4. Risultati
4.1 Esperimenti di perdita e guadagno di funzione del
gene per il recettore 5-HT2B
Per valutare il ruolo del recettore 5-HT2B durante lo sviluppo delle strutture oculari derivanti dal POM di Xenopus, ho effettuato esperimenti di perdita e guadagno di funzione genica.
Per quanto riguarda gli esperimenti di perdita di funzione, ho utilizzato la tecnica del morpholino, oligonucleotide antisenso capace di legarsi in maniera complementare al 5’ dell’ mRNA di un gene specifico, impedendone il legame con il ribosoma e quindi la traduzione in proteina.
Fig. 4.1: meccanismo di funzionamento del morpholino
Il morpholino è stato iniettato in embrioni di Xenopus in un blastomero, a stadio di due cellule, in modo da ottenere, grazie alle caratteristiche della segmentazione di Xenopus, un embrione che abbia un
Insieme al morpholino ho coiniettato il messaggero codificante per la GFP (green fluorescent protein) o per la RFP (red fluorescent protein), traccianti vitali che mi hanno permesso di distinguere il lato iniettato da quello di controllo, utilizzando un microscopio a fluorescenza, e di selezionare quindi gli embrioni in base al lato in cui sono stati iniettati e alla corretta distribuzione dei componenti microiniettati.
Per gli esperimenti di guadagno di funzione ho utilizzato l’mRNA del recettore 5-HT2B sintetizzato in vitro, reso stabile e in grado di sopravvivere all’interno della cellula, coiniettando anche in questo caso il messaggero per la GFP o la RFP.
Nel mio laboratorio sono stati precedentemente utilizzati tre morpholini contro il recettore 5-HT2B. Il primo, 5-HT2B-MO1, è complementare alle prime venticinque basi del messaggero a partire dal sito di inizio della traduzione (ATG); il secondo, 5-HT2B-MO2, è stato disegnato per essere complementare ad una porzione della regione 5’UTR; mentre il terzo, 5-HT2B-MO3, è stato disegnato su una porzione della regione 5’UTR del gene 5-HT2B come riportato in fig. 4.2.
Fig. 4.2: schema dei tre morpholini disegnati nel nostro laboratorio.
Grazie ad esperimenti effettuati precedentemente al mio lavoro di tesi, il MO3 è risultato essere quello con una maggiore specificità e con un minor grado di effetti tossici (Reisoli et al. 2010), perciò tutti i risultati riportati in questa tesi, sono stati ottenuti mediante microiniezione di 5-HT2B-MO3.
4.2 Il gene Pitx2 è espresso nel mesenchima
perioculare di Xenopus
Al fine di visualizzare il POM, ho utilizzato come marcatore molecolare il gene XPitx2. L’espressione di questo gene in Xenopus è riportata in letteratura (Schweickert et al., 2001), ma la sua espressione non è stata descritta in dettaglio. Come prima cosa ho quindi verificato che il gene Pitx2 in Xenopus fosse espresso nel POM così come descritto nel topo, cercando anche di individuare gli stadi migliori in cui effettuare l’analisi. Ho condotto quindi esperimenti di ibridazione in situ “whole mount” su embrioni a vari stadi di sviluppo utilizzando come sonda Pitx2, seguita dal taglio degli embrioni al vibratomo.
Il messaggero per il fattore di trascrizione Pitx2 risulta essere effettivamente espresso a livello del POM dell’occhio di Xenopus a stadio 33-34, mentre la marcatura è assente sia nella retina, sia nella lente
(Figura 4.3).
Figura 4.3: Espressione dell’mRNA del gene Pitx2 a livello dell’occhio di embrioni di Xenopus
processati per ibridazione in situ a stadio 33-34 e tagliati al vibratomo in sezioni coronali. L’espressione è presente nel mesenchima perioculare, che a questi stadi migra nella parte anteriore dell’occhio.
4.3. La perdita di funzione del gene 5-HT2B altera la
morfogenesi oculare.
Ho quindi analizzato gli eventuali effetti a carico dello sviluppo dell’occhio dovuti alla perdita di funzione del recettore 5-HT2B sugli embrioni iniettati con 5-HT2B-MO3.
In particolare ho analizzato l’espressione del gene Pitx2 a livello delle cellule del POM che derivano dalle creste neurali e che migrano verso la parte anteriore dell’occhio. Ho utilizzato la tecnica dell’ibridazione in situ “whole mount” utilizzando la sonda Pitx2 su embrioni a stadio 33-34, stadio in cui le suddette cellule stanno migrando sulla superficie dell’occhio.
