Sistema nervoso: aspetti anatomici,
morfologia neuronale e connettività
sinaptica
1.1 Introduzione
Il sistema nervoso è costituito da un complesso di organi e tessuti specializzati che hanno la funzione di ricevere e riconoscere stimoli provenienti dall’ambiente esterno e dall’interno dell’organismo e di elaborare risposte coordinate volontarie o involontarie. Esso è provvisto di apparati sensoriali, atti a rilevare gli stimoli che si verificano nell’ambiente, di apparati di integrazione, che elaborano i dati sensoriali confrontandoli con le informazioni depositate nella memoria, e di apparati motori, che promuovono i movimenti e altri tipi di attività [15].
In questo capitolo l’attenzione verrà rivolta al sistema nervoso centrale, descrivendo le principali caratteristiche anatomiche, morfologiche e funzionali di questa complessa macchina e ci si soffermerà in maniera specifica sugli aspetti che più interessano in questo lavoro di tesi. Verrà descritta l’organizzazione del sistema nervoso centrale dando alcuni dettagli
in più sulla struttura del mesencefalo e del cervelletto in quanto le cellule analizzate appartengono a queste due regioni.
Si passerà quindi ad una descrizione delle unità che compongono il sistema nervoso, ovvero i neuroni, soffermandosi sui loro aspetti morfologici, e sulla loro strategia di comunicazione soprattutto a livello strutturale e connettivo piuttosto che fisiologico. Si dimostrerà che morfologia e connettività non sono due aspetti disgiunti del sistema nervoso ma sono due facce della stessa medaglia in quanto l’una è influenzata dall’altra.
Verrà illustrato infine il processo di sviluppo neuronale, dato che è fondamentale conoscere come avviene lo sviluppo del sistema nervoso per comprendere come agiscono certi disturbi dell’infanzia, come l’autismo, e possibilmente intervenire per frenare queste disfunzioni.
1.2 Organizzazione del sistema nervoso
Il sistema nervoso di tutti i vertebrati, da quello dei pesci a quello dell’uomo, può essere suddiviso in due parti: il sistema nervoso centrale ed il sistema nervoso periferico.
Con una certa approssimazione si può dire che il sistema nervoso periferico rappresenta l’interfaccia del sistema nervoso centrale con il mondo esterno. Il sistema nervoso periferico include tutti i nervi e tutti i gangli, ovvero tutti quei raggruppamenti di neuroni che stanno sparsi nel corpo al di fuori della colonna vertebrale.
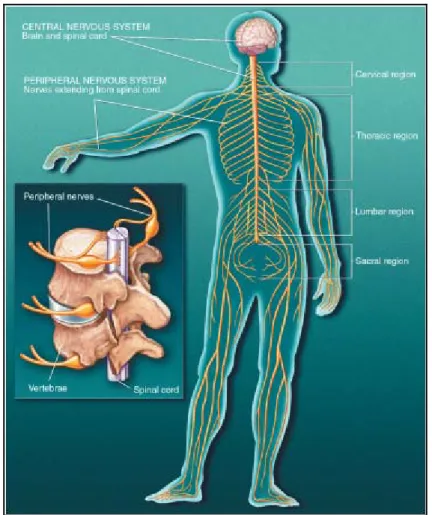
Il sistema nervoso centrale dei mammiferi è formato da tre regioni: il midollo spinale, il cervello ed il cervelletto. Il midollo spinale è contenuto interamente nella colonna vertebrale, mentre le altre strutture si trovano all’interno della scatola cranica (Fig. 1.1).
Figura 1.1 Il sistema nervoso nell’uomo
Le varie regioni del sistema nervoso svolgono compiti diversi e, in un certo senso, complementari. Il midollo spinale è la principale via di comunicazione fra il cervello ed il sistema nervoso periferico. Al midollo afferiscono tutte le informazioni sensoriali provenienti dalla pelle, dai visceri, dai muscoli e dalle articolazioni del tronco e degli arti. Dal midollo, nascono le fibre motorie che giungono ai muscoli e che controllano i movimenti volontari.
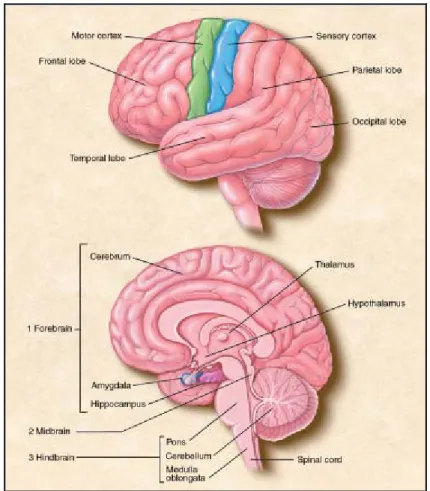
Il cervello è formato da diverse parti: il bulbo, il ponte ed il mesencefalo, che presiedono al controllo delle funzioni vegetative (ad esempio modulano la frequenza del battito cardiaco e determinano quella respiratoria); il diencefalo, formato dall’ipotalamo, con il ruolo di mantenere costante “l’ambiente interno” del corpo (cioè di mantenere attorno a certi valori ottimali caratteristiche come la temperatura corporea e la concentrazione di alcune
sostanze sciolte nel sangue), e dal talamo, attraverso il quale passano tutte le informazioni dirette alla corteccia cerebrale. Infine, ci sono gli emisferi cerebrali: sono due, sono approssimativamente uguali e, nell’uomo, ricoprono quasi tutte le altre parti del cervello. La superficie degli emisferi è rivestita dalla corteccia cerebrale. Ogni emisfero è suddiviso in quattro lobi principali, a loro volta divisi in un gran numero di aree cerebrali ognuna delle quali è specializzata per una certa funzione (Fig. 1.2) [16,17].
