fitogeni (MAPK) quali anisomicina, Rp-cAMPS e pd098059 distruggono la memoria a lungo termine correlata alla paura ma non hanno alcun effetto sulla memoria a breve termine. I ratti saggiati a distanza di un’ora dal trattamento mostrano freezing associato al suono o all’ambiente. Questi studi hanno dimostrato che la via delle MAPK chinasi
è necessaria per la sintesi proteica responsabile delle modificazioni a lungo termine alla base del consolidamento della memoria alla paura (Schafe et al., 2000). Topi transgenici per la cAMP response element binding protein (CREB), mostrano deficit della memoria a lungo termine, ma non di quella a breve termine per il condizionamento alla paura sia uditivo che contestuale. Da tali risultati si può evincere che i fattori di trascrizione CREB siano coinvolti nelle modificazioni dell’espressione genica legate al consolidamento della memoria alla paura ( Abel et al., 1997).
Cambiamenti trascrizionali si osservano sia nell’ippocampo che nell’amigdala in seguito a CFC (Hall et al., 2001). Ratti sottoposti a microinfusioni di actinomicina-D (act-D) nell’amigdala basolaterale, prima di essere condizionati, presentano una riduzione delle risposte apprese a partire da 24 h fino a 48 h dopo le microinfusioni (Bailey et al., 1999). Questi risultati supportano l’ipotesi che la ritenzione a lungo termine di associazioni apprese durante FC richieda la trascrizione di nuovo mRNA e la sintesi di nuove proteine all’interno dell’amigdala (Bailey et al., 1999). Recenti ricerche hanno messo in evidenza una rapida e selettiva induzione dell’espressione di brain derived neurotrophic factor (BDNF) nell’ippocampo durante l’apprendimento alla paura contestuale. L’aumento dell’espressione di BDNF si verifica solamente nell’area CA1, il suo incremento evidenzia il coinvolgimento di questo fattore trofico nella plasticità sinaptica nel sistema nervoso centrale dell’adulto e suggerisce che sia correlato con l’apprendimento ippocampale in vivo (Hall et al., 2001).
Studi recenti basati sulla tecnica dei microarray hanno evidenziato cambiamenti dell’espressione genica nell’ippocampo e nell’amigdala in seguito a fear conditioning. Tramite questi esperimenti si è osservato, analizzando i livelli di espressione genica, che le due regioni coinvolgono
patterns genici distinti (Mei et al., 2005). Nell’ippocampo la maggior parte dei geni regolati appartiene alle categorie funzionali delle proteine segnale, dei fattori di trascrizione e delle proteine coinvolte nell’assemblaggio nella formazione delle vescicole (Mei et al., 2005). Molte delle proteine individuate nell’ippocampo, sono coinvolte anche in malattie neurodegenerative dell’Uomo come ad esempio la proteina precursore dell’amiloide (APP) della malattia di Alzheimer.
Nell’amigdala la maggior parte dei geni la cui espressione risulta modulata in seguito a CFC codifica per proteine strutturali e per proteine di adesione cellulare tipiche delle strutture sinaptiche, dendritiche e assonali come ad esempio l’actina, la spectrina cerebrale, la tubulina e le proteine associate ai microtubuli (Van Rossum and Hanisch, 1999). Sono stati anche individuati geni coinvolti nel turn-over e nella regolazione fisiologica dei recettori ionotropici, come ad esempio la proteina associata al recettore GABAA
(GABARAP). In seguito al CFC è stato osservato che l’espressione di geni specifici della glia viene aumentata (Mei et al., 2005). Questo dato è in linea con la recente osservazione che la glia sia coinvolta nei fenomeni che controllano il numero di sinapsi in vitro e con la possibilità che la glia possa giocare un ruolo importante nei cambiamenti che sono alla base della plasticità sinaptica (Ullian et al., 2001). Infine, in seguito al CFC, vengono ad essere modulati anche molti geni importanti per la formazione o la stabilizzazione della mielina come la proteina basica degli oligodendrociti associata alla mielina (MOBP).