Confrontando il lato “wild type” con il lato microiniettato, ho notato che si verifica una significativa diminuzione del segnale di ibridazione a livello della superficie dell’occhio, mentre la marcatura intorno all’occhio (che corrisponde alle cellule mesenchimali di origine mesodermica che non migrano nella parte anteriore dell’occhio) non risulta alterata (Figura
4.4). Inoltre la marcatura a livello della fessura ottica è maggiormente
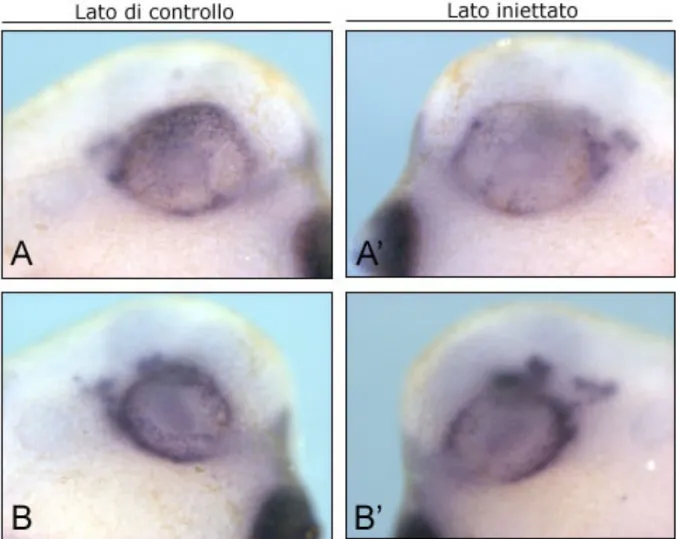
dispersa (come indicato dalle frecce nella figura 4.4.A’), a differenza del lato di controllo in cui le cellule si portano nella fessura e si dispongono in maniera più compatta (freccia in figura 4.4.A). Questo fenotipo si presenta con una frequenza del 43% (n=58).
Al fine di ottenere maggiori informazioni sul ruolo del recettore 5-HT2B sulla morfogenesi oculare, ho analizzato l’epitelio pigmentato della retina. Ho effettuato quindi ulteriori esperimenti di ibridazione in situ su embrioni iniettati con 5-HT2B-MO3 a stadio 33-34, utilizzando come marcatore il gene Dct (dopacromotautomerasi), enzima implicato nella sintesi della melanina che permette la visualizzazione dei precursori dei melanociti dell’epitelio pigmentato.
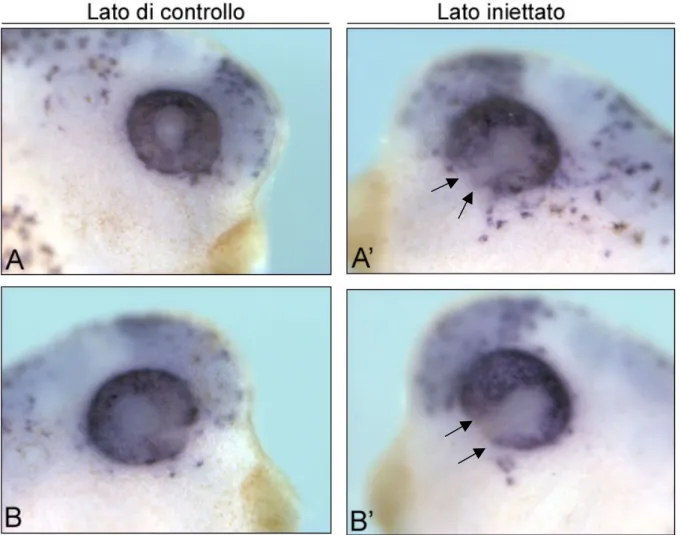
In questi embrioni, confrontando il lato iniettato con quello di controllo, ho osservato come la marcatura sia assente a livello della
fessura ottica, mentre rimane invariata nella parte più dorsale e laterale dell’epitelio pigmentato. Questi esperimenti hanno messo in evidenza come nel lato iniettato con 5-HT2B-MO3 la fessura ottica non si chiuda come invece avviene nel lato di controllo (Figura 4.5). Questo fenotipo si presenta nel 53% degli embrioni iniettati (n=30).
Fig.4.4. La perdita di funzione del recettore 5-HT2B altera il comportamento delle cellule che esprimono Pitx2. Due esempi di embrioni in visione laterale
processati per ibridazione in situ a stadio 33-34, utilizzando come marcatore Pitx2 (A e B lato di controllo di due diversi embrioni, A’ e B’ rispettivi lati iniettati) . Nel lato iniettato la marcatura è minore rispetto al lato di controllo e a livello della fessura ottica è maggiormente dispersa (frecce in A’), a differenza del lato di controllo in cui le cellule si portano nella fessura e si dispongono in maniera più compatta. (freccia in A).