Figura 1.2 La corteccia cerebrale ed una sezione del cervello
1.2.1 Il mesencefalo [18, 19]
Il mesencefalo, insieme alle parti più caudali quali il ponte ed il midollo allungato, forma parte del tronco encefalico. Nel cervello umano maturo, esso
diventa la parte meno differenziata, sia nella sua forma di sviluppo che nell’ambito della propria struttura, tra le tre vescicole. Il mesencefalo è considerato la parte più rostrale del tronco encefalico.
Dal punto di vista evolutivo e filogenetico il mesencefalo umano viene considerato nella sua origine un archipallio, indicando che la sua architettura generale viene ad essere simile a quella dei vertebrati più antichi.
1.2.1.1 Organizzazione morfologica
È possibile distinguere due sezioni del mesencefalo (Fig. 1.3):
• una sezione inferiore, passante per i tubercoli quadrigemini inferiori • una sezione superiore, passante per i tubercoli quadrigemini superiori
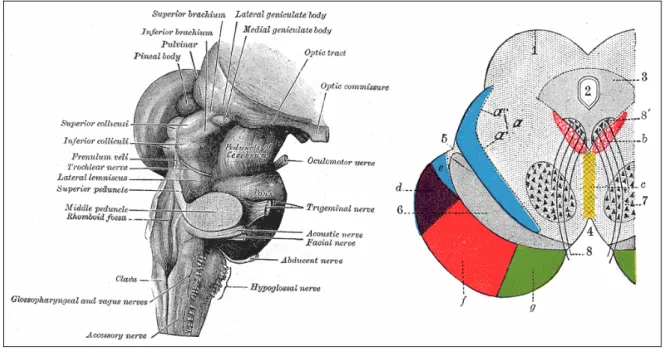
Figura 1.3 Il mesencefalo ed una sua sezione coronale: 1. Corpi quadrigemini. 2. Acquedotto cerebrale. 3. Strato grigio centrale. 4. Spazio interpediculare. 5. Solco laterale. 6. Substantia nigra. 7. Nucleo rosso del tetto. 8. Nervo oculomotore, con 8’,il suo nucleo di origine. a. Lemniscus (in blu) con a’ lemniscus mediale e a" lemniscus laterale. b. Fasciculus medial longitudinale. c. Raphé. d. Fibre temporopontine. e. Porzione del lemniscus mediale, che corre fino al nucleo lentiforme e l’isola. f. Fibre cerebrospinali. g. Fibre frontopontine.
La lamina quadrigemina, costituita dai tubercoli quadrigemini, si trova nella parte dorsale del mesencefalo. La parte ventrale del mesencefalo è chiamata peduncolo cerebrale; al suo interno si distingue un nucleo pigmentato, di colore nero chiamato sostanza nera.
Sia la sezione inferiore che quella superiore sono divise in due regioni, il tetto ed il peduncolo cerebrale separati dall’acquedotto del Silvio.
La sezione inferiore è composta da fasci che sono diretti ai nuclei basilari del ponte, da cui si dipartono milioni di fibre che si incrociano e si dirigono verso il cervelletto; da qui gli impulsi ritornano al mesencefalo per giungere poi al talamo ed ai gangli alla base. A livello della sezione inferiore si trovano due nuclei importanti: il nucleo del nervo trocleare ed il nucleo della radice ascendente del nervo trigemino. Nella sezione superiore sono invece presenti il nucleo del tetto, il nucleo pre-tettale e il nucleo rosso così chiamato per la presenza di un pigmento ricco di ferro. Il nucleo rosso riceve fibre provenienti dal cervelletto e manda fibre al talamo e alla corteccia cerebrale.
A livello della sezione superiore del mesencefalo si trova anche l’emergenza del nervo oculo-motore comune.
1.2.1.2 Funzioni
Il mesencefalo è in rapporto con la regolazione del movimento. Sia nella sezione superiore che inferiore, sono presenti fibre direttamente connesse al cervelletto con il quale avviene uno scambio di informazioni motorie.
Inoltre, la sostanza nera contiene una notevolissima quantità di un mediatore chimico, la dopamina, la cui carenza provoca disfunzioni motorie.
La sezione inferiore inoltre è un centro di riflessi uditivi: dal nucleo al corpo quadrigemino inferiore partono fibre delle vie acustiche riflesse che consentono il movimento della testa in rapporto alla sensibilità uditiva.
La sezione superiore è invece collegata a riflessi visivi: il nervo oculo-motore innerva quasi tutti i muscoli dell’occhio, le fibre che lo compongono si comportano sulla base dei segnali che provengono dalla retina, per generare il riflesso fotomotore oppure il riflesso dell’accomodazione e convergenza.
1.2.2 Il cervelletto
“Nella parte posteriore del nostro cranio, sopra il tronco cerebrale e sotto i due grandi emisferi cerebrali che lo sovrastano come un mantello, c’è una massa di tessuto bianco e grigio, delle dimensioni di una palla da baseball e di forma arcuata. È il cervelletto, il “cervello minore”. Cominciava così l’articolo che, nel 1958, presentava il cervelletto ai lettori di “Scientific American”. Scritto da Ray S. Snider, della Northwestern University, proseguiva osservando: “Al contrario del cervello, in cui l’uomo ha cercato e trovato i centri di controllo di numerose attività mentali di importanza vitale, il cervelletto è ancora una regione ammantata da un sottile mistero, le cui funzioni si nascondono agli occhi dei ricercatori”. Diciassette anni dopo, sempre su “Scientific American”, venne pubblicato un altro articolo in cui l’autore, Rodolfo R.
Linàs, affermava: “Non ci sono più dubbi sul fatto che il cervelletto
rappresenti una regione di controllo cruciale per l’organizzazione dei movimenti”.