1.7. METABOLISMO DELL’ACETIL-L-CARNITINA
La Acetil-L-Carnitina (ALC) è una molecola naturale ubiquitariamente presente nell’organismo in particolare nelle strutture nervose, dove rappresenta l’estere di L-carnitina a corta catena più abbondante (vedere Fig.6) (Shug et al., 1982). Le ultime ricerche sulla L-Carnitina ed i suoi derivati hanno chiarito alcuni aspetti dell’attività di queste molecole nell’influenzare molteplici aspetti del metabolismo cellulare in tutti gli esseri viventi. È metabolita fisiologico che gioca un ruolo di fondamentale importanza nel processo di produzione di energia a partire da lipidi.
ALC si forma dall’acetilazione della Carnitina ad opera della Carnitina Acetil Transferasi (CAT), enzima localizzato nella membrana interna del mitocondrio, che catalizza il trasferimento del gruppo acetile dall’Acetil-CoenzimaA (Acetil-CoA) derivato dalla β-ossidazione lipidica, alla Carnitina (Edwards et al., 1974).
L’Acil-Carnitina, così formatasi, attraversa la membrana mitocondriale ad opera della traslocasi e il suo gruppo acilico è nuovamente trasferito al CoA dalla Carnitina Palmitoil Transferasi CPT II, formando Acil-Coa. Nella matrice mitocondriale questo è β-ossidato con produzione di Acetil-CoA, che entra nel ciclo di Krebs con conseguente produzione di ATP (Sonnewald et al., 1993). ALC può essere trasportata all’interno del mitocondrio per mezzo della Carnitina/ Acil-Carnitina Traslocasi (CT), localizzata a livello della membrana mitocondriale interna, mediante un trasporto uniporto (vedere Fig.7) (Indiveri et al., 1994). ALC, assieme alle altre Acil-carnitine e alla carnitina libera, fa parte del “sistema della carnitina”, in cui si trova anche un complesso enzimatico, localizzato a livello della membrana mitocondriale, che comprende: Carnitina Palmitoil Transferasi (CPT I-II), Carnitina/ Acil-carnitina Traslocasi (CT), Carnitina Acetil Transferasi (CAT).
L-Carnitina, presente fisiologicamente negli organismi animali nella sua forma libera e nelle sue forme esterificate a lunga o corta catena, è l’unica molecola vettrice utilizzata dagli acidi grassi per attraversare la membrana mitocondriale, quindi è fondamentale nel metabolismo lipidico. L-Carnitina infatti permette il trasporto degli acidi grassi a lunga catena dal citosol cellulare ai mitocondri, dove il processo continua attraverso la β-ossidazione, il ciclo di Krebs e la fosforilazione ossidativa fino alla formazione di energia sottoforma di adenosintrifosfato (ATP). Proprio per il ruolo essenziale che riveste ALC , una carenza di L-Carnitina comporta un blocco della β-ossidazione (Edwards et al., 1974). ALC riveste un ruolo cruciale in questo processo in quanto ha la funzione di trasportatore intermedio del gruppo acetilico tra la molecola di L-Carnitina e quella del CoA, ed è in grado di facilitare l’up-take di CoA nel mitocondrio durante il processo di β-ossidazione degli acidi grassi, incrementare la produzione di acetilcolina e stimolare la sintesi dei fosfolipidi di membrana.
Nell’uomo il 25% della Carnitina necessaria per il fabbisogno giornaliero è sintetizzato dall’organismo nel cuore, nel fegato ed in misura minore nel cervello, mentre il restante 75% deve essere introdotto con la dieta. La L-Carnitina è immagazzinata prevalentemente nel tessuto muscolare scheletrico e nel miocardio (Rebouche and Seim 1998). La L-Carnitina e ALC vengono trasportate attraverso le membrane plasmatiche grazie a trasportatori cationici appartenenti alla superfamiglia dei trasportatori di ioni organici (Koepsell, 1998). Due di questi trasportatori sono stati isolati e caratterizzati nel ratto, nel topo e nell’uomo (Wu et al., 1998): OCTN1 (Organic Cation Transporter N1) con bassa affinità per carnitina e ALC, e OCTN2 (Organic Cation Transporter N2), con alta affinità per queste molecole (Tamai et al., 1998). L’elevata capacità di ALC di attraversare la
BEE sottolinea come questa sostanza possa agire in modo rilevante su aspetti funzionali del sistema nervoso attraverso proprietà di tipo neuroprotettivo e neuromodulatorio.