Figura 4.5. La perdita di funzione del gene 5-HT2B altera la distribuzione delle cellule dell’epitelio pigmentato della retina. Due esempi di embrioni a stadio
33-34 in visione laterale dopo ibridazione in situ con la sonda Dct (A e B lato di controllo di due diversi embrioni, A’ e B’ rispettivi lati iniettati). Nel lato iniettato la marcatura manca a livello della fessura ottica (frecce in A’ e B’), mentre nel lato di controllo è presente e uniformemente distribuita (frecce in A e B).
4.4 La sovraespressione del gene 5-HT2B altera
la morfogenesi oculare
Per ottenere un quadro più completo degli effetti del recettore 5-HT2B sulla morfogenesi oculare, ho effettuato esperimenti complementari di guadagno di funzione genica del recettore 5-HT2B. Ho ripetuto quindi esperimenti di ibridazione in situ su embrioni iniettati a stadio 33-34,
avvalendomi degli stessi marcatori utilizzati per gli esperimenti di perdita di funzione.
Embrioni iniettati e ibridati con la sonda Pitx2, hanno sorprendentemente mostrato un fenotipo piuttosto simile a quello riscontrato negli embrioni microiniettati con il morpholino. Si può notare di nuovo come nel lato iniettato ci siano ampie zone dell’occhio in cui queste cellule non sono visibili mentre la marcatura intorno all’occhio sembra essere conservata (Figura 4.6). La frequenza ottenuta di questi fenotipi è del 61,5% (n=91).
Figura 4.6. La sovraespressione del gene 5-HT2B altera o l’espressione di
Pitx2 o la migrazione e/o sopravvivenza delle cellule che lo esprimono. Due
esempi di embrioni in visione laterale processati per ibridazione in situ a stadio 33-34, utilizzando come marcatore Pitx2 (A e B lato di controllo di due diversi embrioni, A’ e B’ rispettivi lati iniettati).
Embrioni in cui è stato sovraespresso il gene 5-HT2B sono stati inoltre processati per ibridazione in situ utilizzando il marcatore Dct
(Figura 4.7). Anche in questo caso ho osservato che per quanto riguarda
le cellule pigmentate la marcatura non è presente a livello della fessura ottica (frecce in figura 4.7 A’ e B’), mentre rimane invariata nella parte più dorsale e laterale dell’epitelio pigmentato. Anche in questo caso il fenotipo ottenuto è molto simile a quello riscontrato in seguito all’abrogazione del gene 5-HT2B. La frequenza ottenuta di questi fenotipi è del 69% (n=78).
Figura 4.7. La sovraespressione del gene 5-HT2B altera la morfogenesi dell’epitelio pigmentato della retina. Due esempi di embrioni in visione laterale
processati per ibridazione in situ a stadio 33-34, utilizzando come marcatore Dct (A e B lato di controllo di due diversi embrioni, A’ e B’ rispettivi lati iniettati). Le frecce in A’ e B’ indicano la mancanza del pigmento a livello della fessura ottica.
4.5. Analisi della muscolatura extraoculare
Le cellule del POM contribuiscono alla formazione della muscolatura extraoculare, in particolare quelle che derivano dalle creste neurali partecipano alla formazione del tessuto connettivo che riveste i sei muscoli extraoculari e che consente l’attacco del muscolo da un lato alla cartilagine orbitale, dall’altro all’occhio. Le cellule che derivano dal mesoderma invece, danno origine alle cellule muscolari.
Al fine di verificare se un’alterazione dell’espressione del gene 5-HT2B potesse influenzare la formazione e/o l’inserzione dei muscoli extraoculari, ho effettuato esperimenti di immunoistochimica “whole mount” su embrioni a stadio 45 iniettati con il 5-HT2B-MO3 o con l’mRNA del recettore 5-HT2B, in modo da poter visualizzare i muscoli estrinseci dell’occhio (Figura 4.8).