Di recente, tuttavia, le sue funzioni sono tornate ad essere oggetto di discussione. La sua massa cospicua e la struttura complessa, suggeriscono che quest’organo abbia un ruolo più penetrante e complesso [20].
1.2.2.1 Organizzazione morfologica [21, 22]
Due componenti chiave dell’organizzazione si stabiliscono durante lo sviluppo del cervelletto: un arrangiamento altamente specifico delle corteccia cerebrale ed una microcircuiteria ugualmente specifica ed uniforme.
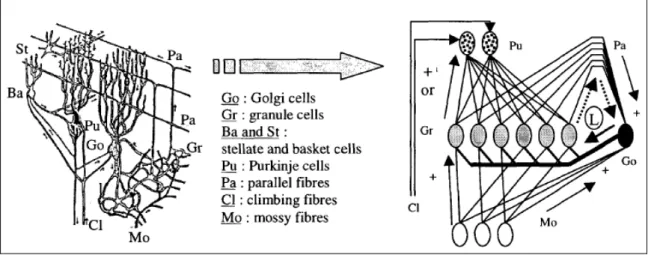
Il cervelletto è diviso in vari lobi ciascuno di quali si è sviluppato progressivamente ed ha assunto un ruolo diverso nella scala filogenetica per controllare funzioni particolarmente importanti per il comportamento ed essenziali per la sopravvivenza delle varie specie animali. Nei mammiferi, si sviluppa la parte recente, o neocerebello, costituita dagli emisferi cerebellari. La corteccia di questi emisferi è costituita da molti tipi di neuroni: le cellule granulari, le cellule di Purkinje, le cellule del Golgi, le cellule stellate superficiali e le cellule dei canestri (Fig. 1.4).
Le cellule granulari sono piccoli neuroni glutaminergici (attivabili da acido glutammico) che ricevono una vasta gamma di afferenze multimodali dal sistema delle fibre muschiose, provenienti dai nuclei pre-cerebellari che ricevono afferenze praticamente da tutte le regioni del sistema nervoso, e le smistano in modo molto ordinato a popolazioni specifiche di cellule di Purkinje attraverso il sistema delle fibre parallele. Le cellule di Pukinje sono le cellule più grandi del cervelletto. Esse sono disposte su un’unica fila all’interno della corteccia e il loro albero dendritico è orientato perpendicolarmente alle fibre parallele. I loro assoni rappresentano l’unica uscita della corteccia cerebellare trasmettendo segnali di controllo ai centri motori e non motori cui è rivolta l’azione cerebellare. Le cellule del Golgi, attivate anch’esse dalle fibre parallele, modulano il flusso di informazioni mediante un’azione inibitoria a livello della sinapsi tra fibre muschiose e cellule dei granuli. I restanti due tipi di cellule corticocerebellari, ovvero le cellule stellate superficiali e le cellule dei canestri, sono attivate anch’esse
dalle fibre parallele ed hanno un’azione inibitoria sulle cellule di Purkinje modulandone l’eccitabilità rispettivamente a livello dendritico e somatico. Un altro aspetto peculiare della corteccia cerebellare è costituito dalla presenza di un secondo grande sistema di afferenze, quello delle fibre rampicanti. Esse provengono dal nucleo olivare inferiore che riceve, in modo somatotopicamente ordinato, informazioni multimodali da tutti i distretti del sistema nervoso centrale. Le fibre rampicanti terminano direttamente sui dendriti delle cellule di Purkinje con una serie di sinapsi multiple che hanno una potente azione eccitatoria.
Basandosi sulla struttura biologica, la corteccia del cervelletto può essere modellata come una complessa rete neurale i cui nodi comunicano tramite segnali eccitatori ed inibitori (Fig. 1.4) [23].
Figura 1.4 Modello biologico (sinistra) e modello computazionale (destra)
1.2.2.2 Funzioni [21,22]
La prima funzione attribuita al cervelletto, come già stato detto, è stata la coordinazione del movimento. Questa ipotesi derivava dall’osservazione che la rimozione di questa struttura poteva provocare immediatamente difficoltà nella coordinazione motoria.
Dalla descrizione morfofunzionale tuttavia, si capisce come la struttura del cervelletto sia estremamente complessa così da far pensare ad un coinvolgimento del cervelletto in funzioni con un elevato livello d’integrazione. Per questo i neuroscienziati hanno cercato di comprendere come il cervelletto fosse implicato in compiti non solo prettamente motori. Negli anni ’80 studi anatomici sui roditori, hanno svelato come la maggior parte delle fibre afferenti al cervelletto provenissero non dagli arti ma dal muso. In base a queste osservazioni James Bower e collaboratori hanno formulato una nuova teoria sulle funzioni del cervelletto che suggerisce un coinvolgimento specifico di quest’organo nel coordinamento del processo di acquisizione dei dati sensoriali da parte del cervello.
Sempre negli anni ’80, partendo dall’osservazione che nell’uomo la porzione emisferica neocerebellare presenta uno sviluppo enorme rispetto ai primati subumani, Leiner e collaboratori, hanno formulato l’ipotesi che questo particolare sviluppo fosse legato al controllo di funzioni cognitive presenti solo nell’uomo, quali il linguaggio e le attività mentali. L’ipotesi prende origine dall’esistenza di cospicue proiezioni di fibre che da numerose aree associative della corteccia frontale e prefrontale, inclusa l’area del Broca, proiettano al nucleo olivare inferiore e ai nuclei pontini da cui partono rispettivamente le fibre rampicanti e le fibre muschiose. La presenza di questo circuito, suggerisce un possibile ruolo del cervelletto nel processo di apprendimento del linguaggio in cui esso potrebbe intervenire sia negli aspetti cognitivi di scelta delle parole e della loro successione, sia negli aspetti più prettamente motori della loro articolazione.