1.8. MODIFICAZIONI DELL’ECCITABILITÀ IPPOCAMPALE LEGATE ALLA SOMMINISTRAZIONE DI ALC
Le modificazioni dell’eccitabilità ippocampale date da somministrazione di ALC sono state studiate mediante tecniche di elettrofisiologia su slices di ippocampo di ratti anziani. L’ALC è stata somministrata 30 minuti prima della stimolazione tetanica (treni di 100 stimolazioni a 100 Hz) delle vie afferenti di Schaffer, a concentrazioni comprese tra 10 µM e 1 mM. La somministrazione di ALC determina un aumento della trasmissione sinaptica basale di circa il 20% e dell’LTP di circa il 150%. Questi dati suggeriscono che l’ALC è in grado di modulare la trasmissione sinaptica eccitatoria in ratti anziani (Castorina et al., 1993). Sembra che questa azione di ALC coinvolga i recettori del glutammato NMDA e i recettori muscarinici. Infatti, l’aggiunta di antagonisti dei recettori del glutammato di tipo NMDA riducono significativamente l’aumento del potenziale post-sinaptico eccitatorio (EPSP) indotto dall’ALC (Castorina et al., 1993). Questo effetto sembra però dovuto al ripristino di carenze che si sviluppano durante l’invecchiamento. La progressiva riduzione età-dipendente dei recettori di tipo NMDA nell’ippocampo è correlata con la comparsa e lo sviluppo di disturbi della memoria associati all’invecchiamento e ad alcune condizioni patologiche come la demenza. Il trattamento per sei mesi con ALC (75 mg/kg/dì) a ratti anziani attenua la perdita neuronale dei recettori NMDA a livello dell’ippocampo. Mentre una singola dose di trattamento con ALC a ratti anziani riduce di poco la perdita neuronale e non ha effetti
su ratti giovani. Questi risultati suggeriscono che ALC possa esercitare un’azione trofica e neuroprotettiva quando è somministrata cronicamente. (Castorina et al., 1994).
Nei nostri laboratori si sono effettuati studi elettrofisiologici sui neuroni sensoriali e studi comportamentali di induzione al nuoto nell’invertebrato Hirudo medicinalis. In queste cellule la stimolazione dei campi afferenti o stimolazioni intracellulari evocano una scarica di potenziali d’azione, seguita da una iperpolarizzazione postuma (AHP). L’AHP è un parametro elettrofisiologico caratteristico di molte cellule nervose sia di invertebrato che di vertebrato. Studi elettrofisiologici hanno analizzato l’effetto dell’ALC sull’AHP dei neuroni meccanocettivi T di H. medicinalis. L’AHP è sostenuta dall’attività della pompa Na+-K+/ATPasi, e in modo minore, per circa un 20-25%, dall’attivazione di una conduttanza K+/Ca2+-dipendente (gK/Ca) (Jansen and Nicholls, 1973). La ripetuta stimolazione dei neuroni T
determina un incremento dell’ampiezza dell’AHP che è sostenuto da un potenziamento dell’attività della Na+-K+/ATPasi. L’incremento dell’ampiezza dell’AHP porta ad una durevole depressione delle connessioni sinaptiche tra le cellule T e i neuroni postsinaptici (Scuri et al., 2002). Il trattamento con ALC 2mM determina un prolungato aumento dell’ampiezza dell’AHP, suggerendo un possibile ruolo della pompa elettrogenica Na+-K+/ATPasi nella modulazione della trasmissione sinaptica. Infatti in presenza di apamina, un antagonista del canale K+/Ca2+ -dipendente (gK/Ca) (Mozzachiodi et al., 2001), l’ALC è ancora in grado di
produrre un aumento dell’ampiezza dell’AHP suggerendo che possa modulare positivamente e in modo sostenuto l’attività della Na+-K+/ATPasi (Scuri et al., 2002).Studi comportamentali hanno dimostrato che una singola somministrazione di ALC induce cambiamenti comportamentali, in particolare blocca la sensitizzazione e riduce la disabitudine indotte da
stimolazione nocicettiva (brushing) nell’induzione al nuoto, dopo 2 ore dalla somministrazione, e che questi effetti permangono per oltre 11 giorni (Zaccardi et al., 2001;2004).