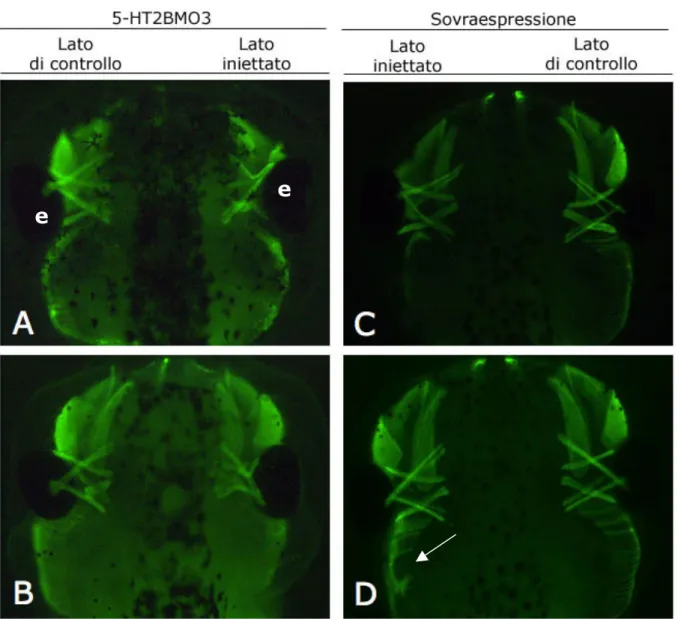
E’ stato possibile osservare che negli embrioni microiniettati con il 5-HT2BMO3, i muscoli si formano, ma si verifica un’alterazione della organizzazione stereotipata e dell’inserzione del muscolo, in particolare la tipica “struttura delle croci” manca nel lato iniettato. La frequenza ottenuta di questi fenotipi è del 34% (n=53).
Per quanto riguarda la sovraespressione invece, non ho evidenziato fenotipi rilevanti e la struttura e l’inserzione dei muscoli sembrano essere inalterate. Nella figura 4.8.D in particolare è possibile notare un’alterazione dei muscoli branchiali, fenotipo già descritto nel nostro laboratorio (Reisoli et al., 2010) ma non di quelli extraoculari. Quest’ultimo risultato conferma il successo dell’iniezione, ma come si può vedere dalle immagini, la muscolatura oculare non è alterata.
Figura 4.8. Esperimenti di perdita di funzione determinano difetti nella organizzazione dei muscoli extraoculari, esperimenti di guadagno di funzione non determinano alterazioni rilevanti. Embrioni a stadio 45 fotografati in
fluorescenza in visione dorsale e processati con l’anticorpo 12/101. Con la lettera “e” sono indicati gli occhi. A e B sono stati microiniettati con il 5-HT2BMO3 e mostrano un’alterazione delle inserzioni dei muscoli extraoculari. C e D sono stati microiniettati con l’mRNA del recettore 5-HT2B e non mostrano alterazioni rilevanti. Nella figura D è possibile notare un’alterazione dei muscoli branchiali (Reisoli et al., 2010, freccia fig.D) ma non di quelli extraoculari.
e
4.6. Esperimenti di recupero funzionale
Per poter dimostrare la specificità dei fenotipi ottenuti a seguito dell’iniezione del morpholino 5-HT2B-MO3 ho condotto esperimenti di recupero di funzione (“rescue”).
Nell’arco della stessa giornata, ho iniettato embrioni fratelli con il morpholino 5-HT2B-MO3, ed ho effettuato l’esperimento di recupero funzionale che consiste nella co-iniezione del messaggero per il recettore 5-HT2B (250pg/embrione) e il morpholino 5-HT2B-MO3 (20ng/embrione). Il morpholino, essendo stato disegnato nella regione del 5’UTR del gene 5-HT2B, si lega esclusivamente ai trascritti endogeni presenti nell’embrione mentre non è in grado di legarsi al messaggero trascritto in vitro che viene microiniettato in quanto questo non contiene la regione 5’UTR del gene.
In seguito, gli embrioni fissati a stadio 33-34, sono stati processati per ibridazione in situ utilizzando come marcatore Pitx2.
I risultati ottenuti mostrano un 24% di fenotipi per l’esperimento di perdita di funzione (n=98) e un 16% di fenotipo (n=60) nell’esperimento di recupero funzionale. Questo esperimento conferma il precedente dato pubblicato in cui si è dimostrato che il fenotipo a livello della cresta neurale cranica derivante dall’abrogazione del recettore 5-HT2B, mediante l’utilizzo di 5-HT2B-MO3, poteva essere efficientemente recuperato dalla coiniezione del messaggero per 5-HT2B (Reisoli et al., 2010). Nel mio caso sarà comunque opportuno ripetere l’esperimento nell’immediato futuro per ottenere un numero maggiore di embrioni ed un recupero di funzione più evidente.
Figura 4.9. Esperimento di “rescue” funzionale. Embrioni fotografati in visione
laterale processati a stadio 33-34 utilizzando come marcatore Pitx2. Nelle figure A e A’ è riportato un embrione co-iniettato con il 5-HT2BMO3 e il messaggero (rispettivamente lato di controllo e lato iniettato), nelle figure B e B’ è riportato un embrione microiniettato con il 5-HT2BMO3 (rispettivamente lato di controllo e lato iniettato).