Infine, importanti studi di neuropsicologia, realizzati da Schmahmann e
Sherman su un elevato numero di pazienti cerebellari, hanno rivelato
importanti deficit, quali deterioramento delle capacità esecutive, depressione dell’umore, disorientamento del linguaggio e alterazioni degli aspetti emotivi
del comportamento, tutti legati a lesioni del cervelletto. Questi studi dimostrano il coinvolgimento del cervelletto nella sfera cognitiva ed affettiva.
1.3 Il neurone
Il tessuto nervoso è costituito da tre componenti fondamentali:
1. le cellule nervose o neuroni, che ne costituiscono le unità funzionali 2. le cellule non nervose o gliali, i cui tipi principali sono gli astrociti, gli
oligodendrociti e la microglia nel sistema nervoso centrale, e le cellule di Schwann nel sistema nervoso periferico
3. il tessuto connettivo fibroso e i vasi venosi e arteriosi.
I neuroni sono le unità funzionali del sistema nervoso. Essi sono cellule eucariote la cui membrana cellulare è altamente eccitabile ed è capace di modificare la distribuzione degli ioni, permettendo così la conduzione di un impulso di tipo elettrico (potenziale d’azione o spike) che costituisce il sistema di comunicazione tra le diverse unità.
In rapporto a queste funzioni di comunicazione, nella cellula nervosa sono notevolmente sviluppate due proprietà: l’eccitabilità e la conducibilità. La prima è definita come la capacità di reagire agli stimoli provenienti dall’ambiente esterno e interno trasformandoli in impulsi nervosi. La seconda è la capacità di trasmettere i segnali ad altre parti della stessa cellula nervosa o ad altri neuroni o alle cellule epiteliali, muscolari, ecc [24].
1.3.1 Anatomia microscopica del neurone [24]
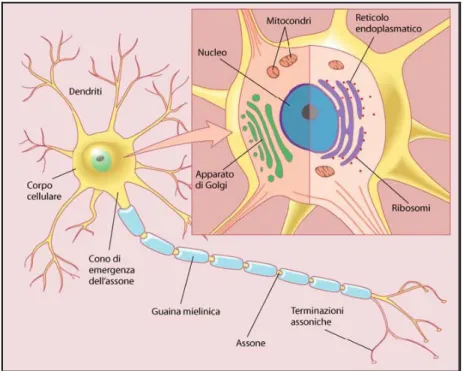
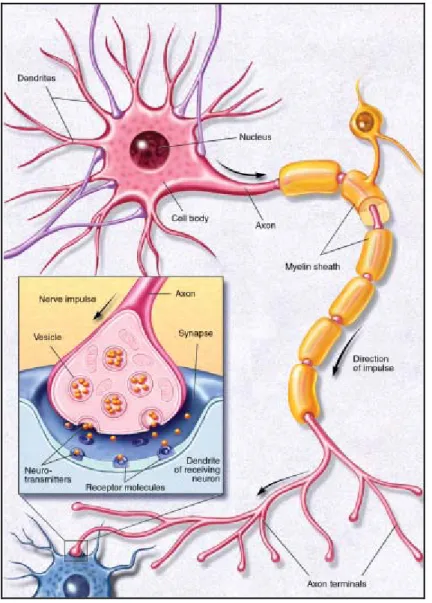
Gran parte dei neuroni è costituita da un corpo cellulare, detto pirenoforo o soma, da uno o più dendrtiti e da un assone (Fig. 1.5).
Figura 1.5 Principali componenti di un neurone
Una caratteristica comune di tutti i neuroni è una precisa polarizzazione nella conduzione dello stimolo, che viaggia sempre in una sola direzione:
dendriti → soma → assone.
I dendriti costituiscono il polo recettore del neurone e l’assone il polo trasmittente, mentre il soma costituisce il centro di elaborazione e integrazione dei messaggi in ingresso (input) provenienti da moltissime cellule, che concretizzerà in un messaggio in uscita (output).
Il soma di ogni neurone contiene il nucleo e il nucleolo. All’interno del nucleo, ricoperto da membrana, si trova il materiale genetico (cromosomi) che conserva le informazioni necessarie allo sviluppo della cellula ed alla sintesi
proteica, indispensabile per il mantenimento e la sopravvivenza della cellula stessa. Il nucleolo produce i ribosomi, necessari per la translazione delle informazioni genetiche nelle proteine.
Il soma è fornito di un apparato biosintetico ben sviluppato che produce i costituenti di membrana, gli enzimi di sintesi e altre sostanze chimiche per le funzioni specializzate delle cellule nervose. L’apparato biosinetico include i corpi di Nissl, il reticolo endoplasmatico e l’apparato di Golgi. I corpi di Nissl sono gruppi di ribosomi usati per la sintesi proteica. Il reticolo endoplasmatico (RE) è un sistema di vescicole utilizzate per trasportare materiale nel citoplasma. Può essere dotato di ribosomi (RE rugoso) o meno (RE liscio). Quando vi sono i ribosomi, il RE è importante per la sintesi proteica. L’Apparato di Golgi è una struttura costituita da membrane, importante per l’immagazzinamento di peptidi e proteine (compresi i neurotrasmettitori) in vescicole.
Il soma è inoltre dotato di numerosi mitocondri, che producono energia in forma di molecole di ATP, e di elementi del citosheletro: i neurofilamenti e i microtubuli.