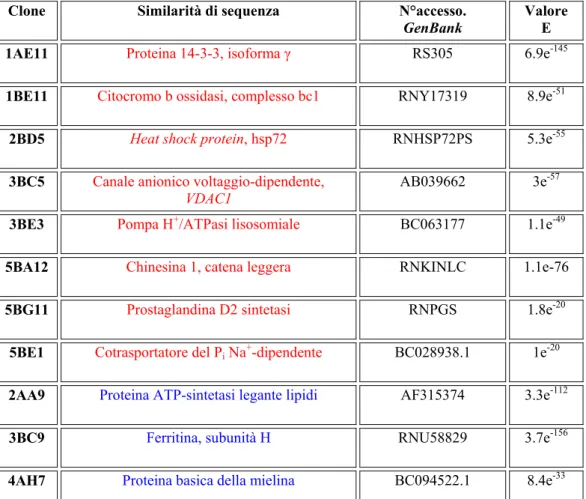
1.9. ALC E MODULAZIONE DELL’ESPRESSIONE GENICA Per individuare gli effetti di ALC a lungo termine nel sistema nervoso di ratto, nei nostri laboratori, sono stati individuati i geni la cui espressione risulta modulata in seguito a trattamento cronico con questo metabolita fisiologico (Traina et al., 2004; 2006). I geni individuati sono stati classificati in base alla loro funzione e sono mostrati in tabella 1.
Tabella 1: cloni differenzialmente espressi in seguito a trattamento cronico con ALC nel
cervello di ratto (in rosso: cloni modulati in senso positivo, in blu: cloni modulati in senso negativo). Per ogni clone sono stati riportati i codici di accesso in banca dati e il valore E delle sequenze che hanno mostrato la più alta similarità dopo analisi bioinformatica.
Clone Similarità di sequenza N°accesso.
GenBank Valore E
1AE11 Proteina 14-3-3, isoforma γ RS305 6.9e-145
1BE11 Citocromo b ossidasi, complesso bc1 RNY17319 8.9e-51
2BD5 Heat shock protein, hsp72 RNHSP72PS 5.3e-55
3BC5 Canale anionico voltaggio-dipendente,
VDAC1 AB039662 3e
-57
3BE3 Pompa H+/ATPasi lisosomiale BC063177 1.1e-49
5BA12 Chinesina 1, catena leggera RNKINLC 1.1e-76
5BG11 Prostaglandina D2 sintetasi RNPGS 1.8e-20
5BE1 Cotrasportatore del Pi Na+-dipendente BC028938.1 1e-20
2AA9 Proteina ATP-sintetasi legante lipidi AF315374 3.3e-112
3BC9 Ferritina, subunità H RNU58829 3.7e-156
Le sequenze dei cloni identificati codificano per proteine coinvolte in varie funzioni cellulari: il canale anionico voltaggio-dipendente (VDAC1), la pompa H+/ATPasi lisosomiale e la proteina ATP-sintetasi legante lipidi, sono coinvolte nel mantenimento dell’omeostasi cellulare; la Heat shock protein (hsp72) e la ferritina subunità H, sono sintetizzate in risposta a stress cellulari e quindi coinvolte in fenomeni di neuroprotezione; la proteina 14-3-3 isoforma γ e la prostaglandina D2 sintetasi sono coinvolte nel differenziamento neuronale; il cotrasportatore del Pi Na+ dipendente è implicato nel tour-over proteico; il citocromo b ossidasi complesso bc1 è un componente della catena mitocondriale di trasporto degli elettroni responsabile del metabolismo energetico delle cellule infine la proteina basica della mielina coinvolta nei processi di mielinizzazione.
Inoltre per verificare l’ipotesi che ALC possa produrre modificazioni dell’espressione genica nel sistema nervoso di H. medicinalis sono state costruite librerie sottrattive soppressive tramite la tecnica SSH (Scuri et al., 2008 submited). Questi studi presentano risultati molto interessanti che indicano i possibili meccanismi molecolari attraverso cui ALC esplica le sue proprietà neuroprotettive e neuromodulatorie sia in ratti giovani sia in H. medicinalis.