I dendriti sono ramificazioni a breve decorso, che si legano al corpo del neurone in modo indistinto e possono avere più ramificazioni successive con relativa diminuzione di diametro. I dendriti di alcuni neuroni possono raggiungere 1 mm di lunghezza e possono essere tanto estesi da costituire più del 90% della superficie dell’intero neurone. I principali organelli citoplasmatici contenuti nei dendriti sono i microtubuli e i neurofilamenti Assieme al soma svolgono funzioni inerenti l’eccitabilità, in quanto su di essi terminano i prolungamenti di altre cellule nervose in regioni di contatto specializzate, dette sinapsi.
L’assone è un prolungamento in generale unico, di forma cilindrica. Esso si diparte dal corpo cellulare in corrispondenza di una protuberanza detta cono
di emergenza dell’assone, prosegue restringendosi per pochi micron, formando il segmento iniziale, ed infine si estende con forma cilindrica quasi regolare fino alla ramificazione terminale, formando le sinapsi.
La sua lunghezza varia molto, può raggiungere estensioni di oltre un metro e comunque è sempre più lungo dei dendriti. Il suo diametro, nell’uomo, può variare da un minimo di 1 μm ad un limite superiore di circa 20 μm, mentre raggiunge un massimo di 500 μm per l’assone gigante di calamaro
Gli assoni possono essere rivestiti da guaine, oppure esserne privi. Nel sistema nervoso periferico gli assoni sono comunque sempre rivestiti dalle cellule di Schwann. Molti degli assoni periferici sono avvolti da una membrana stratificata a spirale, detta guaina mielinica e originata dalle cellule di Schwann. Il citoplasma di queste cellule produce una sostanza biancastra di natura fosfolipidica, detta mielina, che avvolge l’assone come una guaina dalla sua emergenza dal pirenoforo fino allo sfioccamento finale che resta nudo; la guaina è però interrotta lungo l’assone in vari punti detti nodi di Ranvier, che hanno molta importanza nella conduzione dell’impulso nervoso e che rappresentano il punto di confine tra una cellula di Schwann e la successiva.
Nel sistema nervoso centrale la guaina degli assoni mielinizzati è formata dalle cellule dell’oligodendroglia, altri neuroni sono invece privi di guaina mielinica (assoni amielinici). Nel sistema nervoso periferico gli assoni di questo tipo sono sempre rivestiti dalle cellule di Schwann ma sono privi di mielina, mentre nel sistema nervoso centrale sono completamente privi di rivestimento.
L’assone contiene reticolo endoplasmatico liscio e citoscheletro ben sviluppato. La sua funzione principale è quella di trasmettere l’impulso nervoso ma serve anche per trasmettere a tutto il neurone gli elementi metabolici sospesi al suo interno in un fluido detto assoplasma.
1.3.2 Morfologia dei neuroni
La morfologia dei neuroni del cervello di un mammifero è molto eterogenea (Fig. 1.6): si possono annoverare circa 1000 tipi di neuroni diversi, che vengono distinti a seconda della loro forma e del tipo e quantità di ramificazioni.
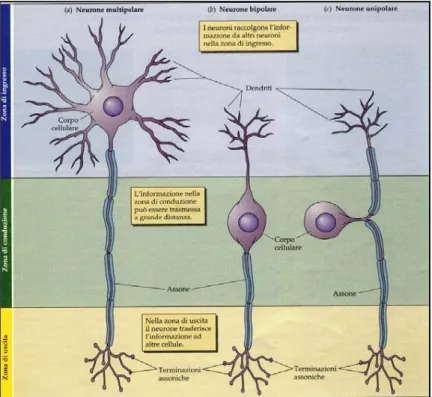
La prima classificazione morfologica dei neuroni è stata proposta da Ramon y
Cajal all’inizio del XX secolo: egli suddivise in sole tre classi le svariate
forme assunte dalla cellula nervosa, in base al numero di estensioni che dipartono dal corpo cellulare [15]:
• Neuroni unipolari: possiedono un soma di forma geometrica regolare e sono provvisti di un solo prolungamento. Al loro interno l’impulso viaggia in un’unica direzione, in particolare si allontana dal soma (centrifugo). Sono presenti nei mammiferi solo nella vita embrionale. • Neuroni bipolari: sono caratterizzati da un soma di forma generalmente
ovoidale da cui si dipartono due prolungamenti in direzioni opposte. • Neuroni multipolari: sono i più rappresentati nel sistema nervoso e
sono caratterizzati da un soma di forma variabile, da un singolo assone e da un gran numero di dendriti.
Figura 1.6 Tipi di neuroni in base alla classificazione di Ramon y Cajal
Una classificazione più complessa può essere effettuata in base al dominio spaziale nel quale si sviluppa la struttura dendritica (Fig. 1.7) [25]: si va da neuroni altamente selettivi che creano sinapsi con un solo assone, a neuroni che riempiono la maggior parte del loro dominio, creando sinapsi con ogni assone che lo attraversa, passando per morfologie intermedie.
La tendenza di un neurone a riempire un dominio spaziale può riflettere la sua propensione a creare pochi contatti con gli assoni di un gran numero di cellule oppure molti contatti con assoni di poche cellule, in base anche alla disposizione geometrica degli assoni rispetto alla struttura dendritica. Ad esempio una cellula di Purkinje (con morfologia dendritica a ventaglio) crea una o due sinapsi con ogni assone che la attraversa ortogonalmente al suo piano di sviluppo, mentre crea fino a 17 sinapsi con ogni assone orientato parallelamente a questo piano.
Figura 1.7 Differenti morfologie neuronale classificate in base al dominio spaziale nel quale si sviluppa la struttura dendritica
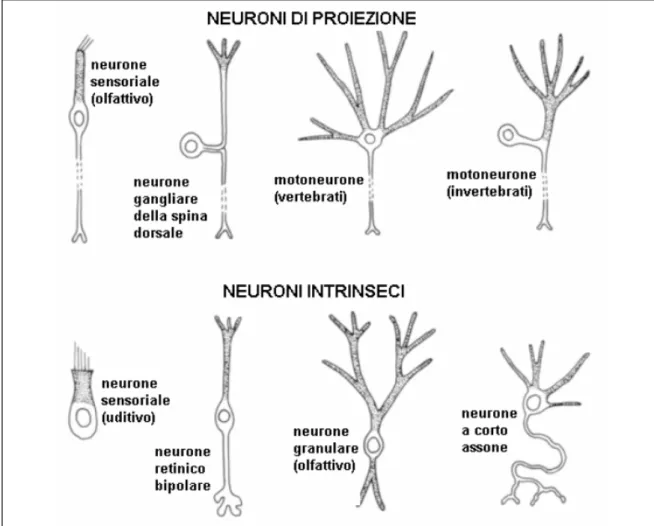
I neuroni possono essere infine classificati in due grandi classi: neuroni che si sviluppano interamente in una regione celebrale del SNC (neuroni intrinseci) e neuroni che collegano, in genere attraverso il proprio assone, regioni diverse (neuroni di proiezione). Alcuni esempi sono mostrati in Fig. 1.8.
Figura 1.8 Esempi di neuroni di proiezione e neuroni intrinseci
Un ultimo aspetto della morfologia neuronale degno di considerazione: i neuroni hanno una struttura simile ai frattali. Se si esaminano a basso ingrandimento si possono osservare ramificazioni asimmetriche (i dendriti) connesse con i corpi cellulari, a ingrandimento leggermente superiore si osservano ramificazioni più piccole a partire da quelle più grandi e così dicendo. Anche se la ramificazione dei neuroni a un certo livello si ferma, il
fatto più notevole è che questi ultimi, ad una certa scala, sono simili (seppur non necessariamente identici) a quelli della struttura vista a ingrandimento maggiore o minore.
1.3.3 Comunicazione tra neuroni: la sinapsi [15, 17]
La sinapsi (dal greco "συν - άπτειν " = "connettere") è la regione attraverso la quale un impulso nervoso si trasmette da una cellula elettricamente eccitabile ad un’altra (Fig. 1.9).
Come sopra riportato, le cellule nervose sono delimitate da un involucro costituito da cellule di nevroglia lungo gran parte della loro superficie, esso mantiene separate le cellule tra loro e da altri tessuti. Solo in punti circoscritti il citoplasma può entrare in stretto contatto: ciò si verifica lungo il soma, i dendriti e specialmente alla terminazione degli assoni. In questi punti si ha una giunzione tra cellule nervose detta appunto sinapsi. Si distinguono una zona pre-sinaptica costituta dalla porzione terminale dell’assone del neurone trasmittente (pre-sinaptico) che si sviluppa in un piccolo slargamento a forma di bottoncino (bottone sinaptico) e una zona post-sinaptica costituita da un ispessimento della membrana cellulare (nastro sinaptico) del neurone ricevente (post-sinaptico) o dell’organo ricevente.
Esistono due tipi di sinapsi: le sinapsi elettriche e le sinapsi chimiche. Nei vertebrati superiori prevalgono le sinapsi di tipo chimico.
Nella sinapsi elettrica la trasmissione dell’impulso avviene attraverso lo scambio ionico. Questo tipo di trasmissione rende la giunzione bidirezionale. La giunzione appare al microscopio come un punto di estrema vicinanza delle membrane. Tali contatti vengono indicati con il termine gap junction.
Le sinapsi elettriche sono particolarmente adatte quando serve rapidità nella trasmissione del segnale e quando è richiesta la sincronizzazione nell’attività di più cellule.
Le sinapsi chimiche sono caratterizzate invece dall’uso di neuro trasmettitori che, passando dal neurone pre-sinaptico a quello post-sinaptico, trasmettono l’impulso nervoso. Questo tipo di trasmissione rende la giunzione unidirezionale.
Figura 1.9 Sinapsi
Quando un impulso arriva a una terminazione assonica, il neurone successivo della via nervosa è influenzato in modo tale che viene a variare la sua probabilità che esso generi a sua volta impulsi. Una sostanza chimica trasmettitrice, è liberata dalla membrana pre-sinaptica terminale, si diffonde
attraverso lo stretto spazio che separa le due cellule e si lega a dei recettori presenti sulla membrana post-sinaptica. Questo legame comporta una depolarizzazione della membrana: vengono rimosse le normali barriere al flusso degli ioni sodio e potassio e ne risulta un flusso locale di ioni sufficiente ad invertire il potenziale che va dal livello della cellula a riposo di −90 millivolt ad un valore di circa 50 millivolt, prima di invertirsi nuovamente per tornare al normale potenziale di riposo. Questo segnale è quello che viene detto potenziale d’azione. Esso si propaga da una cellula all’altra attraverso la depolarizzazione di segmenti successivi di membrana: il risultato è una rapida trasmissione dell’impulso su lunghe distanze.
1.4 Connettività cerebrale
A partire dalla fine degli anni ’70 del secolo scorso, si è affermato uno schema interpretativo dei meccanismi cognitivi che guarda al cervello da un punto di vista computazionale, come un insieme di molte reti, ciascuna delle quali costituita da un certo numero di unità (i nodi) che interagiscono fra loro per elaborare uno o più flussi di informazione (anche in parallelo) [26]. Ciascuno dei nodi (i neuroni) ha una propria dinamica e, se riceve in ingresso uno stimolo, che può essere eccitatorio o inibitorio, a propria volta risponde a chi lo ha inviato ed eccita o inibisce chi gli è collegato.
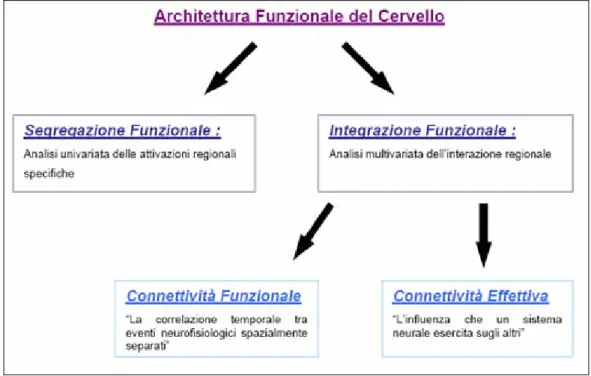
Questo tipo di modello ha influenzato profondamente le indagini del cervello. Inizialmente l’indagine mirava all’individuazione di regioni funzionali specializzate ed ha riguardato soprattutto quell’aspetto che comunemente viene definito segregazione funzionale delle aree cerebrali. Tuttavia, specialmente nelle ultime due decadi, un sempre maggior numero di neuroscienziati ha cercato di stabilire come gruppi di neuroni,
anatomicamente collegati fra di loro e caratterizzati dallo svolgimento di funzioni specifiche e diverse, si integrino secondo una certa architettura organizzativa: la così detta integrazione funzionale. Questi sono due aspetti che coesistono e si integrano all’interno delle funzionalità cerebrali; da un lato c’è la specializzazione delle aree cerebrali nello svolgimento di particolari funzioni specifiche, dall’altro ci sono una integrazione e una comunicazione molto spinte delle diverse regioni che vanno a formare dei veri e propri modelli di connettività.
Su questo aspetto i ricercatori fanno una distinzione sostanziale tra modelli di connettività indicando tre classi: connettività anatomica, connettività effettiva e connettività funzionale [27].
La connettività anatomica è determinata dalle connessioni anatomiche che esistono fra neuroni e fra loro popolazioni, e una sua qualche conoscenza risulta condizione necessaria per uno studio fruttuoso delle altre due tipologie. Per queste ultime le definizioni più largamente adottate sono quelle fornite da
Karl Friston nel 1993 [28]: la connettività funzionale è la correlazione
temporale fra due eventi neurofisiologici spazialmente distanti; mentre la
connettività effettiva è riportata come l’influenza che un sistema neuronale
Figura 1.10 Modelli di connettività
Per comprendere come diverse aree del cervello siano connesse funzionalmente ed “effettivamente” è importante conoscere le connessioni anatomiche tra i neuroni.
Le reti di connessioni possono essere classificate a diversi livelli di scala [29]: • Microscala: neuroni vicini sono strettamente interconnessi, formando
circuiti specializzati che trasmettono all’esterno l’informazione processata localmente attraverso gruppi neuronali o minicolonne
• Mesoscala: le diverse minicolonne appartenenti ad una stessa regione, sono connesse tra loro tramite connessioni orizzontali e verticali formando reti segregate funzionalmente
• Macroscala: gruppi neuronali appartenenti a regioni diverse sono connessi tra loro tramite delle connessioni a lunga distanza
Gli studi sulla connettività anatomica vengono effettuati soprattutto a livello di macroscala. Tali studi hanno messo in evidenza la presenza di pattern non
casuali nelle reti neurali. Tali reti presentano, ad esempio, le seguenti proprietà [30]:
• Presenza di cluster: gruppi di neuroni altamente interconnessi
• Organizzazione gerarchica: gruppi di neuroni altamente interconnessi fanno parte di cluster a livello più alto all’interno dei quali tali gruppi comunicano attraverso pochi nodi
• Presenza di motivi ripetuti: diagrammi specifici di interconnessioni tra neuroni che si ripetono frequentemente nelle reti
• Caratteristiche small-world: elevato livello di clustering e lunghezza delle connessioni breve
• Elevata complessità: bilanciamento tra la presenza di unità neuronali specializzati e la loro integrazione
• Frattalità e autosimilarità: ripetizione, in scala sempre maggiore, di determinati pattern di connessione
La conoscenza di queste caratteristiche topologiche ed organizzative delle reti neurali, forniscono importanti indicazioni per comprendere come gruppi di neuroni o regioni diverse del cervello sono “effettivamente” e funzionalmente connesse.
1.5 Relazione tra morfologia neuronale e connettività
sinaptica
Nella descrizione della morfologia neuronale, è stato visto come la struttura dei neuroni sia estremamente complessa e varia. Il numero e le dimensioni di assone e dendriti cambia da una classe all’altra. Tale complessità e variabilità spinge a trovare una relazione tra morfologia e funzione.
Molti progressi nella formulazione di una relazione struttura-funzione sono stati fatti utilizzando l’approccio dell’ottimizzazione delle connessioni. Per poter minimizzare le connessioni è necessario dare un significato concreto alla funzione costo da minimizzare. Sono state fatte varie ipotesi in proposito ma quella più accreditata attribuisce alla funzione costo il volume delle connessioni. In effetti la formazione di determinati pattern, gli angoli tra le diramazioni dendritiche, i diametri degli assoni e lo spazio occupato da assoni e dendriti,sono spiegati nel miglior modo attraverso la minimizzazione del volume delle connessioni.
Utilizzando dei semplici modelli matematici, si può dimostrare che solamente tenendo conto di tutte le caratteristiche della morfologia neuronale, ovvero assoni che formano diramazioni, dendriti e spine, si ottiene una stima del volume cerebrale [31]. Questo significa che i neuroni sono strutturati in modo tale da costruire una rete di connessioni sufficientemente efficiente da rientrare nel volume cerebrale.
Se i neuroni avessero solo gli assoni, dato che ciascun assone dovrebbe raggiungere tutti i corpi cellulari, il volume occupato dalle connessioni sarebbe molto elevato. La presenza dei dendriti fa sì che il segnale venga portato a metà strada, cioè non al soma ma all’estremità del dendrite, occupando in questo modo un volume minore.
Un’osservazione importante è data dal fatto che gli assoni e i dendriti hanno delle dimensioni tali da equalizzare la convergenza e la divergenza . Questa evidenza deriva dall’osservazione di molte cellule nervose quali quelle della retina, del bulbo olfattivo e del cervelletto.
Un’ulteriore diminuzione del volume delle connessioni, è ottenuta con la presenza di spine sui dendriti: in questo modo, affinché avvenga al sinapsi tra due neuroni, non è necessario che dendrita e assone si tocchino ma basta che vengano in contatto in un punto della spina.
I neuroni inoltre sembrano avere caratteristiche frattali. La geometria frattale sembra essere molto ricorrente negli organismi viventi, forse grazie alla sua alta tolleranza agli errori, alle sue capacità di economia nelle informazioni morfogeniche (e quindi genetiche) e alle sue proprietà di “ottimizzazione” del riempimento dello spazio circostante. Ma il tratto distintivo degli organismi che sembrano mostrare una qualche “frattalità” è la moltiplicazione della superficie esterna e dei punti di contatto con l’ambiente esterno, che per i neuroni sono fondamentali per l’attività elettrofisiologica delle sinapsi [32]. Nonostante queste caratteristiche generali, ogni classe di neuroni ha una sua particolare geometria che soddisfa determinati requisiti di connettività.
La minimizzazione del volume non è l’unico fattore che influisce sulla morfologia dei neuroni. I neuroni infatti hanno una geometria tale da trovare un compromesso tra il ritardo del segnale, l’attenuazione e la velocità di trasmissione dell’informazione da una parte e la minimizzazione del volume dall’altra: se gli assoni e i dendriti avessero un diametro minore, il volume verrebbe ridotto ulteriormente ma si avrebbe una maggiore ritardo del segnale negli assoni, una maggiore attenuazione nei dendriti ed una ridotta capacità di trasmissione dell’informazione a livello delle sinapsi.
Le considerazioni fatte fino ad adesso, sono relative alle connessioni tra neuroni vicini. Un’altra cosa importante è capire come la minimizzazione del
volume delle connessioni viene ottenuta a livello globale, ovvero tra diverse regioni cerebrali. In questo caso le dimensioni della rete vengono mantenute piccole adottando l’architettura small-world: interconnessioni a lunga distanza tra varie colonne di neuroni rade e fitta rete di interconnessioni tra neuroni appartenenti ad una stessa colonna [31].
1.6 Sviluppo del sistema nervoso [16, 17]
Nei vertebrati l’embrione, duranti i primi stadi di crescita, si differenzia in tre strati cellulari formando il disco embrionico: uno strato esterno (ectoderma), uno intermedio (mesoderma) ed uno più interno (endoderma).
Il sistema nervoso ha la sua origine in una regione dell’ectoderma chiamata neuroepitelio: una sezione dell’ectoderma si ispessisce formando una placca neuronale i cui bordi successivamente si arrotolano su sé stessi andando a formare il tubo neuronale che sarà la base della spina dorsale e del cervello (Fig. 1.11).
Questo avviene a partire da tre settimane dal concepimento. Alla quarta settimana si possono già riconoscere le maggiori regioni del cervello. Verso i sei mesi di gestazione sono visibili chiaramente le circonvoluzioni cerebrali, quelle tortuosità del cervello molto importanti per aumentare la superficie, fornendo posto alle più varie attività nervose.
Figura 1.11 Stadi di sviluppo del cervello
I neuroni sono generati nella cosiddetta zona di proliferazione, che circonda il ventricolo centrale di questo cervello embrionale, dalla differenziazione di cellule precursori chiamate neuroblasti. La specializzazione delle cellule embrionali in neuroni dipende dal fenomeno del differenziamento: ogni cellula dell’organismo contiene tutti i geni necessari alla formazione completa dell’intero organismo, ma solo in particolari condizioni ed in presenza di determinati fattori chimici, alcuni geni vengono espressi ed utilizzati, mentre altri rimangono silenti, dando luogo a differenti tipi cellulari.
Una volta generato, ogni neurone migra nella sua posizione finale, attraverso il supporto strutturale di particolari cellule gliali radiali, che esistono solamente in questo stadio dello sviluppo embrionale. Il posizionamento dei neuroni nel cervello embrionale segue una sequenza che va dall’interno all’esterno: i primi neuroni generati migrano negli strati più interni della placca neurale mentre quelli generati successivamente si posizionano in strati più superficiali. Durante (o immediatamente dopo) questa migrazione inizia il processo di maturazione neuronale (Fig 1.12), con l’emissione dal corpo cellulare di lunghe estensioni citoplasmatiche chiamate neuriti, destinati a diventare assoni e dendriti.
Figura 1.12 Processo di maturazione neuronale
Una volta che i neuroni sono giunti a destinazione, si compongono le connessioni appropriate per ogni singola funzione.
Il cervello continua a rielaborare e raffinare le sue strutture e le connessioni tra i neuroni anche dopo la nascita. Questi fenomeni sono necessari affinché l’organismo possa interagire, adattandosi, con le condizioni dell’ambiente circostante. I cambiamenti più evidenti avvengono prevalentemente durante alcune fasi relativamente brevi, nella vita dell’individuo: i periodi critici. Se in questi periodi il cervello grazie all’esperienza, non si “auto-costruisce” in modo adeguato, una volta passato il periodo critico, non è più in grado di adeguarsi. È importante quindi conoscere bene le varie fasi dello sviluppo per poter individuare, e possibilmente in un prossimo futuro curare o prevenire, alcune disfunzioni del sistema nervoso come quelle che causano l’autismo.